
Remodeling in Persistent Atrial Fibrillation: Pathophysiology and Therapeutic Targets—A Systematic Review

Division of Cardiology, Creighton University and CHI Health, 7500 Mercy Rd, Omaha, NE 68124, USA
*
Author to whom correspondence should be addressed.
Physiologia 2023, 3(1), 43-72; https://doi.org/10.3390/physiologia3010004
Submission received: 5 December 2022
/
Revised: 22 December 2022
/
Accepted: 4 January 2023
/
Published: 9 January 2023
(This article belongs to the Special Issue Cardiac Physiology and Pathophysiology)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Atrial fibrillation (AF) is characterized by disorganized rapid atrial electrical activity, which leads to impaired atrial function, adverse hemodynamic effects, and increased thromboembolic risk. The paroxysmal forms of AF can be effectively treated with current pharmacological and non-pharmacological modalities by targeting the arrhythmia triggers. Persistent AF, however, is more difficult to treat due to remodeling processes which may become major factors in the maintenance of the arrhythmia, rendering trigger-targeting treatment options less effective. We will systematically review the recent findings of the development and maintenance of persistent AF, including genetic, cellular, organ level, and systemic processes. As AF remains the most common sustained arrhythmia with the ongoing need to find effective treatment, we will also discuss potential treatment options targeting the remodeling processes.
1. Introduction
Atrial fibrillation (AF) is the most common sustained arrhythmia, affecting a large population and increasing morbidity and mortality. The disorganized, high-frequency atrial activity rapidly leads to a loss of atrial contractile function and an irregular ventricular rate. Long-term consequences of AF include thrombogenicity (increased risk of thromboembolic complications) and remodeling processes of the heart, perpetuating the arrhythmia and leading to maintenance of AF and non-arrhythmogenic issues (such as functional mitral regurgitation). AF—independently of other risk factors—increases mortality 1.5–3.5-fold, dementia 1.4–1.6-fold, causes 20–30% of all ischemic strokes, impairs quality of life in more than 60% of affected patients, and has a 10–40% annual hospitalization rate. Age is one of the strongest risk factors for AF—the predicted incidence and prevalence will continue to increase due to the increasing ratio of elderly in the general population [1].
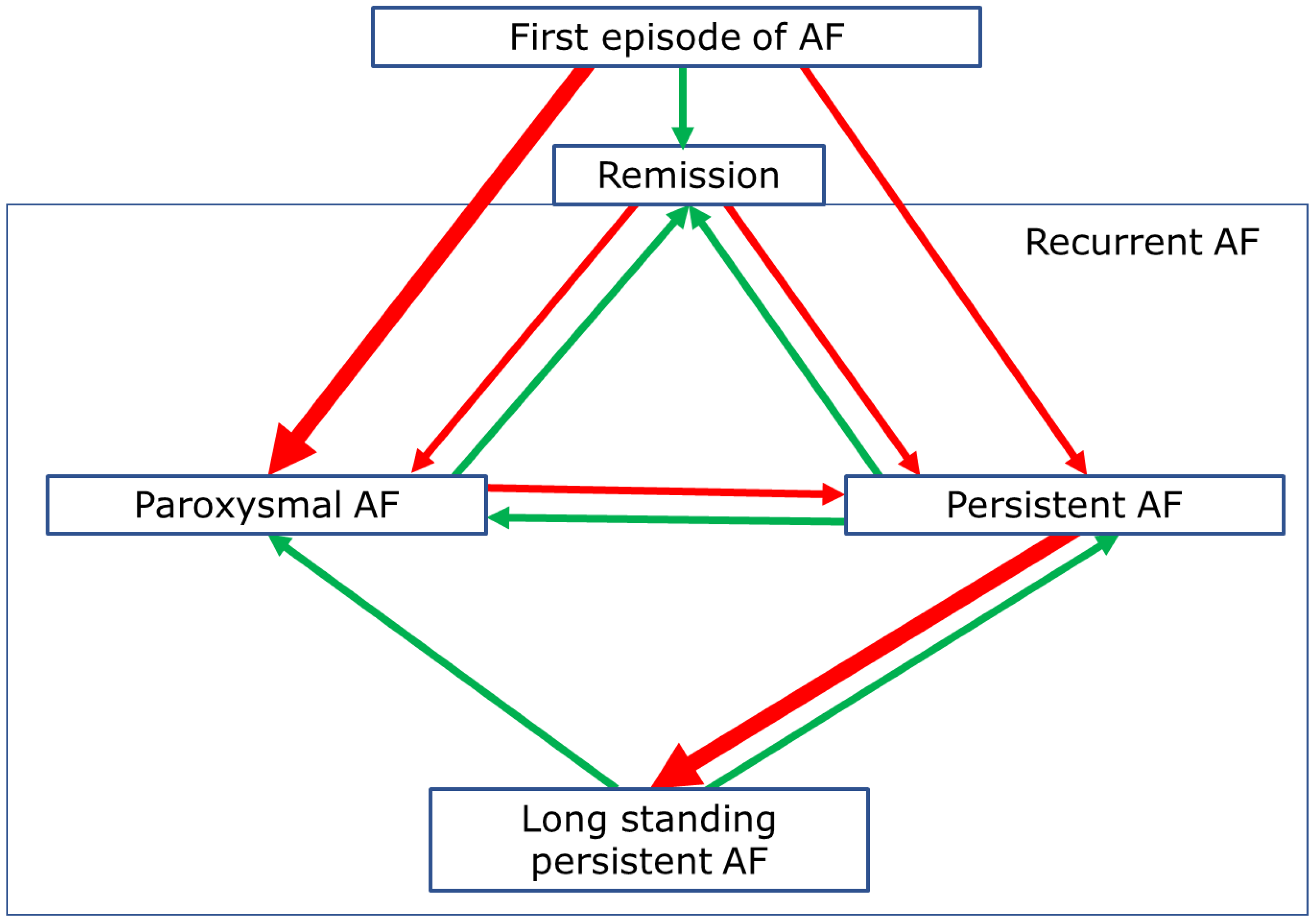
This arrhythmia is a non-homogenous entity, commonly classified according to its temporal characteristics (Figure 1). Recurrent AF is described as paroxysmal (PAF, episodes lasting ≤7 days, terminating spontaneously or with intervention), persistent (PeAF, episodes lasting >7 days), or long-standing persistent (LPeAF, the arrhythmia persisting beyond 1 year).
The pulmonary veins (PV) have been identified as a source of premature atrial contractions, triggering PAF. Ablation of these foci has led to the improvement or elimination of AF in some patients [2]. However, it was recognized early that these patients represented only a small subset of the general AF population, and ablation was much less effective in PeAF. Extensive research and development have improved the safety and efficacy of AF ablation in the past two decades—today, durable PV isolation can be performed by experienced operators in under two hours, using advanced mapping systems and safe energy sources (radiofrequency or cryoablation), often without the use of fluoroscopy. However, expanding these techniques to address non-PV triggers did not consistently improve the efficacy of ablation in PeAF [1].
One of the drivers in the development of non-pharmacological management of AF was the low efficacy of medical arrhythmia management and the lack of new medications. As the ablation procedure also struggles to effectively treat PeAF, new therapeutic targets will need to be identified. Electrophysiological changes of the atrial myocardium—caused by prolonged AF—lead to remodeling and the progressive nature of the arrhythmia, “AF begets AF” [3]. This discovery led to intense research in the pathophysiology of remodeling in PeAF, which may identify new treatment options. The objective of this review is to synthesize the research findings published within the past 10 years.
2. Methods
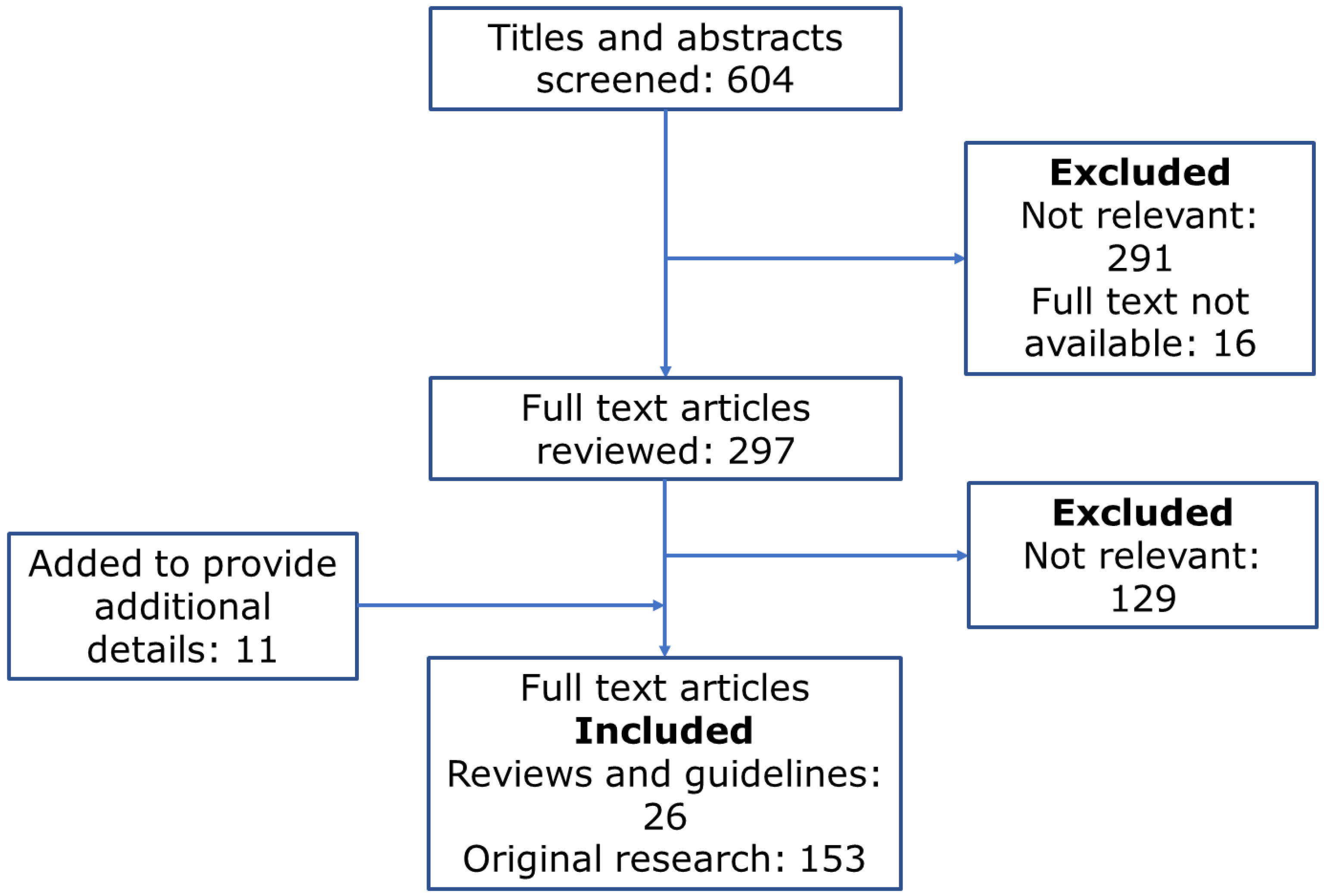
The literature search for this review has been performed according to the PRISMA guidelines [4]. We performed Pubmed, Web of Science, and Cochrane electronic database searches using the terms “persistent atrial fibrillation” and “remodeling.” The search limits were confined to the English language and the date of publication of 2012 or more recent. The databases were accessed on 29 October 2022. Abstracts of all search results were reviewed and screened by each author. Articles with abstracts relevant to this systematic review were then accessed, and all available full-text publications were reviewed. Review articles were included where they were relevant to the discussion of background information (Figure 2).
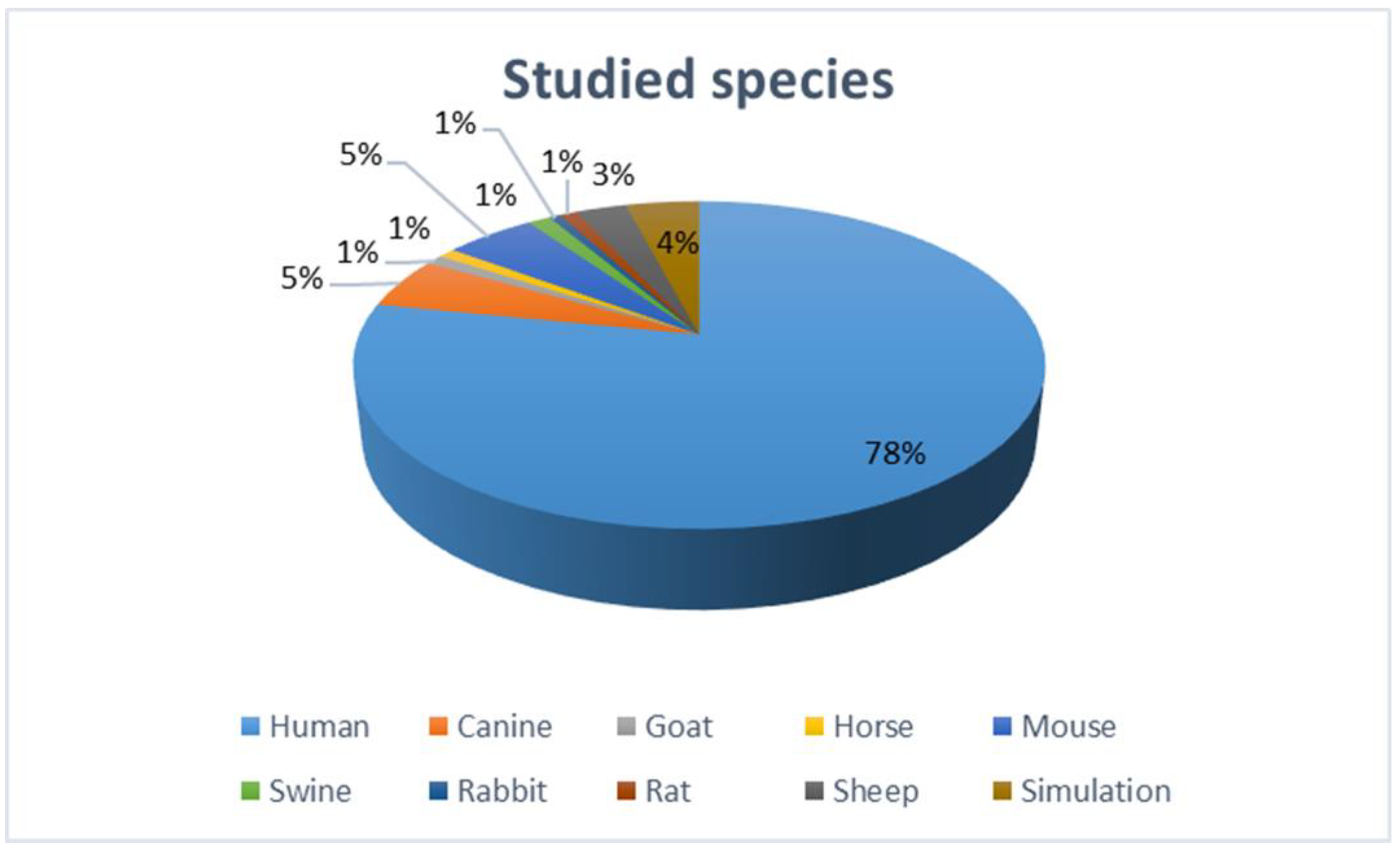
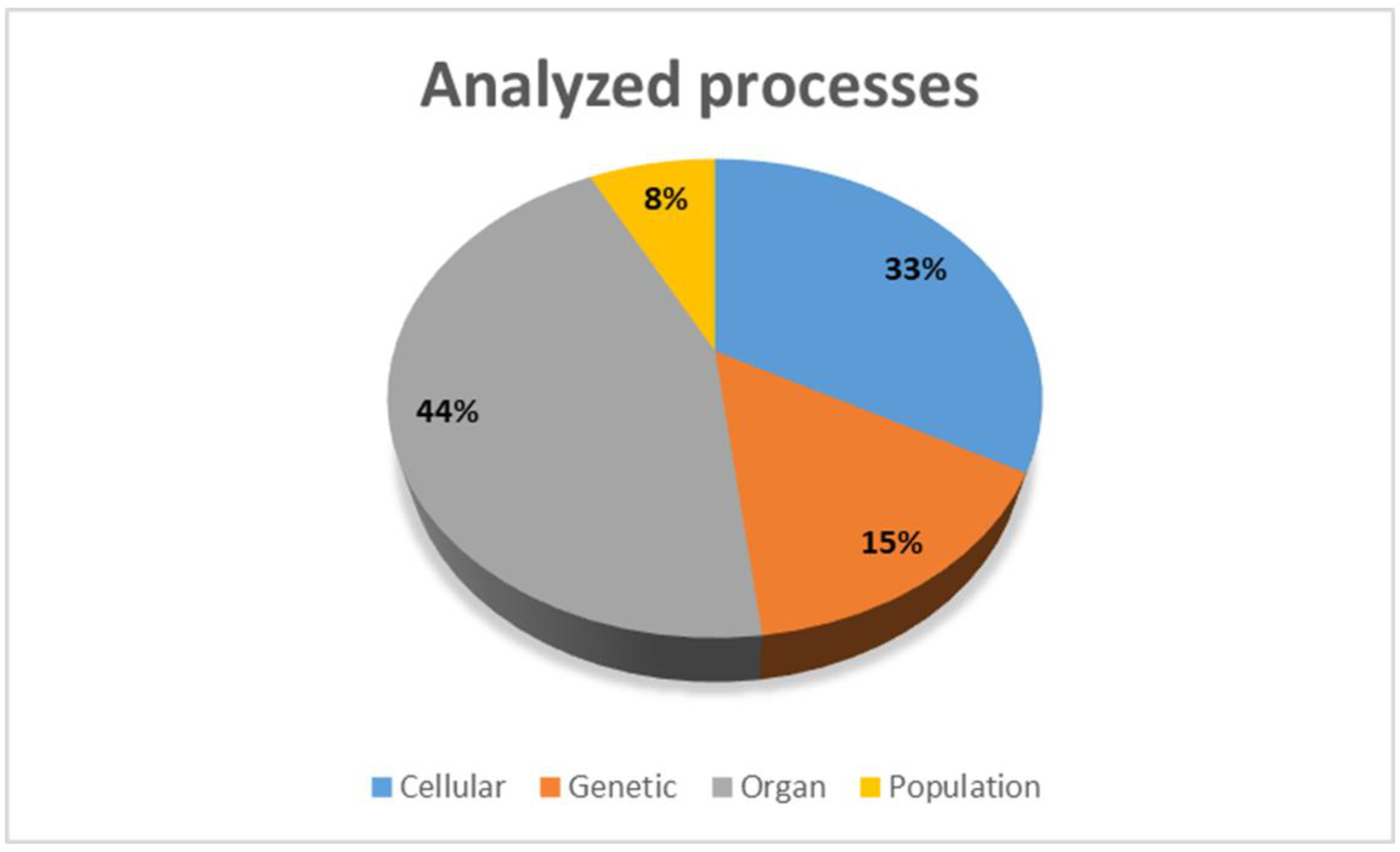
The eligibility criteria for full-text review were experimental or clinical studies of a pathophysiological process in PeAF or a review of this topic. Editorials and short letters were excluded from the full-text review. To provide a comprehensive review of the discussed topic, we included in vivo (human and animal), in vitro, and in silico (simulation) studies (Figure 3 and Figure 4).
Data from review articles were used to briefly summarize known pathophysiological processes. Data from original research articles were used to present novel findings or to expand the knowledge of a given process. In the case of multiple studies investigating the same process with similar results, larger or more recent studies were presented. In the case of studies with conflicting results, each study was presented. Small studies, where the only correlation was assessed without an attempt to clarify the pathophysiological mechanism, were excluded from this review. Structured comparative meta-analysis with statistical methods was not feasible due to the heterogeneity of research methods, study designs, and outcome measures. Synthesis was provided primarily by grouping studies by pathophysiological processes and secondarily by study design and methodology [5].
3. Progression of AF
The progression of AF is individually variable and may be affected by the combined effects of risk factors and treatment (Figure 5).
Several clinical risk factors have a strong correlation with AF progression, such as valvular heart disease, hypertension, coronary artery disease, heart failure, advancing age, male sex, dilated left atrium (LA), congenital heart disease, and acute pericarditis. Strong extracardiac factors include hyperthyroidism and alcohol consumption. More recently identified risk factors are pulmonary disease, sleep apnea, obesity, ethnicity, chronic kidney disease, and diastolic dysfunction [6]. Progression from PAF to PeAF occurred in 5.5% of older patients (mean age 65) each year in the Reappraisal of AF study. The strongest clinical risk factors were prolonged PR interval, impaired LA function, mitral valve regurgitation, and high waist circumference. Male sex, lower levels of factor XIIa: C1-esterase inhibitor and tissue factor pathway inhibitor, higher levels of N-terminal pro-brain natriuretic peptide, proprotein convertase subtilisin/kexin type 9, and peptidoglycan recognition protein 1 were also associated with AF progression [7]. In patients with implanted defibrillators or cardiac resynchronization devices, the rate of progression was 4.3% over a median follow-up of 3.4 years [8].
After the initial discoveries that the pathophysiology of PeAF may be different from its paroxysmal form, research has been intensified to identify processes that may lead to the progression of the disease. The initial focus was on the electrophysiological properties of the arrhythmia, as this was one of the most readily measurable abnormalities. As the complexity and diversity of this condition became apparent, studies have been expanded to include non-electrophysiological factors. The role of genetics, inflammation, fibrotic processes, and contribution from extracardiac remodeling have been under intense investigation recently. Multiple individual steps have been investigated in the pathogenesis and progression of AF. A synthesis suggests initiation by high-frequency excitation, followed by multiple modulating factors, such as oxidative stress, calcium overload, atrial dilatation, inflammation, and myofibroblast activation. These lead to transcriptionally mediated changes in cardiac myocytes and fibroblasts and remodeling of the extracellular matrix [9].
4. Electrophysiological Remodeling
4.1. Arrhythmia Mechanisms—Initiation and Maintenance of AF
AF is initiated by triggers; then, other mechanisms contribute to its perpetuation. The PVs have been identified as common triggers in early clinical/ablation studies [2]. The myocardial cells extending into the PVs have a higher resting membrane potential, a lower depolarization velocity, and a shorter action potential duration—these features make them more sensitive to stressors such as atrial stretch or neurogenic stimulation. The PVs were shown to be prone to developing action potential propagation blocks due to the presence of fibroblasts in a simulation study [10]. Premature beats may occur due to calcium handling abnormalities, leading to delayed afterdepolarization and triggered or automatic arrhythmia mechanisms. Atrial myocytes at the ostium of the PVs have abrupt changes in fiber orientation, promoting reentrant arrhythmia. Non-PV triggers of ectopy, initiating AF, can be observed in 20% of premature beats: ligament of Marshall, vena cava, crista terminalis, interatrial septum, or the atrial appendages. The prevalence of these triggers increases with the duration of AF [11].
The main mechanism of AF maintenance is still debated. Multiple waves propagate seemingly randomly through the atria, breaking up and creating new wavelets. The AF sustains as long as the number of wavelets remains above a critical level (multiple wavelet hypothesis). However, these may not be the primary drivers for maintenance of the arrhythmia, just results of independent mechanisms, such as functional reentry (spiral wave—rotors) or anatomical reentry (intramural reentry due nonuniform conduction—initially due to anatomical orientation of fibers, and later fibrosis) [11].
4.2. Electrical Remodeling
During sustained AF, progressive changes can be observed in the atrial electrical activity with electrocardiography or surface mapping. These changes do not affect all atrial components uniformly. Generally, shortening of the effective refractory period (an effect of cellular level remodeling) leads to increased fibrillatory frequency. Atrial fibrillatory rate, measured on a surface electrocardiogram (ECG), correlated with risk of AF recurrence after pharmacological or direct current cardioversion in a study of 195 horses: rate above 380 fibrillations per minute was significantly associated with AF recurrence (hazard ratio = 2.4, p = 0.01) [12].
Areas with sustained rapid reentrant or focal activity have dominant frequencies (DF) on ECG or intracardiac electrograms (EGM). DF analysis may be used to quantify remodeling—in a study of 40 patients with PeAF, higher DF values correlated with ablation-refractory AF [13]. In a sheep model of induced PeAF, the dominant frequency increased progressively during the paroxysmal-to-persistent AF transition period and stabilized when AF became persistent. The rate of DF increase over time correlated strongly with the time to PeAF. The cellular mechanisms were action potential duration shortening due to functional ion channel protein expression changes (CaV1.2, NaV1.5, and KV4.2 decrease; Kir2.3 increase) [14]. A link between electrical and structural remodeling was analyzed in a canine model of PeAF. Spatial maps of high DF probability were constructed and then correlated with fibrotic regions determined by T1 Mapping on MRI—regions of stable high DF bordered regions of fibrosis 82% of the time (p < 0.05) [15].
LA effective conducting size (a function of LA area, conduction velocity, and refractoriness) has been proposed as a marker for electrical remodeling: it predicted AF inducibility in patients with PeAF after pulmonary vein isolation (PVI) but not in PAF. LA effective conducting size >4 cm was required for the continuation of AF after ablation [16]. Low voltage areas in sinus rhythm show decremental conduction, which were shown to form rotational drivers during AF [17]. Prolonged P wave duration >120 ms (a marker of prolonged atrial depolarization time) after ablation of PAF or PeAF predicts AF recurrence with a sensitivity of 75% and specificity of 69%, hazard ratio 2.00 (p = 0.03) [18].
Mapping limited to the endocardial surface may not fully describe the degree of electrical remodeling. Endocardial-epicardial dissociation could be observed in a swine model of induced AF—this was more pronounced in the left vs. right atrium and persistent vs. acute AF. The LA appendage and posterior LA wall showed the most marked dissociation [19].
Remodeling also affects the right atrium, with electrophysiological changes similar to the LA (except for higher global unipolar voltage in the LA and decreased conduction velocity in the RA) [20]. This explains why atrial electrical remodeling can also be observed using implanted pacemakers and defibrillators (sensing right atrial electrical activity). The atrial activation rate (frequency of the dominant peak in the signal spectrum of bipolar EGMs of AF episodes) was 30% faster in patients who progressed to PeAF in a multicenter longitudinal study of patients with implanted pacemakers or defibrillators over a median follow-up of 3.4 years [8]. The role of the right atrium in the initiation and maintenance of AF is less known. However, higher right ventricular afterload (pulmonary hypertension), affecting right atrial function, was a risk factor for AF, independent of LA and left ventricular remodeling parameters in a study of 2246 elderly patients (mean age 75 years) [21].
Not all areas of the atria are equally prone to or affected by remodeling. The atrioventricular (AV) node showed only a mild decrease in AV nodal function (increased effective refractory period and Wenckebach cycle length) in patients with PeAF, compared to PAF, which was not dependent on the duration of AF. The presence of dual AV node physiology was similar in both groups [22]. In 117 patients with LPeAF, post-cardioversion sinus node recovery time correlated with AF recurrence: recovery time ≥1100 ms was an independent predictor of clinical recurrence of AF (HR 5.426, p < 0.001). The recovery time did not correlate with other electroanatomical remodeling parameters of LA [23]. Patients with a prolonged sinus node recovery time also have higher inducibility of non-PV triggers of AF [24].
5. Genetic Processes in Atrial Remodeling
5.1. Genomic Risk of AF
The first genetic studies in the early 2000s used segregation analysis and candidate gene sequencing to identify mutations associated with AF. Later, genome-wide association studies allowed an unbiased detection of common genetic variants. Most recently, transcriptome analysis (mRNA expression) allowed the study of dynamic changes in the remodeling process [25].
The first single nucleotide variant that was shown to have a significant association with AF is rs2200733 on chromosome 4q25. In a study of 102 patients who underwent AF ablation, 40% carried this variant. Carriers had higher conduction velocity heterogeneity (p = 0.001), complex LA signals (p = 0.002), and arrhythmia recurrence after ablation (p = 0.019). This locus appears to be a regulator for the PITX2 gene (homeodomain transcription factor 2), which has a diverse role in cardiac and extracardiac functions (including development)—however, the exact mechanism of increased AF susceptibility has not yet been elucidated [26]. KCNN3 and ZFHX3 were also suspected as AF susceptibility genes in early studies. In a genome-wide association study of 660 patients with PAF or PeAF, only ZFHX3 was associated with LA dilatation and AF recurrence after catheter ablation [27]. In another similar study, only the calcium signaling pathway (hsa04020) reached statistical significance for correlation with LA size, low voltage areas, and AF type. Within this pathway, there were 39 genes, such as CACNA1C and ryanodine receptor 2 (RyR2), that showed a good correlation with the structural changes and AF type [28]. Variations in the atrial-specific phosphoinositide 3-kinase p110α gene, which reduces the activity of this enzyme, can cause pathological atrial enlargement and increased susceptibility in AF in mouse models [29].
5.2. Transcriptome Changes Affecting AF Risk
Genome-wide mRNA microarray profiling of LA appendages from 239 patients showed that AF susceptibility was associated with decreased expression of the targets of CREB/ATF family, heat-shock factor 1, ATF6, SRF, and E2F1 transcription factors. Persistent AF activity was associated with decreased expression in genes and gene sets related to ion channel function, consistent with reported functional changes, suggesting that transcriptional remodeling marks susceptibility to AF, while the remodeling of ion channel expression occurs later in persistent AF [30]. Altered (lower) expression of circadian genes (BMAL1, CRY2, NR1D1, NR1D2, PER2, RORA, RORC, and TIM) was observed in 15 patients with implanted pacemakers and PeAF, compared to a control pacemaker group [31]. Esophageal Cancer Related Gene-4, a tumor suppressor gene, may also play a role in the pathogenesis of AF. It is homogenously highly expressed in the atria and the conduction system. The expression is decreased in the atrial appendages of AF patients. In a canine model of pacing-induced AF, the atrial expression was significantly decreased compared to controls. In a knock-down mouse model, atrial myocytes showed shortened action potential duration, activated proinflammatory cascades, and genes involved in cardiac remodeling [32].
The risk of AF is paradoxically increased in endurance athletes. A study of intensely exercised mice showed that exercise promotes tumor necrosis factor (TNF)-dependent transcriptome remodeling of the extracellular matrix and mechanosensing pathways in a time-dependent manner. Chronically, both collagen and collagen turnover genes were downregulated, similar to changes observed during aging [33]. In human atrial cells from patients with rheumatic PeAF, upregulation of intracellular chloride channels (type 1, 4, and 5) was noted, co-located with a rise in type IV collagen, suggesting that this mechanism may play a role in the fibrosis of myocyte membrane [34]. TNF-α, p53, EMT, and SYDECAN1 signaling are highly affected pathways in persistent AF [35]. These findings, however, may not be specific to the pathophysiology of AF, as similar changes were observed in atrial myocytes in patients with primary mitral regurgitation (leading to chronic pressure/volume overload of the left atrium), even without AF [36].
Transcriptome changes during remodeling are not uniform in all atrial cells: in a sheep PeAF model, the LA underwent more profound changes in expression than the right atrium. Genes involving extracellular matrix remodeling, inflammation, ion channel, myofibril structure, mitochondria, chromatin remodeling, neural function, and cell proliferation were affected [37].
Compared to the genome analysis, the proteome may be even more closely related to the disease phenotype and may signify the progression of the disease. Proteomic analysis of resected human LA appendage samples showed a difference in the expression of multiple proteins between patients in sinus rhythm of PAF/PeAF, including proteins playing roles in cardiomyocyte necrosis/apoptosis, oxidative stress, and cytoskeleton/protein binding: AKR1A1, LYZ, H2AFY, DDAH1, FGA, FGB, LAMB1, LAMC1, MYL2, MYBPC3, MYL5, MYH10, HNRNPU, DKK3, COPS7A, YWHAQ, and PAICS [38].
5.3. microRNA Abnormalities
MicroRNAs (miRNA, a non-coding RNA) regulate gene expression at the post-transcriptional level and may play a role in the pathogenesis of AF. Patients with PeAF show a different expression profile than those with PAF; the dysregulated pathways include MAPK and TGF-β signaling pathways [39].
Abnormally expressed miRNAs were noted in the pericardial fluid of patients with PeAF—the 3 most dysregulated miRNAs were miR-382-3p, miR-3126-5p, and miR-450a-2-3p, which are involved in pathways associated with cardiac fibrosis, such as the AKT1/glycogen synthase kinase-3β and TGF-β/MAPK1 pathways [40]. Decreased circulating miRNA-21 expression was noted to be associated with AF but not with AF subtypes, suggesting that molecular mechanisms responsible for the onset and progression of AF may be different [41]. However, another study showed a positive correlation between circulating miRNA-21 levels with a higher risk of AF and LA fibrosis, measured by MRI [42]. Upregulation of miR-30d was observed in patients with PeAF and may play a role in ion channel remodeling. Its role in post-transcriptional downregulation of IK,Ach (acetylcholine-sensitive potassium channel) was elucidated in a study using atrial tissue samples from older (60–79 years) male patients: miR-30d was upregulated, whereas KCNJ3, a target gene of miR-30d, was down-regulated. Overexpression of miR-30d suppressed the expression of KCNJ3 mRNA and Kir3.1 protein and, consequently, IK.Ach [43]. miR-425-5p was suggested as a biomarker for atrial fibrosis in AF—plasma miR-425-5p level negatively correlated with LA fibrosis in patients with PeAF; catheter ablation restored the decreased plasma miR-425-5p. miR-425-5p differentiated AF from healthy controls (area under the curve, AUC 0.921) and PeAF from PAF (AUC 0.888). CREB1 was identified to be a direct target for miR-425-5p in a mouse model, suggesting its role in the remodeling process [44].
6. Remodeling of Cellular Functions
6.1. Energy Homeostasis and Oxidative Stress
Frequent atrial depolarization increases the energetic demands of the cells. In a sheep model of persistent AF, glucose metabolism was upregulated, while lipid metabolism was downregulated to sustain tricarboxylic acid cycle anaplerosis. Eplerenone partially reversed the metabolic remodeling [45].
Mitochondrial dysfunction may play a role in the progression of AF—short term, mitochondria may increase the synthesis of adenosine triphosphate; however, longer-term the production, affecting the intracellular ionic equilibrium and energy-requiring enzymatic reactions, impair contraction and relaxation and activation of adenosine monophosphate protein kinase (AMPK), shifting metabolic pathways toward glycolysis and inhibiting anabolic processes. AMPK can also affect ionic channels (such as IK, ATP, and slow inward calcium channels). Dysfunctional mitochondria are also a source of free radicals, such as superoxide anion, which oxidizes multiple intracellular targets, including RyR2 of the sarcoplasmic reticulum and the sarcolemmal inward sodium channel, changing the cardiomyocyte’s excitability. Cytokine release may lead to the activation of fibroblasts and the promotion of fibrosis [46].
Oxidative stress may play a role in remodeling. Cystathionine γ-lyase (CTH) is the primary enzyme producing hydrogen sulfide (H2S), a gasotransmitter that reduces oxidative stress. In a case-control study, patients with PeAF had lower plasma acid-labile sulfide and plasma-free sulfide levels compared to patients without AF. In a follow-up mice model study, CTH-knockout mice had decreased atrial sulfide levels, increased atrial superoxide levels, atrial electrical remodeling, and increased vulnerability for induced PeAF. Rescuing the hydrogen sulfide signaling with diallyl trisulfide or reconstitution with endothelial cell-specific CTH overexpression reduced atrial superoxide, increased sulfide levels, and lowered AF inducibility [47]. Exogenous H2S supplementation in diabetic rats was able to mitigate atrial fibrosis and lowered AF incidence and duration [48].
Reduction of mitochondrial oxidative stress may help to counteract its contribution to remodeling. A crossbred mouse model was used to investigate this mechanism: animals expressing human F1759A-NaV1.5 channels were crossbred with ones expressing human mitochondrial catalase. The persistent Na+ current induced ryanodine receptor oxidation and dysfunction, which was attenuated in animals that also expressed the catalase—reduction in mitochondrial stress, cellular reactive oxygen species, and structural remodeling, were noted [49].
Changes due to intracellular remodeling may also affect response to pharmacological agents. In isolated human atrial myocytes, those obtained from patients with persistent AF showed diminished cAMP response to 5-HT4 receptor stimulation. cAMP and contractile response were recovered after inhibition of PDE3 and PDE4; however, arrhythmic responses were not recovered—regulation of these pathways in different subcellular domains was hypothesized [50].
6.2. Intracellular Ca2+
Intracellular Ca2+ affects multiple cellular functions in atrial cells, including excitability, action potential characteristics, and metabolic pathways as a second messenger. Changes in Ca2+ homeostasis (Ca2+ overload) may promote arrhythmia mechanisms, mainly by triggered activity. In a simulated model of remodeled human atria, the role of calcium handling instability was investigated: this instability led to Ca2+-driven alternans, rendered the atria vulnerable to ectopy-induced arrhythmia, and increased the complexity and persistence of arrhythmias induced by fast pacing [51]. In a simulation model, increased activity of the late Na+ current (INa,L)—although it does not have a major contribution to human atrial myocyte excitability in physiological conditions—led to the cellular accumulation of Ca2+, with subsequent dysregulation of Ca2+/calmodulin-dependent protein kinase II signaling and RyR2-mediated Ca2+ release [52].
Increased Ca2+/calmodulin-dependent protein kinase II activity may be observed in PeAF. In an atrial gene transfer porcine AF model, increased activity was observed in the early stages of AF and heart failure; inhibition preserved atrial contractile function and attenuated atrial hypertrophy, fibrosis, and apoptosis, while it did not affect inflammation or myolysis [53]. Another remodeling pathway—clustering of RyR2 channels—was investigated in human surgical right atrial appendage specimens from patients with PeAF in a nanoscale study using superresolution imaging. Despite advanced macroscale remodeling (atrial dilation and cardiomyocyte hypertrophy), clustering was not observed [54].
6.3. Proteostasis
Heat shock proteins help to maintain proteostasis (homeostasis of protein production, function, and degradation). They act as ATP-independent chaperones to prevent irreversible protein aggregation, stabilize cytoskeletal structures, increase resistance to stress situations and inhibit the activation of proteolytic enzymes. In the atrial tissues of patients with PeAF, exhaustion involving small heat shock proteins was observed. Genetic or pharmacological induction of small heat shock proteins protected against cardiomyocyte remodeling in experimental models for AF [55].
The role of autophagy—which plays an important role in proteostasis by degrading aberrant proteins and organelles—has been investigated in the pathogenesis of AF. Atrial tissue from patients with PeAF reveals activation of autophagy and induction of endoplasmic reticulum stress, which correlates with markers of cardiomyocyte damage. Tachypaced HL-1 atrial cardiomyocytes show gradual activation of autophagy: enhanced LC3B-II expression, autophagic flux, autophagosome formation, and degradation of p62, resulting in a reduction of Ca2+ amplitude. Using a sodium salt of phenyl butyrate protected atrial dog cardiomyocytes from electrical remodeling due to high-frequency pacing. Changes in Ca2+- handling, contractile dysfunction, autophagy, and attenuated AF progression were noted [56]. Autophagy-mediated electrical remodeling (degradation of L-type calcium channel) was also observed in a rabbit model of persistent AF [57].
7. Organ Level Remodeling
7.1. Contractile Function
The electrical remodeling in PeAF also leads to impaired mechanical function due to reduced intracellular Ca2+ transients [58]. These changes in contractile function can be measured using echocardiography as motion velocity or ejection fraction (EF) and can be used as markers of mechanical remodeling.
Low values of LA appendage wall motion velocity (measured by transesophageal echocardiography) predict unsuccessful cardioversion and recurrence of AF [59]. Atrial EF was investigated in a study of 50 patients who underwent AF ablation with pulmonary vein isolation (PVI); LA or LA appendage ejection fraction was lower in patients who had AF recurrence after successful ablation: LA 16% vs. 36%, p < 0.001; LA appendage: 16% vs. 42%, p < 0.001. Patients with atrial/appendage ejection fraction below 23% had a 77.8% AF recurrence rate over a median follow-up of 229 days [60].
7.2. Atrial Geometry and Wall Thickness
Atrial dilatation has been observed frequently in PeAF and may be related due to multiple factors, including chronic pressure and/or volume overload and changes in the cellular composition of atrial tissues. This is a readily measurable clinical parameter, which is discussed in depth in the section describing the reversibility of remodeling. Atrial dilatation also contributes to functional mitral regurgitation.
LA wall thickness may be a sign of compensatory hypertrophy, which correlates with the degree of remodeling in PeAF. In 100 patients who underwent ablation, preprocedural CT scans showed wall thickness ranging from 1.7 to 3.9 mm. LA roof thickness >3.1 mm predicted a strong atrial AF maintenance substrate—poor response to PVI or even complex fractionated atrial electrogram ablation (sensitivity 90.2%, specificity 94.9%) [61]. Increased wall thickness, leading to endo-epicardial dissociation, provides a 3D substrate for arrhythmia maintenance [62,63]. Epicardial breakthrough waves were observed in an epicardial mapping study in a third of patients, even in sinus rhythm, leading to endo-epicardial dyssynchrony and presenting a substrate for AF maintenance [64].
Besides the LA, geometrical changes of other atrial structures have also been evaluated in PeAF. The thickness of the interatrial septum correlates with other parameters with LA remodeling; however, its ability to predict the risk of AF recurrence after ablation is controversial [65,66]. The contribution of right atrial remodeling to PeAF is less known. In the Multi-Ethnic Study of Atherosclerosis study, higher right atrial volume indices, evaluated by MRI, were independently associated with AF (not just persistent) after adjustment for conventional cardiovascular risk factors and LA parameters [67]. The shape of the LA appendage may also affect susceptibility to AF progression. In an echocardiographic study of 225 patients, PeAF was associated with a higher prevalence of non-chicken wing–type appendages. It is unknown whether the shape of the LA appendage affects AF progression or whether PeAF leads to a change in the shape [68].
7.3. Fibrosis
Fibroblasts that are normally present in the myocardium provide a cellular scaffold for normal mechanical function and contribute to the uniformity of the excitable substrate. They also play a role in the regulation of cardiomyocyte function by slowing down the conduction in response to mechanical stretch. In pathological conditions, they may proliferate and differentiate into myofibroblasts, increase production of the extracellular matrix and lead to fibrosis. The fibrotic myocardium is heterogeneous, leading to changes in electrical conduction and functional and structural blocks, creating an arrhythmia substrate [69]. Fibrotic changes of the LA also affect its mechanical functions. Patients with AF and extensive atrial fibrosis had impaired LA reservoir and conduit strain in an MRI study of 105 patients [70].
The source of the fibroblasts in atrial remodeling has been investigated in resected LA appendages obtained from 38 patients with AF who underwent cardiac surgery. Fibrotic depositions positively correlated with LA dimensions. The endothelial–mesenchymal transition was noted by using Western blot and immunofluorescence multilabeling by confirming the transition to a mesenchymal phenotype, which can produce collagen in endocardial (CD31+) cells [71]. Epithelial-to-mesenchymal transition and activation of non-cardiomyocyte mesenchymal cells in the epicardium and perivascular space contribute to remodeling by collagen synthesis and increased turnover of the extracellular matrix [72].
Remodeling may initiate in specific locations. The posterior LA wall and floor around the antrum of the left inferior PV areas were mostly affected by fibrosis in an MRI study. However, only 68% of patients had adequate image quality for analysis [73]. A mapping study of 36 patients showed differences in regional voltage between patients with PAF or PeAF: LA structural remodeling in PeAF was suggested to initiate from the right PVs, the LA septum, roof, or posterior wall [74].
Comorbidities may affect the degree of atrial fibrosis. In a pathological study, the degree of fibrotic changes was most severe in mitral valve disease-related PeAF and correlated with the LA volume index and AF duration. The fibrotic areas mainly consisted of type I collagen, with only a minor cellular component of a few fibrocytes. The cardiomyocytes inside the fibrotic areas showed sarcolemmal damage and connexin 43 redistribution/internalization [75].
The severity and type of fibrosis affect the degree of electrical remodeling. Endomysial fibrosis (surrounding individual muscle fibers) correlated with the complexity of AF pattern (larger number of narrower waves, more breakthroughs, and a higher fractionation index) in PeAF compared to PAF (r = 0.57, p = 0.001) in a study which used epicardial high-density mapping during cardiac surgery. The amount of overall fibrosis did not correlate with AF complexity (r = 0.23, p = 0.20) in this study [76].
The electrophysiology of myofibroblasts has not been studied as extensively as that of cardiomyocytes. Ion channel expression was noted in atrial fibroblasts: voltage-gated Na+, K+, and Ca2+ channels, Ca2+-activated K+ channels, and transient receptor potential channels. A simulation study confirmed that the presence of an L-type Ca2+ current could trigger automaticity in myofibroblasts, and myocyte action potentials were prolonged when coupled to myofibroblasts with Ca2+ handling components. Different spatial myofibroblast distribution patterns increased the vulnerable window to induce arrhythmias [77]. Gap junction proteins (connexins 43, 45, and 40) play a role in cardiac myocyte-to-myocyte interaction, impulse propagation, and determine conduction velocity. They also link fibroblasts to myocytes and may be affected by remodeling [78].
Myofibroblasts contribute to atrial myocyte electromechanical remodeling via physical contact and platelet-derived growth factor (PDGF) signaling in a co-cultured sheep cell study. Myocytes contacting myofibroblasts showed a shortening of action potential duration, a decrease in calcium transients, and were responsive to tachypacing. These effects were blocked by pre-treatment with neutralizing PDGF antibody [79]. Fibroblasts obtained from the right atrial appendages of patients with persistent AF have higher connective tissue growth factor mRNA expression compared to those in sinus rhythm, promoting fibrosis [80]. Activation of cyclin-dependent kinase 1 was suggested as an important intermediary step in this process [81].
The association of biomarkers and imaging parameters with atrial fibrosis has been investigated in several studies. Late gadolinium enhancement is commonly used to assess the amount of fibrosis in cardiac structures. Left intraatrial dyssynchrony—measured with MRI—is associated with LA late gadolinium enhancement (a conventional MRI marker of fibrosis) but can be more easily obtained (analysis time 5 ± 9 min vs. 60 ± 20 min) [82]. The level of myeloperoxidase, a pro-fibrotic enzyme, is elevated in patients with AF; the level in the LA is 10 times higher than in the femoral vein, suggesting a direct origin from the LA [83]. Venous troponin T levels predict low voltage areas in patients with AF, both in PAF and PeAF (AUC 0.675, p < 0.001), however, the odds ratio is low, 1.044 (p = 0.016) [84]. Plasma microfibrillar-associated protein 4 level correlates with the presence and degree of AF (control, paroxysmal or persistent) and the degree of LA enlargement in humans. In a rat model, both plasma and atrial levels of this protein correlated with atrial fibrosis severity [85].
7.4. Epicardial Fat
Multiple population studies have shown the correlation between the amount of epicardial adipose tissue (EAT) with AF risk, independently of total body fat [86,87]. A meta-analysis of 7 studies found that there was a significant difference in the amount of EAT in patients with AF, compared to control groups (32.0 mL difference, 95% confidence interval 21.5–42.5), the highest amount of fat was observed in the PeAF group (29.6 mL difference vs. the PAF group) [88]. In an echocardiography/CT study of 205 patients, the thickness of whole heart and left atrial EAT was increased in AF: whole heart EAT in controls 6.31 ± 0.63 mm vs. 6.76 ± 0.79 mm in PAF vs. 7.01 ± 1.18 mm in PeAF, p < 0.001). LA EAT positively correlated with the LA size, P wave duration, and dispersion. Conversely, LA EAT negatively correlated with LA EF and peak strain rate [89].
EAT may influence LA function and remodeling through paracrine and inflammatory effects. In epicardial adipose samples collected from patients with non-valvular PeAF, 655 mRNAs and 57 long non-coding RNAs were differentially expressed between AF and SR (p < 0.05; fold change >1.5) [90]. Besides the paracrine and electrical effects, a large amount of epicardial fat may also interfere with the mechanical function. In a case-match study of 27 patients with PeAF and cardioembolic stroke, the amount of epicardial fat also correlated with the degree of atrial dysfunction [91].
Remodeling of epicardial fat may also contribute to structural remodeling. In patients with PeAF, fibrotic transformation of the subepicardial fat was observed. This was confirmed in a sheep model of PeAF; a process involving cytotoxic T lymphocytes, leading to adipocyte cell death, was suspected [92].
7.5. Functional Mitral Regurgitation
Functional mitral regurgitation (MR) may occur even with a structurally intact mitral valve in the context of ventricular or atrial dysfunction. MR contributes to elevated atrial pressure and volume overload. Left ventricular dysfunction causes papillary muscle dysfunction and mitral leaflet tethering, resulting in reduced leaflet coaptation. In PeAF, the mechanism of MR is due to atrial enlargement and mitral annular dilatation [93].
In PeAF, both the mitral annular area and the mitral leaflet area increase, even with preserved left ventricular function. Functional MR occurs when the increase in leaflet area fails to compensate for the annular dilatation [94]. Mitral annular contractile dysfunction, disruption of the annular saddle shape, and iatrogenic posterior mitral leaflet tethering also play a role in the mechanism [95].
Functional tricuspid regurgitation may also develop in PeAF: in a study of 83 patients, right atrial dilation was detected in 93%, while only 27% and 12% of them showed dilated or dysfunctional right ventricles, respectively. The end-diastolic tricuspid annular area had the strongest correlation with the minimum volume of the right atrium, suggesting that right atrial dilatation is an important component in functional tricuspid regurgitation. The correlation between RV and tricuspid valve dysfunction was weaker [96].
8. Extracardiac Factors Affecting Remodeling
8.1. Vegetative Nervous System
Autonomic changes can promote and maintain AF through various mechanisms, including the promotion of enhanced automaticity, early and delayed after-depolarizations, and spatially heterogenous abbreviation of refractoriness [86]. Vagus nerve stimulation exacerbated the deterioration of electrical remodeling in a canine PeAF model; ablation of the epicardial neural network blocked or reversed these changes [97].
PeAF also leads to changes in the vegetative nervous system. Both the sympathetic and parasympathetic nervous cardiac activity may increase in PeAF. In a rabbit model, persistent rapid atrial pacing led to heterogeneous autonomous neural remodeling (increased heterogeneity of tyrosine hydroxylase and choline acetyl transferase positive nerves, mostly affecting the right atrium); these changes were suppressed by using rosuvastatin [98]. In a human pathological study, patients with permanent AF had increased density of fibers of the intrinsic cardiac autonomic nervous system, especially atrial sympathetic nerve fibers, M1 muscarinic receptors, β3, and GRK-5 sympathetic receptors [99].
The remodeling process involves multiple levels of the vegetative nervous system. Ganglionated plexi ablation in a canine PeAF model effectively prolonged the atrial effective refractory period for a short time. However, the effects on atrial effective refractory period and suppression of AF induction were only transient—local atrial neuronal remodeling may also play an important role [100]. Local extensive cardioneuroablation (functional left and right atrial denervation, sparing right atrial pacemaker areas) was more effective in the prevention of repolarization changes in a similar canine model [101].
8.2. Renin-Angiotensin-Aldosterone System
The renin-angiotensin-aldosterone system (RAAS)—mainly angiotensin II (Ang II)—is involved in the process of atrial remodeling in AF. Ang II is a promoter of atrial fibrosis in animal models. In a canine model of PeAF, Ang II induced the proliferation of atrial fibroblasts and upregulated the expression of both the connective tissue growth factor and ROCK-1 in a dose-dependent manner. Their expression was upregulated by TGF-β1 and downregulated by simvastatin or losartan. Losartan and simvastatin also attenuated the effects of TGF-β1 [102]. The integral membrane protein angiotensin-converting enzyme 2 (ACE2) plays a protective role as it degrades Ang II into Angiotensin-(1–7) and may diminish this pro-remodeling effect. Plasma ACE2 levels are elevated in patients with AF and have a weak but significant correlation with the degree of LA remodeling: LA bipolar voltage, a high proportion of complex fractionated electrograms, or long LA activation time [103]. In a canine PeAF model, ACE2 overexpression suppressed atrial electrical remodeling and improved atrial function through the TRPM7 signaling pathway [104].
9. Systemic Factors in Remodeling
9.1. Inflammation
Inflammation may contribute to electrical and structural remodeling of the atria. Several inflammatory markers, such as C reactive protein, IL-6, IL-8, TNF, myeloperoxidase, cfDNA, and beta-galactoside binding lectin galectin (galectin-3) levels were elevated in patients with AF in multiple studies, depleted protective heat shock proteins (such as hsp27) were also noted [105]. Only myeloperoxidase had a weak but significant predictive value for AF recurrence after cardioversion or ablation [106]. An immunohistochemical study of patients undergoing open heart surgery showed a site-specific increase of CD45+ inflammatory cells in the atrial myocardium of patients with AF, compared to those in SR—mostly from the monocyte-macrophage lineage [107].
The NLRP3-inflammasome is an intracellular sensor that detects various environmental irritants and mediates caspase-1 activation and interleukin-1β release in immune cells. It may also play a mechanistic role in the association of AF and inflammation: NLRP3-inflammasome activity is increased in atrial cardiomyocytes in patients with PAF or PeAF. A knock-in mouse model expressing constitutively active NLRP3 showed spontaneous premature atrial contractions and inducible AF. The cellular mechanism included ectopic activity, abnormal sarcoplasmic-reticulum Ca2+ release, shortening of the atrial effective refractory period, and atrial hypertrophy. Specific inhibition of NLRP3 attenuated the response. Genetic inhibition of NLRP3 also prevented AF development in another transgenic mice model (CREM) of AF [108].
Inflammation may not play a role in all atrial remodeling processes. Resolvin-D1, an inflammation-resolution promoter, suppressed inflammatory changes and fibrotic/electrical remodeling in the right atria of rats with induced pulmonary hypertension; however, it did not prevent atrial enlargement [109].
9.2. Hypercoagulability
AF induces hypercoagulability; however, the role of the coagulation cascade components in promoting AF remodeling is less known. The presence of low voltage LA areas correlated with 30 differentially abundant proteins of the coagulation and complement cascades (q < 0.05) in an untargeted proteome analysis study of 48 patients (59% with persistent AF) [110]. Protease-activated receptors and factor Xa provoked remodeling in persistent AF animal models, which was prevented by specific inhibitors [86]. Factor Xa promotes thrombus formation and exacerbates inflammation via the activation of protease-activated receptor 2 (PAR-2). Inhibition with rivaroxaban showed a cardioprotective effect against pressure-overload-induced atrial remodeling and AF vulnerability in a mouse model [111].
9.3. Galectin-3
Galectin-3 is expressed in fibrotic tissues and may serve as a marker of fibrosis in patients with persistent AF [112]. Its pentamers bind to TGF-β receptors causing cell surface retention and internalization of the receptors, mediating duration and directionality of signaling through Smad and Akt proteins. Galectin-3 induces atrial fibroblast proliferation, myocyte hypertrophy, and electrical remodeling, which was mitigated with a specific blocker (GMCT) in a sheep model. High levels also predicted AF recurrence after ablation [113].
Elevated levels of galectin-3 were observed in a transesophageal echocardiographic study of 153 patients—patients with PeAF or permanent AF had higher levels compared to patients with PAF, the levels correlated with the degree of LA appendage remodeling; galectin-3 was an independent predictor for LA appendage thrombus (galectin-3 level > 18.95 ng/mL: sensitivity 90.9% and specificity 96.9%) [114].
9.4. Homocysteine
Homocysteine, an amino acid produced in the process of methionine metabolism, regulates the activity of the ion channels through their redox state. Elevated homocysteine levels can lead to electrical remodeling of the cardiomyocytes by affecting Na+ and K+ currents, shortening the action potential duration, and decreasing its rate of adaptation. In population studies, an elevated homocysteine level was an independent risk factor in thromboembolic events and AF. Homocysteine can activate TRCP3, a regulation factor of fibrotic mechanisms, and activate fibrotic changes in the atria [115].
9.5. Uric Acid
Elevated uric acid levels are common in conditions where the risk for AF is also increased: hypertension, type 2 diabetes mellitus, end-stage renal disease, and heart failure. Hyperuricemia may act as an independent risk factor for AF progression by activating xanthine oxidoreductase or lead to intracellular accumulation of soluble uric acid (increasing oxidative stress). The monosodium urate crystals may also activate the NLRP3 inflammasome, which then promotes the maturation of active forms of IL-1β and IL-18, promoting an inflammatory response [116].
The levels of uric acid and components of the RAAS system are elevated in patients with PeAF, compared to patients with PAF or sinus rhythm. The elevated uric acid level in these patients was an independent risk factor for elevated renin, Ang II, and aldosterone [117].
9.6. Gastrointestinal Effects
Elevated liver enzymes, but not hepatic steatosis, were associated with increased AF prevalence in the general adult population in a population study of 3090 patients. The increased levels of serum liver enzymes may correlate with inflammatory, procoagulant, and fibrinogenic mediators; however, these were not separately evaluated [118].
Chenodeoxycholic acid (a bile acid) level was higher in patients with PeAF than in those with PAF or other paroxysmal supraventricular tachycardia. The chenodeoxycholic acid level was positively correlated with the size and proportion of low voltage areas (r = 0.78 and r = 0.89, p < 0.05) in these patients. In a mouse model, chenodeoxycholic acid promoted atrial myocyte apoptosis in a concentration-dependent manner. A suggested mechanism for bile-acid-induced apoptosis is through the activation of the FXR nuclear receptor signaling pathway [119].
In a mouse model, short-chain fatty acid derived from dietary fiber fermentation by gut commensals alleviated AF development via GPR43/NLRP3 signaling [120]. Elevated microbial diversity was noted in patients with PAF or PeAF, compared to patients without AF—this will need further investigation as it may offer a treatment target if the gut microbiome plays a role in AF progression [121].
10. Clinical Risk Factors for AF Progression
10.1. Age
The increasing age of the general population is one of the main factors in increasing AF prevalence. Even without AF, remodeling of the LA can be observed with advancing age: progressive increase in LA dimensions, LA stiffness, and decrease in strain indices were observed with echocardiography; decreased LA voltage, increased LA surface area and volume, increased ratio of low voltage areas was noted with electroanatomical mapping in elderly patients (>65 years old), compared to younger ones [122]. In a study of 122 patients, who underwent ablation for PeAF, increasing age was associated with electric remodeling of the LA, characterized by a decrease in the extent of complex fractionated electrogram area and its index [123].
10.2. Obesity
Obesity is associated with an increased risk of AF; every 5-point increase in body mass index increases the risk of AF incidence by 20–30%, independently of gender, age, diabetes, and hypertension. Due to the high prevalence of obesity, it is the second most important risk factor for AF after hypertension. Processes that contribute to remodeling, independent from common complications of obesity (metabolic syndrome, sleep apnea), are increased epicardial adipose tissue, LA enlargement due to increased filling pressure, and systemic pro-inflammatory state [124].
Impaired conduit and reservoir function was observed even in milder obesity in the Asklepios Study in younger patients (35–55 years, BMI > 30) [125]. Obesity was associated with higher LA volumes but did not correlate with mechanical function in an exercise stress echocardiography study of patients with PAF or PeAF [126].
10.3. Metabolic Syndrome
Metabolic syndrome consists of central obesity, dyslipidemia, hypertension, and impaired glucose tolerance/diabetes mellitus. Hypertension promotes LA remodeling due to compensatory hypertrophy, fibrosis, and inflammation. The effect of diabetes is less clear; gap junction dysfunction and electrical remodeling due to outward K+ channel expression may contribute. Although total cholesterol or LDL level has an inverse relationship with AF incidence, metabolic syndrome leads to unfavorable changes in lipoprotein characteristics (higher oxidized lipid content in VLDL), which has toxic effects on cells expressing VLDL receptors, such as cardiomyocytes [127]. Other elevated biomarkers in metabolic syndrome with a suspected or proven role in AF progression elevated include adiponectin, leptin, TNF-α, IL-6, IL-10, PTX3, ghrelin, uric acid, and oxidized LDL [128].
10.4. Sleep Apnea
Acute effects of sleep apnea are mainly due to hypoxemia, hypercapnia, exaggerated negative intrathoracic pressure, and arousals. Chronic effects include autonomic dysregulation, inflammation, procoagulant state, insulin resistance, endothelial dysfunction, oxidative stress and accelerated atherosclerosis. Sleep apnea increased the risk of AF 4-8-fold in population studies. The prevalence of sleep apnea in the AF population is 43–73%. Increased vagal tone activates the IK,Ach current and decreases the atrial effective refractory period. Apnea episodes may increase cardiac afterload and stimulate myocyte hypertrophy and fibrosis. Measures of atrial electromechanical coupling correlate with sleep apnea severity. Untreated sleep apnea is an independent risk factor for AF recurrence after ablation. The severity of sleep apnea affects the degree of atrial remodeling in PAF; however, when the AF becomes persistent, significant atrial remodeling was observed in a mapping study of 66 patients, independent of sleep apnea severity. Thus, sleep apnea may play an important note in the earlier stages of AF progression [129]. Treatment with nocturnal continuous positive airway pressure reduces LA size and decreases the AF recurrence rate after ablation to a level comparable to patients without sleep apnea [130].
10.5. Gender
Although the overall risk of AF is 1.5-fold higher in men compared to women, the risk for complications such as heart failure or stroke is higher in women. Women also have a higher risk of AF recurrence after ablation [131]. Women have a higher level of fibrosis which may be an explanation for their increased risk for stroke in AF [132]. Fibrotic areas correlated with prothrombotic endocardial changes in an MRI study: decreased wall shear stress, increased oscillatory shear index, endothelial cell activation potential, and relative residence time (residence time of blood near the LA wall) [133]. In a study of pulmonary vein sleeves obtained from 166 patients, females with LPeAF had a higher degree of fibrosis compared to patients without AF, while this difference was less pronounced in males. The TGFβ/Smad3 pathway was upregulated in the female AF group, promoting fibrosis [134]. Overexpression of TGF-β1 led to an increase in atrial fibrosis, myocyte size, and progressive P-wave prolongation in a transgenic goat model [135].
In a study of 70 patients with PeAF or LPeAF, older age, greater left atrial area, and female gender correlated with LA scar, while the duration of AF did not [136]. Gender differences were also noted in an ablation study of 85 patients with persistent AF: ultra-high-density mapping showed a higher incidence of focal and rotational activity in men vs. women (73.4% vs. 38.1%, p = 0.003)—the absence of these areas shows more advanced remodeling [137].
10.6. Race
The role of race as an AF risk factor has not been elucidated yet. Caucasians have higher rates of AF, while electrical remodeling occurs earlier in Black hypertensive patients. Most studies underreport racial characteristics [138].
10.7. Biomarkers
Studies analyzing the correlation of biomarkers with atrial remodeling and AF progression led to further investigations analyzing their role in the pathogenesis: inflammatory and metabolic markers have been discussed in the prior sections. Some markers, such as CRP, may decrease promptly after successful cardioversion, while others may also be affected by common comorbidities (such as N-terminal pro-BNP in patients with AF and concomitant heart failure) [139].
Clinically useful biomarkers in PeAF include those that correlate with AF progression or identify a disease phenotype that may respond to a specific treatment. Atrial natriuretic peptide (ANP), soluble interleukin 1 receptor-like 1 (ST2), and galectin-3 levels correlate with the degree of atrial myopathy [112,139]. Proprotein convertase subtilisin/kexin type 9 and peptidoglycan recognition protein 1 are associated with AF progression [7]. From the inflammatory markers, myeloperoxidase may have a predictive value for AF recurrence when attempting rhythm control [106]. Several biomarkers have been studied to assess the risk of thromboembolic or vascular events beyond the currently used clinical risk scores: D-dimer, antithrombin-III, high sensitivity troponin T, BNP, IL-6, NADPH oxidase 2, and CRP. So far, these only had a modest additional predictive value when used together with clinical risk scores [140].
11. Reversibility of Remodeling
While some processes may be irreversible, several of the described cellular and organ level changes due to remodeling may be reversible by effective AF suppression or by targeting factors participating in remodeling.
Reverse electrical remodeling of the PV ostia and the LA can be observed as early as 2–3 months after the successful ablation of PeAF [141,142]. Reduction in atrial conduction time may correlate with reverse remodeling. The P wave to LA appendage conduction time, measured with transthoracic echocardiography, can be used to predict AF recurrence after ablation of PeAF, with cut-off values of >175 ms within 24 h of ablation (sensitivity 75%, specificity 78%), or >166 ms (sensitivity 75%, specificity 75.6%), when measured at 6 months follow up [143].
A long-term (7 year) MRI study of 42 patients who underwent ablation for PeAF confirmed that reverse LA remodeling occurs after successful ablation in most cases; however, progressive remodeling was observed despite the successful ablation in patients with BMI ≥ 30 or left ventricular hypertrophy [144]. The degree of reverse remodeling is more pronounced in patients with PeAF than in those with PAF [145,146]. Reverse remodeling of the right atrium and ventricle was also observed in an echocardiography study 15 ± 7 months after ablation [147]. Improvement in LV systolic function was also demonstrated after successful ablation of PeAF, even in patients with preserved left ventricular function: EF improved from 56.8 ± 9.8% to 65.1 ± 9.1% (p < 0.0001), along with strain measures [148].
Reverse remodeling of the mitral apparatus and decreased functional MR were observed in 45 patients who underwent successful ablation of PeAF and maintained SR 6 months after the procedure [149]. Patients with PeAF and functional MR respond to ablation better if the functional MR is due to atrial remodeling: The composite endpoint of all-cause death and heart failure hospitalization during the 2-year follow-up of 136 patients was more frequently observed in the ventricular functional MR than in the atrial functional MR group (22.6% vs. 3.7%, p < 0.0001) [150]. Surgical repair of class III-IV functional mitral regurgitation in 82 patients with PeAF was effective to improve heart failure symptoms and decrease LA and ventricular size over a 3-year follow-up. Concomitant surgical ablation to prevent recurrent MR, performed in 63% of patients, was effective with LA size <6 cm [151].
Reverse structural remodeling after successful ablation correlates with long-term arrhythmia-free survival: in a study of 140 patients with enlarged left atria (LA volume index ≥34 mL/m2), who had no AF recurrence 1 year after ablation, 54% had reverse remodeling with normal atrial size at 1 year. At a median follow-up of 44 months, 8% of patients who had reverse structural remodeling had AF recurrence, compared to 40% of patients with persistently enlarged left atria, p < 0.01. However, persistent LA functional abnormalities were observed even in patients where the atrial size decreased [152]. More extensive ablation, such as posterior left atrial wall isolation, is often used in patients with PeAF, and it did not prevent reverse remodeling when rhythm control was successful in a study of 105 patients [153].
Despite the rapidity of structural reverse remodeling after successful ablation, multiple markers of inflammation and collagen turnover show delayed or bimodal response, especially in patients where the AF relapses: in a study of 60 patients, hsCRP and IL-6 decreased after ablation but returned near to baseline levels by 12 months. TIMP-2 increased in patients where the AF did not recur but did not change substantially in the recurrence group. TGF-β1 increased in both groups, but the change was less pronounced in the recurrence group [154].
Medical management with antagonists of the RAAS system may be useful to promote reverse remodeling in addition to ablation. Sacubitril/valsartan was superior to valsartan alone in a randomized controlled study of 64 patients who underwent ablation for PAF or PeAF: atrial sizes decreased over a 24-week period (left: 4.3 ± 0.5 cm to 3.8 ± 0.5 cm, p < 0.001; right: 4.4 ± 0.8 cm to 3.9 ± 0.7 cm, p = 0.017), which was not observed in the valsartan group. The difference in clinical outcomes (AF recurrence, hospitalization, mortality), however, was not significant [155]. In a sheep model, eplerenone mitigated fibrosis and atrial dilatation and prolonged transition to PeAF in 26% of animals, but it did not prevent AF-induced electrical remodeling (rate of dominant frequency and ion channel densities) or AF persistence [156]. In a mouse model of PeAF, eicosapentaenoic acid and pitivastatin ameliorated adverse effects of tachypacing by affecting multiple pathways of electrical remodeling, thrombosis, and oxidative stress [157].
12. Potential Targets for the Treatment of Persistent Atrial Fibrillation
12.1. Simulation Models
Subcellular, cellular, tissue and organ-level models of PeAF have been developed to clarify the initiation, maintenance, and progression of AF. A future goal is to individualize AF therapy based on arrhythmia and substrate characteristics [158]. A mathematical model simulating human atrial cells and tissue analyzed the quantitative contributions of ionic current remodeling seen in PeAF to action potential shortening and rate adaptation. Ionic remodeling was simulated by reducing L-type Ca2+ channel current (ICa,L), increasing inward rectifier K+ current (IK1), and modulating five other ionic currents. A negative effect of Na+/Ca2+ exchange on action potential duration was only observed in PeAF but not in controls. Action potential rate adaptation was affected by IKur in control and by INaCa in the PeAF condition. In a 2D tissue model, a large reduction (70%) of ICaL became a dominant factor leading to a stable spiral wave in PeAF [159].
The heterogeneity of arrhythmia mechanisms maintaining AF affects the efficacy of the methods used for termination. In a simulation using 173 mathematical models, based on data obtained from remodeled human atrial tissue with intersubject variability, the relationship of ion currents, AF maintenance, and the dynamics of functional reentry were analyzed. Self-sustained reentries were maintained in 73% of the simulations; AF maintenance was associated with higher expressions of INa and ICa,L, with no significant differences in other currents. ICa,L blockade promoted AF termination in 30% of the self-sustained models [160].
The amount of fibrosis also correlated with the substrate of the arrhythmia mechanism in simulated atrial tissue: high fibrosis density generated a significant conduction velocity reduction, intensified vulnerability to multiple re-entries, and increased the amount of complex propagation patterns [161].
12.2. Pharmacological Targets
Targeting the arrhythmia mechanisms with ion channel blockers, even atrial selective agents, does not result in efficient rhythm control in patients with PeAF. Upstream therapies may be useful adjuncts. Based on data from simulation and translational studies, components of remodeling and its promoting extracardiac factors may be promising therapeutic targets [162].
The first-generation antiarrhythmics are non-selective (most of them affect multiple ion channels both in the atria and ventricles), with notable extracardiac side effects. Their poor efficacy, even in PAF and side effect profile, led to intense research in more selective agents, specifically targeting ion channels only present in the atrial myocardium: IKAch, IKur, or INa. Single-channel blockers were largely ineffective. Atrial-selective multichannel blockers, such as vernakalant (mostly IKur blocker, less effect on IKAc and INa), were used with success in acute settings for cardioversion. However, as antiarrhythmics only work on the electrophysiological pathways of AF, their efficacy in PeAF is even lower [163].
Due to the non-uniform remodeling of electrophysiological properties of different atrial structures, even targeting atrial selective ion channels may not have the desired homogenous effects. In a horse model of PeAF, the effects of the IK;Ca inhibitor NS8593 were studied using epicardial high-density contact mapping. The AF cycle length was prolonged. However, AF complexity increased due to the slowing of conduction, increased anisotropy, and electrical dissociation in the right atrium. AF complexity did not change in the left atrium, and cardioversion was not achieved [164].
Ca2+ homeostasis may provide a selective target for intervention. Downregulation of ICa, L caused by an alteration in RyR2 kinetics can be observed during atrial remodeling and facilitates triggered activity. Dantrolene, which decreases Ca2+ leak from RyR2, may suppress this arrhythmia mechanism, but no clinical data exists yet [163]. Bepridil, a nonselective Ca2+ channel blocker, can be used to assess the degree of remodeling and predict response to ablation: in 76 patients with PeAF, those who achieved cardioversion with bepridil (63.2%) prior to ablation had less low voltage areas noted during mapping (7.5% vs. 14.0%, p = 0.009) and 1-year AF recurrence-free survival was higher (87.1% vs. 62.3%, p = 0.03) [165]. In a canine model of persistent AF, a combination of bepridil with aprindine (a class Ib antiarrhythmic with atrial and ventricular effect) prevented pacing-induced atrial electrical remodeling [166].
Pharmaceutical agents affecting fibrosis may suppress this mechanism as part of the remodeling process. Large-scale clinical data are available with blockers of the RAAS: angiotensin-converting enzyme inhibitors, Ang II receptor blockers, or mineralocorticoid receptor blockers [163]. Combination with a statin may be even more effective. In a randomized trial of 189 patients with hypertension and non-permanent AF, treatment with fluvastatin and valsartan was more effective in reducing the recurrence rate in non-permanent AF (79.17%) and in delaying the progression to permanent AF (8.33%) during 24 months of follow up, compared to treatment with a calcium channel blocker (43.9% and 31.7%) or valsartan alone (70.5% and 14.3%) [167].
Sodium–glucose cotransporter-2 (SGLT2) inhibitors, used in the treatment of diabetes mellitus and cardiomyopathy/congestive heart failure, have a pleiotropic effect which may make them candidates for upstream therapy in AF. In a diabetic rat model, treatment with empagliflozin decreased the amount of diabetes-induced atrial remodeling, atrial fibrosis, and AF inducibility. In addition, the treatment also suppressed diabetes-induced mitochondrial changes (respiratory function, membrane potential, and biogenesis), likely via the peroxisome proliferator-activated receptor-c coactivator 1α/nuclear respiratory factor-1 (NRF-1)/mitochondrial transcription factor A (Tfam) signaling pathway [168]. In a canine model, canagliflozin suppressed rapid pacing-induced AF susceptibility, atrial fibrosis, and oxidative stress [169].
Connexin 40, which is an important part of gap junctions in the atria (but not the ventricles), may also become a target for pharmaceutical intervention: rotigaptide, a peptide antiarrhythmic, increases the conduction velocity in animal models. Decreasing inhomogenous conduction may act against reentry; however, these agents so far have shown no consistent benefit in AF [163].
12.3. Ablation of Persistent AF
Ablation directly affects the electroanatomical substrate and is very effective in the treatment of PAF. However, once remodeling occurs in PeAF, the efficacy decreases, and there is a need to identify patients in whom the remodeling processes play a greater role, and more extensive ablation is needed. Clinical risk scores may help to determine the degree of atrial remodeling. In 152 patients who underwent mapping and ablation of long-standing PeAF, CHA2DS2VASC ≥ 3, CAAP-AF ≥ 7, and DR-FLASH ≥ 4 predicted the presence of low voltage areas, while ATLAS score ≤ 7 predicted the absence of these [170]. LA stiffness, assessed by echocardiography, provided an additional tool to assess the risk of AF recurrence after ablation [171]. Recurrence of AF after epicardial, thoracoscopic ablation for PeAF was predicted by longer mean AF duration, larger left atrial volume, and higher BNP and SERCA levels in a study of 27 patients [172].
Quantification of electrical remodeling may help to guide ablation. In a simulated left atrial bilayer model of PeAF with fibrotic remodeling, machine learning (random forest classifier) was used to determine AF termination with combined ablation targets: PVI alone, box ablation, ablation of the largest or all fibrotic areas, ablation of the largest or all driver hotspots. The predictive value for ablation success was 0.73–0.83 with the addition of patient-specific and ablation pattern-specific lesion metrics [173]. In a study of 118 patients with PeAF, those assigned to a computational modeling-guided ablation (additional lesion sets beyond empirical ablation) had better long-term outcomes than the empirical group: clinical recurrence rate was significantly lower, 20.8% vs. 40.0%, p = 0.042. There were no differences in procedural time or complications [174].
Increased right atrial conduction time may show remodeling and predict recurrence of AF after ablation: in a study of 155 patients, long high right atrium to coronary sinus conduction time (>151 ms) was a significant risk factor for the recurrence of atrial tachyarrhythmia [175]. Inter-atrial conduction time ≥123 ms had a similar predictive value for AF recurrence after PVI for persistent AF [176].
12.4. Cardiac Resynchronization Therapy
Cardiac resynchronization therapy with biventricular pacing (CRT) is a well-established treatment modality for patients with congestive heart failure, severe cardiomyopathy, and intraventricular conduction delay. Reverse cardiac remodeling, including improvement in the parameters of atrial myopathy, has been observed in responders [139]. These, however, did not consistently predict lower AF incidence in the CRT trials. Even when LV reverse remodeling is observed, the LA may not be affected to the same degree: while >40% of responder patients achieved complete reverse remodeling (LV and LA) in the MADIT-CRT trial, 11% only had incomplete LA reverse remodeling, despite good LV response [177]. Reverse electrical remodeling (reduction of P wave duration, 142.7 ms vs. 133.1 ms; p < 0.001) was observed in a study of 41 CRT patients with >92% biventricular pacing over an average of 55 months follow-up, which did not correlate with the time to AF onset after CRT implant [178].
AF may interfere with CRT due to rapidly conducted beats, which leads to decreased amount of biventricular pacing. The presence of an atrioventricular block (either due to underlying disease or atrioventricular node ablation) may promote CRT. In a study of 75 patients with CRT and complete atrioventricular block, reverse LA remodeling and decreased duration of AF episodes were observed during a 2-year follow-up: average LA diameter decreased from 52 to 48 mm (p < 0.001), the ratio of patients experiencing AF episodes >48 h duration decreased from 76% (first 3 months of CRT) to 33% at the end of the follow up [179].
13. Conclusions
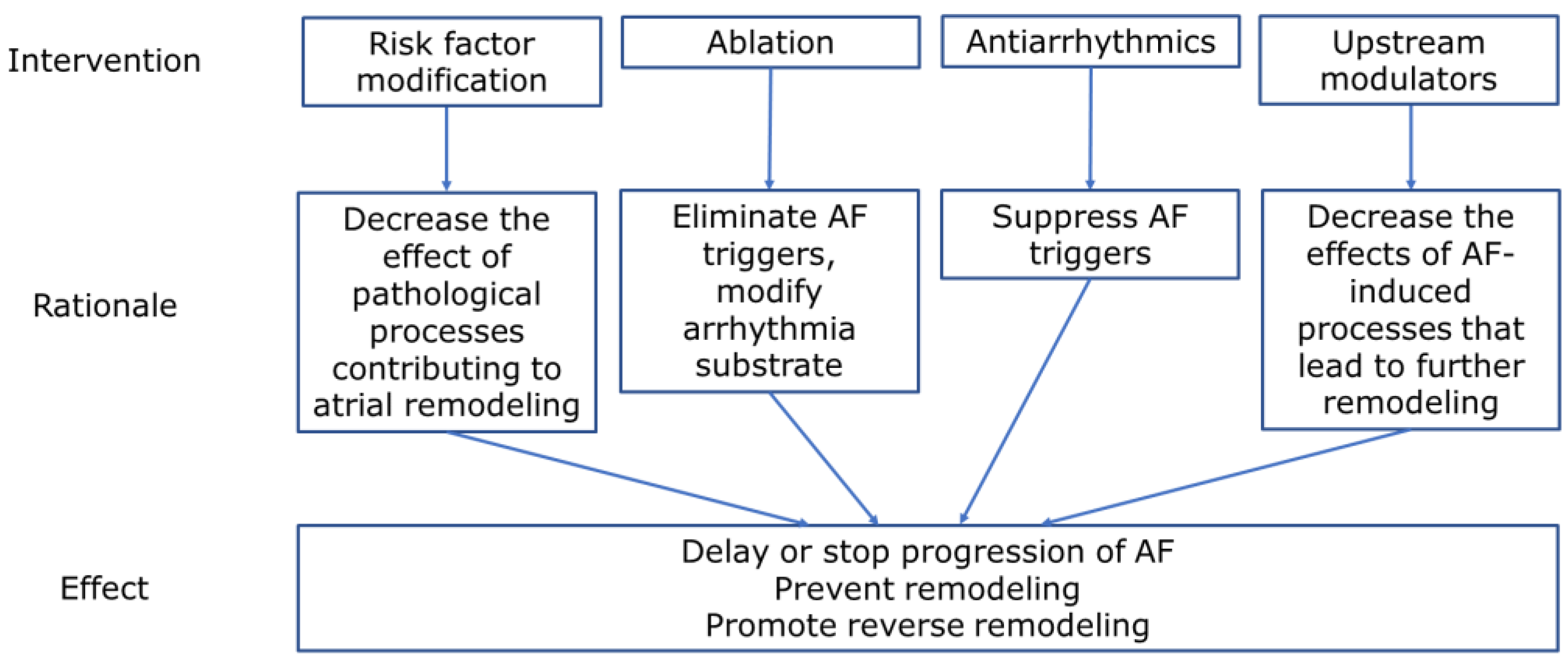
Abundant data now suggests that atrial/cardiac remodeling in PeAF occurs on genetic, biochemical/metabolic (cellular), and organ levels. Even extracardiac organ systems are involved and lead to systemic activation of processes, affecting cardiac remodeling. We have reviewed the novel pathophysiological findings in these processes, including extracardiac factors, models developed to study remodeling, reversibility of changes, and potential targets for therapies to prevent or reverse remodeling. The findings highlight the complexity of the pathophysiology—persistent AF is not just an ‘electrical disease,’ and its treatment should involve all aspects of its pathophysiology (Figure 6).
Important findings from recent studies highlighted the complexity of the AF progression process and the role of remodeling in the transformation from PAF to PeAF. As electrical remodeling does not affect all atrial components uniformly, it is unlikely that effective treatment can be achieved only by using antiarrhythmic medications targeting ion channels. Components of atrial remodeling can be targeted using existing medications: the RAAS system is a very promising target for upstream therapies, and statins had a synergistic effect in experimental studies. SGPLT2 inhibitors may affect multiple metabolic processes in atrial remodeling. As the anticoagulation pathway also has a role in the remodeling process, factor Xa inhibitors may modulate this risk, expanding the indication for anticoagulation. Simulation models, based on individual patient characteristics, may help to assess the degree of remodeling and identify targets for ablation beyond pulmonary vein isolation.
As the global burden of AF will continue to increase due to the high prevalence of its risk factors and the aging population, the demand will remain high for basic, translational, and clinical studies to clarify the pathophysiology of AF progression and to develop cost-effective treatment options.
14. Limitations
We used the PRISMA guidelines for the literature search (Supplementary Materials). Due to the very heterogenous study designs noted after screening the results of the search (multiple pathophysiological processes studied using in silico, in vitro, and in vivo studies, in multiple species), structured statistical meta-analysis was not feasible.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/physiologia3010004/s1, Table S1: PRISMA-P checklist [180].
Author Contributions
Conceptualization, A.R.; writing—original draft preparation, I.B.; writing review and editing, A.R. and I.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Publicly available datasets were analyzed in this study (Pubmed, Web of Science, and Cochrane electronic databases). https://pubmed.ncbi.nlm.nih.gov. https://www.webofknowledge.com. https://www.cochranelibrary.com.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hindricks, G.; Potpara, T.; Dagres, N.; Arbelo, E.; Bax, J.J.; Blomström-Lundqvist, C.; Boriani, G.; Castella, M.; Dan, G.-A.; Dilaveris, P.E.; et al. 2020 ESC Guidelines for the diagnosis and management of atrial fibrillation developed in collaboration with the european association for cardio-thoracic surgery (EACTS). Eur. Heart J. 2021, 42, 373–498. [Google Scholar] [CrossRef] [PubMed]
- Haïssaguerre, M.; Jaïs, P.; Shah, D.C.; Takahashi, A.; Hocini, M.; Quiniou, G.; Garrigue, S.; le Mouroux, A.; le Métayer, P.; Clémenty, J. Spontaneous initiation of atrial fibrillation by ectopic beats originating in the pulmonary veins. N. Engl. J. Med. 1998, 339, 659–666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wijffels, M.C.E.F.; Kirchhof, C.J.H.J.; Dorland, R.; Allessie, M.A. Atrial fibrillation begets atrial fibrillation. Circulation 1995, 92, 1954–1968. [Google Scholar] [CrossRef] [PubMed]
- Moher, D.; Shamseer, L.; Clarke, M.; Ghersi, D.; Liberati, A.; Petticrew, M.; Shekelle, P.; Stewart, L.A. Preferred reporting items for systematic review and meta-analysis protocols (PRISMA-P) 2015 Statement. Syst. Rev. 2015, 4, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campbell, M.; McKenzie, J.E.; Sowden, A.; Katikireddi, S.V.; Brennan, S.E.; Ellis, S.; Hartmann-Boyce, J.; Ryan, R.; Shepperd, S.; Thomas, J.; et al. Synthesis without meta-analysis (SWiM) in systematic reviews: Reporting guideline. BMJ 2020, 368, l6890. [Google Scholar] [CrossRef] [Green Version]
- Vlachos, K.; Letsas, K.P.; Korantzopoulos, P.; Liu, T.; Georgopoulos, S.; Bakalakos, A.; Karamichalakis, N.; Xydonas, S.; Efremidis, M.; Sideris, A. Prediction of atrial fibrillation development and progression: Current perspectives. World J. Cardiol. 2016, 8, 267–276. [Google Scholar] [CrossRef]
- Nguyen, B.-O.; Weberndorfer, V.; Crijns, H.J.; Geelhoed, B.; ten Cate, H.; Spronk, H.; Kroon, A.; de With, R.; Al-Jazairi, M.; Maass, A.H.; et al. Prevalence and determinants of atrial fibrillation progression in paroxysmal atrial fibrillation. Heart 2022, in press. [Google Scholar] [CrossRef]
- Lillo-Castellano, J.M.; González-Ferrer, J.J.; Marina-Breysse, M.; Martínez-Ferrer, J.B.; Pérez-Álvarez, L.; Alzueta, J.; Martínez, J.G.; Rodríguez, A.; Rodríguez-Pérez, J.C.; Anguera, I.; et al. Personalized monitoring of electrical remodelling during atrial fibrillation progression via remote transmissions from implantable devices. EP Eur. 2020, 22, 704–715. [Google Scholar] [CrossRef]
- Jalife, J.; Kaur, K. Atrial remodeling, fibrosis, and atrial fibrillation. Trends Cardiovasc. Med. 2015, 25, 475–484. [Google Scholar] [CrossRef] [Green Version]
- Sánchez, J.; Gomez, J.F.; Martinez-Mateu, L.; Romero, L.; Saiz, J.; Trenor, B. Heterogeneous effects of fibroblast-myocyte coupling in different regions of the human atria under conditions of atrial fibrillation. Front. Physiol. 2019, 10, 847. [Google Scholar] [CrossRef]
- Cheniti, G.; Vlachos, K.; Pambrun, T.; Hooks, D.; Frontera, A.; Takigawa, M.; Bourier, F.; Kitamura, T.; Lam, A.; Martin, C.; et al. Atrial fibrillation mechanisms and implications for catheter ablation. Front. Physiol. 2018, 9, 1458. [Google Scholar] [CrossRef] [Green Version]
- Buhl, R.; Hesselkilde, E.M.; Carstensen, H.; Hopster-Iversen, C.; van Loon, G.; Decloedt, A.; van Steenkiste, G.; Marr, C.M.; Reef, V.B.; Schwarzwald, C.C.; et al. Atrial fibrillatory rate as predictor of recurrence of atrial fibrillation in horses treated medically or with electrical cardioversion. Equine Vet. J. 2022, 54, 1013–1022. [Google Scholar] [CrossRef]
- Luca, A.; Pittet, A.; Buttu, A.; McCann, A.; Vesin, J.-M.; Pascale, P.; le Bloa, M.; Herrera, C.; Park, C.-I.; Rollin, A.; et al. Severe and uniform bi-atrial remodeling measured by dominant frequency analysis in persistent atrial fibrillation unresponsive to ablation. J. Interv. Card. Electrophysiol. 2020, 59, 431–440. [Google Scholar] [CrossRef]
- Martins, R.P.; Kaur, K.; Hwang, E.; Ramirez, R.J.; Willis, B.C.; Filgueiras-Rama, D.; Ennis, S.R.; Takemoto, Y.; Ponce-Balbuena, D.; Zarzoso, M.; et al. Dominant frequency increase rate predicts transition from paroxysmal to long-term persistent atrial fibrillation. Circulation 2014, 129, 1472–1482. [Google Scholar] [CrossRef]
- Angel, N.; Kholmovski, E.G.; Ghafoori, E.; Dosdall, D.J.; MacLeod, R.S.; Ranjan, R. Regions of high dominant frequency in chronic atrial fibrillation anchored to areas of atrial fibrosis. Proc. Comput. Cardiol. 2019, 46, 10. [Google Scholar]
- Williams, S.E.; O’Neill, L.; Roney, C.H.; Julia, J.; Metzner, A.; Reißmann, B.; Mukherjee, R.K.; Sim, I.; Whitaker, J.; Wright, M.; et al. Left atrial effective conducting size predicts atrial fibrillation vulnerability in persistent but not paroxysmal atrial fibrillation. J. Cardiovasc. Electrophysiol. 2019, 30, 1416–1427. [Google Scholar] [CrossRef] [Green Version]
- Honarbakhsh, S.; Schilling, R.J.; Orini, M.; Providencia, R.; Keating, E.; Finlay, M.; Sporton, S.; Chow, A.; Earley, M.J.; Lambiase, P.D.; et al. Structural remodeling and conduction velocity dynamics in the human left atrium: Relationship with reentrant mechanisms sustaining atrial fibrillation. Heart Rhythm 2019, 16, 18–25. [Google Scholar] [CrossRef] [Green Version]
- Ohguchi, S.; Inden, Y.; Yanagisawa, S.; Shigematsu, T.; Yasuda, K.; Katagiri, K.; Oguri, M.; Murohara, T. Long P-wave duration immediately after pulmonary vein isolation on radiofrequency catheter ablation for atrial fibrillation predicts clinical recurrence: Correlation with atrial remodeling in persistent atrial fibrillation. Heart Vessel. 2022, 37, 476–488. [Google Scholar] [CrossRef]
- Walters, T.E.; Lee, G.; Lee, A.; Sievers, R.; Kalman, J.M.; Gerstenfeld, E.P. Site-specific epicardium-to-endocardium dissociation of electrical activation in a swine model of atrial fibrillation. JACC Clin. Electrophysiol. 2020, 6, 830–845. [Google Scholar] [CrossRef]
- Prabhu, S.; Voskoboinik, A.; McLellan, A.J.A.; Peck, K.Y.; Pathik, B.; Nalliah, C.J.; Wong, G.R.; Azzopardi, S.M.; Lee, G.; Mariani, J.; et al. A Comparison of the electrophysiologic and electroanatomic characteristics between the right and left atrium in persistent atrial fibrillation: Is the right atrium a window into the left? J. Cardiovasc. Electrophysiol. 2017, 28, 1109–1116. [Google Scholar] [CrossRef]
- Parikh, R.R.; Norby, F.L.; Wang, W.; Thenappan, T.; Prins, K.W.; Van’t Hof, J.R.; Lutsey, P.L.; Solomon, S.D.; Shah, A.M.; Chen, L.Y. Association of right ventricular afterload with atrial fibrillation risk in older adults. Chest 2022, 162, 884–893. [Google Scholar] [CrossRef] [PubMed]
- Sairaku, A.; Nakano, Y.; Suenari, K.; Tokuyama, T.; Kawazoe, H.; Matsumura, H.; Tomomori, S.; Amioka, M.; Kihara, Y. Electrical remodeling of the atrioventricular node caused by persistent atrial fibrillation in humans. J. Cardiovasc. Electrophysiol. 2016, 27, 918–922. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Shim, J.; Uhm, J.-S.; Joung, B.; Lee, M.-H.; Pak, H.-N. Post-shock sinus node recovery time is an independent predictor of recurrence after catheter ablation of longstanding persistent atrial fibrillation. Int. J. Cardiol. 2013, 168, 1937–1942. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, K.; Nagata, Y.; Nitta, G.; Okata, S.; Nagase, M.; Miyazaki, R.; Nagamine, S.; Kaneko, M.; Lee, T.; Nozato, T.; et al. Impact of the sinus node recovery time after termination of atrial fibrillation during catheter ablation on clinical outcomes in patients with persistent atrial fibrillation. PLoS ONE 2021, 16, e0259750. [Google Scholar] [CrossRef]
- Steenman, M. Insight into atrial fibrillation through analysis of the coding transcriptome in humans. Biophys. Rev. 2020, 12, 817–826. [Google Scholar] [CrossRef]
- Wong, G.R.; Nalliah, C.J.; Lee, G.; Voskoboinik, A.; Prabhu, S.; Parameswaran, R.; Sugumar, H.; Anderson, R.D.; Ling, L.-H.; McLellan, A.; et al. Genetic Susceptibility to atrial fibrillation is associated with atrial electrical remodeling and adverse post-ablation outcome. JACC Clin. Electrophysiol. 2020, 6, 1509–1521. [Google Scholar] [CrossRef]
- Husser, D.; Büttner, P.; Ueberham, L.; Dinov, B.; Sommer, P.; Arya, A.; Hindricks, G.; Bollmann, A. Association of atrial fibrillation susceptibility genes, atrial fibrillation phenotypes and response to catheter ablation: A Gene-based analysis of GWAS data. J. Transl. Med. 2017, 15, 71. [Google Scholar] [CrossRef] [Green Version]
- Husser, D.; Ueberham, L.; Dinov, B.; Kosiuk, J.; Kornej, J.; Hindricks, G.; Shoemaker, M.B.; Roden, D.M.; Bollmann, A.; Büttner, P. Genomic contributors to atrial electroanatomical remodeling and atrial fibrillation progression: Pathway enrichment analysis of GWAS data. Sci. Rep. 2016, 6, 36630. [Google Scholar] [CrossRef] [Green Version]
- Ezeani, M.; Prabhu, S. PI3K(P110α) as a determinant and gene therapy for atrial enlargement in atrial fibrillation. Mol. Cell. Biochem. 2022, in press. [Google Scholar] [CrossRef]
- Deshmukh, A.; Barnard, J.; Sun, H.; Newton, D.; Castel, L.; Pettersson, G.; Johnston, D.; Roselli, E.; Gillinov, A.M.; McCurry, K.; et al. Left atrial transcriptional changes associated with atrial fibrillation susceptibility and persistence. Circ. Arrhythm. Electrophysiol. 2015, 8, 32–41. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.-L.; Chuang, J.-H.; Wang, H.-T.; Chen, H.-C.; Liu, W.-H.; Yang, M.-Y. Altered expression of circadian clock genes in patients with atrial fibrillation is associated with atrial high-rate episodes and left atrial remodeling. Diagnostics 2021, 11, 90. [Google Scholar] [CrossRef]
- Huang, L.; Yu, H.; Fan, X.; Li, X.; Mao, L.; Cheng, J.; Zeng, X.; Dang, X. A potential role of esophageal cancer related Gene-4 for atrial fibrillation. Sci. Rep. 2017, 7, 2717. [Google Scholar] [CrossRef] [Green Version]
- Oh, Y.; Yang, S.; Liu, X.; Jana, S.; Izaddoustdar, F.; Gao, X.; Debi, R.; Kim, D.-K.; Kim, K.-H.; Yang, P.; et al. Transcriptomic bioinformatic analyses of atria uncover involvement of pathways related to strain and post-translational modification of collagen in increased atrial fibrillation vulnerability in intensely exercised mice. Front. Physiol. 2020, 11, 605671. [Google Scholar] [CrossRef]
- Jiang, Y.-Y.; Hou, H.-T.; Yang, Q.; Liu, X.-C.; He, G.-W. Chloride channels are involved in the development of atrial fibrillation—A transcriptomic and proteomic study. Sci. Rep. 2017, 7, 10215. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Bektik, E.; Sakon, P.; Wang, X.; Huang, S.; Meng, X.; Chen, M.; Han, W.; Chen, J.; Wang, Y.; et al. Integrated analysis of the MicroRNA–MRNA network predicts potential regulators of atrial fibrillation in humans. Cells 2022, 11, 2629. [Google Scholar] [CrossRef]
- Çubukçuoğlu Deniz, G.; Durdu, S.; Doğan, Y.; Erdemli, E.; Özdağ, H.; Akar, A.R. Molecular signatures of human chronic atrial fibrillation in primary mitral regurgitation. Cardiovasc. Ther. 2021, 2021, 5516185. [Google Scholar] [CrossRef]
- Alvarez-Franco, A.; Rouco, R.; Ramirez, R.J.; Guerrero-Serna, G.; Tiana, M.; Cogliati, S.; Kaur, K.; Saeed, M.; Magni, R.; Enriquez, J.A.; et al. Transcriptome and proteome mapping in the sheep atria reveal molecular featurets of atrial fibrillation progression. Cardiovasc. Res. 2021, 117, 1760–1775. [Google Scholar] [CrossRef]
- Liu, B.; Li, X.; Zhao, C.; Wang, Y.; Lv, M.; Shi, X.; Han, C.; Pandey, P.; Qian, C.; Guo, C.; et al. Proteomic analysis of atrial appendages revealed the pathophysiological changes of atrial fibrillation. Front. Physiol. 2020, 11, 573433. [Google Scholar] [CrossRef]
- Gao, Y.; Liu, Y.; Fu, Y.; Wang, Q.; Liu, Z.; Hu, R.; Yang, X.; Chen, M. The potential regulatory role of hsa_circ_0004104 in the persistency of atrial fibrillation by promoting cardiac fibrosis via TGF-β pathway. BMC Cardiovasc. Disord. 2021, 21, 25. [Google Scholar] [CrossRef]
- Liu, L.; Chen, Y.; Shu, J.; Tang, C.-E.; Jiang, Y.; Luo, F. Identification of MicroRNAs enriched in exosomes in human pericardial fluid of patients with atrial fibrillation based on bioinformatic analysis. J. Thorac. Dis. 2020, 12, 5617–5627. [Google Scholar] [CrossRef]
- Galenko, O.; Jacobs, V.; Knight, S.; Taylor, M.; Cutler, M.J.; Muhlestein, J.B.; Carlquist, J.L.; Knowlton, K.U.; Jared Bunch, T. The role of MicroRNAs in the development, regulation, and treatment of atrial fibrillation. J. Int. Card. Electrophysiol. 2019, 55, 297–305. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Zhang, F.; Zhang, Y.-L.; Yang, X.-C. Relationship between circulating MiRNA-21, atrial fibrosis, and atrial fibrillation in patients with atrial enlargement. Ann. Palliat. Med. 2021, 10, 12742–12749. [Google Scholar] [CrossRef] [PubMed]
- Morishima, M.; Iwata, E.; Nakada, C.; Tsukamoto, Y.; Takanari, H.; Miyamoto, S.; Moriyama, M.; Ono, K. Atrial fibrillation-mediated upregulation of MiR-30d regulates myocardial electrical remodeling of the G-protein-gated K+ channel, IK.ACh. Circ. J. 2016, 80, 1346–1355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, F.; Ren, W.; Zhang, X.; Wu, P.; Fan, J. MiR-425-5p is negatively associated with atrial fibrosis and promotes atrial remodeling by targeting CREB1 in atrial fibrillation. J. Cardiol. 2022, 79, 202–210. [Google Scholar] [CrossRef]
- Jie, Q.-Q.; Li, G.; Duan, J.-B.; Li, X.-B.; Yang, W.; Chu, Y.-P.; Yu, S.-D.; Liu, X.-Y.; Wang, C.-Y.; Liu, F.-F.; et al. Remodeling of myocardial energy and metabolic homeostasis in a sheep model of persistent atrial fibrillation. Biochem. Biophys. Res. Commun. 2019, 517, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Muszyński, P.; Bonda, T.A. Mitochondrial dysfunction in atrial fibrillation—Mechanisms and pharmacological interventions. J. Clin. Med. 2021, 10, 2385. [Google Scholar] [CrossRef] [PubMed]
- Watts, M.; Kolluru, G.K.; Dherange, P.; Pardue, S.; Si, M.; Shen, X.; Trosclair, K.; Glawe, J.; Al-Yafeai, Z.; Iqbal, M.; et al. Decreased bioavailability of hydrogen sulfide links vascular endothelium and atrial remodeling in atrial fibrillation. Redox Biol. 2021, 38, 101817. [Google Scholar] [CrossRef]
- Xue, X.; Ling, X.; Xi, W.; Wang, P.; Sun, J.; Yang, Q.; Xiao, J. Exogenous hydrogen sulfide reduces atrial remodeling and atrial fibrillation induced by diabetes mellitus via activation of the PI3K/Akt/ENOS pathway. Mol. Med. Rep. 2020, 22, 1759–1766. [Google Scholar] [CrossRef]
- Avula, U.M.R.; Dridi, H.; Chen, B.; Yuan, Q.; Katchman, A.N.; Reiken, S.R.; Desai, A.D.; Parsons, S.; Baksh, H.; Ma, E.; et al. Attenuating persistent sodium current-induced atrial myopathy and fibrillation by preventing mitochondrial oxidative stress. JCI Insight 2021, 6, e147371. [Google Scholar] [CrossRef]
- Dolce, B.; Christ, T.; Grammatika Pavlidou, N.; Yildirim, Y.; Reichenspurner, H.; Eschenhagen, T.; Nikolaev, V.O.; Kaumann, A.J.; Molina, C.E. Impact of phosphodiesterases PDE3 and PDE4 on 5-hydroxytryptamine receptor4-mediated increase of CAMP in human atrial fibrillation. Naunyn Schmiedebergs Arch. Pharmacol. 2021, 394, 291–298. [Google Scholar] [CrossRef]
- Chang, K.C.; Trayanova, N.A. Mechanisms of arrhythmogenesis related to calcium-driven alternans in a model of human atrial fibrillation. Sci. Rep. 2016, 6, 36395. [Google Scholar] [CrossRef]
- Onal, B.; Gratz, D.; Hund, T.J. Ca2+/Calmodulin-dependent Kinase II-dependent regulation of atrial myocyte late Na+ current, Ca2+ cycling, and excitability: A mathematical modeling study. Am. J. Physiol. Heart Circ. Physiol. 2017, 313, H1227–H1239. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Finet, J.E.; Wolfram, J.A.; Anderson, M.E.; Ai, X.; Donahue, J.K. Calcium/Calmodulin-dependent protein Kinase II causes atrial structural remodeling associated with atrial fibrillation and heart failure. Heart Rhythm 2019, 16, 1080–1088. [Google Scholar] [CrossRef]
- Munro, M.L.; van Hout, I.; Aitken-Buck, H.M.; Sugunesegran, R.; Bhagwat, K.; Davis, P.J.; Lamberts, R.R.; Coffey, S.; Soeller, C.; Jones, P.P. Human atrial fibrillation is not associated with remodeling of ryanodine receptor clusters. Front. Cell Dev. Biol. 2021, 9, 633704. [Google Scholar] [CrossRef]
- Hu, X.; van Marion, D.M.S.; Wiersma, M.; Zhang, D.; Brundel, B.J.J.M. The protective role of small heat shock proteins in cardiac diseases: Key role in atrial fibrillation. Cell Stress Chaperones 2017, 22, 665–674. [Google Scholar] [CrossRef] [Green Version]
- Wiersma, M.; Meijering, R.A.M.; Qi, X.; Zhang, D.; Liu, T.; Hoogstra-Berends, F.; Sibon, O.C.M.; Henning, R.H.; Nattel, S.; Brundel, B.J.J.M. Endoplasmic reticulum stress is associated with autophagy and cardiomyocyte remodeling in experimental and human atrial fibrillation. J. Am. Heart Assoc. 2017, 6, e006458. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Y.; Zhao, J.; Gong, Y.; Wang, D.; Wang, X.; Yun, F.; Liu, Z.; Zhang, S.; Li, W.; Zhao, X.; et al. Autophagy exacerbates electrical remodeling in atrial fibrillation by Ubiquitin-dependent degradation of L-type calcium channel. Cell Death Dis. 2018, 9, 873. [Google Scholar] [CrossRef] [Green Version]
- Adeniran, I.; MacIver, D.H.; Garratt, C.J.; Ye, J.; Hancox, J.C.; Zhang, H. Effects of persistent atrial fibrillation-induced electrical remodeling on atrial electro-mechanics—Insights from a 3D model of the human atria. PLoS ONE 2015, 10, e0142397. [Google Scholar] [CrossRef] [Green Version]
- Wałek, P.; Sielski, J.; Gorczyca, I.; Roskal-Wałek, J.; Starzyk, K.; Jaskulska-Niedziela, E.; Bartkowiak, R.; Wożakowska-Kapłon, B. Left atrial mechanical remodelling assessed as the velocity of left atrium appendage wall motion during atrial fibrillation is associated with maintenance of sinus rhythm after electrical cardioversion in patients with persistent atrial fibrillation. PLoS ONE 2020, 15, e0228239. [Google Scholar] [CrossRef]
- Kaufmann, R.; Rezar, R.; Strohmer, B.; Wernly, B.; Lichtenauer, M.; Hitzl, W.; Meissnitzer, M.; Hergan, K.; Granitz, M. Left atrial ejection fraction assessed by prior cardiac CT predicts recurrence of atrial fibrillation after pulmonary vein isolation. J. Clin. Med. 2021, 10, 752. [Google Scholar] [CrossRef]
- Zuo, K.; Li, K.; Liu, M.; Li, J.; Liu, X.; Liu, X.; Zhong, J.; Yang, X. Correlation of left atrial wall thickness and atrial remodeling in atrial fibrillation. Medicine 2019, 98, e15170. [Google Scholar] [CrossRef] [PubMed]
- Verheule, S.; Eckstein, J.; Linz, D.; Maesen, B.; Bidar, E.; Gharaviri, A.; Schotten, U. Role of endo-epicardial dissociation of electrical activity and transmural conduction in the development of persistent atrial fibrillation. Prog. Biophys. Mol. Biol. 2014, 115, 173–185. [Google Scholar] [CrossRef] [PubMed]
- Maesen, B.; Zeemering, S.; Afonso, C.; Eckstein, J.; Burton, R.A.B.; van Hunnik, A.; Stuckey, D.J.; Tyler, D.; Maessen, J.; Grau, V.; et al. Rearrangement of atrial bundle architecture and consequent changes in anisotropy of conduction constitute the 3-dimensional substrate for atrial fibrillation. Circ. Arrhythm. Electrophysiol. 2013, 6, 967–975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mouws, E.M.J.P.; Lanters, E.A.H.; Teuwen, C.P.; van der Does, L.J.M.E.; Kik, C.; Knops, P.; Bekkers, J.A.; Bogers, A.J.J.C.; de Groot, N.M.S. Epicardial breakthrough waves during sinus rhythm. Circ. Arrhythm. Electrophysiol. 2017, 10, e005145. [Google Scholar] [CrossRef] [PubMed]
- Lim, H.E.; Na, J.O.; Im, S.I.; Choi, C.U.; Kim, S.H.; Kim, J.W.; Kim, E.J.; Han, S.W.; Rha, S.-W.; Park, C.G.; et al. Interatrial septal thickness as a marker of structural and functional remodeling of the left atrium in patients with atrial fibrillation. Korean J. Intern. Med. 2015, 30, 808–820. [Google Scholar] [CrossRef] [Green Version]
- Park, Y.M.; Park, H.C.; Ban, J.-E.; Choi, J.-I.; Lim, H.E.; Park, S.W.; Kim, Y.-H. Interatrial septal thickness is associated with the extent of left atrial complex fractionated atrial electrograms and acute procedural outcome in patients with persistent atrial fibrillation. Europace 2015, 17, 1700–1707. [Google Scholar] [CrossRef]
- Xie, E.; Yu, R.; Ambale-Venkatesh, B.; Bakhshi, H.; Heckbert, S.R.; Soliman, E.Z.; Bluemke, D.A.; Kawut, S.M.; Wu, C.O.; Nazarian, S.; et al. Association of right atrial structure with incident atrial fibrillation: A longitudinal cohort cardiovascular magnetic resonance study from the multi-ethnic study of atherosclerosis (MESA). J. Cardiovasc. Magn. Reson. 2020, 22, 36. [Google Scholar] [CrossRef]
- Kishima, H.; Mine, T.; Takahashi, S.; Ashida, K.; Ishihara, M.; Masuyama, T. Morphologic remodeling of left atrial appendage in patients with atrial fibrillation. Heart Rhythm 2016, 13, 1823–1828. [Google Scholar] [CrossRef]
- Platonov, P.G. Atrial fibrosis: An obligatory component of arrhythmia mechanisms in atrial fibrillation? J. Geriatr. Cardiol. 2017, 14, 233–237. [Google Scholar] [CrossRef]
- Hopman, L.H.G.A.; Mulder, M.J.; van der Laan, A.M.; Demirkiran, A.; Bhagirath, P.; van Rossum, A.C.; Allaart, C.P.; Götte, M.J.W. Impaired left atrial reservoir and conduit strain in patients with atrial fibrillation and extensive left atrial fibrosis. J. Cardiovasc. Magn. Reson. 2021, 23, 131. [Google Scholar] [CrossRef]
- Kato, T.; Sekiguchi, A.; Sagara, K.; Tanabe, H.; Takamura, M.; Kaneko, S.; Aizawa, T.; Fu, L.-T.; Yamashita, T. Endothelial–mesenchymal transition in human atrial fibrillation. J. Cardiol. 2017, 69, 706–711. [Google Scholar] [CrossRef]
- Van den Berg, N.W.E.; Kawasaki, M.; Fabrizi, B.; Nariswari, F.A.; Verduijn, A.C.; Neefs, J.; Wesselink, R.; Al-Shama, R.F.M.; van der Wal, A.C.; de Boer, O.J.; et al. Epicardial and endothelial cell activation concurs with extracellular matrix remodeling in atrial fibrillation. Clin. Transl. Med. 2021, 11, e558. [Google Scholar] [CrossRef]
- Benito, E.M.; Cabanelas, N.; Nuñez-Garcia, M.; Alarcón, F.; Figueras, I.; Ventura, R.M.; Soto-Iglesias, D.; Guasch, E.; Prat-Gonzalez, S.; Perea, R.J.; et al. Preferential regional distribution of atrial fibrosis in posterior wall around left inferior pulmonary vein as identified by late gadolinium enhancement cardiac magnetic resonance in patients with atrial fibrillation. EP Eur. 2018, 20, 1959–1965. [Google Scholar] [CrossRef]
- Kogawa, R.; Okumura, Y.; Watanabe, I.; Nagashima, K.; Takahashi, K.; Iso, K.; Watanabe, R.; Arai, M.; Kurokawa, S.; Ohkubo, K.; et al. Left atrial remodeling: Regional differences between paroxysmal and persistent atrial fibrillation. J. Arrhythm. 2017, 33, 483–487. [Google Scholar] [CrossRef]
- Callegari, S.; Macchi, E.; Monaco, R.; Magnani, L.; Tafuni, A.; Croci, S.; Nicastro, M.; Garrapa, V.; Banchini, A.; Becchi, G.; et al. Clinicopathological bird’s-eye view of left atrial myocardial fibrosis in 121 patients with persistent atrial fibrillation. Circ. Arrhythm. Electrophysiol. 2020, 13, e007588. [Google Scholar] [CrossRef]
- Maesen, B.; Verheule, S.; Zeemering, S.; la Meir, M.; Nijs, J.; Lumeij, S.; Lau, D.H.; Granier, M.; Crijns, H.J.; Maessen, J.G.; et al. Endomysial fibrosis, rather than overall connective tissue content, is the main determinant of conduction disturbances in human atrial fibrillation. EP Eur. 2022, 24, 1015–1024. [Google Scholar] [CrossRef]
- Sánchez, J.; Trenor, B.; Saiz, J.; Dössel, O.; Loewe, A. Fibrotic remodeling during persistent atrial fibrillation: In silico investigation of the role of calcium for human atrial myofibroblast electrophysiology. Cells 2021, 10, 2852. [Google Scholar] [CrossRef]
- Szilágyi, J.; Sághy, L. Atrial remodeling in atrial fibrillation. Comorbidities and markers of disease progression predict catheter ablation outcome. Curr. Cardiol. Rev. 2021, 17, 217–229. [Google Scholar] [CrossRef]
- Musa, H.; Kaur, K.; O’Connell, R.; Klos, M.; Guerrero-Serna, G.; Avula, U.M.R.; Herron, T.J.; Kalifa, J.; Anumonwo, J.M.B.; Jalife, J. Inhibition of platelet-derived growth factor-AB signaling prevents electromechanical remodeling of adult atrial myocytes that contact myofibroblasts. Heart Rhythm 2013, 10, 1044–1051. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Guo, Y.; Chen, Q.; Cheng, X.; Xiang, G.; Chen, M.; Wu, H.; Huang, Q.; Zhu, P.; Zhang, J. TGFβ1 and HGF regulate CTGF expression in human atrial fibroblasts and are involved in atrial remodelling in patients with rheumatic heart disease. J. Cell. Mol. Med. 2019, 23, 3032–3039. [Google Scholar] [CrossRef] [Green Version]
- Sai, C.; Yunhan, J.; Zhao, J.; Yu, Z.; Yun, Z.; Zhezhe, C.; Fuqin, T.; Yingbin, X.; Ruiyan, M. Cyclin dependent kinase 1 (CDK1) activates cardiac fibroblasts via directly phosphorylating paxillin at Ser244. Int. Heart J. 2019, 60, 374–383. [Google Scholar] [CrossRef] [PubMed]
- Ciuffo, L.A.; Lima, J.; Vasconcellos, H.D.; de Balouch, M.; Tao, S.; Nazarian, S.; Spragg, D.D.; Marine, J.E.; Berger, R.D.; Calkins, H.; et al. Intra-atrial dyssynchrony using cardiac magnetic resonance to quantify tissue remodeling in patients with atrial fibrillation. Arq. Bras. Cardiol. 2019, 4, 441–450. [Google Scholar] [CrossRef] [PubMed]
- Holzwirth, E.; Kornej, J.; Erbs, S.; Obradovic, D.; Bollmann, A.; Hindricks, G.; Thiele, H.; Büttner, P. Myeloperoxidase in atrial fibrillation: Association with progression, origin and influence of renin-angiotensin system antagonists. Clin. Res. Cardiol. 2020, 109, 324–330. [Google Scholar] [CrossRef] [PubMed]
- Kornej, J.; Zeynalova, S.; Büttner, P.; Burkhardt, R.; Bae, Y.J.; Willenberg, A.; Baber, R.; Thaler, A.; Hindricks, G.; Loeffler, M.; et al. Differentiation of atrial fibrillation progression phenotypes using troponin, T. Int. J. Cardiol. 2019, 297, 61–65. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Li, H.; Kou, W.; Tang, K.; Zhao, D.; Zhang, J.; Zhuang, J.; Zhao, Y.; Ji, S.; Peng, W.; et al. Increased plasma microfibrillar-associated protein 4 is associated with atrial fibrillation and more advanced left atrial remodelling. Arch. Med. Sci. 2019, 15, 632–640. [Google Scholar] [CrossRef]
- Lau, D.H.; Schotten, U.; Mahajan, R.; Antic, N.A.; Hatem, S.N.; Pathak, R.K.; Hendriks, J.M.L.; Kalman, J.M.; Sanders, P. Novel mechanisms in the pathogenesis of atrial fibrillation: Practical applications. Eur. Heart J. 2016, 37, 1573–1581. [Google Scholar] [CrossRef] [Green Version]
- Oba, K.; Maeda, M.; Maimaituxun, G.; Yamaguchi, S.; Arasaki, O.; Fukuda, D.; Yagi, S.; Hirata, Y.; Nishio, S.; Iwase, T.; et al. Effect of the epicardial adipose tissue volume on the prevalence of paroxysmal and persistent atrial fibrillation. Circ. J. 2018, 82, 1778–1787. [Google Scholar] [CrossRef] [Green Version]
- Gaeta, M.; Bandera, F.; Tassinari, F.; Capasso, L.; Cargnelutti, M.; Pelissero, G.; Malavazos, A.E.; Ricci, C. Is epicardial fat depot associated with atrial fibrillation? A systematic review and meta-analysis. EP Eur. 2017, 19, 747–752. [Google Scholar] [CrossRef]
- Chen, Q.; Chen, X.; Wang, J.; Zhong, J.; Zhang, H.; Wu, B.; Zheng, Z.; Xie, X.; Zhu, J.; Tang, X.; et al. Redistribution of adipose tissue is associated with left atrial remodeling and dysfunction in patients with atrial fibrillation. Front. Cardiovasc. Med. 2022, 9, 969513. [Google Scholar] [CrossRef]
- Zhao, L.; Ma, Z.; Guo, Z.; Zheng, M.; Li, K.; Yang, X. Analysis of long non-coding RNA and MRNA profiles in epicardial adipose tissue of patients with atrial fibrillation. Biomed. Pharmacother. 2020, 121, 109634. [Google Scholar] [CrossRef]
- Tsao, H.-M.; Hu, W.-C.; Tsai, P.-H.; Lee, C.-L.; Liu, F.-C.; Wang, H.-H.; Lo, L.-W.; Chang, S.-L.; Chao, T.-F.; Chen, S.-A. The abundance of epicardial adipose tissue surrounding left atrium is associated with the occurrence of stroke in patients with atrial fibrillation. Medicine 2016, 95, e3260. [Google Scholar] [CrossRef]
- Haemers, P.; Hamdi, H.; Guedj, K.; Suffee, N.; Farahmand, P.; Popovic, N.; Claus, P.; LePrince, P.; Nicoletti, A.; Jalife, J.; et al. Atrial fibrillation is associated with the fibrotic remodelling of adipose tissue in the subepicardium of human and sheep atria. Eur. Heart J. 2017, 38, 53–61. [Google Scholar] [CrossRef] [Green Version]
- Uno, G.; Omori, T.; Shimada, S.; Rader, F.; Siegel, R.J.; Shiota, T. Differences in mitral valve geometry between atrial and ventricular functional mitral regurgitation in patients with atrial fibrillation: A 3D transoesophageal echocardiography study. Eur. Heart J. Cardiovasc. Imaging 2021, 22, 1106–1116. [Google Scholar] [CrossRef]
- Kim, D.-H.; Heo, R.; Handschumacher, M.D.; Lee, S.; Choi, Y.-S.; Kim, K.-R.; Shin, Y.; Park, H.-K.; Bischoff, J.; Aikawa, E.; et al. Mitral valve adaptation to isolated annular dilation. JACC Cardiovasc. Imaging 2019, 12, 665–677. [Google Scholar] [CrossRef]
- Machino-Ohtsuka, T.; Seo, Y.; Ishizu, T.; Sato, K.; Sugano, A.; Yamamoto, M.; Hamada-Harimura, Y.; Aonuma, K. Novel mechanistic insights into atrial functional mitral regurgitation—3-dimensional echocardiographic study. Circ. J. 2016, 80, 2240–2248. [Google Scholar] [CrossRef] [Green Version]
- Guta, A.C.; Badano, L.P.; Tomaselli, M.; Mihalcea, D.; Bartos, D.; Parati, G.; Muraru, D. The pathophysiological link between right atrial remodeling and functional tricuspid regurgitation in patients with atrial fibrillation: A three-dimensional echocardiography study. J. Am. Soc. Echocardiogr. 2021, 34, 585–594.e1. [Google Scholar] [CrossRef]
- Tang, B. Atrial fibrillation electrical remodelling via ablation of the epicardial neural networks and suprathreshold stimulation of vagosympathetic nerve. Med. Sci. Monit. 2015, 21, 82–89. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.; Wang, X.; Li, Z.; Wang, W.; Zhang, Y.; Liu, J.; Hou, Y. Atrial heterogeneous autonomic neural remodeling in rabbits with experimental atrial fibrillation and the effect of intervention by rosuvastatin. Pacing Clin. Electrophysiol. 2016, 39, 598–606. [Google Scholar] [CrossRef]
- De Oliveira, Í.M.; Silva, E.L.d.; Martins, Y.d.O.; Rocha, H.A.L.; Scanavacca, M.I.; Gutierrez, P.S. Cardiac autonomic nervous system remodeling may play a role in atrial fibrillation: A study of the autonomic nervous system and myocardial receptors. Arq. Bras. Cardiol. 2021, 117, 999–1007. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, M.; Zhang, Y.; Xie, X.; Wang, W.; Li, Z.; Gao, M.; Wang, Z.; Hou, Y. Long-term effects of ganglionated plexi ablation on electrophysiological characteristics and neuron remodeling in target atrial tissues in a canine model. Circ. Arrhythm. Electrophysiol. 2015, 8, 1276–1283. [Google Scholar] [CrossRef]
- Moss, E.; Cardinal, R.; Yin, Y.; Pagé, P. Biatrial neuroablation attenuates atrial remodeling and vulnerability to atrial fibrillation in canine chronic rapid atrial pacing. Auton. Neurosci. 2015, 189, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.-J.; Yao, F.-J.; Lu, G.-H.; Xu, C.-G.; Xu, Z.; Tang, K.; Cheng, Y.-J.; Gao, X.-R.; Wu, S.-H. The role of the Rho/ROCK pathway in Ang II and TGF-Β1-induced atrial remodeling. PLoS ONE 2016, 11, e0161625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walters, T.E.; Kalman, J.M.; Patel, S.K.; Mearns, M.; Velkoska, E.; Burrell, L.M. Angiotensin converting enzyme 2 activity and human atrial fibrillation: Increased plasma angiotensin converting enzyme 2 activity is associated with atrial fibrillation and more advanced left atrial structural remodelling. Europace 2016, 19, 1280–1287. [Google Scholar] [CrossRef]
- Zhou, T.; Han, Z.; Gu, J.; Chen, S.; Fan, Y.; Zhang, H.; Yin, Y.; Zhang, J.; Wang, C. Angiotensin-converting enzyme-2 overexpression improves atrial electrical remodeling through TRPM7 signaling pathway. Oncotarget 2017, 8, 78726–78733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ihara, K.; Sasano, T. Role of inflammation in the pathogenesis of atrial fibrillation. Front. Physiol. 2022, 13, 862164. [Google Scholar] [CrossRef]
- Pauklin, P.; Zilmer, M.; Eha, J.; Tootsi, K.; Kals, M.; Kampus, P. Markers of inflammation, oxidative stress, and fibrosis in patients with atrial fibrillation. Oxid. Med. Cell. Longev. 2022, 2022, 4556671. [Google Scholar] [CrossRef]
- Smorodinova, N.; Bláha, M.; Melenovský, V.; Rozsívalová, K.; Přidal, J.; Ďurišová, M.; Pirk, J.; Kautzner, J.; Kučera, T. Analysis of immune cell populations in atrial myocardium of patients with atrial fibrillation or sinus rhythm. PLoS ONE 2017, 12, e0172691. [Google Scholar] [CrossRef] [Green Version]
- Yao, C.; Veleva, T.; Scott, L.; Cao, S.; Li, L.; Chen, G.; Jeyabal, P.; Pan, X.; Alsina, K.M.; Abu-Taha, I.; et al. Enhanced cardiomyocyte NLRP3 inflammasome signaling promotes atrial fibrillation. Circulation 2018, 138, 2227–2242. [Google Scholar] [CrossRef]
- Hiram, R.; Xiong, F.; Naud, P.; Xiao, J.; Sirois, M.; Tanguay, J.-F.; Tardif, J.-C.; Nattel, S. The inflammation-resolution promoting molecule resolvin-D1 prevents atrial proarrhythmic remodelling in experimental right heart disease. Cardiovasc. Res. 2021, 117, 1776–1789. [Google Scholar] [CrossRef]
- Kornej, J.; Büttner, P.; Hammer, E.; Engelmann, B.; Dinov, B.; Sommer, P.; Husser, D.; Hindricks, G.; Völker, U.; Bollmann, A. Circulating proteomic patterns in AF related left atrial remodeling indicate involvement of coagulation and complement cascade. PLoS ONE 2018, 13, e0198461. [Google Scholar] [CrossRef] [Green Version]
- Kondo, H.; Abe, I.; Fukui, A.; Saito, S.; Miyoshi, M.; Aoki, K.; Shinohara, T.; Teshima, Y.; Yufu, K.; Takahashi, N. Possible role of rivaroxaban in attenuating pressure-overload-induced atrial fibrosis and fibrillation. J. Cardiol. 2018, 71, 310–319. [Google Scholar] [CrossRef]
- Wałek, P.; Grabowska, U.; Cieśla, E.; Sielski, J.; Roskal-Wałek, J.; Wożakowska-Kapłon, B. Analysis of the correlation of galectin-3 concentration with the measurements of echocardiographic parameters assessing left atrial remodeling and function in patients with persistent atrial fibrillation. Biomolecules 2021, 11, 1108. [Google Scholar] [CrossRef]
- Takemoto, Y.; Ramirez, R.J.; Yokokawa, M.; Kaur, K.; Ponce-Balbuena, D.; Sinno, M.C.; Willis, B.C.; Ghanbari, H.; Ennis, S.R.; Guerrero-Serna, G.; et al. Galectin-3 regulates atrial fibrillation remodeling and predicts catheter ablation outcomes. JACC Basic Transl. Sci. 2016, 1, 143–154. [Google Scholar] [CrossRef] [Green Version]
- Tang, Z.; Zeng, L.; Lin, Y.; Han, Z.; Gu, J.; Wang, C.; Zhang, H. Circulating galectin-3 is associated with left atrial appendage remodelling and thrombus formation in patients with atrial fibrillation. Heart Lung Circ. 2019, 28, 923–931. [Google Scholar] [CrossRef]
- Ivanov, V.; Smereka, Y.; Rasputin, V.; Dmytriiev, K. Homocysteine and atrial fibrillation: Novel evidences and insights. Monaldi Arch. Chest Dis. 2022, in press. [Google Scholar] [CrossRef]
- Taufiq, F.; Li, P.; Miake, J.; Hisatome, I. Hyperuricemia as a risk factor for atrial fibrillation due to soluble and crystalized uric acid. Circ. Rep. 2019, 1, 469–473. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.-D.; Liu, J.; Zhang, Y.-C.; Wang, Y.; Wang, Y.; Ma, D. Correlation between the elevated uric acid levels and circulating renin-angiotensin-aldosterone system activation in patients with atrial fibrillation. Cardiovasc. Diagn. Ther. 2021, 11, 50–55. [Google Scholar] [CrossRef]
- Markus, M.R.P.; Meffert, P.J.; Baumeister, S.E.; Lieb, W.; Siewert, U.; Schipf, S.; Koch, M.; Kors, J.A.; Felix, S.B.; Dörr, M.; et al. Association between hepatic steatosis and serum liver enzyme levels with atrial fibrillation in the general population. Atherosclerosis 2016, 245, 123–131. [Google Scholar] [CrossRef]
- Wang, X.; Li, Z.; Zang, M.; Yao, T.; Mao, J.; Pu, J. Circulating primary bile acid is correlated with structural remodeling in atrial fibrillation. J. Interv. Card. Electrophysiol. 2020, 57, 371–377. [Google Scholar] [CrossRef]
- Zuo, K.; Fang, C.; Liu, Z.; Fu, Y.; Liu, Y.; Liu, L.; Wang, Y.; Yin, X.; Liu, X.; Li, J.; et al. Commensal microbe-derived SCFA alleviates atrial fibrillation via GPR43/NLRP3 signaling. Int. J. Biol. Sci. 2022, 18, 4219–4232. [Google Scholar] [CrossRef]
- Zuo, K.; Yin, X.; Li, K.; Zhang, J.; Wang, P.; Jiao, J.; Liu, Z.; Liu, X.; Liu, J.; Li, J.; et al. Different types of atrial fibrillation share patterns of gut microbiota dysbiosis. mSphere 2020, 5, e00071-20. [Google Scholar] [CrossRef] [PubMed]
- Lin, K.; Chen, K.; Li, S.; Cai, M.; Yuan, M.; Wang, Y.; Zhang, X.; Wei, M.; Yan, M.-L.; Ma, X.-X.; et al. Impaired left atrial performance resulting from age-related arial fibrillation is associated with increased fibrosis burden: Insights from a clinical study combining with an in vivo experiment. Front. Cardiovasc. Med. 2021, 7, 615065. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.M.; Lee, D.I.; Park, H.C.; Shim, J.; Choi, J.; Park, S.W.; Kim, Y. The extent of complex fractionated atrial electrograms in the left atrium reflects age-related electrical remodeling in patients with persistent atrial fibrillation. J. Arrhythm. 2019, 35, 805–812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Javed, S.; Gupta, D.; Lip, G.Y.H. Obesity and atrial fibrillation: Making inroads through fat. Eur. Heart J. Cardiovasc. Pharmacother. 2021, 7, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Chirinos, J.A.; Sardana, M.; Satija, V.; Gillebert, T.C.; de Buyzere, M.L.; Chahwala, J.; de Bacquer, D.; Segers, P.; Rietzschel, E.R. Effect of obesity on left atrial strain in persons aged 35–55 years (The Asklepios study). Am. J. Cardiol. 2019, 123, 854–861. [Google Scholar] [CrossRef] [PubMed]
- Mishima, R.S.; Ariyaratnam, J.P.; Pitman, B.M.; Malik, V.; Emami, M.; McNamee, O.; Stokes, M.B.; Lau, D.H.; Sanders, P.; Elliott, A.D. Cardiorespiratory fitness, obesity and left atrial function in patients with atrial fibrillation. IJC Heart Vasc. 2022, 42, 101083. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-C.; Lin, Y.-H. The pathogenic role of very low density lipoprotein on atrial remodeling in the metabolic syndrome. Int. J. Mol. Sci. 2020, 21, 891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rafaqat, S. Biomarkers of metabolic syndrome: Role in pathogenesis and pathophysiology of atrial fibrillation. J. Atr. Fibrillation 2021, 14, 20200495. [Google Scholar] [CrossRef]
- Nalliah, C.J.; Wong, G.R.; Lee, G.; Voskoboinik, A.; Kee, K.; Goldin, J.; Watts, T.; Linz, D.; Wirth, D.; Parameswaran, R.; et al. Sleep apnoea has a dose-dependent effect on atrial remodelling in paroxysmal but not persistent atrial fibrillation: A high-density mapping study. EP Eur. 2021, 23, 691–700. [Google Scholar] [CrossRef]
- Vizzardi, E.; Sciatti, E.; Bonadei, I.; D’Aloia, A.; Curnis, A.; Metra, M. Obstructive sleep apnoea—Hypopnoea and arrhythmias. J. Cardiovasc. Med. 2017, 18, 490–500. [Google Scholar] [CrossRef]
- Wong, G.R.; Nalliah, C.J.; Lee, G.; Voskoboinik, A.; Chieng, D.; Prabhu, S.; Parameswaran, R.; Sugumar, H.; Al-Kaisey, A.; McLellan, A.; et al. Sex-related differences in atrial remodeling in patients with atrial fibrillation: Relationship to ablation outcomes. Circ. Arrhythm. Electrophysiol. 2022, 15, e009925. [Google Scholar] [CrossRef]
- Akoum, N.; Mahnkopf, C.; Kholmovski, E.G.; Brachmann, J.; Marrouche, N.F. Age and sex differences in atrial fibrosis among patients with atrial fibrillation. EP Eur. 2018, 20, 1086–1092. [Google Scholar] [CrossRef]
- Paliwal, N.; Ali, R.L.; Salvador, M.; O’Hara, R.; Yu, R.; Daimee, U.A.; Akhtar, T.; Pandey, P.; Spragg, D.D.; Calkins, H.; et al. Presence of left atrial fibrosis may contribute to aberrant hemodynamics and increased risk of stroke in atrial fibrillation patients. Front. Physiol. 2021, 12, 657452. [Google Scholar] [CrossRef]
- Li, Z.; Wang, Z.; Yin, Z.; Zhang, Y.; Xue, X.; Han, J.; Zhu, Y.; Zhang, J.; Emmert, M.Y.; Wang, H. Gender differences in fibrosis remodeling in patients with long-standing persistent atrial fibrillation. Oncotarget 2017, 8, 53714–53729. [Google Scholar] [CrossRef] [Green Version]
- Polejaeva, I.A.; Ranjan, R.; Davies, C.J.; Regouski, M.; Hall, J.; Olsen, A.L.; Meng, Q.; Rutigliano, H.M.; Dosdall, D.J.; Angel, N.A.; et al. Increased susceptibility to atrial fibrillation secondary to atrial fibrosis in transgenic goats expressing transforming growth factor-Β1. J. Cardiovasc. Electrophysiol. 2016, 27, 1220–1229. [Google Scholar] [CrossRef] [Green Version]
- Ammar-Busch, S.; Buiatti, A.; Tatzber, A.; Reents, T.; Bourier, F.; Semmler, V.; Telishevska, M.; Hessling, G.; Deisenhofer, I. Predictors of low voltage areas in persistent atrial fibrillation: Is it really a matter of time? J. Interv. Card. Electrophysiol. 2020, 57, 345–352. [Google Scholar] [CrossRef]
- Ríos-Muñoz, G.R.; Soto, N.; Ávila, P.; Carta, A.; Atienza, F.; Datino, T.; González-Torrecilla, E.; Fernández-Avilés, F.; Arenal, Á. Structural remodeling and rotational activity in persistent/long-lasting atrial fibrillation: Gender-effect differences and impact on post-ablation outcome. Front. Cardiovasc. Med. 2022, 9, 819429. [Google Scholar] [CrossRef]
- Lau, C.-P.; Tse, H.-F.; Siu, C.-W.; Gbadebo, D. Atrial electrical and structural remodeling: Implications for racial differences in atrial fibrillation. J. Cardiovasc. Electrophysiol. 2012, 23, s36–s40. [Google Scholar] [CrossRef]
- Dmour, B.-A.; Miftode, R.-S.; Iliescu Halitchi, D.; Anton-Paduraru, D.T.; Iliescu Halitchi, C.-O.; Miftode, I.-L.; Mitu, O.; Costache, A.-D.; Stafie, C.-S.; Costache, I.I. Latest insights into mechanisms behind atrial cardiomyopathy: It is not always about ventricular function. Diagnostics 2021, 11, 449. [Google Scholar] [CrossRef]
- Wan, D.; Andrade, J.; Laksman, Z. Thromboembolic risk stratification in atrial fibrillation—Beyond clinical risk scores. Rev. Cardiovasc. Med. 2021, 22, 353. [Google Scholar] [CrossRef]
- Maille, B.; Das, M.; Hussein, A.; Shaw, M.; Chaturvedi, V.; Williams, E.; Morgan, M.; Ronayne, C.; Snowdon, R.L.; Gupta, D. Reverse electrical and structural remodeling of the left atrium occurs early after pulmonary vein isolation for persistent atrial fibrillation. J. Interv. Card. Electrophysiol. 2020, 58, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.-H.; Li, H.-K.; Couri, D.M.; Araoz, P.A.; Lee, Y.-H.; Ma, C.-S.; Packer, D.L.; Cha, Y.-M. Reversal of pulmonary vein remodeling after catheter ablation of atrial fibrillation. J. Geriatr. Cardiol. 2016, 13, 163–168. [Google Scholar] [CrossRef] [PubMed]
- Hori, Y.; Nakahara, S.; Fukuda, R.; Sato, H.; Ukaji, T.; Koshikawa, Y.; Nishiyama, N.; Ishikawa, T.; Kobayashi, S.; Taguchi, I. Atrial reverse remodeling represented by the atrial conduction time in persistent atrial fibrillation patients after catheter ablation: Its impact on predicting late atrial fibrillation recurrence. J. Cardiol. 2020, 75, 521–528. [Google Scholar] [CrossRef] [PubMed]
- Kriatselis, C.; Unruh, T.; Kaufmann, J.; Gerds-Li, J.-H.; Kelle, S.; Gebker, R.; Jahnke, C.; Paetsch, I.; Pieske, B. Long-term left atrial remodeling after ablation of persistent atrial fibrillation: 7-year follow-up by cardiovascular magnetic resonance imaging. J. Interv. Card. Electrophysiol. 2020, 58, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Hwang, J.; Park, H.-S.; Han, S.; Jun, S.-W.; Kang, N.-Y.; Jeon, J.-H.; Choi, S.-W.; Lee, C.H.; Kim, I.-C.; Cho, Y.-K.; et al. The impact of catheter ablation of atrial fibrillation on the left atrial volume and function: Study using three-dimensional echocardiography. J. Interv. Card. Electrophysiol. 2020, 57, 87–95. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Liu, Q.; Yang, Y.; Zhang, C.; Yin, H.; Wu, J.; Yao, L.; Jin, L.; Yang, J.; Feng, L.; et al. Effect of radiofrequency catheter ablation on left atrial structure and function in patients with different types of atrial fibrillation. Sci. Rep. 2022, 12, 9511. [Google Scholar] [CrossRef]
- Liżewska-Springer, A.; Dąbrowska-Kugacka, A.; Lewicka, E.; Królak, T.; Drelich, Ł.; Kozłowski, D.; Raczak, G. Echocardiographic assessment in patients with atrial fibrillation (AF) and normal systolic left ventricular function before and after catheter ablation: If AF begets AF, does pulmonary vein isolation terminate the vicious circle? Cardiol. J. 2020, 27, 126–135. [Google Scholar] [CrossRef] [Green Version]
- Nagai, T.; Arakawa, J.; Hamabe, A.; Tabata, H. Improvement of left ventricular function after successful radiofrequency catheter ablation in persistent atrial fibrillation with preserved left ventricular ejection fraction: A comprehensive echocardiographic assessment using two-dimensional speckle tracking analysis. J. Echocardiogr. 2019, 17, 95–103. [Google Scholar] [CrossRef]
- Nishino, S.; Watanabe, N.; Ashikaga, K.; Morihisa, K.; Kuriyama, N.; Asada, Y.; Shibata, Y. Reverse remodeling of the mitral valve complex after radiofrequency catheter ablation for atrial fibrillation. Circ. Cardiovasc. Imaging 2019, 12, e009317. [Google Scholar] [CrossRef]
- Masuda, M.; Sekiya, K.; Asai, M.; Iida, O.; Okamoto, S.; Ishihara, T.; Nanto, K.; Kanda, T.; Tsujimura, T.; Matsuda, Y.; et al. Influence of catheter ablation for atrial fibrillation on atrial and ventricular functional mitral regurgitation. ESC Heart Fail. 2022, 9, 1901–1913. [Google Scholar] [CrossRef]
- Chen, J.; Wang, Y.; Lv, M.; Yang, Z.; Zhu, S.; Wei, L.; Hong, T.; Ding, W.; Lin, Y.; Wang, C. Mitral Valve Repair and surgical ablation for atrial functional mitral regurgitation. Ann. Transl. Med. 2020, 8, 1420. [Google Scholar] [CrossRef]
- Kawakami, H.; Inoue, K.; Nagai, T.; Fujii, A.; Sasaki, Y.; Shikano, Y.; Sakuoka, N.; Miyazaki, M.; Takasuka, Y.; Ikeda, S.; et al. Persistence of left atrial abnormalities despite left atrial volume normalization after successful ablation of atrial fibrillation. J. Arrhythm. 2021, 37, 1318–1329. [Google Scholar] [CrossRef]
- Miwa, Y.; Mohri, T.; Katsume, Y.; Tashiro, M.; Momose, Y.; Nonoguchi, N.; Hoshida, K.; Togashi, I.; Hagiwara, Y.; Maeda, A.; et al. Left atrial reverse remodeling following the modified box isolation with centerline in patients with persistent atrial fibrillation. Int. Heart J. 2021, 62, 21–108. [Google Scholar] [CrossRef]
- Sasaki, N.; Okumura, Y.; Watanabe, I.; Mano, H.; Nagashima, K.; Sonoda, K.; Kogawa, R.; Ohkubo, K.; Nakai, T.; Hirayama, A. Increased levels of inflammatory and extracellular matrix turnover biomarkers persist despite reverse atrial structural remodeling during the first year after atrial fibrillation ablation. J. Interv. Card. Electrophysiol. 2014, 39, 241–249. [Google Scholar] [CrossRef]
- Yang, L.; Zhang, M.; Hao, Z.; Wang, N.; Zhang, M. Sacubitril/valsartan attenuates atrial structural remodelling in atrial fibrillation patients. ESC Heart Fail. 2022, 9, 2428–2434. [Google Scholar] [CrossRef]
- Takemoto, Y.; Ramirez, R.J.; Kaur, K.; Salvador-Montañés, O.; Ponce-Balbuena, D.; Ramos-Mondragón, R.; Ennis, S.R.; Guerrero-Serna, G.; Berenfeld, O.; Jalife, J. Eplerenone reduces atrial fibrillation burden without preventing atrial electrical remodeling. J. Am. Coll. Cardiol. 2017, 70, 2893–2905. [Google Scholar] [CrossRef]
- Tong, M.; Wang, J.; Ji, Y.; Chen, X.; Wang, J.; Wang, S.; Ruan, L.; Cui, H.; Zhou, Y.; Zhang, Q.; et al. Effect of eicosapentaenoic acid and pitavastatin on electrophysiology and anticoagulant gene expression in mice with rapid atrial pacing. Exp. Ther. Med. 2017, 14, 2310–2316. [Google Scholar] [CrossRef] [Green Version]
- Heijman, J.; Sutanto, H.; Crijns, H.J.G.M.; Nattel, S.; Trayanova, N.A. Computational models of atrial fibrillation: Achievements, challenges, and perspectives for improving clinical care. Cardiovasc. Res. 2021, 117, 1682–1699. [Google Scholar] [CrossRef]
- Lee, Y.-S.; Hwang, M.; Song, J.-S.; Li, C.; Joung, B.; Sobie, E.A.; Pak, H.-N. The contribution of ionic currents to rate-dependent action potential duration and pattern of reentry in a mathematical model of human atrial fibrillation. PLoS ONE 2016, 11, e0150779. [Google Scholar] [CrossRef] [Green Version]
- Liberos, A.; Bueno-Orovio, A.; Rodrigo, M.; Ravens, U.; Hernandez-Romero, I.; Fernandez-Aviles, F.; Guillem, M.S.; Rodriguez, B.; Climent, A.M. Balance between sodium and calcium currents underlying chronic atrial fibrillation termination: An in silico intersubject variability study. Heart Rhythm 2016, 13, 2358–2365. [Google Scholar] [CrossRef] [Green Version]
- Palacio, L.C.; Ugarte, J.P.; Saiz, J.; Tobón, C. The effects of fibrotic cell type and its density on atrial fibrillation dynamics: An in silico study. Cells 2021, 10, 2769. [Google Scholar] [CrossRef] [PubMed]
- Jost, N.; Christ, T.; Magyar, J. New strategies for the treatment of atrial fibrillation. Pharmaceuticals 2021, 14, 926. [Google Scholar] [CrossRef] [PubMed]
- Calvo, D.; Filgueiras-Rama, D.; Jalife, J. Mechanisms and drug development in atrial fibrillation. Pharmacol. Rev. 2018, 70, 505–525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fenner, M.F.; Gatta, G.; Sattler, S.; Kuiper, M.; Hesselkilde, E.M.; Adler, D.M.T.; Smerup, M.; Schotten, U.; Sørensen, U.; Diness, J.G.; et al. Inhibition of small-conductance calcium-activated potassium current (IK,Ca) leads to differential atrial electrophysiological effects in a horse model of persistent atrial fibrillation. Front. Physiol. 2021, 12, 614483. [Google Scholar] [CrossRef] [PubMed]
- Yakabe, D.; Fukuyama, Y.; Araki, M.; Nakamura, T. Responsiveness to bepridil predicts atrial substrate in patients with persistent atrial fibrillation. J. Arrhythm. 2021, 37, 79–87. [Google Scholar] [CrossRef]
- Nakatani, Y.; Sakamoto, T.; Nishida, K.; Kataoka, N.; Yamaguchi, Y.; Sakabe, M.; Fujiki, A.; Mizumaki, K.; Inoue, H. Bepridil enhances aprindine-induced prolongation of atrial effective refractory period in a canine atrial rapid pacing model. J. Cardiol. 2015, 66, 445–450. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Z.; Niu, X.; Dong, Z.; Qi, W.; Liu, E.; Liu, T.; Li, L.; Liang, Y.; Li, G. Upstream therapeutic strategies of valsartan and fluvastatin on hypertensive patients with non-permanent atrial fibrillation. Cardiovasc. Ther. 2018, 36, e12478. [Google Scholar] [CrossRef] [Green Version]
- Shao, Q.; Meng, L.; Lee, S.; Tse, G.; Gong, M.; Zhang, Z.; Zhao, J.; Zhao, Y.; Li, G.; Liu, T. Empagliflozin, a sodium glucose co-transporter-2 inhibitor, alleviates atrial remodeling and improves mitochondrial function in high-fat diet/streptozotocin-induced diabetic rats. Cardiovasc. Diabetol. 2019, 18, 165. [Google Scholar] [CrossRef] [Green Version]
- Nishinarita, R.; Niwano, S.; Niwano, H.; Nakamura, H.; Saito, D.; Sato, T.; Matsuura, G.; Arakawa, Y.; Kobayashi, S.; Shirakawa, Y.; et al. Canagliflozin suppresses atrial remodeling in a canine atrial fibrillation model. J. Am. Heart Assoc. 2021, 10, e017483. [Google Scholar] [CrossRef]
- Kiedrowicz, R.M.; Wielusiński, M.; Wojtarowicz, A.; Kaźmierczak, J. Atrial fibrillation risk scores to evaluate left atrial substrate based on voltage analysis in long-standing persistent type of arrhythmia. Kardiol. Pol. 2021, 79, 525–530. [Google Scholar] [CrossRef]
- Marchandise, S.; Garnir, Q.; Scavée, C.; Varnavas, V.; le Polain de Waroux, J.-B.; Wauters, A.; Beauloye, C.; Roelants, V.; Gerber, B.L. Prediction of left atrial fibrosis and success of catheter ablation by speckle tracking echocardiography in patients imaged in persistent atrial fibrillation. Front. Cardiovasc. Med. 2022, 9, 856796. [Google Scholar] [CrossRef]
- Sardu, C.; Santulli, G.; Guerra, G.; Trotta, M.C.; Santamaria, M.; Sacra, C.; Testa, N.; Ducceschi, V.; Gatta, G.; d’Amico, M.; et al. Modulation of SERCA in patients with persistent atrial fibrillation treated by epicardial thoracoscopic ablation: The CAMAF study. J. Clin. Med. 2020, 9, 544. [Google Scholar] [CrossRef] [Green Version]
- Roney, C.H.; Beach, M.L.; Mehta, A.M.; Sim, I.; Corrado, C.; Bendikas, R.; Solis-Lemus, J.A.; Razeghi, O.; Whitaker, J.; O’Neill, L.; et al. In silico comparison of left atrial ablation techniques that target the anatomical, structural, and electrical substrates of atrial fibrillation. Front. Physiol. 2020, 11, 1145. [Google Scholar] [CrossRef]
- Kim, I.-S.; Lim, B.; Shim, J.; Hwang, M.; Yu, H.T.; Kim, T.-H.; Uhm, J.-S.; Kim, S.-H.; Joung, B.; On, Y.K.; et al. Clinical usefulness of computational modeling-guided persistent atrial fibrillation ablation: Updated outcome of multicenter randomized study. Front. Physiol. 2019, 10, 1512. [Google Scholar] [CrossRef]
- Kanemaru, Y.; Arima, Y.; Kaikita, K.; Kiyama, T.; Kaneko, S.; Ito, M.; Yamabe, H.; Motozato, K.; Yamanaga, K.; Fujisue, K.; et al. Elongation of the high right atrium to coronary sinus conduction time predicts the recurrence of atrial fibrillation after radiofrequency catheter ablation. Int. J. Cardiol. 2020, 300, 147–153. [Google Scholar] [CrossRef] [Green Version]
- Higuchi, S.; Ejima, K.; Shoda, M.; Yamamoto, E.; Iwanami, Y.; Yagishita, D.; Hagiwara, N. Impact of a prolonged interatrial conduction time for predicting the recurrence of atrial fibrillation after circumferential pulmonary vein isolation of persistent atrial fibrillation. Heart Vessel. 2019, 34, 616–624. [Google Scholar] [CrossRef]
- Mathias, A.; Moss, A.J.; McNitt, S.; Zareba, W.; Goldenberg, I.; Solomon, S.D.; Kutyifa, V. Clinical implications of complete left-sided reverse remodeling with cardiac resynchronization therapy. J. Am. Coll. Cardiol. 2016, 68, 1268–1276. [Google Scholar] [CrossRef]
- Alexander, B.; Sadiq, F.; Azimi, K.; Glover, B.; Antiperovitch, P.; Hopman, W.M.; Jaff, Z.; Baranchuk, A. Reverse atrial electrical remodeling induced by cardiac resynchronization therapy. J. Electrocardiol. 2017, 50, 610–614. [Google Scholar] [CrossRef]
- Kugler, L.; Markendorf, S.; Bachmann, M.; Eriksson, U. Cardiac resynchronization therapy in the presence of total atrioventricular block reduces long-lasting atrial fibrillation episodes. J. Arrhythm. 2022, 38, 723–729. [Google Scholar] [CrossRef]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. BMJ 2021, 372, n71. [Google Scholar] [CrossRef]
Figure 1.
Classification of atrial fibrillation based on its temporal pattern. Red arrows indicate deterioration, green arrows indicate improvement. The width of the arrows represents the most common clinical course. After a first episode, AF commonly becomes paroxysmal. Remission or primary persistent AF may occasionally occur. Paroxysmal AF may remain paroxysmal, go into remission, or progress into persistent AF. Persistent AF, if not treated effectively, becomes long-standing persistent AF. Without active treatment, persistent or long-standing persistent AF almost never improves to the paroxysmal form or remission.
Figure 1.
Classification of atrial fibrillation based on its temporal pattern. Red arrows indicate deterioration, green arrows indicate improvement. The width of the arrows represents the most common clinical course. After a first episode, AF commonly becomes paroxysmal. Remission or primary persistent AF may occasionally occur. Paroxysmal AF may remain paroxysmal, go into remission, or progress into persistent AF. Persistent AF, if not treated effectively, becomes long-standing persistent AF. Without active treatment, persistent or long-standing persistent AF almost never improves to the paroxysmal form or remission.

Figure 2.
Literature search process.

Figure 3.
Species analyzed in the cited literature. One study may have investigated more than one species. The majority (78%) of the studies investigated human experimental or clinical processes. Several studies (4%) included mathematical models and simulations. Canine and mouse models were the most studied animal models of persistent AF in the past 10 years (5% each).
Figure 3.
Species analyzed in the cited literature. One study may have investigated more than one species. The majority (78%) of the studies investigated human experimental or clinical processes. Several studies (4%) included mathematical models and simulations. Canine and mouse models were the most studied animal models of persistent AF in the past 10 years (5% each).

Figure 4.
Pathophysiological processes analyzed in the reviewed studies. One study may have investigated more than one process. The majority of the studies investigated organ (44%) and cellular level (33%) processes. Genetic level analysis was pursued in 15% of the studies, while general characteristics of patient populations with persistent AF were investigated in 8% of the studies.
Figure 4.
Pathophysiological processes analyzed in the reviewed studies. One study may have investigated more than one process. The majority of the studies investigated organ (44%) and cellular level (33%) processes. Genetic level analysis was pursued in 15% of the studies, while general characteristics of patient populations with persistent AF were investigated in 8% of the studies.

Figure 5.
Examples of the clinical course of AF in patients with implanted dual chamber cardiac pacemakers or defibrillators. These devices provide the capability of continuous heart rhythm monitoring after implant. Pacing provides treatment for symptomatic bradycardia and may also enable the use of medical or non-pharmacological treatment for AF if a low heart rate was limiting their use prior to the device implant. One-year follow-up is marked with the green line. Daily AF burden varies from 0 to 24 h. (a) Remission of paroxysmal atrial fibrillation a few weeks after device implant. (b) The pacemaker was implanted in a patient with persistent atrial fibrillation. Cardioversion was performed several weeks after the implant, and the AF remained in remission thereafter. (c) Paroxysmal atrial fibrillation with a stable burden after implant. (d) Progression of paroxysmal AF into persistent AF.
Figure 5.
Examples of the clinical course of AF in patients with implanted dual chamber cardiac pacemakers or defibrillators. These devices provide the capability of continuous heart rhythm monitoring after implant. Pacing provides treatment for symptomatic bradycardia and may also enable the use of medical or non-pharmacological treatment for AF if a low heart rate was limiting their use prior to the device implant. One-year follow-up is marked with the green line. Daily AF burden varies from 0 to 24 h. (a) Remission of paroxysmal atrial fibrillation a few weeks after device implant. (b) The pacemaker was implanted in a patient with persistent atrial fibrillation. Cardioversion was performed several weeks after the implant, and the AF remained in remission thereafter. (c) Paroxysmal atrial fibrillation with a stable burden after implant. (d) Progression of paroxysmal AF into persistent AF.

Figure 6.
Treatment pathways to prevent the progression of atrial fibrillation.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Roka, A.; Burright, I. Remodeling in Persistent Atrial Fibrillation: Pathophysiology and Therapeutic Targets—A Systematic Review. Physiologia 2023, 3, 43-72. https://doi.org/10.3390/physiologia3010004
AMA Style
Roka A, Burright I. Remodeling in Persistent Atrial Fibrillation: Pathophysiology and Therapeutic Targets—A Systematic Review. Physiologia. 2023; 3(1):43-72. https://doi.org/10.3390/physiologia3010004
Chicago/Turabian StyleRoka, Attila, and Isaac Burright. 2023. "Remodeling in Persistent Atrial Fibrillation: Pathophysiology and Therapeutic Targets—A Systematic Review" Physiologia 3, no. 1: 43-72. https://doi.org/10.3390/physiologia3010004