

Linking Phyllosphere and Rhizosphere Microbiome to the Plant–Insect Interplay: The New Dimension of Tripartite Interaction


Abstract
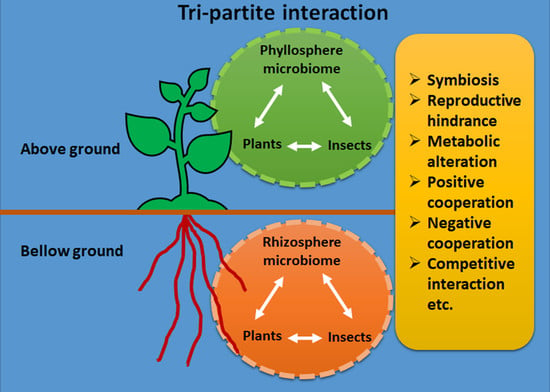
:
1. Introduction
2. Major Insect Pests to the Crop Plants
3. Molecular Mechanism of Plant–Insect Interplay
4. Plant-Associated Microbiome
4.1. Microbiome of Phyllosphere

{kind=link}
{kind=link}
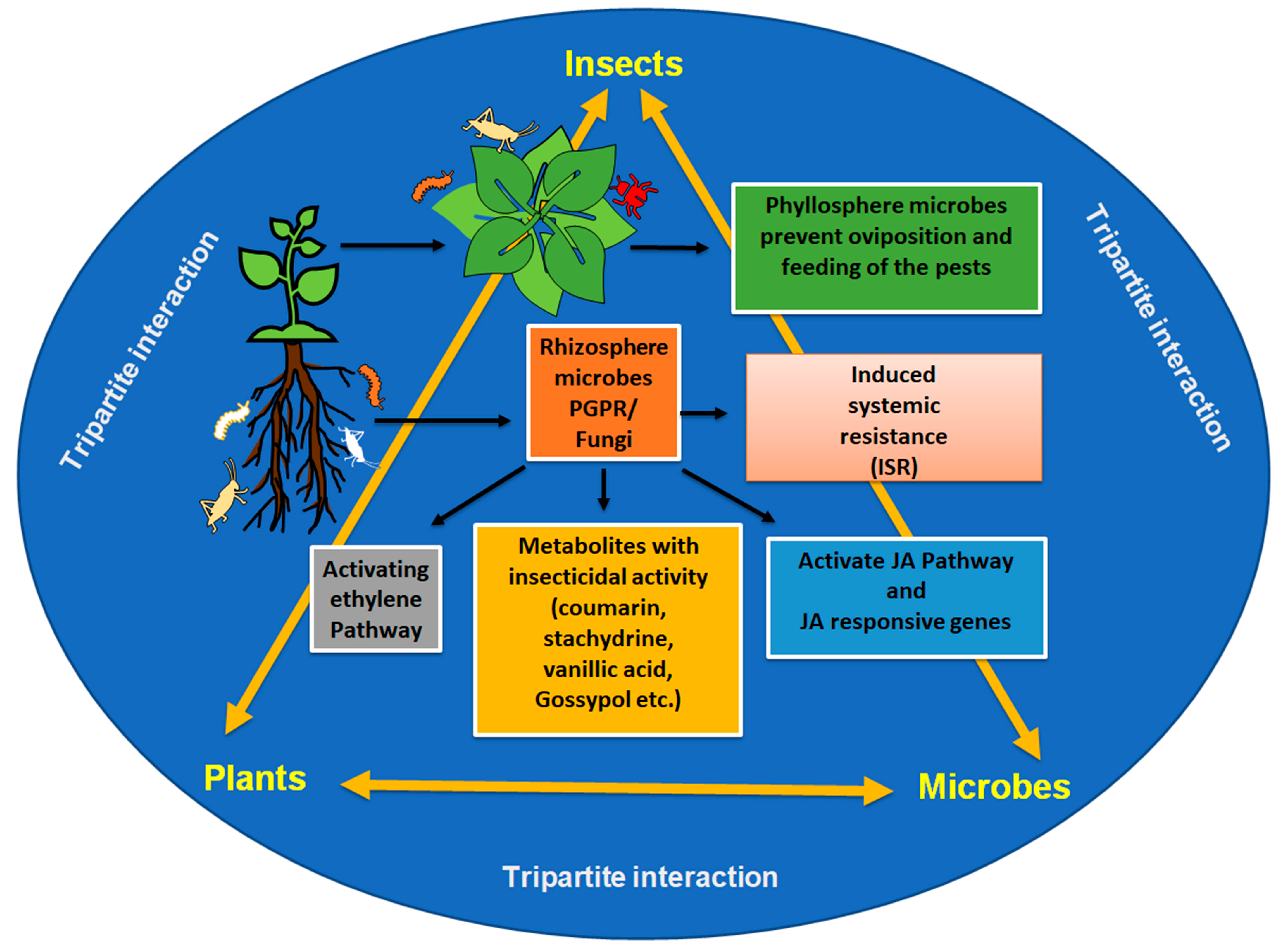{kind=link}
| Phyllosphere Microbes | Examples | Reference |
|---|---|---|
| Bacteria (considered to be the most abundant microbes in phyllosphere) | Proteobacteria (Alphaproteobacteria, Betaproteobacteria, and Gammaproteobacteria), Actinobacteria, Firmicutes, and Bacteroidetes are common in phyllosphere | Thapa et al. [55] |
| Bacillus and Pseudomonas may be prevalent genera | Thapa et al. [55] | |
| Methylotrophic bacteria are prevalent in phyllosphere | Sivakumar et al. [73] | |
| Methane-producing bacteria are also present | Iguchi et al. [61] | |
| Bacteria involved in nitrogen cycle may be present in phyllosphere including diazotrophic bacteria | Papen et al. [65], Furnkranz et al. [66] | |
| Bacterial pathogen may be present like Pseudomonas syringae | Sivakumar et al. [73] | |
| Biosurfactant producing bacteria may be present | Sivakumar et al. [73] | |
| Actinomycetes is also present in phyllosphere | Sivakumar et al. [73] | |
| Fungi | Epiphytic and endophytic fungi may be involved in decomposition of litter, recycling of carbon and nitrogen. Endophytic fungi may be involved in resistance against biotic and abiotic stress | Sivakumar et al. [73] |
| Yeast like fungi and filamentous fungi are present | Sivakumar et al. [73] | |
| Genus of Aspergillus, Penicillium, Alternaria, Trichoderma, Fusarium are observed | Sivakumar et al. [73] |
4.2. Microbiome of Rhizosphere
4.3. The Effect of Phyllosphere and Rhizosphere Microbiome on Plant–Insect Interaction
4.3.1. Phyllosphere Interaction
4.3.2. Effect of Plant Growth Promoting Rhizobacteria (PGPR)/Rhizosphere Bacteria on Plant–Insect Interaction
5. Plant–Microbe Interaction Shaping the Ecological Dynamics
6. Conclusions and Future Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Saikkonen, K.; Neuvonen, S. European pine sawfly and microbial interactions mediated by the host plant. In Sawfly Life History Adaptations to Woody Plants; Wagner, M., Raffa, K., Eds.; Academic Press, Inc.: New York, NY, USA, 1993; pp. 431–450. [Google Scholar]
- Sugio, A.; Dubreuil, G.; Giron, D.; Simon, J.C. Plant–insect interactions under bacterial influence: Ecological implications and underlying mechanisms. J. Exp. Bot. 2015, 66, 467–478. [Google Scholar] [CrossRef] [PubMed]
- Wielkopolan, B.; Obrępalska-Stęplowska, A. Three-way interaction among plants, bacteria, and coleopteran insects. Planta 2016, 244, 313–332. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.M.; Coupland, G.; Dolan, L.; Harberd, N.; Jones, J.; Martin, C.; Sablowski, R.; Amey, A. Plant Biology; Garland Science, Taylor & Francis Group: Boca Raton, FL, USA, 2010. [Google Scholar]
- Lappalainen, J.; Helander, M.L.; Palokangas, P. The performance of the autumnal moth is lower on trees infected by birch rust. Myco. Res. 1995, 99, 994–996. [Google Scholar] [CrossRef]
- Franco, F.P.; Túler, A.C.; Gallan, D.Z.; Gonçalves, F.G. Fungal phytopathogen modulates plant and insect responses to promote its dissemination. ISME J. 2021, 15, 3522–3533. [Google Scholar] [CrossRef] [PubMed]
- Sturz, A.V.; Christie, B.R.; Matheson, B.G.; Arsenault, W.J.; Buchanan, N.A. Endophytic bacterial communities in the periderm of potato tubers and their potential to improve resistance to soilborne plant pathogens. Plant Pathol. 1999, 48, 360–369. [Google Scholar] [CrossRef]
- Stout, M.J.; Thaler, J.S.; Thomma, B.P.H.J. Plant-mediated interactions between pathogenic microorganisms and herbivorous arthropods. Annu. Rev. Entomol. 2006, 51, 663–689. [Google Scholar] [CrossRef]
- Humphrey, P.T.; Nguyen, T.T.; Villalobos, M.M.; Whiteman, N.K. Diversity and abundance of phyllosphere bacteria are linked to insect herbivory. Mol. Ecol. 2014, 23, 1497–1515. [Google Scholar] [CrossRef]
- Bulgarelli, D.; Schlaeppi, K.; Spaepen, S.; van Themaat, E.V.L.; Schulze-Lefert, P. Structure and functions of the bacterial microbiota of plants. Annu. Rev. Plant. Biol. 2013, 64, 807–838. [Google Scholar] [CrossRef]
- Dhaliwal, G.S.; Jindal, V.; Dhawan, A.K. Insect pest problems and crop losses: Changing trends. Indian J. Ecol. 2010, 37, 1–7. [Google Scholar]
- IPPC Secretariat. Scientific Review of the Impact of Climate Change on Plant Pests—A Global Challenge to Prevent and Mitigate Plant Pest Risks in Agriculture, Forestry and Ecosystems; FAO on behalf of the IPPC Secretariat: Rome, Italy, 2021. [Google Scholar]
- Bishop, C.D.; Erezyilmaz, D.F.; Flatt, T.; Georgiou, C.D.; Hadfield, M.G.; Heyland, A.; Hodin, J.; Jacobs, M.W.; Maslakova, S.A.; Pires, A.; et al. What is metamorphosis? Integr. Comp. Biol. 2006, 46, 655–661. [Google Scholar] [CrossRef]
- Truman, J.W. The evolution of insect metamorphosis. Curr. Biol. 2019, 29, R1252–R1268. [Google Scholar] [CrossRef]
- App, B.A.; Manglitz, G.R. Insects and related pests. Alfalfa Sci. Technol. 1972, 15, 527–554. [Google Scholar]
- Reitz, S.R.; Gao, Y.; Kirk, W.D.; Hoddle, M.S.; Leiss, K.A.; Funderburk, J.E. Invasion biology, ecology, and management of western flower thrips. Annu. Rev. Entomol. 2020, 65, 17–37. [Google Scholar] [CrossRef]
- Rotenberg, D.; Baumann, A.A.; Ben-Mahmoud, S.; Christiaens, O.; Dermauw, W.; Ioannidis, P.; Jacobs, C.G.; Vargas Jentzsch, I.M.; Oliver, J.E.; Poelchau, M.F.; et al. Genome-enabled insights into the biology of thrips as crop pests. BMC Biol. 2020, 18, 169. [Google Scholar] [CrossRef]
- Gullan, P.J.; Kosztarab, M. Adaptations in scale insects. Annu. Rev. Entomol. 1997, 42, 23–50. [Google Scholar] [CrossRef]
- Mansour, R.; Grissa-Lebdi, K.; Suma, P.; Mazzeo, G.; Russo, A. Key scale insects (Hemiptera: Coccoidea) of high economic importance in a Mediterranean area: Host plants, bio-ecological characteristics, natural enemies and pest management strategies—A review. Plant Prot. Sci. 2017, 53, 1–14. [Google Scholar] [CrossRef]
- Joshi, M.D.; Butani, P.G.; Patel, V.N.; Jeyakumar, P. Cotton mealy bug, Phenacoccus solenopsis Tinsley-a review. Agric. Rev. 2010, 31, 113–119. [Google Scholar]
- Mokhtar, A.H. Biochemical and anatomical characteristics of some mango tree cultivars infested by two mealybugs, planococcus citri (risso) and icerya seychellarum (westood) in Egypt. Egypt. J. Agric. Res. 2022, 100, 570–580. [Google Scholar] [CrossRef]
- Prabhakar, M.; Prasad, Y.G.; Thirupathi, M.; Sreedevi, G.; Dharajothi, B.; Venkateswarlu, B. Use of ground based hyperspectral remote sensing for detection of stress in cotton caused by leafhopper (Hemiptera: Cicadellidae). Comput. Electron. Agric. 2011, 79, 189–198. [Google Scholar] [CrossRef]
- Shorthouse, J.D. INSECTS AND OTHER ANIMALS|Overview of Insects. Encyclopedia of Rose Science; Elsevier: Amsterdam, The Netherlands, 2005; pp. 415–425. [Google Scholar]
- Zhao, X.; Chen, S.; Wang, S.; Shan, W.; Wang, X.; Lin, Y.; Su, F.; Yang, Z.; Yu, X. Defensive responses of tea plants (Camellia sinensis) against tea green leafhopper attack: A multi-omics study. Front. Plant Sci. 2020, 10, 1705. [Google Scholar] [CrossRef]
- Zhang, J.; Guan, W.; Huang, C.; Hu, Y.; Chen, Y.; Guo, J.; Zhou, C.; Chen, R.; Du, B.; Zhu, L.; et al. Combining next-generation sequencing and single-molecule sequencing to explore brown plant hopper responses to contrasting genotypes of japonica rice. BMC Genom. 2019, 20, 682. [Google Scholar] [CrossRef] [PubMed]
- Thein, H.W.; Yamagata, Y.; Van Mai, T.; Yasui, H. Four resistance alleles derived from Oryza longistaminata (A. Chev. & Roehrich) against green rice leafhopper, Nephotettix cincticeps (Uhler) identified using novel introgression lines. Breed Sci. 2019, 69, 573–584. [Google Scholar] [PubMed]
- Will, T.; van Bel, A.J. Physical and chemical interactions between aphids and plants. J. Exp. Bot. 2006, 57, 729–737. [Google Scholar] [CrossRef]
- Giordanengo, P.; Brunissen, L.; Rusterucci, C.; Vincent, C.; van Bel, A.; Dinant, S.; Girousse, C.; Faucher, M.; Bonnemain, J.L. Compatible plant-aphid interactions: How aphids manipulate plant responses. Comptes Rendus Biol. 2010, 333, 516–523. [Google Scholar] [CrossRef] [PubMed]
- Cun, Z. Identification of New Chickpea Virus and Control of Chickpea Virus Disease. Evid-Based Complment. Altern. Med 2022, 2022, 6465505. [Google Scholar] [CrossRef]
- Hidayat, P.; Harleni, H.; Maharani, Y.; Triwidodo, H. Biology and demography statistic aphids Rhopalosiphum rufiabdominale (Sasaki) and Tetraneura nigriabdominalis (Sasaki) (Hemiptera: Aphididae) in rice roots. Indones. J. Entomol. 2019, 16, 180–186. [Google Scholar] [CrossRef]
- Cranshaw, W.; Wainwright-Evans, S. Cannabis sativa as a host of rice root aphid (Hemiptera: Aphididae) in North America. J. Integr. Pest Manag. 2020, 11, 15. [Google Scholar] [CrossRef]
- War, A.R.; Taggar, G.K.; Hussain, B.; Taggar, M.S.; Nair, R.M.; Sharma, H.C. Plant defence against herbivory and insect adaptations. AoB Plants 2018, 10, ply037. [Google Scholar]
- Alborn, H.T.; Hansen, T.V.; Jones, T.H.; Bennett, D.C.; Tumlinson, J.H.; Schmelz, E.A.; Teal, P.E. Disulfooxy fatty acids from the American bird grasshopper Schistocerca americana, elicitors of plant volatiles. Proc. Natl. Acad. Sci. USA 2007, 104, 12976–12981. [Google Scholar] [CrossRef]
- Gust, A.A.; Nürnberger, T. A plant surface receptor for sensing insect herbivory. Proc. Natl. Acad. Sci. USA 2020, 117, 32839–32841. [Google Scholar] [CrossRef]
- Malook, S.U.; Maqbool, S.; Hafeez, M.; Karunarathna, S.C.; Suwannarach, N. Molecular and Biochemical Mechanisms of Elicitors in Pest Resistance. Life 2022, 12, 844. [Google Scholar] [CrossRef]
- Coppola, M.; Di Lelio, I.; Romanelli, A.; Gualtieri, L.; Molisso, D.; Ruocco, M.; Avitabile, C.; Natale, R.; Cascone, P.; Guerrieri, E.; et al. Tomato plants treated with systemin peptide show enhanced levels of direct and indirect defense associated with increased expression of defense-related genes. Plants 2019, 8, 395. [Google Scholar] [CrossRef]
- Steinbrenner, A.D.; Muñoz-Amatriaín, M.; Venegas, J.M.A.; Lo, S.; Shi, D.; Holton, N.; Zipfel, C.; Abagyan, R.; Huffaker, A.; Close, T.J.; et al. A receptor for herbivore-associated molecular patterns mediates plant immunity. Biorxiv 2019, 679803. [Google Scholar]
- Klauser, D.; Desurmont, G.A.; Glauser, G.; Vallat, A.; Flury, P.; Boller, T.; Turlings, T.C.; Bartels, S. The Arabidopsis Pep-PEPR system is induced by herbivore feeding and contributes to JA-mediated plant defence against herbivory. J. Exp. Bot. 2015, 66, 5327–5336. [Google Scholar] [CrossRef]
- Gouhier-Darimont, C.; Stahl, E.; Glauser, G.; Reymond, P. The Arabidopsis lectin receptor kinase LecRK-I 8 is involved in insect egg perception. Front. Plant Sci. 2019, 10, 623. [Google Scholar] [CrossRef]
- Chen, C.Y.; Mao, Y.B. Research advances in plant–insect molecular interaction. F1000Res 2020, 9, 198. [Google Scholar] [CrossRef]
- Stotz, H.U.; Koch, T.; Biedermann, A.; Weniger, K.; Boland, W.; Mitchell-Olds, T. Evidence for regulation of resistance in Arabidopsis to Egyptian cotton worm by salicylic and jasmonic acid signaling pathways. Planta 2002, 214, 648–652. [Google Scholar] [CrossRef]
- Alfonso, E.; Stahl, E.; Glauser, G.; Bellani, E.; Raaymakers, T.M.; Van den Ackerveken, G.; Zeier, J.; Reymond, P. Insect eggs trigger systemic acquired resistance against a fungal and an oomycete pathogen. New Phytol. 2021, 232, 2491–2505. [Google Scholar] [CrossRef]
- Mollah, M.M.I.; Choi, H.W.; Yeam, I.; Lee, J.M.; Kim, Y. Salicylic acid, a plant hormone, suppresses phytophagous insect immune response by interrupting HMG-Like DSP1. Front. Physiol. 2021, 12, 744272. [Google Scholar] [CrossRef]
- Wu, J.; Hettenhausen, C.; Meldau, S.; Baldwin, I.T. Herbivory rapidly activates MAPK signaling in attacked and unattacked leaf regions but not between leaves of Nicotiana attenuata. Plant Cell 2007, 19, 1096–1122. [Google Scholar] [CrossRef]
- Kundu, P.; Vadassery, J. Role of WRKY transcription factors in plant defense against lepidopteran insect herbivores: An overview. J. Plant Biochem. Biotechnol. 2021, 30, 698–707. [Google Scholar] [CrossRef]
- Sperdouli, I.; Andreadis, S.S.; Adamakis, I.D.S.; Moustaka, J.; Koutsogeorgiou, E.I.; Moustakas, M. Reactive Oxygen Species Initiate Defence Responses of Potato Photosystem II to Sap-Sucking Insect Feeding. Insects 2022, 13, 409. [Google Scholar] [CrossRef] [PubMed]
- Mostafa, S.; Wang, Y.; Zeng, W.; Jin, B. Plant responses to Herbivory, Wounding, and infection. Int. J. Mol. Sci. 2022, 23, 7031. [Google Scholar] [CrossRef] [PubMed]
- Parmagnani, A.S.; Maffei, M.E. Calcium Signaling in Plant-Insect Interactions. Plants 2022, 11, 2689. [Google Scholar] [CrossRef]
- Heyer, M.; Scholz, S.S.; Reichelt, M.; Kunert, G.; Oelmüller, R.; Mithöfer, A. The Ca2+ sensor proteins CML37 and CML42 antagonistically regulate plant stress responses by altering phytohormone signals. Plant Mol. Biol. 2022, 109, 611–625. [Google Scholar] [CrossRef]
- Barrera, S.E.; Sarango-Flóres, S.W.; Montenegro-Gómez, S.P. The phyllosphere microbiome and its potential application in horticultural crops. A review. Rev. Colomb. Cienc. Hortícolas 2019, 13, 384–396. [Google Scholar] [CrossRef]
- Vacher, C.; Hampe, A.; Porté, A.J.; Sauer, U.; Compant, S.; Morris, C.E. The Phyllosphere: Microbial Jungle at the Plant–Climate Interface. Annu. Rev. Ecol. Evol. Syst. 2016, 47, 1–24. [Google Scholar] [CrossRef]
- Ruinen, J. The phyllosphere: I. An ecologically neglected milieu. Plant Soil 1961, 15, 81–109. [Google Scholar] [CrossRef]
- Morris, C.E. Phyllosphere. eLS 2002, 1–8. [Google Scholar]
- Doan, H.K.; Leveau, J.H. Artificial surfaces in phyllosphere microbiology. Phytopathology 2015, 105, 1036–1042. [Google Scholar] [CrossRef]
- Thapa, S.; Prasanna, R. Prospecting the characteristics and significance of the phyllosphere microbiome. Ann. Microbiol. 2018, 68, 229–245. [Google Scholar] [CrossRef]
- Morris, C.; Kinkel, L. Phyllosphere microbiology. In Fifty Years of Phyllosphere Microbiology: Significant Contributions to Research in Related Fields; Lindow, S.E., Hecht-Poinar, E.I., Elliott, V.J., Eds.; APS Press: St. Paul, MO, USA, 2002; pp. 365–375. [Google Scholar]
- Gouka, L.; Raaijmakers, J.M.; Cordovez, V. Ecology and functional potential of phyllosphere yeasts. Trends Plant Sci. 2022, 27, 1109–1123. [Google Scholar] [CrossRef]
- Rao, N.S.S. Soil Microbiology. Oxford & IBH Publishing Co. Pvt Ltd.: New Delhi, India, 1999. [Google Scholar]
- Yurimoto, H.; Shiraishi, K.; Saka, Y. Physiology of Methylotrophs Living in the Phyllosphere. Microorganisms 2021, 9, 809. [Google Scholar] [CrossRef]
- Abanda-Nkpwatt, D.; Musch, M.; Tschiersch, J.; Boettner, M.; Schwab, W. Molecular interaction between Methylobacterium extorquens and seedlings: Growth promotion, methanol consumption, and localization of the methanol emission site. J. Exp. Bot. 2006, 57, 4025–4032. [Google Scholar] [CrossRef]
- Iguchi, H.; Yurimoto, H.; Sakai, Y. Interactions of methylotrophs with plants and other heterotrophic bacteria. Microorganisms 2015, 3, 137–151. [Google Scholar] [CrossRef]
- Gibson, L.; Crombie, A.T.; McNamara, N.P.; Murrell, J.C. Isoprene-degrading bacteria associated with the phyllosphere of Salix fragilis, a high isoprene-emitting willow of the Northern Hemisphere. Environ. Microbiol 2021, 16, 17. [Google Scholar] [CrossRef]
- Crombie, A.T.; Larke-Mejia, N.L.; Emery, H.; Dawson, R.; Pratscher, J.; Murphy, G.P.; McGenity, T.J.; Murrell, J.C. Poplar phyllosphere harbors disparate isoprene-degrading bacteria. Proc. Natl. Acad. Sci. USA 2018, 115, 13081–130816. [Google Scholar] [CrossRef]
- Carrión, O.; Gibson, L.; Elias, D.M.O.; McNamara, N.P.; van Alen, T.A.; Huub, J.M.; Op den Camp, H.J.; Supramaniam, C.V.; McGenity, T.J.; Murrell, J.C. Diversity of isoprene-degrading bacteria in phyllosphere and soil communities from a high isoprene-emitting environment: A Malaysian oil palm plantation. Microbiome 2020, 8, 81. [Google Scholar] [CrossRef]
- Papen, H.; Gessler, A.; Zumbusch, E.; Rennenberg, H. Chemolithoautotrophic nitrifiers in the phyllosphere of a spruce ecosystem receiving high atmospheric nitrogen input. Curr. Microbiol. 2002, 44, 56–60. [Google Scholar] [CrossRef]
- Fu¨rnkranz, M.; Wane, W.; Richter, A.; Abell, G.; Rasche, F.; Sessitsch, A. Nitrogen fixation by phyllosphere bacteria associated with higher plants and their colonizing epiphytes of a tropical lowland rainforest of Costa Rica. ISME J. 2008, 2, 561–570. [Google Scholar] [CrossRef]
- Thompson, I.P.; Bailey, M.J.; Fenlon, J.S.; Fermor, T.R.; Lilley, A.K.; Lynch, J.M.; McCormack, P.J.; McQuilken, M.P.; Purdy, K.J.; Rainey, P.B.; et al. Quantitative and qualitative seasonal changes in the microbial community from the phyllosphere of sugar beet (Beta vulgaris). Plant Soil 1993, 150, 177–191. [Google Scholar] [CrossRef]
- Bringel, F.; Couée, I. Pivotal roles of phyllosphere microorganisms at the interface between plant functioning and atmospheric trace gas dynamics. Front. Microbiol. 2015, 6, 486. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, J.L.; Sundin, G.W. Effect of Solar UV-B Radiation on a Phyllosphere Bacterial Community. Appl. Environ. Microbiol. 2001, 67, 5488–5496. [Google Scholar] [CrossRef] [PubMed]
- Elad, Y. Effect of climate change on phyllosphere microflora: Plant-microorganism interactions. Climate Change: Global Risks, Challenges and Decisions. In IOP Conference Series. Earth and Environmental Science; IOP Publishing: Bristol, UK, 2009; p. 6. [Google Scholar]
- Aydogan, E.L.; Moser, G.; Müller, C.; Kämpfer, P.; Glaeser, S.P. Long-Term Warming Shifts the Composition of Bacterial Communities in the Phyllosphere of Galium album in a Permanent Grassland Field-Experiment. Front Microbiol. 2018, 9, 144. [Google Scholar] [CrossRef]
- Brighigna, L.; Gori, A.; Gonneli, S.; Favilli, F. The influence of air pollution on the phyllosphere microflora composition of Tillandsia leaves (Bromeliaceae). Rev. Bio. Trop. 2000, 48, 511–517. [Google Scholar] [CrossRef]
- Sivakumar, N.; Sathishkumar, R.; Selvakumar, G.; Shyamkumar, R.; Arjunekumar, K. Phyllospheric Microbiomes: Diversity, Ecological Significance, and Biotechnological Applications. In Plant Microbiomes for Sustainable Agriculture; Yadav, A.N., Singh, J., Rastegari, A.A., Yadav, N., Eds.; Springer Nature: Cham, Switzerland, 2020; Volume 25, pp. 113–172. [Google Scholar]
- Lagos, M.L.; Maruyama, F.; Nannipieri, P.; Mora, M.L.; Ogram, A.; Jorquera, M.A. Current overview on the study of bacteria in the rhizosphere by modern molecular techniques: A mini-review. J. Soil Sci. Plant Nutr. 2015, 15, 504–523. [Google Scholar]
- Wang, Y.; Zhang, M.; Li, S.; Li, P.; Lang, Z. Effects of Insect-Resistant Maize HGK60 on Community Diversity of Bacteria and Fungi in Rhizosphere Soil. Plants 2022, 11, 2824. [Google Scholar] [CrossRef]
- Prashar, P.; Kapoor, N.; Sachdeva, S. Rhizosphere: Its structure, bacterial diversity and significance. Rev. Environ. Sci. Biotechnol. 2014, 13, 63–77. [Google Scholar] [CrossRef]
- Hiltner, L. About new experiences and problems in the field of Bodenbakteriologie. Works Ger. Agric. Soc. 1904, 98, 59–78. [Google Scholar]
- Mendes, R.; Garbeva, P.; Raaijmakers, J.M. The rhizosphere microbiome: Significance of plant beneficial, plant pathogenic, and human pathogenic microorganisms. FEMS Microbiol. Rev. 2013, 37, 634–663. [Google Scholar] [CrossRef]
- Shaikh, S.S.; Sayyed, R.Z.; Reddy, M.S. Plant Growth-Promoting Rhizobacteria: An Eco-friendly Approach for Sustainable Agroecosystem. In Plant, Soil and Microbes; Hakeem, K.R., Akhtar, M.S., Abdullah, S.N.A., Eds.; Springer International Publishing: Berlin/Heidelberg, Germany, 2016; Volume 1: Implication in Crop Science, pp. 181–201. [Google Scholar]
- Wang, E.; Liu, X.; Si, Z.; Li, X.; Bi, J.; Dong, W.; Chen, M.; Wang, S.; Zhang, J.; Song, A.; et al. Volatile Organic Compounds from Rice Rhizosphere Bacteria Inhibit Growth of the Pathogen Rhizoctonia solani. Agriculture 2021, 11, 368. [Google Scholar] [CrossRef]
- Rashid, M.H.O.; Chung, Y.R. Induction of systemic resistance against insect herbivores in plants by beneficial soil microbes. Front. Plant Sci. 2017, 8, 1816. [Google Scholar] [CrossRef]
- El-Maraghy, S.S.; Tohamy, A.T.; Hussein, K.A. Plant protection properties of the Plant Growth-Promoting Fungi (PGPF): Mechanisms and potentiality. Curr. Res. Environ. Appl. Mycol. 2021, 11, 391–415. [Google Scholar] [CrossRef]
- Akinola, S.A.; Babalola, O.O. The fungal and archaeal community within plant rhizosphere: A review on their contribution to crop safety. J. Plant Nutri. 2021, 44, 600–618. [Google Scholar] [CrossRef]
- Cui, J.; Bai, L.; Liua, X.; Jie, W.; Cai, B. Arbuscular mycorrhizal fungal communities in the rhizosphere of a continuous cropping soybean system at the seedling stage. Braz. J. Microbiol. 2018, 49, 240–247. [Google Scholar] [CrossRef]
- Powell, J.R.; Rillig, M.C. Biodiversity of arbuscular mycorrhizal fungi and ecosystem function. New Phytol. 2018, 220, 1059–1075. [Google Scholar] [CrossRef]
- Dong, C.J.; Wang, L.L.; Li, Q.; Shang, Q.M. Bacterial communities in the rhizosphere, phyllosphere and endosphere of tomato plants. PLoS ONE 2019, 14, e0223847. [Google Scholar] [CrossRef]
- Simon, M.; Hilker, M. Does Rust Infection of Willow Affect Feeding and Oviposition Behavior of Willow Leaf Beetles? J. Insect Behav. 2005, 18, 115–129. [Google Scholar] [CrossRef]
- Martin, D.S.; Jones, C.P.; Yao, K.F.; Lee, L.E. An epiphytic yeast (Sporobolomyces roseus) influencing in oviposition preference of the European corn borer (Ostrinia nubilalis) on maize. Acta Oecol. 1993, 14, 563–574. [Google Scholar]
- Blandino, M.; Scarpino, V.; Vanara, F.; Sulyok, M.; Krska, R.; Reyneri, A. Role of the European corn borer (Ostrinia nubilalis) on contamination of maize with 13 Fusarium mycotoxins. Food addit. Contam. Chem. Annal. 2015, 32, 533–543. [Google Scholar] [CrossRef]
- Pineda, A.; Zheng, S.J.; van Loon, J.J.A.; Dicke, M. Rhizobacteria modify plant–aphid interactions: A case of induced systemic susceptibility. Plant Biol. 2012, 14 (Suppl. S1), 83–90. [Google Scholar] [CrossRef] [PubMed]
- Disi, J.; Simmons, J.; Zebelo, S. Plant Growth-Promoting Rhizobacteria-Induced Defense Against Insect Herbivores. In Field Crops: Sustainable Management by PGPR; Maheshwari, D.K., Dheeman, S., Eds.; Springer Nature: Cham, Switzerland, 2019; pp. 385–410. [Google Scholar]
- Van de Mortel, J.E.; de Vos, R.C.; Dekkers, E.; Pineda, A.; Guillod, L.; Bouwmeester, K.; van Loon, J.J.; Dicke, M.; Raaijmakers, J.M. Metabolic and transcriptomic changes induced in Arabidopsis by the rhizobacterium Pseudomonas fluorescens SS101. Plant Physiol. 2012, 160, 2173–2188. [Google Scholar] [CrossRef]
- Bano, A.; Muqarab, R. Plant defence induced by PGPR against Spodoptera litura in tomato (Solanum lycopersicum L.). Plant Biol. 2017, 19, 406–412. [Google Scholar] [CrossRef] [PubMed]
- Zebelo, S.; Song, Y.; Kloepper, J.W.; Fadamiro, H. Rhizobacteria activates (+)-δ-cadinene synthase genes and induces systemic resistance in cotton against beet armyworm (Spodoptera exigua). Plant Cell Environ. 2016, 39, 935–943. [Google Scholar] [CrossRef] [PubMed]
- Naeem, M.; Aslam, Z.; Khaliq, A.; Ahmed, J.N.; Nawaz, A.; Hussain, M. Plant growth promoting rhizobacteria reduce aphid population and enhance the productivity of bread wheat. Braz. J. Microbiol. 2018, 49 (Suppl. S1), 9–14. [Google Scholar] [CrossRef]
- Shavit, R.; Ofek-Lalzar, M.; Burdman, S.; Morin, S. Inoculation of tomato plants with rhizobacteria enhances the performance of the phloem-feeding insect Bemisia tabaci . Front. Plant Sci. 2013, 4, 306. [Google Scholar] [CrossRef]
- Valenzuela-Soto, J.H.; Estrada-Hernández, M.G.; Ibarra-Laclette, E.; Délano-Frier, J.P. Inoculation of tomato plants (Solanum lycopersicum) with growth-promoting Bacillus subtilis retards whitefly Bemisia tabaci development. Planta 2010, 231, 397–410. [Google Scholar] [CrossRef]
- Zehnder, G.; Kloepper, J.; Yao, C.; Wei, G. Induction of systemic resistance in cucumber against cucumber beetles (Coleoptera: Chrysomelidae) by plant growth promoting rhizobacteria. J. Econ. Entomol. 1997, 90, 391–396. [Google Scholar] [CrossRef]
- Pangesti, N.; Reichelt, M.; van de Mortel, J.E.; Kapsomenou, E.; Gershenzon, J.; van Loon, J.J.; Dicke, M.; Pineda, A. Jasmonic acid and ethylene signaling pathways regulate glucosinolate levels in plants during rhizobacteria-induced systemic resistance against a leaf-chewing herbivore. J. Chem. Ecol. 2016, 42, 1212–1225. [Google Scholar]
- Pangesti, N.; Pineda, A.; Dicke, M.; Van Loon, J.J.A. Variation in plant-mediated interactions between rhizobacteria and caterpillars: Potential role of soil composition. Plant Biol. 2015, 17, 474–483. [Google Scholar] [CrossRef]
- Ling, S.; Zhao, Y.; Sun, S.; Zheng, D.; Sun, X.; Zeng, R.; Chen, D.; Song, Y. Enhanced anti-herbivore defense of tomato plants against Spodoptera litura by their rhizosphere bacteria. BMC Plant Biol. 2022, 22, 254. [Google Scholar] [CrossRef]
- Selvaraj, A.; Thangavel, K.; Uthandi, S. Arbuscular mycorrhizal fungi (Glomus intraradices) and diazotrophic bacterium (Rhizobium BMBS) primed defense in blackgram against herbivorous insect (Spodoptera litura) infestation. Microbiol. Res. 2020, 231, 126355. [Google Scholar] [CrossRef]
- Jiang, D.; Tan, M.; Wu, S.; Zheng, L.; Wang, Q.; Wang, G.; Yan, S. Defense responses of arbuscular mycorrhizal fungus-colonized poplar seedlings against gypsy moth larvae: A multiomics study. Hortic. Res. 2021, 8, 245. [Google Scholar] [CrossRef]
- Barber, N.A.; Kiers, E.T.; Hazzard, R.V.; Adler, L.S. Context-dependency of arbuscular mycorrhizal fungi on plant–insect interactions in an agroecosystem. Front. Plant. Sci. 2013, 4, 338. [Google Scholar] [CrossRef]
- Bhar, A.; Chakraborty, A.; Roy, A. Plant responses to biotic stress: Old memories matter. Plants 2022, 11, 84. [Google Scholar] [CrossRef]
- Rangel, L.I.; Bolton, M.D. The unsung roles of microbial secondary metabolite effectors in the plant disease cacophony. Curr. Opin. Plant Biol. 2022, 68, 102233. [Google Scholar] [CrossRef]
- Mayoral-Peña, Z.; Lázaro-Vidal, V.; Fornoni, J.; Álvarez-Martínez, R.; Garrido, E. Studying Plant–Insect Interactions through the Analyses of the Diversity, Composition, and Functional Inference of Their Bacteriomes. Microorganisms 2022, 11, 40. [Google Scholar] [CrossRef]
- Koprivova, A.; Kopriva, S. Plant secondary metabolites altering root microbiome composition and function. Curr. Opin. Plant Biol. 2022, 67, 102227. [Google Scholar] [CrossRef]


| Rhizosphere Microbes | Examples | Reference |
|---|---|---|
| Bacteria | More common genera found in the rhizosphere are Pseudomonas, Bacillus, Azotobacter, Arthobacter, Rhizobium, Agrobacterium, Flavobacter, and Micrococcus | Prashar et al. [76] |
| Plant growth-promoting rhizobacteria (PGPR) are bacteria that promote plant growth. Nitrogen fixing bacteria are present. | Shaikh et al. [79] Mendes et al. [78] | |
| Fungi | Plant growth-promoting fungi (PGPF) are also present in the rhizosphere | Rashid et al. [81] |
| Mycorrhizal fungi are present in rhizosphere | Cui et al. [84] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Das, T.; Bhattacharyya, A.; Bhar, A. Linking Phyllosphere and Rhizosphere Microbiome to the Plant–Insect Interplay: The New Dimension of Tripartite Interaction. Physiologia 2023, 3, 129-144. https://doi.org/10.3390/physiologia3010011
Das T, Bhattacharyya A, Bhar A. Linking Phyllosphere and Rhizosphere Microbiome to the Plant–Insect Interplay: The New Dimension of Tripartite Interaction. Physiologia. 2023; 3(1):129-144. https://doi.org/10.3390/physiologia3010011
Chicago/Turabian StyleDas, Tanaya, Anindya Bhattacharyya, and Anirban Bhar. 2023. "Linking Phyllosphere and Rhizosphere Microbiome to the Plant–Insect Interplay: The New Dimension of Tripartite Interaction" Physiologia 3, no. 1: 129-144. https://doi.org/10.3390/physiologia3010011