Characterization of Redox Sensitive Brown Algal Mannitol-1-Phosphatases


Abstract
:1. Introduction
2. Materials and Methods
2.1. Mannitol Production by Ectocarpus sp. M1Pases in Escherichia coli
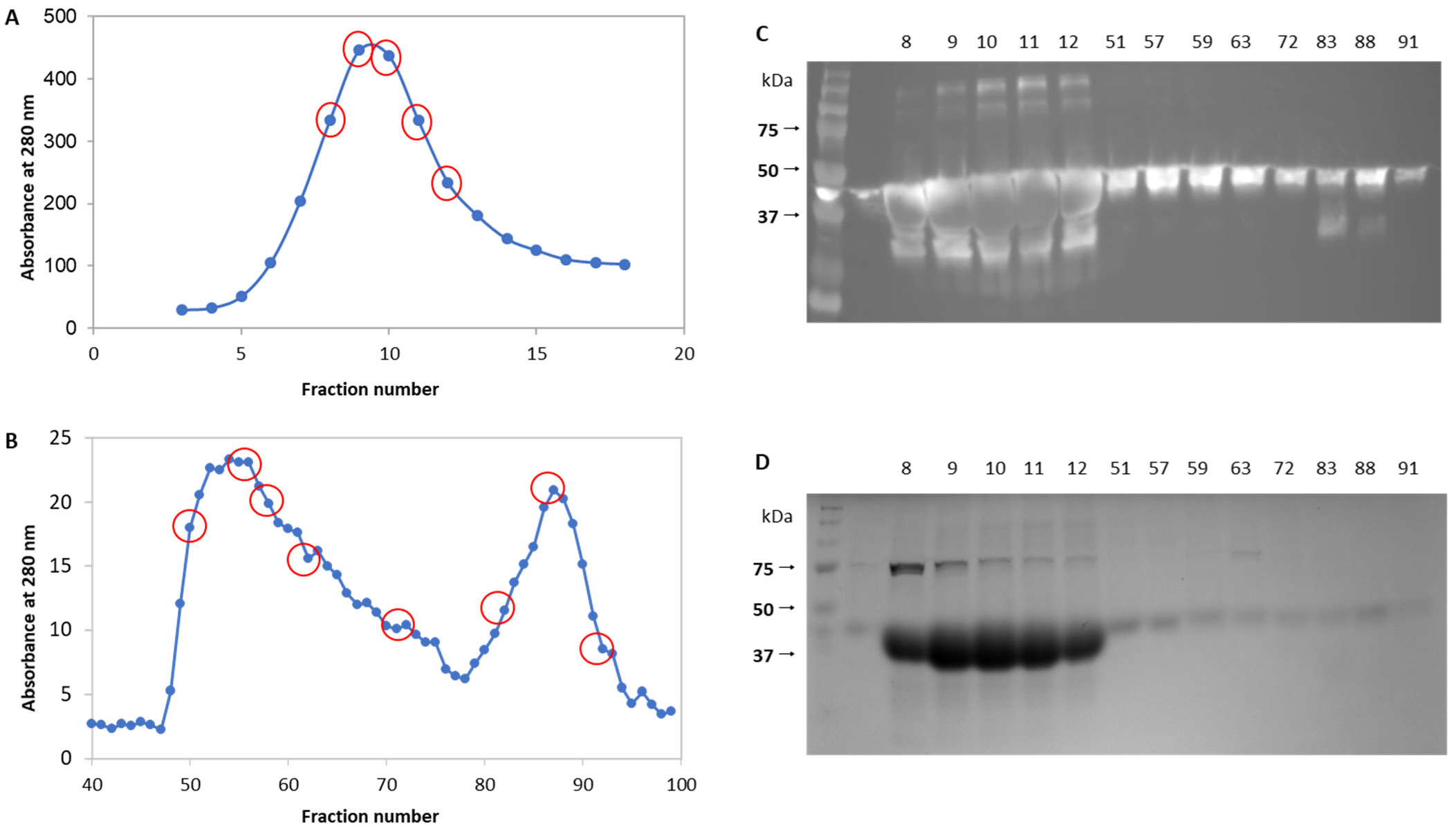
2.2. Production and Purification of EsM1Pase1short
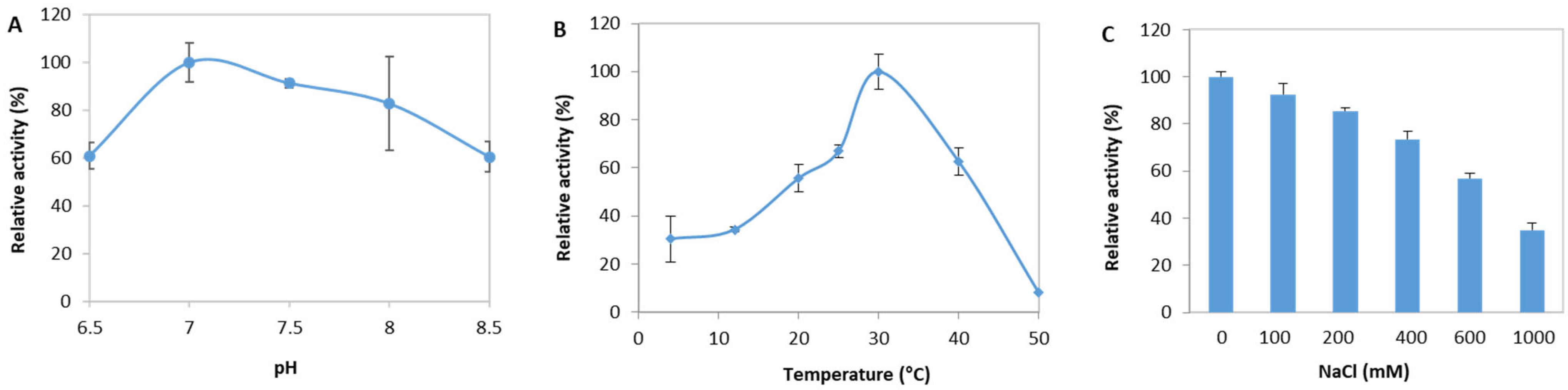
2.3. Biochemical Characterization of EsM1Pase1short
2.4. Retrieval of Sequences, Prediction of Peptide Signal, Determination of Potential Subcellular Localization, and Phylogenetic Analysis
3. Results and Discussion
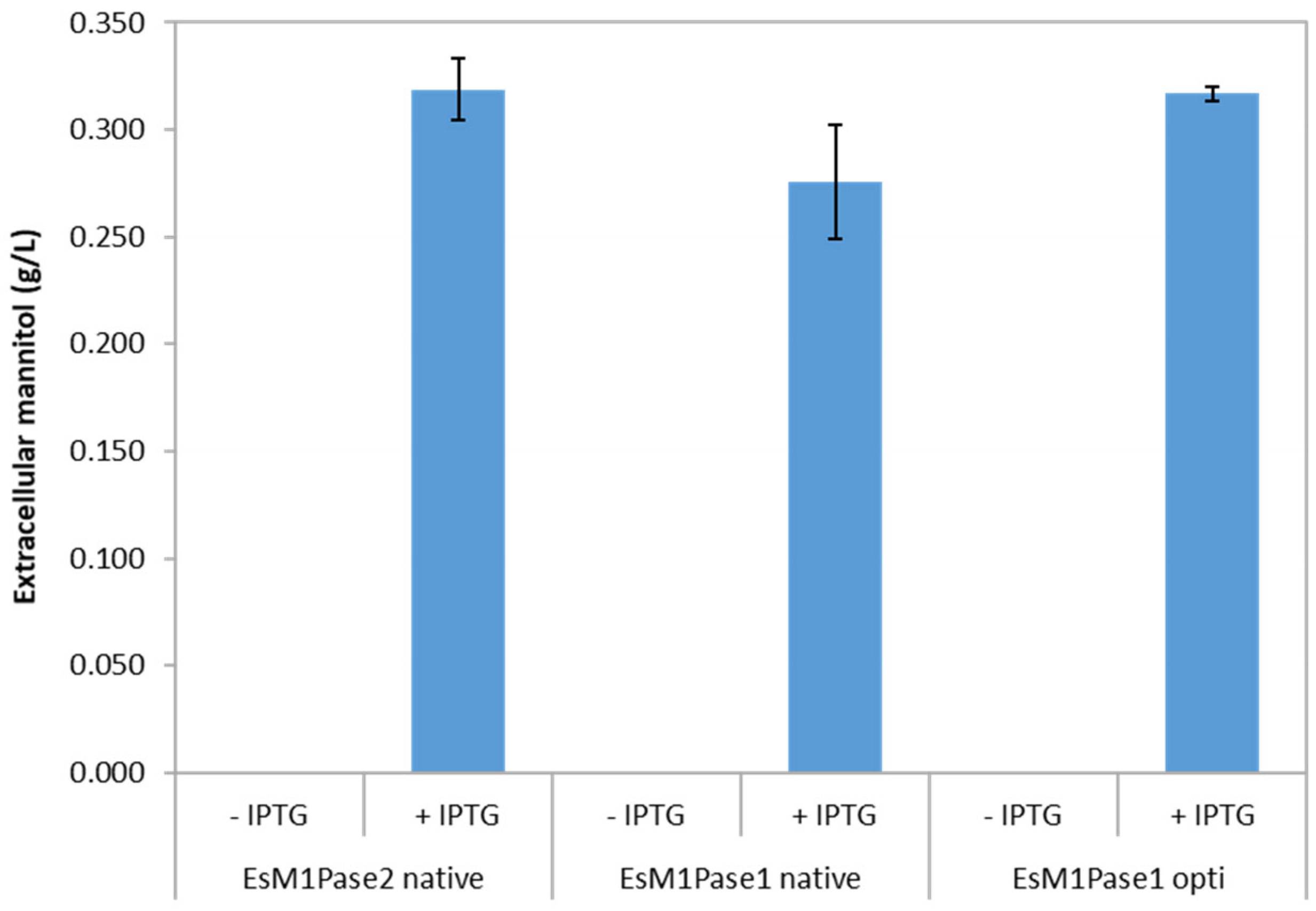
3.1. EsM1Pase1 Is a Bona Fide M1Pase
3.2. Biochemical Characterization of EsM1Pase1short Recombinant Proteins
3.3. Evolution and Potential Localization of M1Pases in Brown Algae
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Krause-Jensen, D.; Lavery, P.; Serrano, O.; Marbà, N.; Masque, P.; Duarte, C.M. Sequestration of macroalgal carbon: The elephant in the Blue Carbon room. Biol. Lett. 2018, 14, 20180236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawai, S.; Murata, K. Biofuel production based on carbohydrates from both brown and red macroalgae: Recent developments in key biotechnologies. Int. J. Mol. Sci. 2016, 17, 145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khalil, H.A.; Saurabh, C.K.; Tye, Y.Y.; Lai, T.K.; Easa, A.M.; Rosamah, E.; Fazita, M.R.N.; Syakir, M.I.; Adnan, A.S.; Fizree, H.M.; et al. Seaweed based sustainable films and composites for food and pharmaceutical applications: A review. Renew. Sust. Energ. Rev. 2017, 77, 353–362. [Google Scholar] [CrossRef]
- Sudhakar, K.; Mamat, R.; Samykano, M.; Azmi, W.H.; Ishak, W.F.W.; Yusaf, T. An overview of marine macroalgae as bioresource. Renew. Sust. Energ. Rev. 2018, 91, 165–179. [Google Scholar] [CrossRef]
- Wei, N.; Quarterman, J.; Jin, Y.S. Marine macroalgae: An untapped resource for producing fuels and chemicals. Trends Biotechnol. 2013, 31, 70–77. [Google Scholar] [CrossRef]
- Rioux, L.-E.; Turgeon, S. Seaweed carbohydrates. In Seaweed Sustainability: Food and Non-Food Applications Book; Academic Press: Cambridge, MA, USA, 2018; pp. 141–192. [Google Scholar]
- Reed, R.H.; Davison, I.R.; Chudek, J.A.; Foster, R. The osmotic role of mannitol in the Phaeophyta: An appraisal. Phycologia 1985, 24, 35–47. [Google Scholar] [CrossRef]
- Cock, J.M.; Sterck, L.; Rouzé, P.; Scornet, D.; Allen, A.E.; Amoutzias, G.; Anthouard, V.; Artiguenave, F.; Aury, J.M.; Badger, J.H.; et al. The Ectocarpus genome and the independent evolution of multicellularity in the brown algae. Nature 2010, 465, 617–621. [Google Scholar] [CrossRef] [Green Version]
- Ye, N.; Zhang, X.; Miao, M.; Fan, X.; Zheng, Y.; Xu, D.; Wang, J.; Zhou, L.; Wang, D.; Gao, Y. Saccharina genomes provide novel insight into kelp biology. Nat. Commun. 2015, 6, 6986. [Google Scholar] [CrossRef] [Green Version]
- Tenhaken, R.; Voglas, E.; Cock, J.M.; Neu, V.; Huber, C.G. Characterization of GDP-mannose dehydrogenase from the brown alga Ectocarpus siliculosus providing the precursor for the alginate polymer. J. Biol. Chem. 2011, 286, 16707–16715. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P.; Shao, Z.; Jin, W.; Duan, D. Comparative characterization of two GDP-mannose dehydrogenase genes from Saccharina japonica (Laminariales, Phaeophyceae). BMC Plant Biol. 2016, 16, 62. [Google Scholar] [CrossRef]
- Chi, S.; Liu, T.; Wang, X.; Wang, R.; Wang, S.; Wang, G.; Shan, G.; Liu, C. Functional genomics analysis reveals the biosynthesis pathways of important cellular components (alginate and fucoidan) of Saccharina. Curr. Genet. 2018, 64, 259–273. [Google Scholar] [CrossRef] [Green Version]
- Fischl, R.; Bertelsen, K.; Gaillard, F.; Coelho, S.; Michel, G.; Klinger, M.; Boyen, C.; Czjzek, M.; Hervé, C. The cell-wall active mannuronan C5-epimerases in the model brown alga Ectocarpus: From gene context to recombinant protein. Glycobiology 2016, 26, 973–983. [Google Scholar] [CrossRef] [Green Version]
- Inoue, A.; Satoh, A.; Morishita, M.; Tokunaga, Y.; Miyakawa, T.; Tanokura, M.; Ojima, T. Functional heterologous expression and characterization of mannuronan C5-epimerase from the brown alga Saccharina japonica. Algal Res. 2016, 16, 282–291. [Google Scholar] [CrossRef] [Green Version]
- Pengyan, Z.; Chang, L.; Zhanru, S.; Fuli, L.; Jianting, Y.; Delin, D. Genome-wide transcriptome profiling and characterization of mannuronan C5-epimerases in Saccharina japonica. Algal Res. 2021, 60, 102491. [Google Scholar] [CrossRef]
- Inoue, A.; Ojima, T. Functional identification of alginate lyase from the brown alga Saccharina japonica. Sci. Rep. 2019, 9, 4937. [Google Scholar] [CrossRef] [Green Version]
- Inoue, A.; Ojima, T. Functional identification of the 4-deoxy-L-erythro-5-hexoseulose uronate reductase from a brown alga, Saccharina japonica. Biochem. Biophys. Res. Commun. 2021, 545, 112–118. [Google Scholar] [CrossRef]
- Groisillier, A. Cloning and expression strategies for the post-genomic analysis of brown algae. In Protocols for Macroalgae Research; CRC Press: Boca Raton, FL, USA, 2018; pp. 453–468. [Google Scholar]
- Gravot, A.; Dittami, S.M.; Rousvoal, S.; Lugan, R.; Eggert, A.; Collén, J.; Boyen, C.; Bouchereau, A.; Tonon, T. Diurnal oscillations of metabolite abundances and genomes analysis provide new insights into central metabolic processes of the brown alga Ectocarpus siliculosus. New Phytol. 2010, 188, 98–110. [Google Scholar] [CrossRef]
- Schiener, P.; Black, K.D.; Stanley, M.S.; Green, D.H. The seasonal variation in the chemical composition of the kelp species Laminaria digitata, Laminaria hyperborea, Saccharina latissima and Alaria esculenta. J. Appl. Phycol. 2015, 27, 363–373. [Google Scholar] [CrossRef]
- Michel, G.; Tonon, T.; Scornet, D.; Cock, J.M.; Kloareg, B. Central and storage carbon metabolism of the brown alga Ectocarpus siliculosus: Insights into the origin and evolution of storage carbohydrates in Eukaryotes. New Phytol. 2010, 188, 82–97. [Google Scholar] [CrossRef]
- Rousvoal, S.; Groisillier, A.; Dittami, S.M.; Michel, G.; Boyen, C.; Tonon, T. Mannitol-1-phosphate dehydrogenase activity in Ectocarpus siliculosus, a key role for mannitol synthesis in brown algae. Planta 2011, 233, 261–273. [Google Scholar] [CrossRef]
- Bonin, P.; Groisillier, A.; Raimbault, A.; Guibert, A.; Boyen, C.; Tonon, T. Molecular and biochemical characterization of mannitol-1-phosphate dehydrogenase from the model brown alga Ectocarpus sp. Phytochemistry 2015, 117, 509–520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Groisillier, A.; Shao, Z.; Michel, G.; Goulitquer, S.; Bonin, P.; Krahulec, S.; Nidetzky, B.; Duan, D.; Boyen, C.; Tonon, T. Mannitol metabolism in brown algae involves a new phosphatase family. J. Exp. Bot. 2014, 65, 559–570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rathor, P.; Borza, T.; Liu, Y.; Qin, Y.; Stone, S.; Zhang, J.; Hui, J.P.; Berrue, F.; Groisillier, A.; Tonon, T.; et al. Low mannitol concentrations in Arabidopsis thaliana expressing Ectocarpus genes improve salt tolerance. Plants 2020, 9, 1508. [Google Scholar] [CrossRef] [PubMed]
- Hanko, E.K.R.; Sherlock, G.; Minton, N.P.; Malys, N. Biosensor-informed engineering of Cupriavidus necator H16 for autotrophic D-mannitol production. Metab. Eng. 2022, 72, 24–34. [Google Scholar] [CrossRef] [PubMed]
- Chi, S.; Liu, T.; Liu, C.; Jin, Y.; Yin, H.; Xu, X.; Li, Y. Characterization of mannitol metabolism genes in Saccharina explains its key role in mannitol biosynthesis and evolutionary significance in Laminariales. bioRxiv 2018. Available online: https://www.biorxiv.org/content/10.1101/243402v1.article-info (accessed on 7 November 2022).
- Iwamoto, K.; Kawanobe, H.; Ikawa, T.; Shiraiwa, Y. Purification and characterization of mannitol-1-phosphatase dehydrogenase in the red alga Caloglossa continua. Plant Physiol. 2001, 133, 893–900. [Google Scholar] [CrossRef] [Green Version]
- Ikawa, T.; Watanabe, T.; Nisizawa, K. Enzymes involved in last steps of biosynthesis of mannitol in brown algae. Plant Cell Physiol. 1972, 13, 1017–1029. [Google Scholar]
- Liberator, P.; Anderson, J.; Feiglin, M.; Sardana, M.; Griffin, P.; Schmatz, D.; Myers, R.W. Molecular cloning and functional expression of mannitol-1-phosphatase from the apicomplexan parasite Eimeria tenella. J. Biol. Chem. 1998, 273, 4237–4244. [Google Scholar] [CrossRef] [Green Version]
- Sand, M.; Rodrigues, M.; González, J.M.; de Crécy-Lagard, V.; Santos, H.; Müller, V.; Averhoff, B. Mannitol-1-phosphate dehydrogenases/phosphatases: A family of novel bifunctional enzymes for bacterial adaptation to osmotic stress. Environ. Microbiol. 2015, 17, 711–719. [Google Scholar] [CrossRef]
- Madsen, M.A.; Semerdzhiev, S.; Amtmann, A.; Tonon, T. Engineering mannitol biosynthesis in Escherichia coli and Synechococcus sp. PCC 7002 using a green algal fusion protein. ACS Synth. Biol. 2018, 12, 2833–2840. [Google Scholar] [CrossRef] [Green Version]
- Gschloessl, B.; Guermeur, Y.; Cock, J.M. HECTAR: A method to predict subcellular targeting in heterokonts. BMC Bioinform. 2008, 9, 393. [Google Scholar] [CrossRef] [Green Version]
- Lemesle-Varloot, L.; Henrissat, B.; Gaboriaud, C.; Bissery, V.; Morgat, A.; Mornon, J.P. Hydrophobic cluster analysis: Procedures to derive structural and functional information from 2-D-representation of protein sequences. Biochimie 1990, 72, 555–574. [Google Scholar] [CrossRef]
- Groisillier, A.; Tonon, T. Determination of Recombinant Mannitol-1-phosphatase Activity from Ectocarpus sp. Bio-Protocol 2016, 6, e1896. [Google Scholar] [CrossRef]
- Johnson, M.T.J.; Carpenter, E.J.; Tian, Z.; Bruskiewich, R.; Burris, J.N.; Carrigan, C.T.; Chase, M.W.; Clarke, N.D.; Covshoff, S.; Depamphilis, C.W.; et al. Evaluating methods for isolating total RNA and predicting the success of sequencing phylogenetically diverse plant transcriptomes. PLoS ONE 2012, 11, e50226. [Google Scholar] [CrossRef]
- Gruber, A.; Rocap, G.; Kroth, G.; Armbrust, E.V.; Mock, T. Plastid proteome prediction for diatoms and other algae with secondary plastids of the red lineage. Plant J. 2015, 81, 519–528. [Google Scholar] [CrossRef] [Green Version]
- Tonon, T.; Li, Y.; McQueen-Mason, S.M. Mannitol biosynthesis in algae: More widespread and diverse than previously thought. New Phytol. 2017, 213, 1573–1579. [Google Scholar] [CrossRef] [Green Version]
- Seifried, A.; Bergeron, A.; Boivin, B.; Gohla, A. Reversible oxidation controls the activity and oligomeric state of the mammalian phosphoglycolate phosphatase AUM. Free Radic. Biol. Med. 2016, 97, 75–84. [Google Scholar] [CrossRef]
- Eggert, A.; Raimund, S.; Van Den Daele, K.; Karsten, U. Biochemical characterization of mannitol metabolism in the unicellular red alga Dixoniella grisea (Rhodellophyceae). Eur. J. Phycol. 2006, 41, 1–9. [Google Scholar] [CrossRef]
- Dittami, S.M.; Scornet, D.; Petit, J.L.; Ségurens, B.; Da Silva, C.; Corre, E.; Dondrup, M.; Glatting, K.H.; König, R.; Sterck, L.; et al. Global expression analysis of the brown alga Ectocarpus siliculosus (Phaeophycaea) reveals large-scale reprogramming of the transcriptome in response to abiotic stress. Genome Biol. 2009, 10, R66. [Google Scholar] [CrossRef] [Green Version]
- Poliner, E.; Panchy, N.; Newton, L.; Wu, G.; Lapinsky, A.; Bullard, B.; Zienkiewicz, A.; Benning, C.; Shiu, S.-H.; Farré, E.M. Transcriptional coordination of physiological responses in Nannochloropsis oceanica CCMP1779 under light/dark cycles. Plant J. 2015, 83, 1097–1113. [Google Scholar] [CrossRef]
- Badis, Y.; Scornet, D.; Harada, M.; Caillard, C.; Godfroy, O.; Raphalen, M.; Gachon, C.M.M.; Coelho, S.M.; Motomura, T.; Nagasato, C.; et al. Targeted CRISPR-Cas9-based gene knockouts in the model brown alga Ectocarpus. New Phytol. 2021, 231, 2077–2091. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mannitol-1-P | Glucose-1-P | Glucose-6-P | Fructose-6-P | Fructose-1-P | Mannose-6-P | |
|---|---|---|---|---|---|---|
| S. japonica (SjaM1Pase1) 1 | 100 | 27.5 | 21.3 | 8.7 | n.d. | n.d. |
| S. japonica (SjaM1Pase2) 1 | 89.8 | 100 | 69.6 | 34.8 | n.d. | n.d. |
| Ectocarpus sp. (EsM1Pase1short) 2 | 100 | 0 | 0 | 0 | 0 | 0 |
| Ectocarpus sp. (EsM1Pase2) 3 | 100 | 0 | 0 | 0 | 0 | 0 |
| Specific Activity (μmol mg Protein−1 min−1) | Km (mM) | Kcat (s−1) | pH | Optimum Temperature (°C) | |
|---|---|---|---|---|---|
| S. japonica (SjaM1Pase1) 1 | 144.93 | 0.83 | 128.02 | 8.5 | 50 |
| S. japonica (SjaM1Pase2) 1 | 6.60 | 0.02 | 8.14 | 8.5 | 30 |
| Ectocarpus sp. (EsM1Pase1short) 2 | 1.19 | 0.43 | 0.8 | 7.0 | 30 |
| Ectocarpus sp. (EsM1Pase2) | 0.08 2 (0.03) 3 | 1.3 2 (0.67) 3 | 0.05 2 (0.02) 3 | 7.0 3 | 30 3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Le Strat, Y.; Tonon, T.; Leblanc, C.; Groisillier, A. Characterization of Redox Sensitive Brown Algal Mannitol-1-Phosphatases. Phycology 2023, 3, 1-12. https://doi.org/10.3390/phycology3010001
Le Strat Y, Tonon T, Leblanc C, Groisillier A. Characterization of Redox Sensitive Brown Algal Mannitol-1-Phosphatases. Phycology. 2023; 3(1):1-12. https://doi.org/10.3390/phycology3010001
Chicago/Turabian StyleLe Strat, Yoran, Thierry Tonon, Catherine Leblanc, and Agnès Groisillier. 2023. "Characterization of Redox Sensitive Brown Algal Mannitol-1-Phosphatases" Phycology 3, no. 1: 1-12. https://doi.org/10.3390/phycology3010001