
Effect of Growth Hormone Exon-5 Polymorphism on Growth Traits, Body Measurements, Slaughter and Carcass Characteristics, and Meat Quality in Meat-Type Lambs in Turkey

Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Approval
2.2. Animals and Feeding Regimens
2.3. DNA Extraction, Primer Design, PCR Amplification, SSCP, and DNA Sequencing
2.3.1. DNA Extraction
2.3.2. Primer Design, PCR Amplification, and SSCP Analysis
2.3.3. DNA Sequencing
2.4. Live Weight, Linear Body Measurement, and In Vivo Ultrasonographic Measurements
2.5. Slaughter and Carcass Characteristics and Meat Quality Assessment
2.6. Statistical Analysis
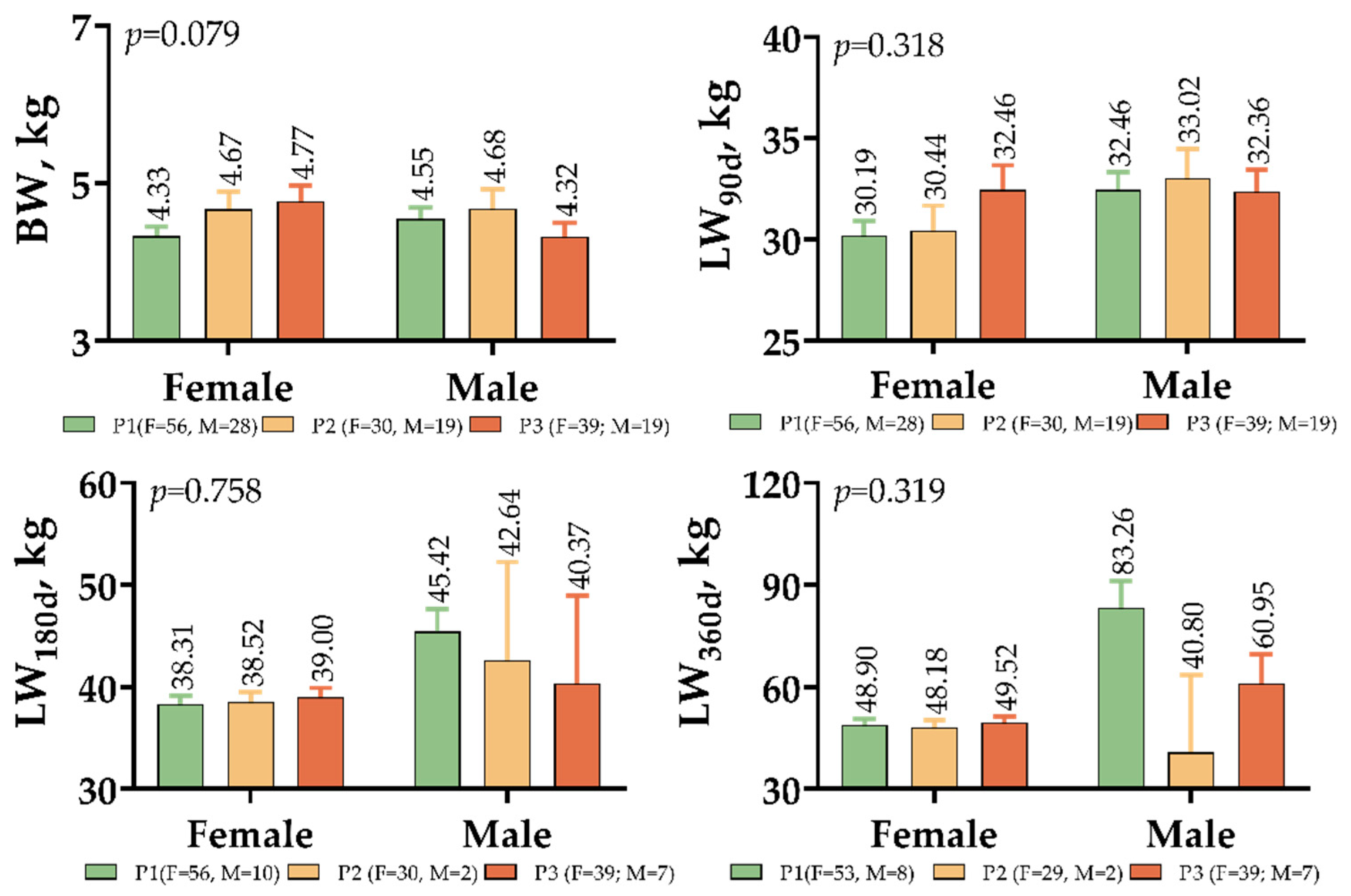
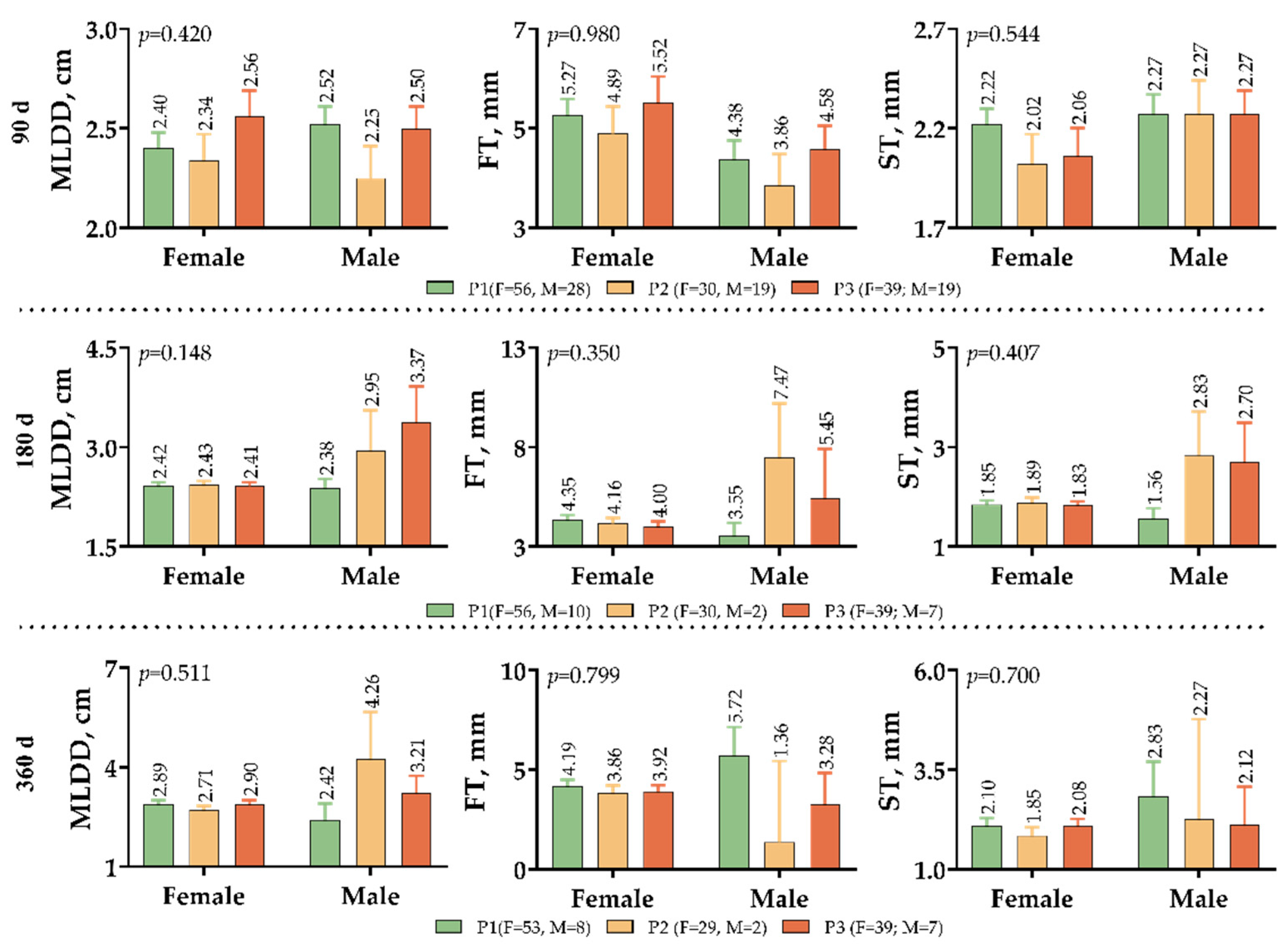
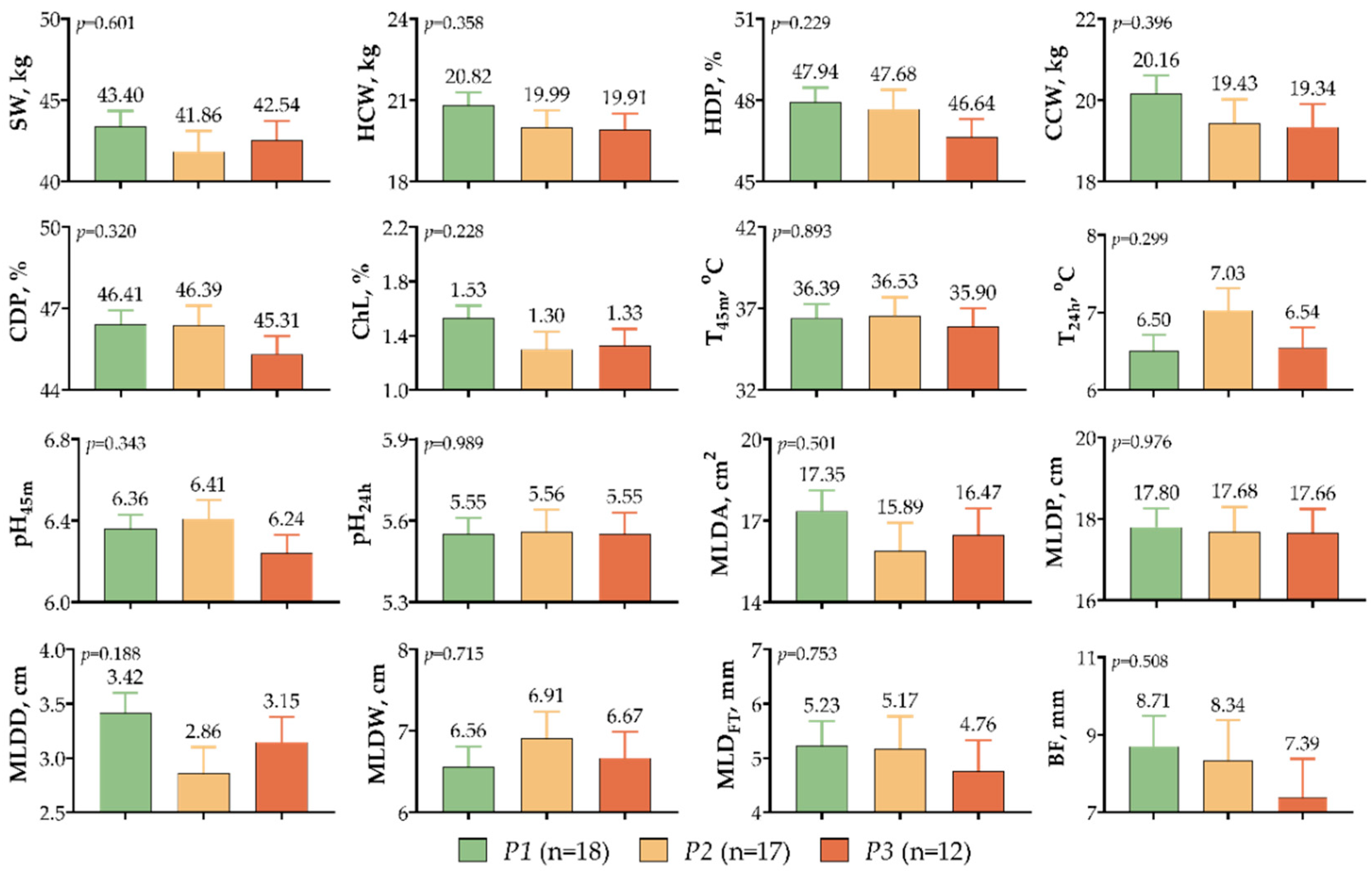
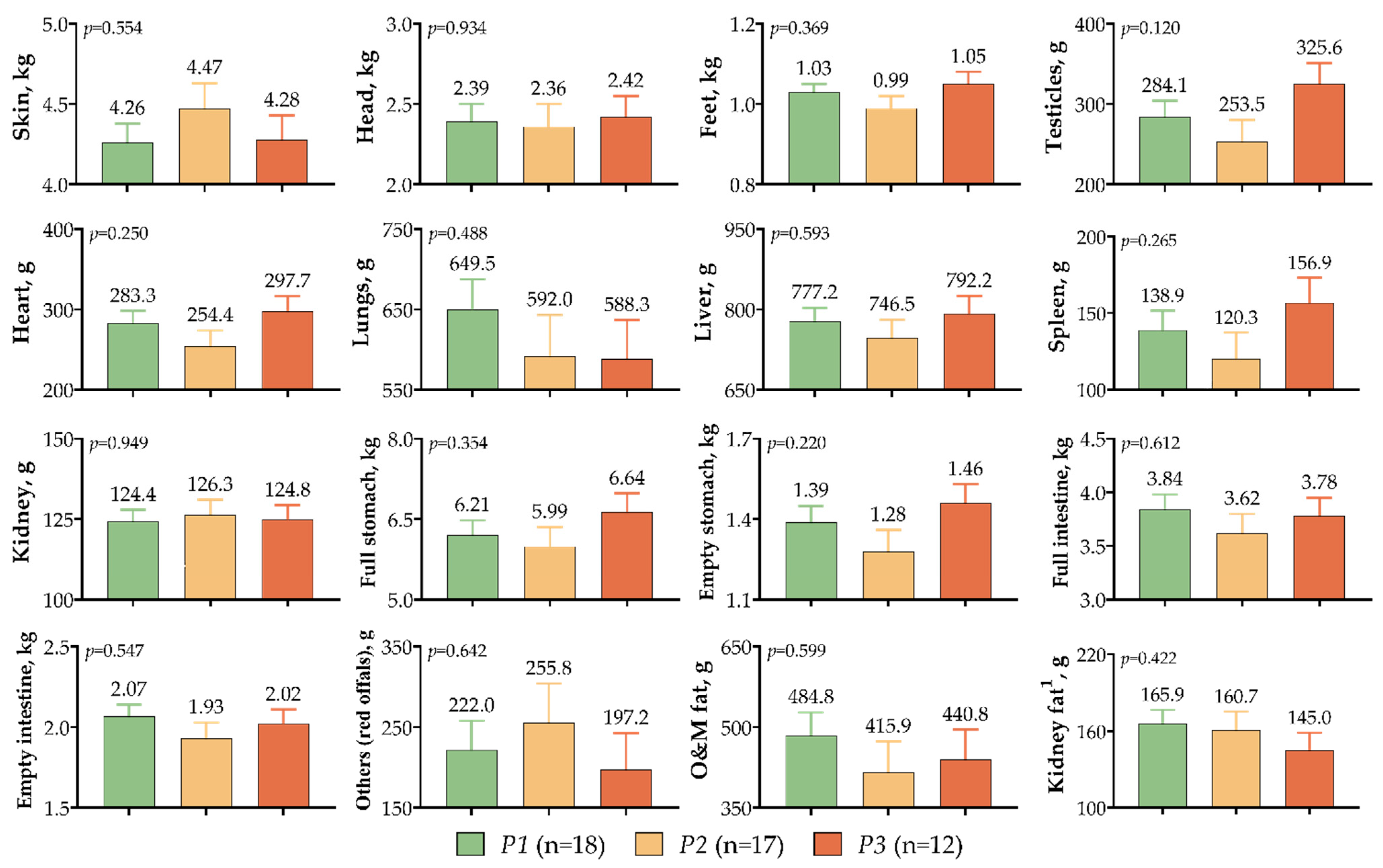
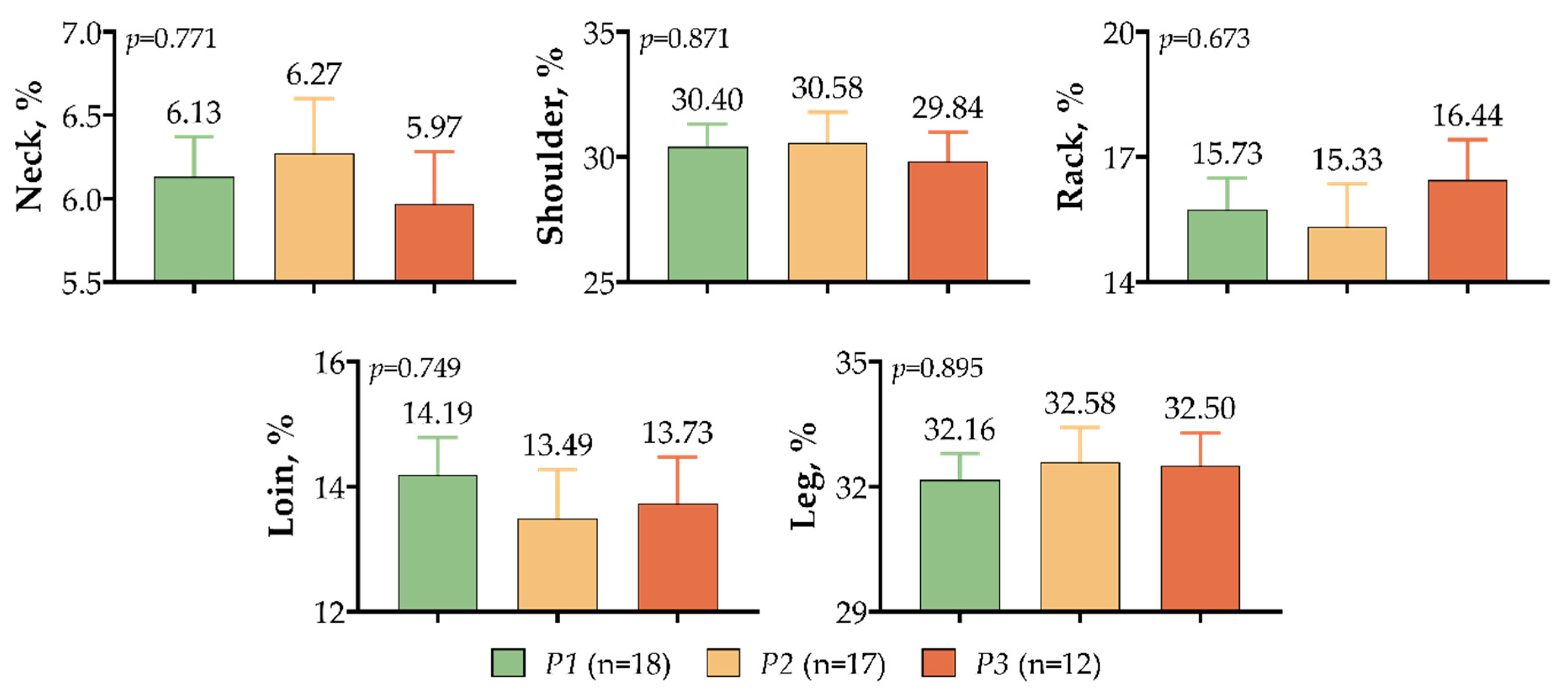
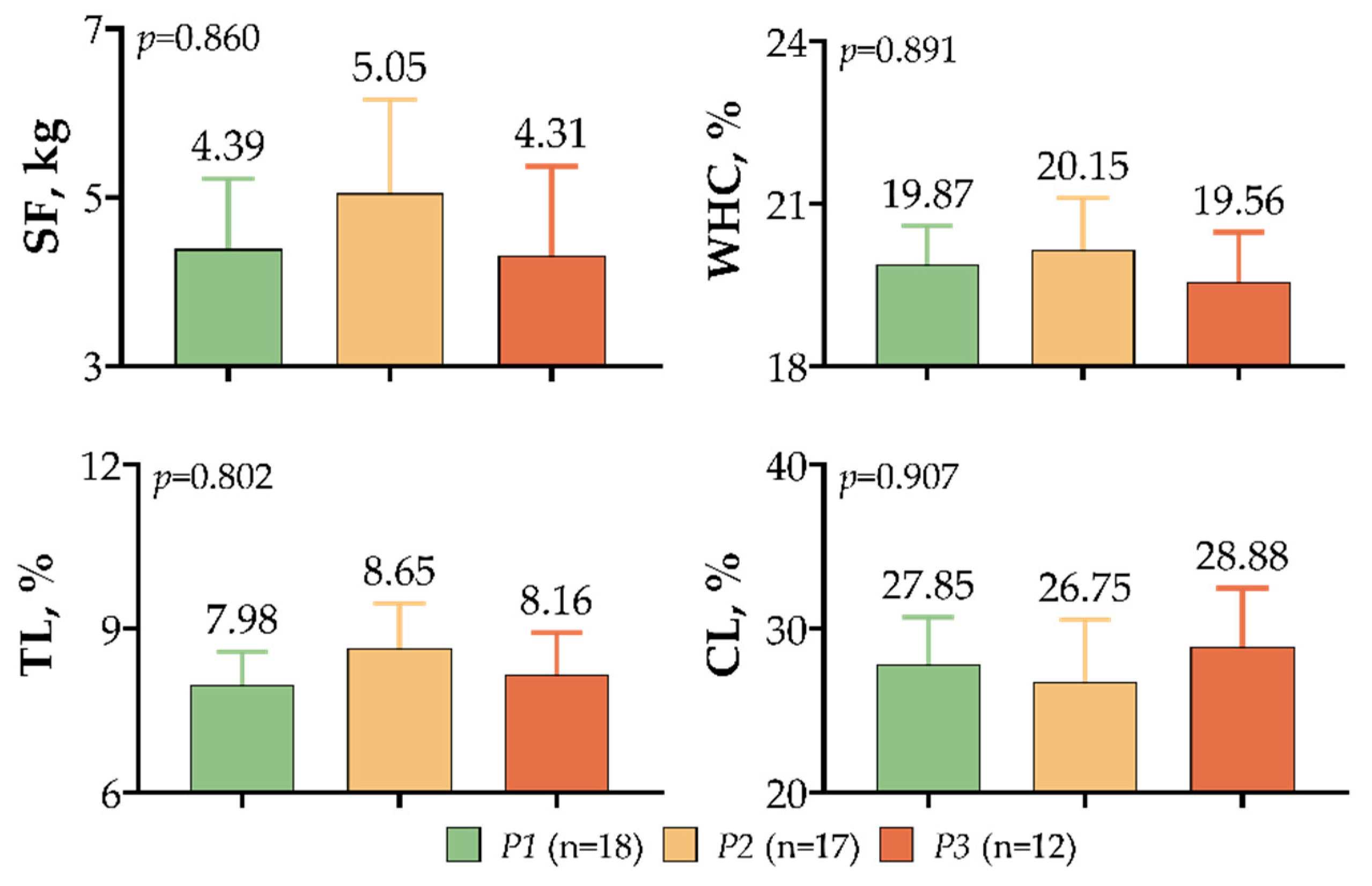
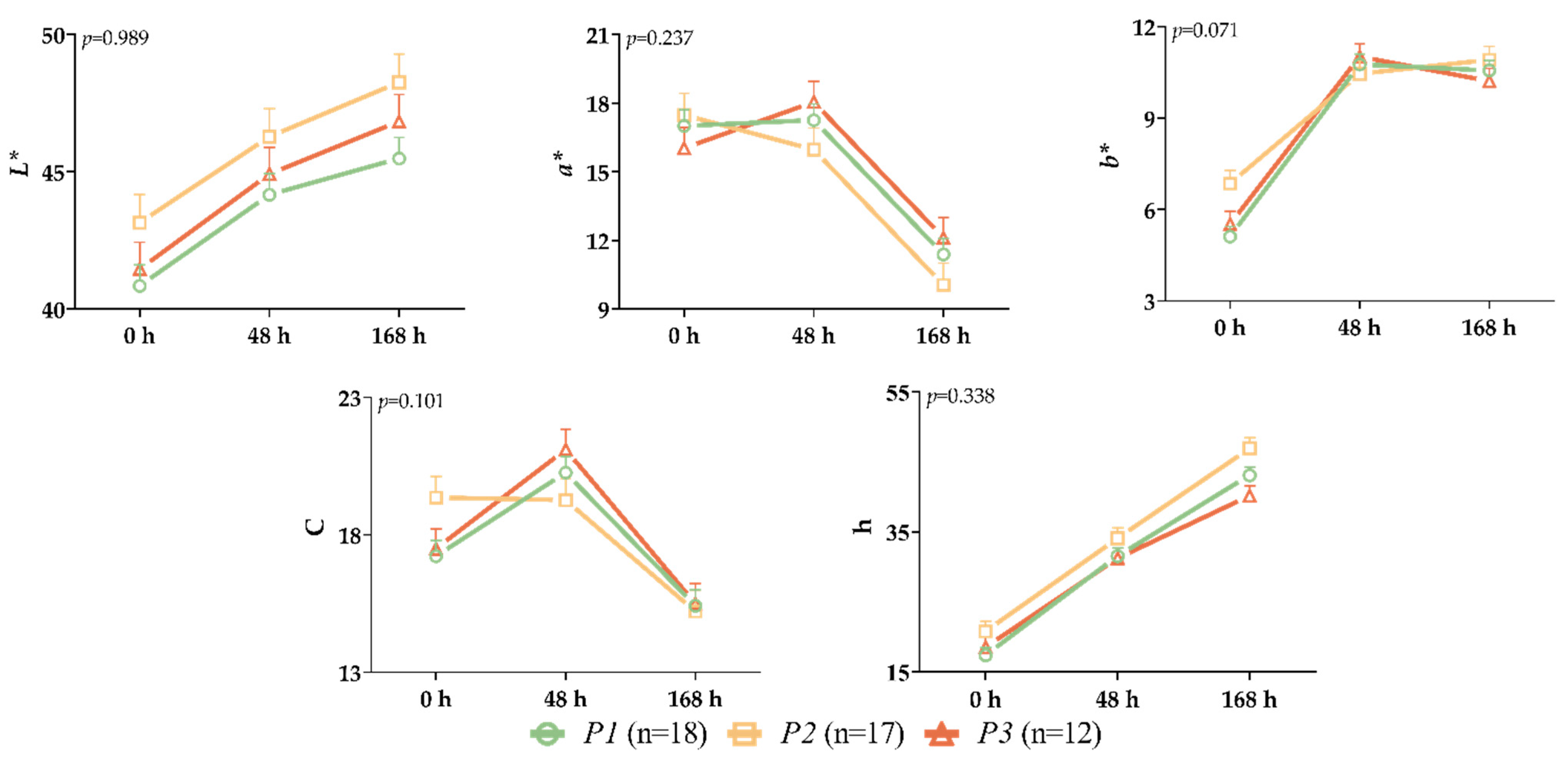
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Flores, D.R.M.; Fonseca, A.F.P.; Schmitt, J.; Tonetto, C.J.; Junior, A.G.R.; Hammerschmitt, R.K.; Facco, D.B.; Brunetto, G.; Nörnberg, J.L. Lambs fed with increasing levels of grape pomace silage: Effects on meat quality. Small Rumin. Res. 2021, 195, 106234. [Google Scholar] [CrossRef]
- Wang, K.; Cui, Y.; Wang, Z.; Yan, H.; Meng, Z.; Zhu, H.; Qu, L.; Lan, X.; Pan, C. One 16 bp insertion/deletion (indel) within the KDM6A gene revealing strong associations with growth traits in goat. Gene 2019, 686, 16–20. [Google Scholar] [CrossRef] [PubMed]
- Erdenee, S.; Akhatayeva, Z.; Pan, C.; Cai, Y.; Xu, H.; Chen, H.; Lan, X. An insertion/deletion within the CREB1 gene identified using the RNA-sequencing is associated with sheep body morphometric traits. Gene 2021, 775, 145444. [Google Scholar] [CrossRef] [PubMed]
- Gorlov, I.F.; Fedunin, A.A.; Randelin, D.A.; Sulimova, G.E. Polymorphisms of bGH, RORC, and DGAT1 genes in Russian beef cattle breeds. Russ. J. Genet. 2014, 50, 1302–1307. [Google Scholar] [CrossRef]
- Gorlov, I.F.; Shirokova, N.V.; Kolosov, Y.A.; Kolosov, A.Y.; Getmantseva, L.V.; Slozhenkina, M.I.; Mosolova, N.I.; Anisimova, E.Y.; Ponomariov, V.V. Polymorphism of CLPG gene in three sheep breeds grown in the steppe zone of the Russian Federation. J. Adv. Vet. Anim. Res. 2020, 7, 51–55. [Google Scholar] [CrossRef]
- Lina, M.; Qing, M.; Yingkang, L.; Zhifen, W. Developmental changes of GH gene in Tan sheep and correlation analysis with slaughter traits. Anim. Husb. Feed Sci. 2017, 9, 209–212. [Google Scholar] [CrossRef]
- Byrne, C.R.; Wilson, B.W.; Ward, K.A. The isolation and characterisation of the ovine growth hormone gene. Aust. J. Biol. Sci. 1987, 40, 459–470. [Google Scholar] [CrossRef] [Green Version]
- Singh, L.V.; Sharma, A.; Kumari, N.; Kaur, N.; Jayakumar, S.; Dixit, S.P.; Gupta, N.; Gupta, S.C. Comparative sequence analysis in the exon 5 of growth hormone gene in the various livestock species of India. Anim. Biotechnol. 2014, 25, 69–72. [Google Scholar] [CrossRef]
- Yao, J.; Aggrey, S.E.; Zadworny, D.; Hayes, J.F.; Kühnlein, U. Sequence variations in the bovine growth hormone gene characterized by single-strand conformation polymorphism (SSCP) analysis and their association with milk production traits in Holsteins. Genetics 1996, 144, 1809–1816. [Google Scholar] [CrossRef]
- Sugita, H.; Ardiyanti, A.; Yokota, S.; Yonekura, S.; Hirayama, T.; Shoji, N.; Yamauchi, E.; Suzuki, K.; Katoh, K.; Roh, S.G. Effect of single nucleotide polymorphisms in GH gene promoter region on carcass traits and intramuscular fatty acid compositions in Japanese Black cattle. Livest. Sci. 2014, 165, 15–21. [Google Scholar] [CrossRef]
- An, X.P.; Hou, J.X.; Wang, L.X.; Li, G.; Wang, J.G.; Song, Y.X.; Zhou, G.Q.; Han, D.; Ling, L.; Cao, B.Y. Novel polymorphisms of the growth hormone gene and their effect on growth traits in Chinese goats. Meat Sci. 2010, 86, 758–763. [Google Scholar] [CrossRef] [PubMed]
- Bahrami, A.; Behzadi, S.; Miraei-Ashtiani, S.R.; Roh, S.; Katoh, K. Genetic polymorphisms and protein structures in growth hormone, growth hormone receptor, ghrelin, insulin-like growth factor 1 and leptin in Mehraban sheep. Gene 2013, 527, 397–404. [Google Scholar] [CrossRef] [PubMed]
- Oğan, M. Karacabey Merinoslarında önemli verim özelliklerini seleksiyonla geliştirme olanakları 1. Çeşitli özellikler bakımından performans düzeyleri. Lalahan Hayvancılık Araştırma Enstitüsü Derg. 1994, 34, 47–58, (In Turkish with an Abstract in English). [Google Scholar]
- Koyuncu, M.; İpek, A.; Tuncel, E.; Akgündüz, V. Some yield characteristics of genotype groups obtained by crossbreeding Kıvırcık with imported mutton sheep breeds (Hampshire Down, Lincoln and Blackhead German). Turk. J. Vet. Anim. Sci. 1999, 23, 423–427, (In Turkish with an Abstract in English). [Google Scholar]
- Kader Esen, S.; Esen, S.; Karadağ, O.; Önenç, A.; Elmaci, C. Slaughter and carcass characteristics of Kıvırcık, Karacabey Merino, Ramlıç, German Black-Head Mutton× Kıvırcık and Hampshire Down× Merino crossbreed lambs reared under intensive conditions. Turk. J. Vet. Anim. Sci. 2020, 44, 1155–1163. [Google Scholar] [CrossRef]
- Gupta, N.; Ahlawat, S.P.S.; Kumar, D.; Gupta, S.C.; Pandey, A.; Malik, G. Single nucleotide polymorphism in growth hormone gene exon-4 and exon-5 using PCR-SSCP in Black Bengal goats–A prolific meat breed of India. Meat Sci. 2007, 76, 658–665. [Google Scholar] [CrossRef]
- An, X.; Wang, L.; Hou, J.; Li, G.; Song, Y.; Wang, J.; Yang, M.; Cui, Y.; Cao, B. Novel polymorphisms of goat growth hormone and growth hormone receptor genes and their effects on growth traits. Mol. Biol. Rep. 2011, 38, 4037–4043. [Google Scholar] [CrossRef]
- Kader Esen, V.; Elmaci, C. The estimation of live weight from body measurements in different meat-type lambs. J. Agric. Sci. 2021, 27, 469–475. [Google Scholar] [CrossRef]
- Kader Esen, S.; Esen, S.; Karadağ, O.; Önenç, A.; Elmaci, C. Genotypic characterization of meat-type lambs expressing the callipyge gene in Turkey: I. Carcass characteristics and retail yield. Turk. J. Vet. Anim. Sci. 2022, 46, 157–164. [Google Scholar] [CrossRef]
- Kader Esen, V.; Esen, S.; Karadag, O.; Elmaci, C. Genotypic characterization of meat-type lambs expressing the callipyge gene in Turkey: II. Effect on body indexes. Small Rumin. Res. 2022, 208, 106633. [Google Scholar] [CrossRef]
- Green, M.R.; Sambrook, J. Molecular Cloning: A Laboratory Manual, 4th ed.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2012. [Google Scholar]
- Byun, S.O.; Fang, Q.; Zhou, H.; Hickford, J.G.H. An effective method for silver-staining DNA in large numbers of polyacrylamide gels. Anal. Biochem. 2009, 385, 174–175. [Google Scholar] [CrossRef] [PubMed]
- Yılmaz, O.; Cemal, I.; Karaca, O. Estimation of mature live weight using some body measurements in Karya sheep. Trop. Anim. Health Prod. 2013, 45, 397–403. [Google Scholar] [CrossRef] [PubMed]
- Esen, S.; Karadag, H.; Kaptan, C.; Eseceli, H. Effect of activated clinoptilolite and inactive brewer’s yeast mixture on loin eye muscle and body indexes in fattening period. Med. Weter. 2020, 76, 626–630. [Google Scholar] [CrossRef]
- Yakan, A.; Ates, C.T.; Alasahan, S.; Odabasioglu, F.; Unal, N.; Ozturk, O.H.; Gungor, O.F.; Ozbeyaz, C. Damascus kids’ slaughter, carcass, and meat quality traits in different production systems using antioxidant supplementation. Small Rumin. Res. 2016, 136, 45–53. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [Green Version]
- Knowles, S.O.; Grace, N.D.; Rounce, J.R.; Realini, C.E. Quality, nutrient and sensory characteristics of aged meat from lambs supplemented with selenomethionine. Food Res. Int. 2020, 137, 109655. [Google Scholar] [CrossRef]
- Honikel, K.O.; Hamm, R. Measurement of water-holding capacity and juiciness. In Quality Attributes and Their Measurement in Meat, Poultry and Fish Products; Pearson, A.M., Dutson, T.R., Eds.; Springer: Boston, MA, USA, 1994; pp. 125–161. [Google Scholar]
- Choi, Y.M.; Kim, B.C. Muscle fiber characteristics, myofibrillar protein isoforms, and meat quality. Livest. Sci. 2009, 122, 105–118. [Google Scholar] [CrossRef]
- Gonzales-Barron, U.; Santos-Rodrigues, G.; Piedra, R.B.; Coelho-Fernandes, S.; Osoro, K.; Celaya, R.; Maurício, R.S.; Pires, J.; Tolsdorf, A.; Geß, A.; et al. Quality attributes of lamb meat from European breeds: Effects of intrinsic properties and storage. Small Rumin. Res. 2021, 198, 106354. [Google Scholar] [CrossRef]
- Minitab, I. Statistical Software for Windows, release 17; Minitab Inc.: State College, PA, USA, 2014. [Google Scholar]
- Sun, H.Z.; Guan, L.L. Genomics for Food Security with Efficient and Sustainable Livestock Production, Comprehensive Foodomics; Elsevier: Amsterdam, The Netherlands, 2020. [Google Scholar] [CrossRef]
- Eggen, A.; Hocquette, J.F. Genomic approaches to economic trait loci and tissue expression profiling: Application to muscle biochemistry and beef quality. Meat Sci. 2004, 66, 1–9. [Google Scholar] [CrossRef]
- Mucha, S.; Mrode, R.; Coffey, M.; Kizilaslan, M.; Desire, S.; Conington, J. Genome-wide association study of conformation and milk yield in mixed-breed dairy goats. J. Dairy Sci. 2018, 101, 2213–2225. [Google Scholar] [CrossRef] [Green Version]
- Moioli, B.; Andrea, M.D.; Pilla, F. Candidate genes affecting sheep and goat milk quality. Small Rumin. Res. 2007, 68, 179–192. [Google Scholar] [CrossRef]
- Bastos, E.; Cravador, A.; Azevedo, J.; Guedes-Pinto, H. Single strand conformation polymorphism (SSCP) detection in six genes in the Portuguese indigenous sheep breed “Churra da Terra Quente”. Biotechnol. Agron. Soc. Environ. 2001, 5, 7–15. [Google Scholar]
- Seevagan, M.; Jeichitra, V.; Rajendran, R.; Tirumurugaan, K.G. Polymorphism of growth hormone (GH) gene in Vembur and Kilakarsal breeds of sheep. Indian J. Small Rumin. 2015, 21, 196–199. [Google Scholar] [CrossRef]
- Gorlov, I.F.; Kolosov, Y.A.; Shirokova, N.V.; Getmantseva, L.V.; Slozhenkina, M.I.; Mosolova, N.I.; Leonova, M.A.; Kolosov, A.Y.; Zlobina, E.Y. Association of the growth hormone gene polymorphism with growth traits in Salsk sheep breed. Small Rumin. Res. 2017, 150, 11–14. [Google Scholar] [CrossRef]
- Hua, G.H.; Chen, S.L.; Yu, J.N.; Cai, K.L.; Wu, C.J.; Li, Q.L.; Zhang, C.Y.; Liang, A.X.; Han, L.; Geng, L.Y.; et al. Polymorphism of the growth hormone gene and its association with growth traits in Boer goat bucks. Meat Sci. 2009, 81, 391–395. [Google Scholar] [CrossRef]
- Jia, J.L.; Zhang, L.P.; Wu, J.P.; Ha, Z.J.; Li, W.W. Study of the correlation between GH gene polymorphism and growth traits in sheep. Genet. Mol. Res. 2014, 13, 7190–7200. [Google Scholar] [CrossRef]
- Moradian, C.; Mohamadi, N.; Razavi-Sheshdeh, S.A.; Hajihosseinlo, A.; Ashrafi, F. Effects of genetic polymorphism at the growth hormone gene on growth traits in Makooei sheep. Eur. J. Exp. Biol. 2013, 3, 101–105. [Google Scholar]
- Abdelmoneim, T.S.; Brooks, P.H.; Afifi, M.; Swelum, A.A.A. Sequencing of growth hormone gene for detection of polymorphisms and their relationship with body weight in Harri sheep. Indian J. Anim. Res. 2017, 51, 205–211. [Google Scholar]
- Al Qasimi, R.H.; Hassan, A.F.; Khudair, B.Y. Effect of IGF-1 and GH genes polymorphism on weights and body measurements of Awassi lambs in different ages. Basrah J. Agric. Sci. 2019, 32, 39–46. [Google Scholar] [CrossRef]
- Machado, A.L.; Meira, A.N.; Jucá, A.D.F.; Azevedo, H.C.; Muniz, E.N.; Coutinho, L.L.; Mourao, G.B.; Pedrosa, V.B.; Pinto, L.F.B. Variants in GH, IGF1, and LEP genes associated with body traits in Santa Inês sheep. Sci. Agric. 2021, 78, e20190216. [Google Scholar] [CrossRef]
- Lee, J.H.; Lee, Y.M.; Lee, J.Y.; Oh, D.Y.; Jeong, D.J.; Kim, J.J. Identification of single nucleotide polymorphisms (SNPs) of the bovine growth hormone (bGH) gene associated with growth and carcass traits in Hanwoo. Asian-Australas. J. Anim. Sci. 2013, 26, 1359–1364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sedykh, T.A.; Gizatullin, R.S.; Dolmatova, I.Y.; Gusev, I.V.; Kalashnikova, L.A. Growth hormone gene polymorphism in relation to beef cattle carcass quality. Russ. Agric. Sci. 2020, 46, 289–294. [Google Scholar] [CrossRef]
- Özay, M. Kıvırcık Irkı Kuzularda Büyüme ve bel Gözü Kası Özellikleri ile Büyüme Hormonu gen Polimorfizmi Arasındaki ilişkiler (Yayın No: 574445). Master’s Thesis, Aydın Adnan Menderes, Aydın, Turkey, 2019. (In Turkish with an Abstract in English). [Google Scholar]
- Devesa, J.; Almengló, C.; Devesa, P. Multiple effects of growth hormone in the body: Is it really the hormone for growth? Clin. Med. Insights Endocrinol. Diabetes 2016, 9, CMED-S38201. [Google Scholar] [CrossRef] [Green Version]
- Meira, A.N.; Montenegro, H.; Coutinho, L.L.; Mourão, G.B.; Azevedo, H.C.; Muniz, E.N.; Machado, A.L.; Pedrosa, V.B.; Pinto, L.F.B. Single nucleotide polymorphisms in the growth hormone and IGF type-1 (IGF1) genes associated with carcass traits in Santa Ines sheep. Animal 2019, 13, 460–468. [Google Scholar] [CrossRef] [PubMed]
- Agung, P.P.; Anwar, S.; Putra, W.P.B.; Zein, M.S.A.; Wulandari, A.S.; Said, S.; Sudiro, A. Association of growth hormone (GH) gene polymorphism with growth and carcass in Sumba Ongole (SO) cattle. J. Indones. Trop. Anim. Agric. 2017, 42, 153–159. [Google Scholar] [CrossRef] [Green Version]
- Akçay, A.; Akyüz, B.; Bayram, D. Determination of the AluI polymorphism effect of bovine growth hormone gene on carcass traits in Zavot cattle with analysis of covariance. Turk. J. Vet. Anim. Sci. 2015, 39, 16–22. [Google Scholar] [CrossRef] [Green Version]
- Han, S.H.; Cho, I.C.; Ko, M.S.; Jeong, H.Y.; Oh, H.S.; Lee, S.S. Effects of POU1F1 and GH1 genotypes on carcass traits in Hanwoo cattle. Genes Genom. 2010, 32, 105–109. [Google Scholar] [CrossRef]
- Pereira, A.P.; Alencar, M.M.D.; Oliveira, H.N.D.; Regitano, L.C.D.A. Association of GH and IGF-1 polymorphisms with growth traits in a synthetic beef cattle breed. Genet. Mol. Biol. 2005, 28, 230–236. [Google Scholar] [CrossRef] [Green Version]
- Costello, S.; O’Doherty, E.; Troy, D.J.; Ernst, C.W.; Kim, K.S.; Stapleton, P.; Sweeney, T.; Mullen, A.M. Association of polymorphisms in the calpain I, calpain II and growth hormone genes with tenderness in bovine M. Longissimus dorsi. Meat Sci. 2007, 75, 551–557. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | DM | CP | CA | EE | CF | NDF | ADF |
|---|---|---|---|---|---|---|---|
| Concentrate feed | 89.44 | 12.65 | 9.43 | 3.22 | 9.66 | 39.90 | 12.95 |
| Alfalfa hay | 87.49 | 23.20 | 14.51 | 2.07 | 20.38 | 35.43 | 23.00 |
| Vetch–wheat mixture hay | 89.49 | 8.71 | 5.66 | 1.26 | 41.86 | 63.59 | 46.68 |
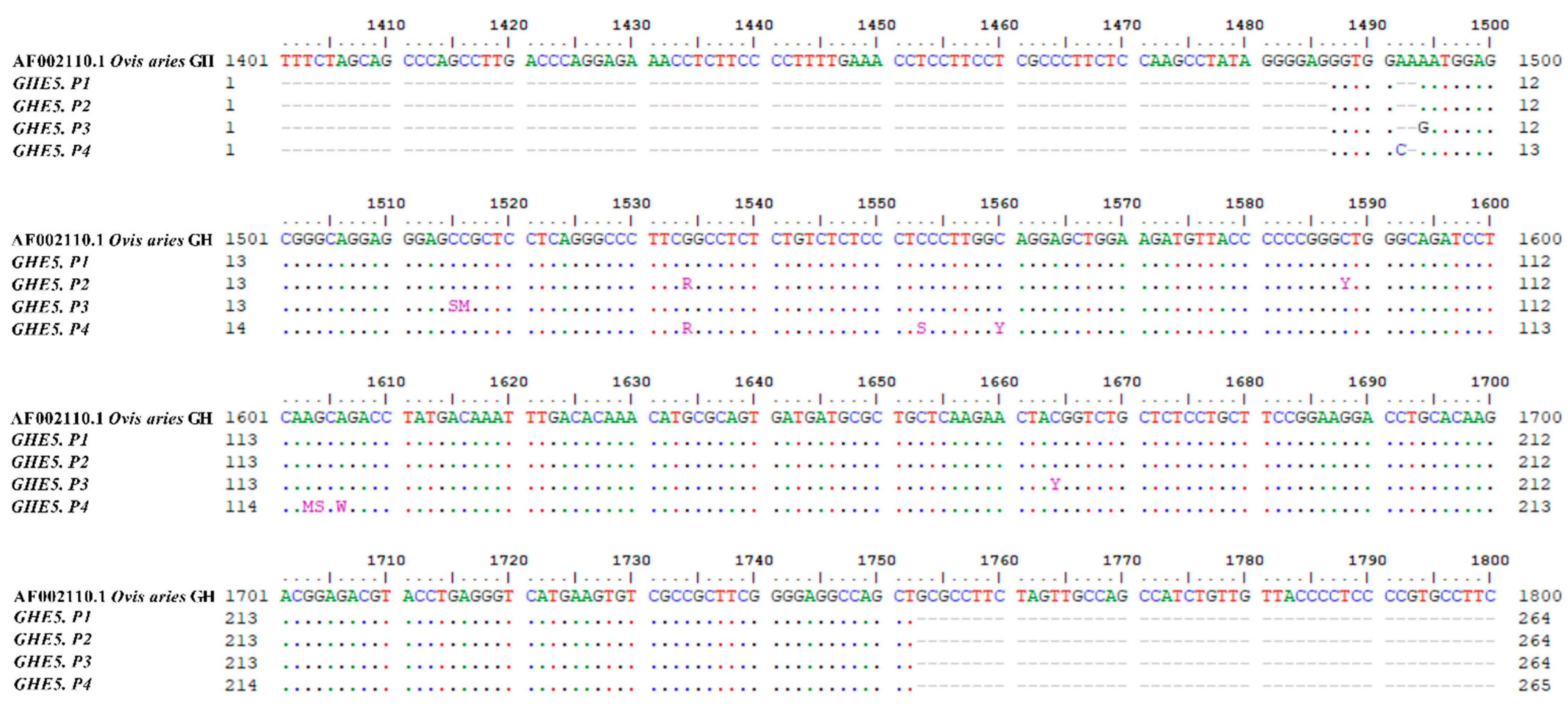
| No | Position 1 | Nucleotide Sequences | Chromosome Location 2 | SNP rs ID | Amino Acid Changing | |||
|---|---|---|---|---|---|---|---|---|
| P1 | P2 | P3 | P4 | |||||
| 1 | c.1563-71 | - | - | - | C | 11:14,855,698 | - | - |
| 2 | c.1563-70 | - | - | - | - | 11:14,855,699 | - | - |
| 3 | c.1563-69 | A | A | G | A | 11:14,855,701 | - | - |
| 4 | c.1563-48 | C | C | S(G/C) | C | 11:14,855,722 | - | - |
| 5 | c.1563-47 | C | C | M(A/C) | C | 11:14,855,723 | - | - |
| 6 | c.1563-29 | G | R(A/G) | G | R(A/G) | 11:14,855,741 | - | - |
| 7 | c.1563-10 | C | C | C | S(G/C) | 11:14,855,760 | - | - |
| 8 | c.1563-3 | C | C | C | Y(C/T) | 11:14,855,767 | - | - |
| 9 | c.1588 | C | Y(C/T) | C | C | 11:14,855,795 | - | p.Ala160Val |
| 10 | c.1603 | A | A | A | M(A/C) | 11:14,855,809 | - | p.Lys165Thr |
| 11 | c.1604 | G | G | G | S(G/C) | 11:14,855,810 | - | p.Lys165Thr |
| 12 | c.1606 | A | A | A | W(A/T) | 11:14,855,812 | - | p.Gln166Leu |
| 13 | c.1664 | C | C | Y(C/T) | C | 11:14,855,871 | rs.596456087 | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Esen, V.K.; Elmacı, C. Effect of Growth Hormone Exon-5 Polymorphism on Growth Traits, Body Measurements, Slaughter and Carcass Characteristics, and Meat Quality in Meat-Type Lambs in Turkey. Ruminants 2022, 2, 420-434. https://doi.org/10.3390/ruminants2040029
Esen VK, Elmacı C. Effect of Growth Hormone Exon-5 Polymorphism on Growth Traits, Body Measurements, Slaughter and Carcass Characteristics, and Meat Quality in Meat-Type Lambs in Turkey. Ruminants. 2022; 2(4):420-434. https://doi.org/10.3390/ruminants2040029
Chicago/Turabian StyleEsen, Vasfiye Kader, and Cengiz Elmacı. 2022. "Effect of Growth Hormone Exon-5 Polymorphism on Growth Traits, Body Measurements, Slaughter and Carcass Characteristics, and Meat Quality in Meat-Type Lambs in Turkey" Ruminants 2, no. 4: 420-434. https://doi.org/10.3390/ruminants2040029