Higher Frequencies of T-Cells Expressing NK-Cell Markers and Chemokine Receptors in Parkinson’s Disease

 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cohorts
2.2. Surface Marker Staining of PBMCs
2.3. Statistics
3. Results
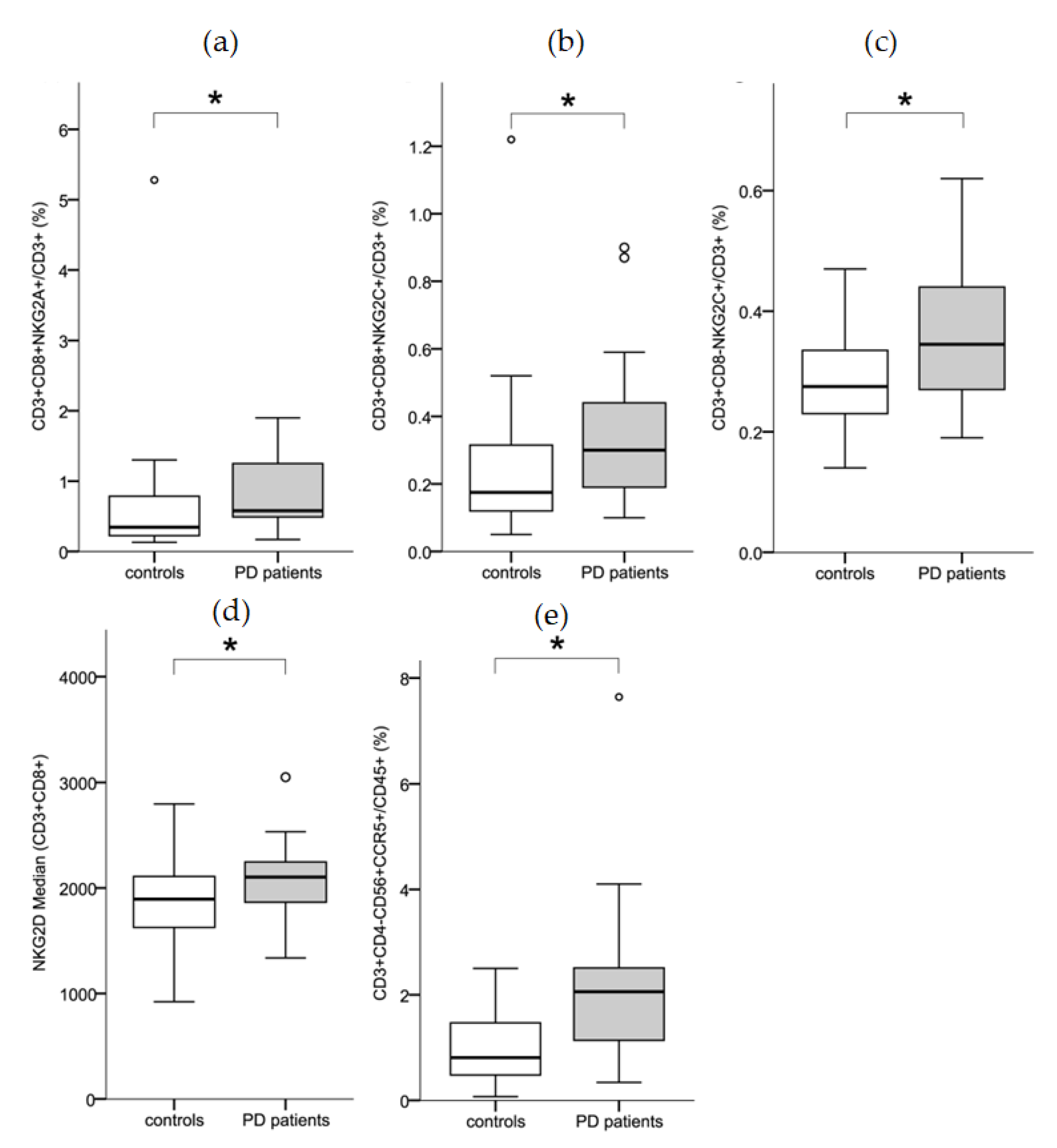
3.1. T-Cell Activation Status
3.2. Chemokine Receptor Expression
3.3. NK-Cell Differentiation States
4. Discussion
4.1. NK-Cell Receptor-Expressing T-Cells in PD Patients vs. Controls
4.2. Chemokine Receptor Expression on T-Cells
4.3. Observations on NK-Cells
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Adriaensen, W.; Matheï, C.; Vaes, B.; van Pottelbergh, G.; Wallemacq, P.; Degryse, J.-M. Interleukin-6 as a first-rated serum inflammatory marker to predict mortality and hospitalization in the oldest old: A regression and CART approach in the BELFRAIL study. Exp. Gerontol. 2015, 69, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Adriaensen, W.; Mathei, C.; Van Pottelbergh, G.; Vaes, B.; Legrand, D.; Wallemacq, P.; Degryse, J.-M. Significance of serum immune markers in identification of global functional impairment in the oldest old: Cross-sectional results from the BELFRAIL study. AGE 2014, 36, 457–467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruunsgaard, H. Effects of tumor necrosis factor-alpha and interleukin-6 in elderly populations. Eur. Cytokine Netw. 2002, 13, 389–391. [Google Scholar] [PubMed]
- Ferrari, C.C.; Tarelli, R. Parkinson’s Disease and Systemic Inflammation. Park. Dis. 2011, 2011, 436813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kempuraj, D.; Thangavel, R.; Natteru, P.A.; Selvakumar, G.P.; Saeed, D.; Zahoor, H.; Zaheer, S.; Iyer, S.S.; Zaheer, A. Neuroinflammation Induces Neurodegeneration. J. Neurol. Neurosurg. Spine 2016, 1, 1003. [Google Scholar]
- Kortekaas, R.; Leenders, K.L.; Van Oostrom, J.C.H.; Vaalburg, W.; Bart, J.; Willemsen, A.T.M.; Hendrikse, N.H. Blood-brain barrier dysfunction in parkinsonian midbrain in vivo. Ann. Neurol. 2005, 57, 176–179. [Google Scholar] [CrossRef]
- Mogi, M.; Harada, M.; Kondo, T.; Riederer, P.; Inagaki, H.; Minami, M.; Nagatsu, T. Interleukin-1 beta, interleukin-6, epidermal growth factor and transforming growth factor-alpha are elevated in the brain from parkinsonian patients. Neurosci. Lett. 1994, 180, 147–150. [Google Scholar] [CrossRef]
- Blum-Degena, D.; Müller, T.; Kuhn, W.; Gerlach, M.; Przuntek, H.; Riederer, P. Interleukin-1β and interleukin-6 are elevated in the cerebrospinal fluid of Alzheimer’s and de novo Parkinson’s disease patients. Neurosci. Lett. 1995, 202, 17–20. [Google Scholar] [CrossRef]
- Muller, T.; Blum-Degen, D.; Przuntek, H.; Kuhn, W. Interleukin-6 levels in cerebrospinal fluid inversely correlate to severity of Parkinson’s disease. Acta Neurol. Scand. 1998, 98, 142–144. [Google Scholar] [CrossRef]
- Dobbs, R.J.; Charlett, A.; Purkiss, A.G.; Dobbs, S.M.; Weller, C.; Peterson, D.W. Association of circulating TNF-α and IL-6 with ageing and parkinsonism. Acta Neurol. Scand. 1999, 100, 34–41. [Google Scholar] [CrossRef]
- Dufek, M.; Hamanová, M.; Lokaj, J.; Goldemund, D.; Rektorová, I.; Michálková, Z.; Sheardová, K.; Rektor, I. Serum inflammatory biomarkers in Parkinson’s disease. Park. Relat. Disord. 2009, 15, 318–320. [Google Scholar] [CrossRef] [PubMed]
- Mihara, T.; Nakashima, M.; Kuroiwa, A.; Akitake, Y.; Ono, K.; Hosokawa, M.; Yamada, T.; Takahashi, M. Natural killer cells of Parkinson’s disease patients are set up for activation: A possible role for innate immunity in the pathogenesis of this disease. Parkinsonism Relat. Disord. 2008, 14, 46–51. [Google Scholar] [CrossRef] [PubMed]
- Niwa, F.; Kuriyama, N.; Nakagawa, M.; Imanishi, J. Effects of peripheral lymphocyte subpopulations and the clinical correlation with Parkinson’s disease. Geriatr. Gerontol. Int. 2012, 12, 102–107. [Google Scholar] [CrossRef] [PubMed]
- Earls, R.H.; Menees, K.B.; Chung, J.; Gutekunst, C.-A.; Lee, H.J.; Hazim, M.G.; Rada, B.; Wood, L.B.; Lee, J.-K. NK cells clear α-synuclein and the depletion of NK cells exacerbates synuclein pathology in a mouse model of α-synucleinopathy. Proc. Natl. Acad. Sci. USA 2020, 117, 1762–1771. [Google Scholar] [CrossRef]
- Earls, R.; Menees, K.B.; Chung, J.; Barber, J.; Gutekunst, C.-A.; Hazim, M.G.; Lee, J.-K. Intrastriatal injection of preformed alpha-synuclein fibrils alters central and peripheral immune cell profiles in non-transgenic mice. J. Neuroinflamm. 2019, 16, 250. [Google Scholar] [CrossRef] [Green Version]
- Earls, R.H.; Lee, J.-K. The role of natural killer cells in Parkinson’s disease. Exp. Mol. Med. 2020, 52, 1517–1525. [Google Scholar] [CrossRef]
- Freishtat, R.J.; Mitchell, L.W.; Ghimbovschi, S.D.; Meyers, S.B.; Hoffman, E.P. NKG2A and CD56 Are Coexpressed on Activated TH2 but Not TH1 Lymphocytes. Hum. Immunol. 2005, 66, 1223–1234. [Google Scholar] [CrossRef] [Green Version]
- Graham, C.M.; Christensen, J.R.; Thomas, D.B. Differential induction of CD94 and NKG2 in CD4 helper T cells. A consequence of influenza virus infection and interferon-gamma? Immunology 2007, 121, 238–247. [Google Scholar] [CrossRef]
- Cush, S.S.; Flaño, E. KLRG1+NKG2A+ CD8 T Cells Mediate Protection and Participate in Memory Responses during γ-Herpesvirus Infection. J. Immunol. 2011, 186, 4051–4058. [Google Scholar] [CrossRef] [Green Version]
- Piet, B.; De Bree, G.J.; Smids-Dierdorp, B.S.; Van Der Loos, C.M.; Remmerswaal, E.; von der Thusen, J.; Van Haarst, J.M.W.; Eerenberg, J.P.; Brinke, A.T.; Van Der Bij, W.; et al. CD8+ T cells with an intraepithelial phenotype upregulate cytotoxic function upon influenza infection in human lung. J. Clin. Investig. 2011, 121, 2254–2263. [Google Scholar] [CrossRef] [Green Version]
- Zaguia, F.; Saikali, P.; Ludwin, S.; Newcombe, J.; Beauseigle, D.; McCrea, E.; Duquette, P.; Prat, A.; Antel, J.P.; Arbour, N. Cytotoxic NKG2C+ CD4 T Cells Target Oligodendrocytes in Multiple Sclerosis. J. Immunol. 2013, 190, 2510–2518. [Google Scholar] [CrossRef] [PubMed]
- Björkander, S.; Hell, L.; Johansson, M.A.; Forsberg, M.M.; Lasaviciute, G.; Roos, S.; Holmlund, U.; Sverremark-Ekström, E. Staphylococcus aureus-derived factors induce IL-10, IFN-γ and IL-17A-expressing FOXP3+CD161+ T-helper cells in a partly monocyte-dependent manner. Sci. Rep. 2016, 6, 22083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pesenacker, A.M.; Bending, D.; Ursu, S.; Wu, Q.; Nistala, K.; Wedderburn, L. CD161 defines the subset of FoxP3+ T cells capable of producing proinflammatory cytokines. Blood 2013, 121, 2647–2658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schirmer, L.; Rothhammer, V.; Hemmer, B.; Korn, T. Enriched CD161highCCR6+γδ T Cells in the Cerebrospinal Fluid of Patients With Multiple Sclerosis. JAMA Neurol. 2013, 70, 345–351. [Google Scholar] [CrossRef] [Green Version]
- Kouli, A.; Jensen, M.; Papastavrou, V.; Scott, K.M.; Kolenda, C.; Parker, C.; Solim, I.H.; Camacho, M.; Martin-Ruiz, C.; Williams-Gray, C.H. T lymphocyte senescence is attenuated in Parkinson’s disease. J. Neuroinflamm. 2021, 18, 228. [Google Scholar] [CrossRef]
- Billerbeck, E.; Kang, Y.-H.; Walker, L.; Lockstone, H.; Grafmueller, S.; Fleming, V.; Flint, J.; Willberg, C.B.; Bengsch, B.; Seigel, B.; et al. Analysis of CD161 expression on human CD8+ T cells defines a distinct functional subset with tissue-homing properties. Proc. Natl. Acad. Sci. USA 2010, 107, 3006–3011. [Google Scholar] [CrossRef] [Green Version]
- Sallusto, F.; Impellizzieri, D.; Basso, C.; Laroni, A.; Uccelli, A.; Lanzavecchia, A.; Engelhardt, B. T-cell trafficking in the central nervous system. Immunol. Rev. 2012, 248, 216–227. [Google Scholar] [CrossRef]
- Reboldi, A.; Coisne, C.; Baumjohann, D.; Benvenuto, F.; Bottinelli, D.; Lira, S.A.; Uccelli, A.; Lanzavecchia, A.; Engelhardt, B.; Sallusto, F. C-C chemokine receptor 6-regulated entry of TH-17 cells into the CNS through the choroid plexus is required for the initiation of EAE. Nat. Immunol. 2009, 10, 514–523. [Google Scholar] [CrossRef]
- Yan, Z.; Yang, W.; Wei, H.; Dean, M.N.; Standaert, D.G.; Cutter, G.R.; Benveniste, E.N.; Qin, H. Dysregulation of the Adaptive Immune System in Patients With Early-Stage Parkinson Disease. Neurol. Neuroimmunol. Neuroinflamm. 2021, 8, e1036. [Google Scholar] [CrossRef]
- Subramanian, S.; Ayala, P.; Wadsworth, T.L.; Harris, C.J.; Vandenbark, A.A.; Quinn, J.F.; Offner, H. CCR6: A Biomarker for Alzheimer’s-like Disease in a Triple Transgenic Mouse Model. J. Alzheimer’s Dis. 2010, 22, 619–629. [Google Scholar] [CrossRef] [Green Version]
- Bulati, M.; Buffa, S.; Martorana, A.; Gervasi, F.; Camarda, C.; Azzarello, D.M.; Monastero, R.; Caruso, C.; Colonna-Romano, G. Double Negative (IgG+IgD−CD27−) B Cells are Increased in a Cohort of Moderate-Severe Alzheimer’s Disease Patients and Show a Pro-Inflammatory Trafficking Receptor Phenotype. J. Alzheimer’s Dis. 2015, 44, 1241–1251. [Google Scholar] [CrossRef] [PubMed]
- Goldeck, D.; Larbi, A.; Pellicanó, M.; Alam, I.; Zerr, I.; Schmidt, C.; Fulop, T.; Pawelec, G. Enhanced Chemokine Receptor Expression on Leukocytes of Patients with Alzheimer’s Disease. PLoS ONE 2013, 8, e66664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Man, S.-M.; Ma, Y.-R.; Shang, D.-S.; Zhao, W.-D.; Li, B.; Guo, D.-W.; Fang, W.-G.; Zhu, L.; Chen, Y.-H. Peripheral T cells overexpress MIP-1α to enhance its transendothelial migration in Alzheimer’s disease. Neurobiol. Aging 2007, 28, 485–496. [Google Scholar] [CrossRef] [PubMed]
- Ubogu, E.E.; Callahan, M.K.; Tucky, B.H.; Ransohoff, R.M. CCR5 expression on monocytes and T cells: Modulation by transmigration across the blood–brain barrier in vitro. Cell. Immunol. 2006, 243, 19–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.; Shang, D.-S.; Zhao, W.-D.; Tian, L.; Li, B.; Fang, W.-G.; Zhu, L.; Man, S.-M.; Chen, Y.-H. Amyloid β Interaction with Receptor for Advanced Glycation End Products Up-Regulates Brain Endothelial CCR5 Expression and Promotes T Cells Crossing the Blood-Brain Barrier. J. Immunol. 2009, 182, 5778–5788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, C.J.; Park, M.H.; Hwang, J.Y.; Kim, J.H.; Yun, N.Y.; Oh, S.Y.; Song, J.K.; Seo, H.O.; Kim, Y.-B.; Oh, K.-W.; et al. CCR5 deficiency accelerates lipopolysaccharide-induced astrogliosis, amyloid-beta deposit and impaired memory function. Oncotarget 2016, 7, 11984–11999. [Google Scholar] [CrossRef] [Green Version]
- Van De Berg, P.J.E.J.; Yong, S.-L.; Remmerswaal, E.; van Lier, R.; Berge, I.J.M.T. Cytomegalovirus-Induced Effector T Cells Cause Endothelial Cell Damage. Clin. Vaccine Immunol. 2012, 19, 772–779. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Cui, G.; Zhu, M.; Kang, X.; Guo, H. Neuroinflammation in Alzheimer’s disease: Chemokines produced by astrocytes and chemokine receptors. Int. J. Clin. Exp. Pathol. 2014, 7, 8342–8355. [Google Scholar]
- Reale, M.; Iarlori, C.; Thomas, A.; Gambi, D.; Perfetti, B.; Di Nicola, M.; Onofrj, M. Peripheral cytokines profile in Parkinson’s disease. Brain Behav. Immun. 2009, 23, 55–63. [Google Scholar] [CrossRef]
- Galiano-Landeira, J.; Torra, A.; Vila, M.; Bové, J. CD8 T cell nigral infiltration precedes synucleinopathy in early stages of Parkinson’s disease. Brain 2020, 143, 3717–3733. [Google Scholar] [CrossRef]
- Suvas, S.; Azkur, A.K.; Rouse, B.T. Qa-1b and CD94-NKG2a interaction regulate cytolytic activity of herpes simplex virus-specific memory CD8+ T cells in the latently infected trigeminal ganglia. J. Immunol. 2006, 176, 1703–1711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gumá, M.; Busch, L.K.; Salazar-Fontana, L.I.; Bellosillo, B.; Morte, C.; García, P.; López-Botet, M. The CD94/NKG2C killer lectin-like receptor constitutes an alternative activation pathway for a subset of CD8+ T cells. Eur. J. Immunol. 2005, 35, 2071–2080. [Google Scholar] [CrossRef] [PubMed]
- van Bijnen, S.T.; Withaar, M.; Preijers, F.; van der Meer, A.; de Witte, T.; Muus, P.; Dolstra, H. T cells expressing the activating NK-cell receptors KIR2DS4, NKG2C and NKG2D are elevated in paroxysmal nocturnal hemoglobinuria and cytotoxic toward hematopoietic progenitor cell lines. Exp. Hematol. 2011, 39, 751–762.e3. [Google Scholar] [CrossRef] [PubMed]
- Appel, S.H. CD4+ T cells mediate cytotoxicity in neurodegenerative diseases. J. Clin. Investig. 2009, 119, 13–15. [Google Scholar] [CrossRef] [PubMed]
- Groh, V.; Brühl, A.; El-Gabalawy, H.; Nelson, J.L.; Spies, T. Stimulation of T cell autoreactivity by anomalous expression of NKG2D and its MIC ligands in rheumatoid arthritis. Proc. Natl. Acad. Sci. USA 2003, 100, 9452–9457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberts, T.K.; Eugenin, E.A.; Lopez, L.; Romero, I.A.; Weksler, B.B.; Couraud, P.-O.; Berman, J.W. CCL2 disrupts the adherens junction: Implications for neuroinflammation. Lab. Investig. 2012, 92, 1213–1233. [Google Scholar] [CrossRef] [Green Version]
- Zecca, L.; Wilms, H.; Geick, S.; Claasen, J.-H.; Brandenburg, L.-O.; Holzknecht, C.; Panizza, M.L.; Zucca, F.A.; Deuschl, G.; Sievers, J.; et al. Human neuromelanin induces neuroinflammation and neurodegeneration in the rat substantia nigra: Implications for Parkinson’s disease. Acta Neuropathol. 2008, 116, 47–55. [Google Scholar] [CrossRef]
- Rentzos, M.; Nikolaou, C.; Andreadou, E.; Paraskevas, G.P.; Rombos, A.; Zoga, M.; Tsoutsou, A.; Boufidou, F.; Kapaki, E.; Vassilopoulos, D. Circulating interleukin-15 and RANTES chemokine in Parkinson’s disease. Acta Neurol. Scand. 2007, 116, 374–379. [Google Scholar] [CrossRef]
- Gangemi, S.; Basile, G.; Merendino, R.A.; Epifanio, A.; Di Pasquale, G.; Ferlazzo, B.; Nicita-Mauro, V.; Morgante, L. Effect of levodopa on interleukin-15 and RANTES circulating levels in patients affected by Parkinson’s disease. Mediat. Inflamm. 2003, 12, 251–253. [Google Scholar] [CrossRef] [Green Version]
- Tang, P.; Chong, L.; Li, X.; Liu, Y.; Liu, P.; Hou, C.; Li, R. Correlation between Serum RANTES Levels and the Severity of Parkinson’s Disease. Oxidative Med. Cell. Longev. 2014, 2014, 208408. [Google Scholar] [CrossRef] [Green Version]
- Reale, M.; Greig, N.H.; Kamal, M.A. Peripheral chemo-cytokine profiles in Alzheimer’s and Parkinson’s diseases. Mini Rev. Med. Chem. 2009, 9, 1229–1241. [Google Scholar] [CrossRef] [PubMed]
- Reale, M.; Iarlori, C.; Feliciani, C.; Gambi, D. Peripheral chemokine receptors, their ligands, cytokines and Alzheimer’s disease. J. Alzheimer’s Dis. 2008, 14, 147–159. [Google Scholar] [CrossRef] [PubMed]
- Dhanwani, R.; Lima-Junior, J.R.; Sethi, A.; Pham, J.; Williams, G.; Frazier, A.; Xu, Y.; Amara, A.W.; Standaert, D.G.; Goldman, J.G.; et al. Transcriptional analysis of peripheral memory T cells reveals Parkinson’s disease-specific gene signatures. npj Park. Dis. 2022, 8, 30. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Huang, Y.; Bao, T.; Liu, C.; Liu, X.; Chen, X. The role of Th17 cells/IL-17A in AD, PD, ALS and the strategic therapy targeting on IL-17A. J. Neuroinflamm. 2022, 19, 98. [Google Scholar] [CrossRef]
- Cao, J.-J.; Li, K.-S.; Shen, Y.-Q. Activated Immune Cells in Parkinson’s Disease. J. Neuroimmune Pharmacol. 2011, 6, 323–329. [Google Scholar] [CrossRef]
- Cardona, S.M.; Garcia, J.A.; Cardona, A.E. The Fine Balance of Chemokines During Disease: Trafficking, Inflammation, and Homeostasis. Methods Mol. Biol. 2013, 1013, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Sallusto, F.; Lenig, D.; Mackay, C.; Lanzavecchia, A. Flexible Programs of Chemokine Receptor Expression on Human Polarized T Helper 1 and 2 Lymphocytes. J. Exp. Med. 1998, 187, 875–883. [Google Scholar] [CrossRef]
- Martin-Blondel, G.; Brassat, D.; Bauer, J.; Lassmann, H.; Liblau, R. CCR5 blockade for neuroinflammatory diseases—Beyond control of HIV. Nat. Rev. Neurol. 2016, 12, 95–105. [Google Scholar] [CrossRef]
- Choi, D.-Y.; Lee, M.K.; Hong, J.T. Lack of CCR5 modifies glial phenotypes and population of the nigral dopaminergic neurons, but not MPTP-induced dopaminergic neurodegeneration. Neurobiol. Dis. 2013, 49, 159–168. [Google Scholar] [CrossRef]
- Nissen, S.K.; Farmen, K.; Carstensen, M.; Schulte, C.; Goldeck, D.; Brockmann, K.; Romero-Ramos, M. Changes in CD163+, CD11b+, and CCR2+ peripheral monocytes relate to Parkinson’s disease and cognition. Brain Behav. Immun. 2022, 101, 182–193. [Google Scholar] [CrossRef]
- Cen, L.; Yang, C.; Huang, S.; Zhou, M.; Tang, X.; Li, K.; Guo, W.; Wu, Z.; Mo, M.; Xiao, Y.; et al. Peripheral Lymphocyte Subsets as a Marker of Parkinson’s Disease in a Chinese Population. Neurosci. Bull. 2017, 33, 493–500. [Google Scholar] [CrossRef] [PubMed]
- Zhu, B.; Yin, D.; Zhao, H.; Zhang, L. The immunology of Parkinson’s disease. Semin. Immunopathol. 2022, 44, 659–672. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.; Dai, S.-B.; Jiang, S.-S.; Yang, W.-Y.; Yan, Y.-Q.; Lin, Z.-H.; Dong, J.-X.; Liu, Y.; Zheng, R.; Chen, Y.; et al. Specific immune status in Parkinson’s disease at different ages of onset. NPJ Park. Dis. 2022, 8, 5. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Markers | Cells | PD Patients % (Median (Range)) | Control % (Median (Range)) | p Values 1 |
|---|---|---|---|---|
| CD3-CD56+ 2 | Total NK-cells | 5.27 (0.51–11.52) | 5.42 (2.25–14.54) | 0.77 |
| CD3-CD56highCD16low 2 | Immature NK-cells | 0.30 (0.06–0.70) | 0.24 (0.09–0.98) | 0.52 |
| CD3-CD56lowCD16high 2 | Mature NK-cells | 4.72 (0.33–10.68) | 4.93 (1.55–13.92) | 0.67 |
| CD3-CD56lowCD16high CD57+ 2 | Late stage mature NK-cells | 2.44 (0.27–5.77) | 3.00 (0.71–11.15) | 0.20 |
| CD3-CD56lowCD16high CD161+ 2 | CD161+ mature NK-cells | 3.89 (0.28–9.01) | 4.53 (1.19–12.51) | 0.57 |
| CD3-CD56lowCD16high CCR5+ 3 | CCR5 expressing mature NK-cells | 0.10 (0.04–1.00) | 0.08 (0.02–0.58) | 0.44 |
| CD3-CD56lowCD16high CCR5 median 4 | CCR5 expressing mature NK-cells | 899 (232–543) | 347 (202–541) | 0.37 |
| CD3-CD56lowCD16high CXCR2 median 4 | CXCR2 expressing mature NK-cells | 1008 (717–1585) | 382 (538–1998) | 0.15 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Goldeck, D.; Schulte, C.; Teixeira dos Santos, M.C.; Scheller, D.; Öttinger, L.; Pawelec, G.; Deuschle, C.; Berg, D.; Nogueira da Costa, A.; Maetzler, W. Higher Frequencies of T-Cells Expressing NK-Cell Markers and Chemokine Receptors in Parkinson’s Disease. J. Ageing Longev. 2023, 3, 1-10. https://doi.org/10.3390/jal3010001
Goldeck D, Schulte C, Teixeira dos Santos MC, Scheller D, Öttinger L, Pawelec G, Deuschle C, Berg D, Nogueira da Costa A, Maetzler W. Higher Frequencies of T-Cells Expressing NK-Cell Markers and Chemokine Receptors in Parkinson’s Disease. Journal of Ageing and Longevity. 2023; 3(1):1-10. https://doi.org/10.3390/jal3010001
Chicago/Turabian StyleGoldeck, David, Claudia Schulte, Marcia Cristina Teixeira dos Santos, Dieter Scheller, Lilly Öttinger, Graham Pawelec, Christian Deuschle, Daniela Berg, Andre Nogueira da Costa, and Walter Maetzler. 2023. "Higher Frequencies of T-Cells Expressing NK-Cell Markers and Chemokine Receptors in Parkinson’s Disease" Journal of Ageing and Longevity 3, no. 1: 1-10. https://doi.org/10.3390/jal3010001