

An In Vitro Assay to Quantify Effects of Micro- and Nano-Plastics on Human Gene Transcription


1
Branch Bioanalytics and Bioprocesses, Fraunhofer Institute for Cell Therapy and Immunology (IZI-BB), 14476 Potsdam, Germany
2
Institute of Biotechnology, Faculty III, Technical University Berlin, 10623 Berlin, Germany
*
Author to whom correspondence should be addressed.
Microplastics 2023, 2(1), 122-131; https://doi.org/10.3390/microplastics2010009
Submission received: 8 November 2022
/
Revised: 2 December 2022
/
Accepted: 13 February 2023
/
Published: 15 February 2023
(This article belongs to the Special Issue Ecotoxicological Impacts of Micro(Nano)plastics in the Environment: Biotic and Abiotic Interactions)
Abstract
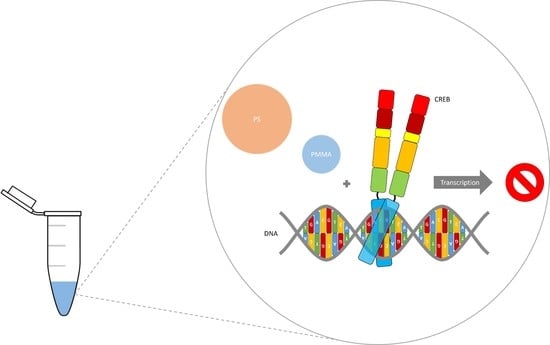
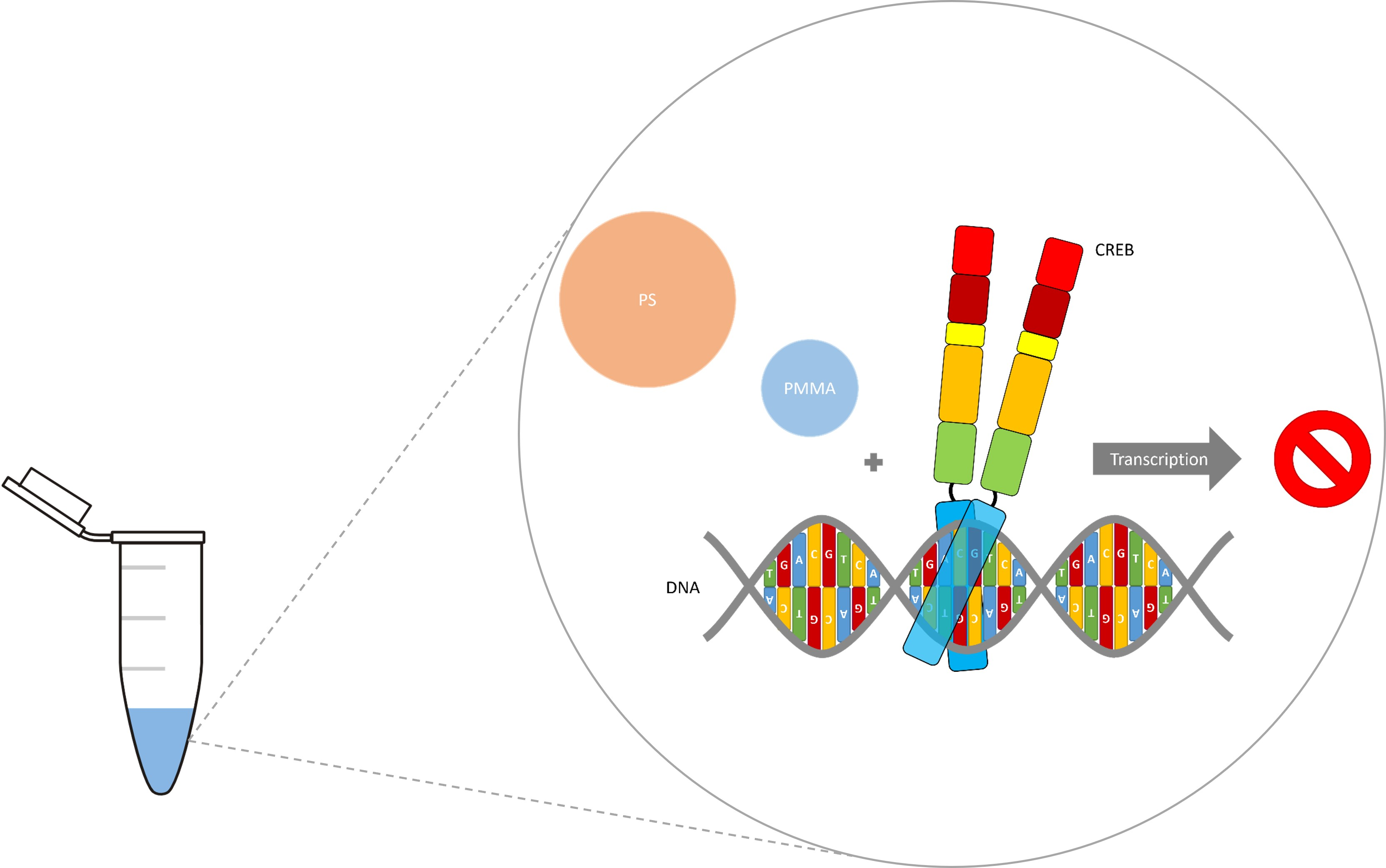
:In today’s age, plastic waste is a major problem for our environment. The decomposition of plastic waste causes widespread contamination in all types of ecosystems worldwide. Micro-plastics in the lower micrometer size range and especially nano-plastics can become internalized by cells and thus become a threat to human health. To investigate the effects of internalized micro- and nano-plastics on human gene transcription, we used an in vitro assay to quantify CREB (cAMP response element binding protein) mediated transcription. Here we show that CREB mediated gene expression was mainly but not exclusively induced by phosphorylation. In addition, the amount of CREB affected transcription was also studied. We were also able to show that the strong CREB mediated stimulation of transcription was diminished by micro- and nano-plastics in any chosen setting. This indicates a threat to human health via the deregulation of transcription induced by internalized micro- and nano-plastics. However, this established quantifiable in vitro transcription test system could help to screen for toxic substances and non-toxic alternatives.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
In today’s age, plastic waste is a major problem for our environment. Since the middle of the 20th century, when the industrial manufacturing of plastics began, they have become incredibly prevalent in both industry and daily life. Plastics’ chemistry allows for the production of practical goods, such as packaging, that is resistant to chemicals and temperature. Additionally, plastics usually have a hydrophobic property, which makes them ideal vessels for toxic compounds and long-lasting organic contaminants [1]. However, the decomposition of plastic waste causes widespread contamination in all types of ecosystems worldwide. In addition, due to the slow decomposition process, the load of micro- and nano-plastics will increase in the near future. Plastics break down step by step into smaller fragments of sizes that are barely visible to the naked eye as they age and decompose in the environment. This decomposition is mostly due to physical and chemical degradation but also due to microbiological breakdown [2]. Due to their worldwide circulation and small sizes, micro- and nano-plastics are considered to be a risk for human health and thus, have caused a sudden surge in studies [3,4]. This surge united scientists from all over the world and from various different disciplines, from chemistry and engineering to biology and exposure sciences, to study how plastics, and especially micro- and nano-plastics, are affecting the environment and the possible hazards they can bring to its inhabitants.
Micro-plastics can be defined as small plastic particles <5 mm in size, while for nano-plastics, there is still no consensus in the literature on their size [5]. Micro- and nano-plastics can originate from primary sources, such as polystyrene (PS) particles in cosmetic and medical products or polymethylmethacrylate (PMMA) copolymer fibers used in the textile and clothing industry. In addition, they can originate from secondary sources, which can become brittle and fragment into micro- and nano-plastics when exposed to physical, chemical, and biological processes [6].
In the marine environment, micro- and nano-plastics can be found in the water, on the ocean floor, and along the shorelines [7]. Since degradation of micro- and nano-plastics is very slow in aquatic environments, they enter the food chain through direct and indirect pathways, causing potential health risks for humans [8]. However, besides food from aquatic environments, other exposure sources of micro- and nano-plastics to humans have been identified; these include sources in terrestrial and atmospheric environments worldwide, and from daily consumed goods like salt, drinking water, and cosmetics [9]. Almost all types of terrestrial environments, and especially agricultural and farmland environments, including greenhouses, home gardens, coastal and industrial soils, as well as floodplain soils, are contaminated by micro- and nano-plastics [10].
Finally, it is now known that atmospheric environments are newly discovered mediums through which micro- and nano-plastics spread easily to other environments worldwide. Thus, micro- and nano-plastics have been measured in atmospheric fallout in metropolises as well as in less populated regions. In addition, micro- and nano-plastics have also been consistently found in indoor air [11,12]. This omnipresent prevalence of micro- and nano-plastics shows that appropriate actions must be taken to identify potential health risks and to identify their countermeasures, as concerns have been raised about possible long term health risks from continuous micro- and nano-plastic exposure.
Recent studies have emphasized potential negative consequences of this omnipresent exposure to micro- and nano-plastic, manifesting in physical stress and damage, inflammation, oxidative stress, immune dysfunction, impaired gut microbiota and intestinal homeostasis, or altered motion in cell cultures, organisms and animal models [13,14,15]. Their ingestion together with food, but also by inhalation and dermal contact, has been shown for various organisms ranging from single cell organisms to humans [16]. From a molecular biology perspective, micro-plastics in the lower micrometer size range, and especially nano-plastics, can become internalized by cells and thus become a threat to human health [17].
To investigate the effects of internalized micro- and nano-plastics, we used an in vitro assay to quantify CREB (cAMP response element binding protein) mediated transcription. The transcription factor CREB was discovered through its ability to control the somatostatin gene expression. The ubiquitously expressed CREB regulates gene expression via binding to CRE (cAMP responsive element), found in many eukaryotic and viral gene promoters [18,19,20]. Additionally, CREB activity is increased by phosphorylation at one or more sites within the KID-domain (kinase-inducible domain) [21]. Thus, PKA (protein kinase A), CaMKIV (calcium/calmodulin-dependent protein kinase type IV), MSK (mitogen- and stress-activated kinase), and RSK (ribosomal S6 kinase) have been shown to enhance CREB mediated transcription [22,23,24]. Phosphorylation of CREB (pCREB) at Ser133 allows the recruitment of CREB’s coactivator CBP (CREB binding protein), which induces increased transcription via its acetyltransferase activity and by facilitating interaction with the basal transcriptional machinery [25,26]. CREB is involved in a large number of biological processes, including long-term neuronal plasticity, cell survival, circadian rhythms, adaptation to drugs, and hormonal regulation of metabolism [27]. Thus, the aim of this study was to investigate the effects of internalized micro- and nano-plastics on human gene transcription via in vitro quantification of CREB mediated gene expression and determine its potential threat to human health.
2. Materials and Methods
2.1. Purification
Human full-length CREB fused to a N-terminal 6xHis-tag was expressed in E. coli BL21-CodonPlus-RIL (catalog number: 230245, Agilent Technologies, Frankfurt am Main, Germany) in liquid culture (LB medium with 34 μg/mL chloramphenicol) at 37 °C to mid-log phase (0.4 OD600 nm) and purified through affinity chromatography using Ni-NTA agarose matrix according to the manufacturer’s instructions (catalog number: 30210; Qiagen, Hilden, Germany), with the exception that the standard buffer was replaced by IVT buffer (20 mM HEPES pH 7.9 at 25 °C, 100 mM potassium chloride, 0.2 mM EDTA, and 0.5 mM dithiothreitol) + 10 mM/250 mM imidazole (wash/elution buffer). The purified protein was dialyzed against IVT buffer for 24 h with double buffer exchange in a 1:1000 ratio using D-Tube Dialyzer Maxi, MWCO 12–14 kDa (catalog number: 71510; Millipore, Darmstadt, Germany) and quantified with Pierce BCA Protein Assay Kit (catalog number: 23225; Thermo Fisher Scientific, Darmstadt, Germany) with bovine serum albumin (Carl Roth GmbH + Co. KG, Karlsruhe, Germany) as standard.
2.2. Western Blot
After running SDS-PAGE, semi-dry western blot analysis was performed. The gel consisted of a resolving gel, which was covered with a stacking gel. For the stacking gel, a polymerization density of 5% was used, 12% for the resolving gel 12%. The proteins separated by SDS-PAGE were transferred to a blotting buffer (4 mM Glycine, 4.8 mM Tris pH 8.5, 0.4% SDS, 20% methanol) activated nitrocellulose membrane (catalog number: 77010; Thermo Fisher Scientific, Darmstadt, Germany) with 1.3 mA/cm2 for 1 h. After blotting, the membrane was blocked with 5% bovine serum albumin (Carl Roth GmbH + Co. KG, Karlsruhe, Germany) in TBS with Tween (catalog number: J77500.K8, Thermo Fisher Scientific, Darmstadt, Germany) for 1 h at room temperature, and then incubated for 1 h at room temperature with the primary antibody dissolved in 5% bovine serum albumin in TBS with Tween. After washing thrice for 5 min, the membrane was incubated with the secondary antibody dissolved in 5% bovine serum albumin in TBS with Tween for 1 h at room temperature and washed again thrice for 5 min. As primary antibodies, pCREB antibody (1:1000 dilution, catalog number: 9198S; Cell Signaling, Leiden, The Netherlands) and His RGS antibody (1:1000 dilution, catalog number: 30210; Qiagen, Hilden, Germany) were used. IgG-Alkaline Phosphatase antibody (1:1000 dilution, catalog number: A3687 and A1293; Sigma-Aldrich Chemie GmbH, Taufkirchen, Germany) served as the secondary antibody. For detection and visualization, Western Blue® Stabilized Substrate for Alkaline Phosphatase (Promega Corporation, Fitchburg, WI, USA) was used.
2.3. Electrophoretic Mobility Shift Assay
Two pmol double-stranded CRE (5′-6FAM-gATCCggCTgACgTCATCAAgCTA; TIB MOLBIOL, Berlin, Germany) was incubated in 20 µL IVT buffer (20 mM HEPES pH 7.9 at 25 °C, 100 mM potassium chloride, 0.2 mM EDTA, and 0.5 mM dithiothreitol) and TA buffer (33 mM Tris acetate pH 7.9, 66 mM potassium acetate, 10 mM magnesium acetate, 0.5 mM dithiothreitol) and 100 μg/mL nuclease-free bovine serum albumin (Carl Roth GmbH + Co. KG, Karlsruhe, Germany) with increasing amounts of CREB and pCREB (54 and 108 pmol) for 20 min at 30 °C and separated on a native 2% agarose gel (catalog number: V3121; Promega Corporation, Fitchburg, WI, USA) with 0.5× Tris-borate-EDTA buffer (catalog number: B52; Thermo Fisher Scientific, Darmstadt, Germany) as the running buffer. The gel was scanned at 487 nm using Typhoon 9400 Fluorescence Imager (GE Healthcare, Braunschweig, Germany).
2.4. In Vitro Phosphorylation of CREB with PKA
The assay was realized through incubation of 347 pmol CREB (12.84 µg) with 3.7 mM ATP (catalog number: R0441; Thermo Fisher Scientific, Darmstadt, Germany), 224 ng murine PKA (Biaffin GmbH & Co KG, Kassel, Germany) in IVT buffer (20 mM HEPES pH 7.9 at 25 °C, 100 mM potassium chloride, 0.2 mM EDTA, and 0.5 mM dithiothreitol) for 20 min at 30 °C. CCE buffer (1 M Tris pH 6.8, 5.7% SDS, 14% glycerol (87%), 7% β-mercaptoethanol, and bromophenol blue) was added to 6.42 µg pCREB and incubated for 5 min at 95 °C. This aliquot was used for western blot analysis. The remaining pCREB was placed on ice and used for electrophoretic mobility shift assay and in vitro transcription experiments the same day, although pCREB activity was stable for several days at 4 °C.
2.5. In Vitro Transcription Assay
The assay was performed with the HeLaScribe® Nuclear Extract in vitro Transcription System according to the manufactures instructions (catalog number: E3110, Promega Corporation, Fitchburg, WI, USA), with the exception that CREB, pCREB, and IVT buffer (20 mM HEPES pH 7.9 at 25 °C, 100 mM potassium chloride, 0.2 mM EDTA, and 0.5 mM dithiothreitol) with murine PKA (Biaffin GmbH & Co. KG, Kassel, Germany) and ATP (catalog number: R0441; Thermo Fisher Scientific, Darmstadt, Germany) were supplemented instead of nuclease-free water. Therefore, CREB, pCREB, and IVT buffer with PKA/ATP were heated for 15 min at 65 °C to inactivate PKA; CREB was not affected. Subsequently, the mix was cooled to room temperature and pre-incubated with 100 ng/μL plasmid (CRE-MGFP, CMV-MGFP, or TATA-MGFP) from the Cignal CRE Reporter Assay Kit (catalog number: 336841; Qiagen, Hilden, Germany) with 4 mM magnesium chloride for 20 min at 30 °C (7). After incubation, residual components were added to 4 μL HeLaScribe® Nuclear Extract. For micro- and nano-plastic in vitro transcription, PS (1 mm, microParticles GmbH, Berlin, Germany) or PMMA (100 nm, microParticles GmbH, Berlin, Germany) were added in the same step. In vitro transcription was stopped after 60 min at 30 °C with HeLa Extract Stop Solution. RNA was extracted via phenol:chloroform:isoamyl alcohol (Carl Roth GmbH + Co. KG, Karlsruhe, Germany), followed by ethanol precipitation, and suspended in 50 µL nuclease-free water. Template plasmids were removed by DNase I (catalog number: 18047019; Thermo Fisher Scientific, Darmstadt, Germany) for 30 min at 37 °C, followed by phenol:chloroform:isoamyl alcohol extraction and ethanol precipitation. DNase I treatment was performed four times and controlled by real-time PCR (CT > 30).
2.6. cDNA Synthesis
Reverse transcription was achieved using the GoScript™ Reverse Transcription System (catalog number: A5003; Promega Corporation, Fitchburg, WI, USA) according to the manufactures instructions, except that each reaction was doubled. Thus 10 µL RNA was preheated with 20 pmol reverse primer, and the residual components were added to a final volume of 30 µL with 4 mM magnesium chloride. Reverse transcription was stopped by phenol:chloroform:isoamyl alcohol (Carl Roth GmbH + Co. KG, Karlsruhe, Germany) extraction, followed by ethanol precipitation, and the obtained pellet suspended in 10 µL nuclease-free water.
2.7. Quantification of Synthezised cDNA
Real-time PCR was carried out using LightCycler® 2.0 Instrument (Roche, Mannheim, Germany) and Maxima SYBR Green/ROX qPCR Master Mix (2×) (catalog number: K0221; Thermo Fisher Scientific, Darmstadt, Germany) according to the manufacturer’s instructions, with the exception that 2 µL cDNA was used in a final volume of 20 µL. The MGFP gene was targeted for amplification with primers 5′ CCgTgAACggCCACAAATTC and 5′ TACACgAAgCAgTCgTCCAC (TIB MOLBIOL, Berlin. Germany). Evaluation of the relative transcription fold change was accomplished by the 2∆∆CT method.
2.8. Statistics
Data are presented as means ± SD, and n is the number of biological replicates. Statistical analysis was performed by using 2-way ANOVA with Dunnett’s multiple comparisons test using Prism 9.1.2 (GraphPad, San Diego, CA, USA). Statistical significance is denoted as ns = not significant, p < 0.05 (*), p < 0.01 (**), p < 0.001 (***), and p < 0.0001 (****).
3. Results
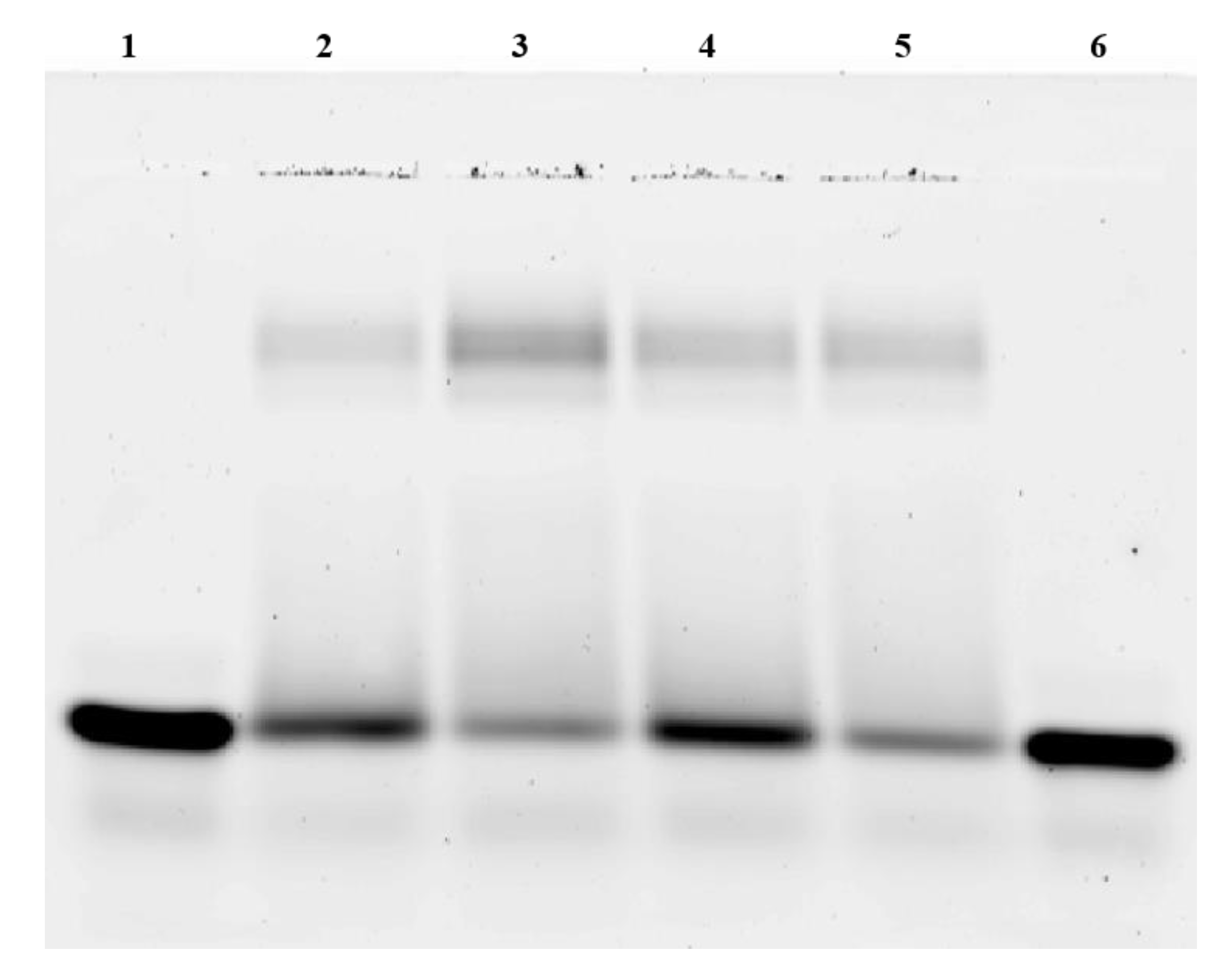
CREB was expressed in E. coli and purified through affinity chromatography. The purity of CREB was tested by SDS-PAGE and western blot, and was shown to be above 85%. After in vitro phosphorylation of CREB via PKA, we verified CREB’s phosphorylation using western blot via a phospho-specific antibody and its binding activity using electrophoretic mobility shift assay (EMSA).
3.1. CREB Binds Independent of Phosphorylation
To test the CRE-binding activity of CREB, EMSA experiments of phosphorylated and non-phosphorylated CREB were performed. Therefore, increasing concentrations of pCREB and CREB were used. Western blot using pCREB antibodies verified the phosphorylation of CREB by PKA. The EMSA results are shown in Figure 1. The CREB-CRE complex that was seen in native 2% agarose gels was in two bands with increasing signal intensities. Binding was detected at a minimum of 16 pmol CREB. At CREB concentrations of 108 pmol, 87% of free CRE (2 pmol) were bound. The data shows that the phosphorylation of CREB had no influence on CRE binding. Phosphorylated CREB showed an identical binding as non-phosphorylated CREB. In addition, increasing amounts of CREB and pCREB were capable to bind CRE in a dose-dependent manner.
3.2. CREB-Mediated Transcription
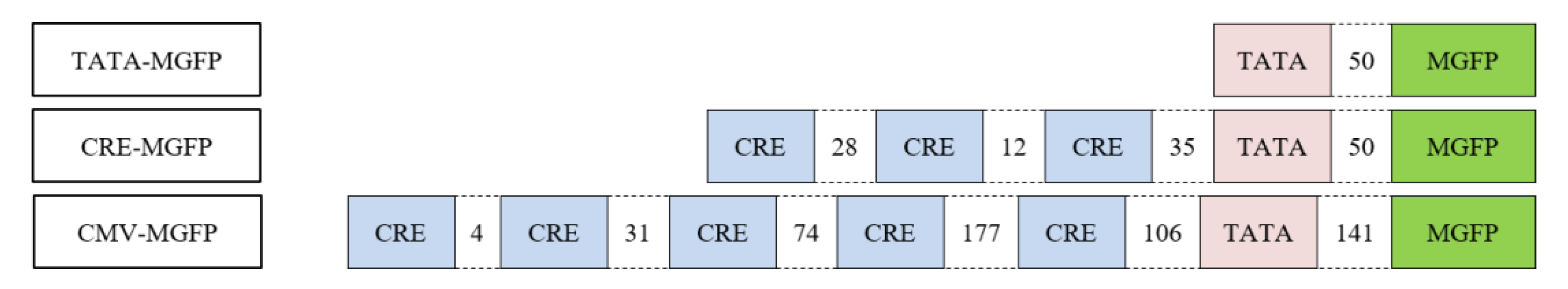
After CREB and pCREB binding was confirmed, in vitro transcription assays were performed. The plasmids used in in vitro transcription experiments were a part of a reporter assay kit for the quantitative assessment of signal transduction pathway activation, in which TATA-MGFP is the negative control, CMV-MGFP is the constitutively expressing positive control, and CRE-MGFP is an inducible reporter construct (Figure 2).
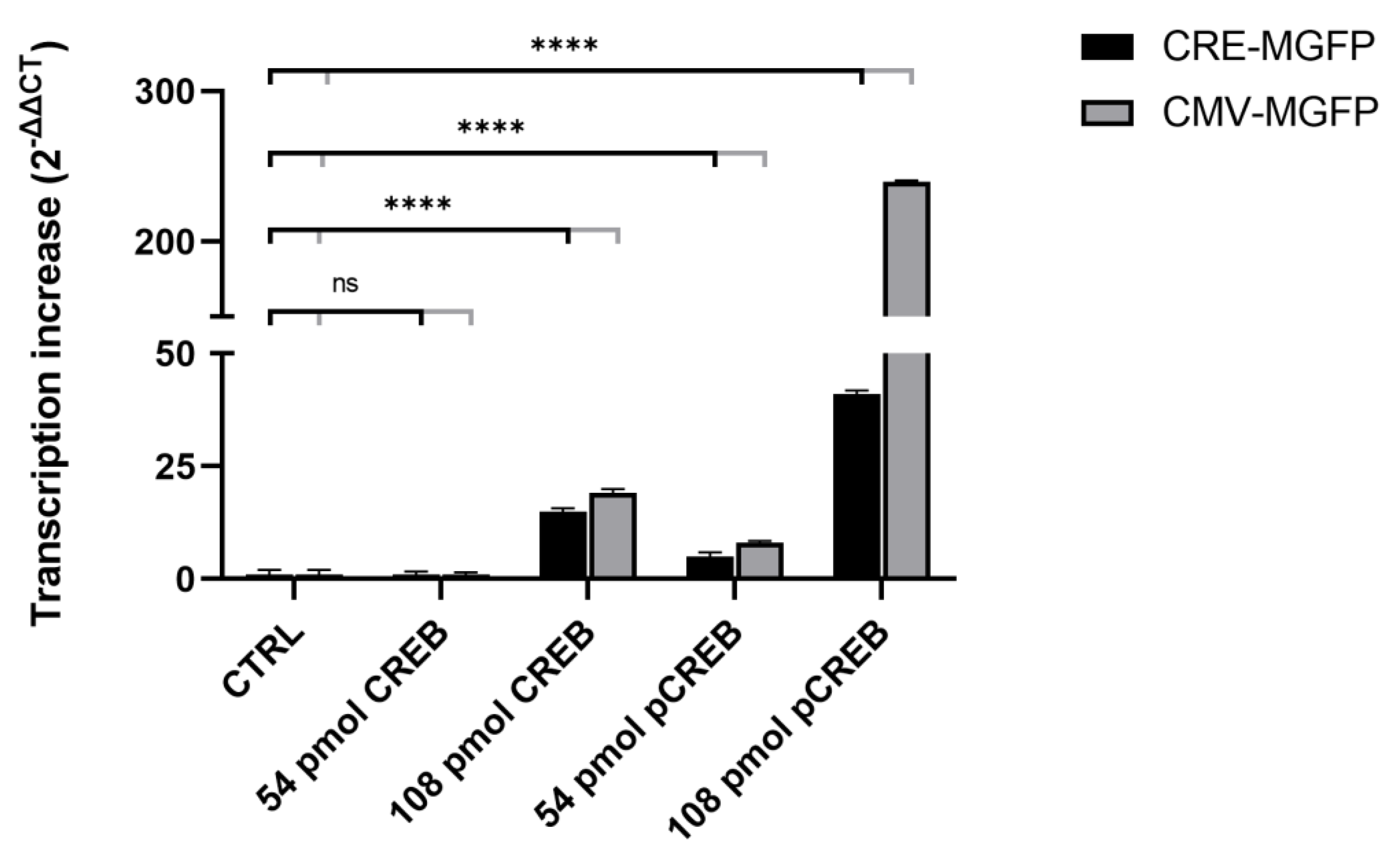
Plasmids were tested with and without CREB, as well as with pCREB. CREB and pCREB concentrations ranged from 0–108 pmol. Through adding CREB or pCREB, increasing amounts of RNA should be generated. Template DNA was removed by DNase I treatment and controlled by real-time PCR (CT > 30). Evaluation of the relative transcription fold change was accomplished by the 2∆∆CT method via the TATA-MGFP negative control. In vitro transcriptions were performed four times (n = 4, biological replicates). The results are shown in Figure 3. The data shows the arithmetic mean as well as the standard deviation. Samples without CREB and pCREB were normalized to 1.00 ± 1.00.
CRE-MGFP as an inducible reporter construct with concentrations of 54 pmol CREB resulted in a non-significant change of 1.29 ± 0.63, while 54 pmol pCREB showed a significant transcription increase of 5.19 ± 0.91. In vitro transcriptions with 108 pmol CREB lead to a significant enhanced gene expression of 14.55 ± 0.71, while 108 pmol pCREB increased transcriptional activity significantly by 41.14 ± 0.86. The data shows that the phosphorylation of CREB increases CREB-mediated transcription. In addition, the data show that this increase is not exclusively due to phosphorylation. Increasing amounts of CREB and pCREB were capable to significantly enhance transcription in a dose-dependent manner. On the other hand, CMV-MGFP, being the constitutively expressing positive control, and CREB with a concentration of 54 pmol does not show a significant change in expression (1.34 ± 0.48), while 54 pmol pCREB elicits a significant increased transcription of 8 ± 0.43. In vitro transcriptions with 108 pmol CREB resulted in a significantly enhanced gene expression of 18.83 ± 0.96, while 108 pmol pCREB showed a significant transcription increase of 239.69 ± 0.72. The data illustrate an even more increased CREB mediated transcription upon phosphorylation than with CRE-MGFP. Again, the data shows that this transcription increase is not exclusively due to phosphorylation but also due to a CREB/pCREB dose-dependency.
3.3. Micro- and Nanoplastic In Vitro Transcription
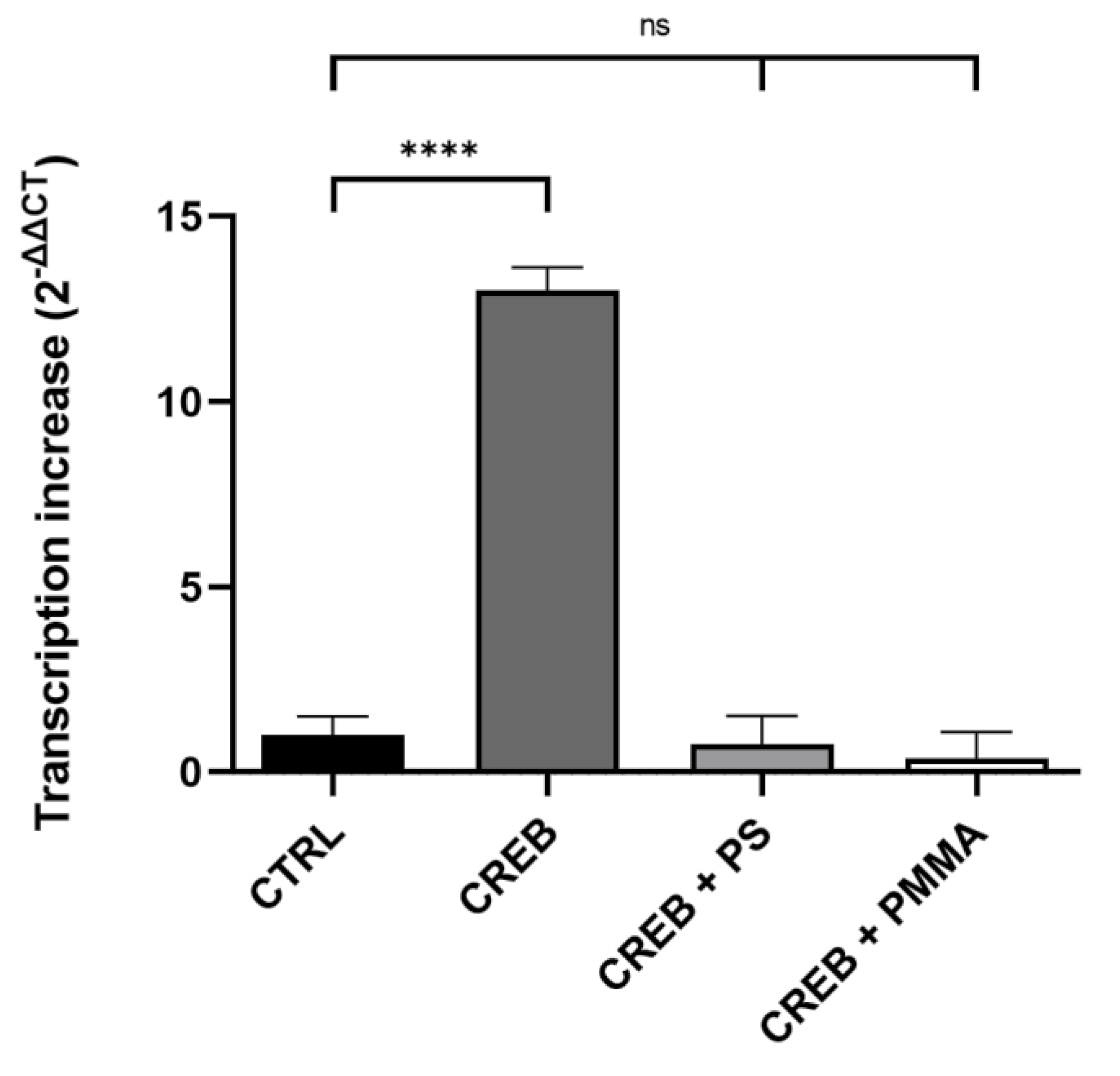
After in vitro transcription confirmed a strong and significant stimulation of CREB mediated transcription, we raised the question if this assay could be used as test system for internalized toxic substances, such as micro- and nano-plastics. Since PS and PMMA are commonly used in cosmetic/medical products and textile/clothing industry, they pollute the environment in different sizes and shapes. Thus, both were used in a different particle sizes to determine the effects of micro- and nano-plastics independent of polymer type. In addition, they were used at environment-like concentrations that were used in the literature for cell culture experiments [28]. More precisely, the CRE-MGFP promoter and CREB were used with or without polystyrene (PS, 100 µg/mL, 1 µm) or polymethylmethacrylate (PMMA, 100 µg/mL, 100 nm). The micro- and nano-plastic in vitro transcription results are shown in Figure 4.
CREB lead to a significantly enhanced gene expression of 13.18 ± 1.25. The addition of PS decreased the CREB-mediated transcription non-significantly to 0.76 ± 1.51 (Figure 4). This corresponds to a decrease of around 94%. On the other hand, the addition of PMMA led to a non-significantly decreased transcription of 0.37 ± 1.42, a decrease of almost 97%. Thus, the data shows that PS and PMMA are able to abolish CREB mediated transcription.
4. Discussion
Whether or not the phosphorylation status of CREB influences CRE binding has been previously debated. Some groups argued that only phosphorylated CREB binds to CRE [29]. In the EMSA experiments performed here, the opposite was demonstrated, thus is in line with the majority of previous experiments analyzing CRE binding of phosphorylated and non-phosphorylated CREB [30,31]. Non-phosphorylated CREB was able to bind CRE, indicating that the PKA modification of CREB had no effect on the DNA binding properties.
This paper is the first that has quantified CREB mediated transcription in relation to the amount of CREB. Transcription activity increased upon phosphorylation of CREB but was not exclusively due to it. This phosphorylation independent gene expression increase is probably facilitated by CREB’s Q2 domain that interacts with the basal transcriptional machinery [32]. Upon phosphorylation, CREB seems to be able to recruit CBP as a coactivator, stabilizing the pre-initiation complex formed at the promoter, resulting in an even higher transcription increase [33]. Several groups showed elevated CMV activation by the cAMP signaling pathway and the stress-activated MAP protein kinase pathway [34,35]. However, the transcription quantification of CMV mediated by CREB and phosphorylated CREB was unknown so far. Our results indicate that the more CRE elements that are present in a promotor, the more it can be activated by CREB/pCREB. Taken together, this would mean for gene regulation that rapid responses occur via phosphorylation, and long-lasting responses tend to occur via the new synthesis of CREB. Additionally, even small amounts of phosphorylated CREB can result in a biological effect.
Furthermore, we wanted to investigate the effects of internalized micro- and nano-plastics. Therefore, we used our new in vitro transcription quantification system to determine the effect of PS and PMMA on human gene expression. Addition of 100 µg/mL PS or PMMA (0.01% w/w) totally abolished the CREB-mediated transcription and showed PS’s and PMMA’s negative effect on human transcription. Ramsperger et al. were able to show cell line dependent negative effects of polystyrene concentrations starting from 150 µg/mL [28]. Thus, our in vitro transcription quantification system is in line with these previous experiments, considering that in vitro transcription, as an isolated process, is probably more error-prone to micro- and nano-plastics than a whole cell that internalizes them.
In addition, a meta-analysis with effect data, available in the literature, derived a concentration of 6650 microplastic (<5 mm) particles per m−3 in the marine environment, resulting in a 0.04% microplastic concentration [36]. Considering this and the results of our experiments with micro- and nano-plastic concentration of 0.01%, the urgent need to screen for safe and non-toxic plastics, as well as plastic alternatives, is highlighted. Micro- and nano-plastic contamination in marine ecosystems has been receiving more attention recently, not only because of the known fact that they are prone to accumulate in marine ecosystems, but also because their occurrence has a negative influence on marine life, which in turn indirectly affects human health via the consumption of seafood [37].
However, micro- and nano-plastic contamination is not only a problem in marine ecosystems but also in freshwater ecosystems. As a recent study shows, micro- and nano-plastic contamination in freshwater ecosystems causes toxicity issues for fish and invertebrates, such as mollusks, and also drinking water for humans [38]. Especially through drinking water, both bottled and tap water, humans are exposed to the omnipresent threat of micro- and nano-plastics on a daily basis.
Despite the potential harmful effects on human health, little research has been done on micro- and nano-plastics in drinking water, and earlier studies tended to concentrate more on micro- and nano-plastics in the freshwater used to produce drinking water than on the water that was actually consumed. However, a recent study was able to quantify an average micro- and nano-plastic content of 0.174 particles/L in drinking water for consumption [39]. Again, this coupled with our results emphasize the urgent need to screen for safe and non-toxic plastics as well as plastic alternatives, in order to overcome potential human health risks.
Since, the human’s exposure to micro- and nano-plastics passes through different routes (ingestion, inhalation, and dermal contact) they are able to cause a variety toxicity issues. Inhaled micro- and nano-plastics can translocate into the respiratory epithelium via diffusion, direct cellular penetration, or active cellular uptake, and induce interstitial fibrosis and granulomatous lesions in the lungs of workers involved in plastic processing. Dermal contact with micro- and nano-plastics is known to induce low inflammatory reactions, fibrous encapsulation, and oxidative stress in humans [40]. Since studies on ingested micro- and nano-plastics in humans are challenging, a meta-analysis revealed that exposure to micro- and nano-plastics, and especially to PS particles, cause inflammation, oxidative stress, lysosomal dysfunction, and apoptosis/genotoxicity in cultured cells. However, these toxic effects have been observed at high concentrations, which may not be relevant compared to real-life exposures in humans [15]. Nevertheless, the present study shows that even lower concentrations than possibly prevalent in the environment are able to negatively affect human gene transcription as an isolated process. It is known that cell organelles and proteins can adsorb to the surface of micro- and nano-plastics, which leads to a change in tertiary and secondary structures and results in functional changes [41]. These functional changes can lead to dysfunctional organelles, resulting in cell malfunction, and can thus become a serious threat for human health. Since this new in vitro transcription quantification system is able to measure the effects of a wide array of plastics (micro- and nano-plastics) on human transcription, the right precautionary measures for each type could be determined.
5. Conclusions
In today’s age, plastic waste is a major problem for our environment and our health. Specifically, micro- and nano-plastics can become internalized by cells and thus become a threat to human health. In this study, we were able to show that the strong CREB mediated stimulation of human gene transcription was diminished by micro- and nano-plastics. This indicates a threat to human health via the deregulation of transcription induced by internalized micro- and nano-plastics. The quantifiable in vitro transcription test system established in this study could be used to screen for toxic and non-toxic substances, as well as to screen for safe exposure concentrations.
Author Contributions
A.P., D.D., C.W. and H.S. designed the experiments. A.P., D.D. and C.W. performed the experiments. A.P. wrote the paper together with C.W. and H.S. All authors discussed the results and commented on the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Casillas, G.; Hubbard, B.C.; Telfer, J.; Zarate-Bermudez, M.; Muianga, C.; Zarus, G.M.; Carroll, Y.; Ellis, A.; Hunter, C.M. Microplastics Scoping Review of Environmental and Human Exposure Data. Microplastics 2023, 2, 78–92. [Google Scholar] [CrossRef]
- Ru, J.; Huo, Y.; Yang, Y. Microbial Degradation and Valorization of Plastic Wastes. Front. Microbiol. 2020, 11, 442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frias, J.; Nash, R. Microplastics: Finding a consensus on the definition. Mar. Pollut. Bull. 2019, 138, 145–147. [Google Scholar] [CrossRef] [PubMed]
- Campanale, C.; Massarelli, C.; Savino, I.; Locaputo, V.; Uricchio, V.F. A Detailed Review Study on Potential Effects of Microplastics and Additives of Concern on Human Health. Int. J. Environ. Res. Public Health 2020, 17, 1212. [Google Scholar] [CrossRef] [Green Version]
- Ricciardi, M.; Pironti, C.; Motta, O.; Miele, Y.; Proto, A.; Montano, L. Microplastics in the Aquatic Environment: Occurrence, Persistence, Analysis, and Human Exposure. Water 2021, 13, 973. [Google Scholar] [CrossRef]
- Akdogan, Z.; Guven, B. Microplastics in the environment: A critical review of current understanding and identification of future research needs. Environ. Pollut. 2019, 254, 113011. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, A.; Ligthart, T.; Boukris, E.; van Harmelen, T. Sources, transport, and accumulation of different types of plastic litter in aquatic environments: A review study. Mar. Pollut. Bull. 2019, 143, 92–100. [Google Scholar] [CrossRef]
- Issac, M.N.; Kandasubramanian, B. Effect of microplastics in water and aquatic systems. Environ. Sci. Pollut. Res. Int. 2021, 28, 19544–19562. [Google Scholar] [CrossRef]
- Usman, S.; Abdull Razis, A.F.; Shaari, K.; Amal, M.N.A.; Saad, M.Z.; Mat Isa, N.; Nazarudin, M.F.; Zulkifli, S.Z.; Sutra, J.; Ibrahim, M.A. Microplastics Pollution as an Invisible Potential Threat to Food Safety and Security, Policy Challenges and the Way Forward. Int. J. Environ. Res. Public Health 2020, 17, 9591. [Google Scholar] [CrossRef]
- Xu, B.; Liu, F.; Cryder, Z.; Huang, D.; Lu, Z.; He, Y.; Wang, H.; Lu, Z.; Brookes, P.C.; Tang, C.; et al. Microplastics in the soil environment: Occurrence, risks, interactions and fate—A review. Crit. Rev. Environ. Sci. Technol. 2020, 50, 2175–2222. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhao, Y.; Du, F.; Cai, H.; Wang, G.; Shi, H. Microplastic Fallout in Different Indoor Environments. Environ. Sci. Technol. 2020, 54, 6530–6539. [Google Scholar] [CrossRef] [PubMed]
- Hirt, N.; Body-Malapel, M. Immunotoxicity and intestinal effects of nano- and microplastics: A review of the literature. Part. Fibre Toxicol. 2020, 17, 57. [Google Scholar] [CrossRef] [PubMed]
- Brandts, I.; Teles, M.; Tvarijonaviciute, A.; Pereira, M.; Martins, M.; Tort, L.; Oliveira, M. Effects of polymethylmethacrylate nanoplastics on Dicentrarchus labrax. Genomics 2018, 110, 435–441. [Google Scholar] [CrossRef]
- Van Raamsdonk, L.W.D.; van der Zande, M.; Koelmans, A.A.; Hoogenboom, R.L.A.P.; Peters, R.J.B.; Groot, M.J.; Peijnenburg, A.A.C.M.; Weesepoel, Y.J.A. Current Insights into Monitoring, Bioaccumulation, and Potential Health Effects of Microplastics Present in the Food Chain. Foods 2020, 9, 72. [Google Scholar] [CrossRef] [Green Version]
- Heddagaard, F.E.; Møller, P. Hazard assessment of small-size plastic particles: Is the conceptual framework of particle toxicology useful? Food Chem. Toxicol. 2020, 136, 111106. [Google Scholar] [CrossRef]
- Nor, N.H.M.; Kooi, M.; Diepens, N.J.; Koelmans, A.A. Lifetime Accumulation of Microplastic in Children and Adults. Environ. Sci. Technol. 2021, 55, 5084–5096. [Google Scholar] [CrossRef]
- Ramsperger, A.F.R.M.; Narayana, V.K.B.; Gross, W.; Mohanraj, J.; Thelakkat, M.; Greiner, A.; Schmalz, H.; Kress, H.; Laforsch, C. Environmental exposure enhances the internalization of microplastic particles into cells. Sci. Adv. 2020, 6, eabd1211. [Google Scholar] [CrossRef]
- Quinn, P.G.; Granner, D.K. Cyclic AMP-dependent protein kinase regulates transcription of the phosphoenolpyruvate carboxykinase gene but not binding of nuclear factors to the cyclic AMP regulatory element. Mol. Cell. Biol. 1990, 10, 3357–3364. [Google Scholar] [CrossRef]
- Moldave, K. (Ed.) . Progress in Nucleic Acid Research and Molecular Biology, 1st ed.; Elsevier: Amsterdam, The Netherlands, 2000. [Google Scholar]
- Gonzalez, G.A.; Montminy, M.R. Cyclic AMP stimulates somatostatin gene transcription by phosphorylation of CREB at serine 133. Cell 1989, 59, 675–680. [Google Scholar] [CrossRef]
- Gonzalez, G.A.; Menzel, P.; Leonard, J.; Fischer, W.H.; Montminy, M.R. Characterization of motifs which are critical for activity of the cyclic AMP-responsive transcription factor CREB. Mol. Cell. Biol. 1991, 11, 1306–1312. [Google Scholar] [CrossRef] [PubMed]
- Naqvi, S.; Martin, K.J.; Arthur, J.S.C. CREB phosphorylation at Ser133 regulates transcription via distinct mechanisms downstream of cAMP and MAPK signalling. Biochem. J. 2014, 458, 469–479. [Google Scholar] [CrossRef] [PubMed]
- Pierrat, B.; Correia, J.S.; Mary, J.L.; Tomás-Zuber, M.; Lesslauer, W. RSK-B, a novel ribosomal S6 kinase family member, is a CREB kinase under dominant control of p38alpha mitogen-activated protein kinase (p38alphaMAPK). J. Biol. Chem. 1998, 273, 29661–29671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, P.; Enslen, H.; Myung, P.S.; Maurer, R.A. Differential activation of CREB by Ca2+/calmodulin-dependent protein kinases type II and type IV involves phosphorylation of a site that negatively regulates activity. Genes Dev. 1994, 8, 2527–2539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chrivia, J.C.; Kwok, R.P.S.; Lamb, N.; Hagiwara, M.; Montminy, M.R.; Goodman, R.H. Phosphorylated CREB binds specifically to the nuclear protein CBP. Nature 1993, 365, 855–859. [Google Scholar] [CrossRef]
- McManus, K.J.; Hendzel, M.J. CBP, a transcriptional coactivator and acetyltransferase. Biochem. Cell Biol. 2001, 79, 253–266. [Google Scholar] [CrossRef]
- Lonze, B.E.; Ginty, D.D. Function and Regulation of CREB Family Transcription Factors in the Nervous System. Neuron 2002, 35, 605–623. [Google Scholar] [CrossRef] [Green Version]
- Ramsperger, A.; Jasinski, J.; Völkl, M.; Witzmann, T.; Meinhart, M.; Jérôme, V.; Kretschmer, W.; Freitag, R.; Senker, J.; Fery, A.; et al. Supposedly identical microplastic particles substantially differ in their material properties influencing particle-cell interactions and cellular responses. J. Hazard. Mater. 2021, 425, 127961. [Google Scholar] [CrossRef]
- Yamamoto, K.K.; Gonzalez, G.A.; Biggs, W.H.; Montminy, M.R. Phosphorylation-induced binding and transcriptional efficacy of nuclear factor CREB. Nature 1988, 334, 494–498. [Google Scholar] [CrossRef]
- Brindle, P.; Linke, S.; Montminy, M. Protein-kinase-A-dependent activator in transcription factor CREB reveals new role for CREM repressors. Nature 1993, 364, 821–824. [Google Scholar] [CrossRef]
- Richards, J.P.; Bächinger, H.P.; Goodman, R.H.; Brennan, R.G. Analysis of the structural properties of cAMP-responsive element-binding protein (CREB) and phosphorylated CREB. J. Biol. Chem. 1996, 271, 13716–13723. [Google Scholar] [CrossRef]
- Xing, L.; Gopal, V.K.; Quinn, P.G. cAMP Response Element-binding Protein (CREB) Interacts with Transcription Factors IIB and IID. J. Biol. Chem. 1995, 270, 17488–17493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.-A.; Lu, J.; Quinn, P.G. Distinct cAMP response element-binding protein (CREB) domains stimulate different steps in a concerted mechanism of transcription activation. Proc. Natl. Acad. Sci. USA 2000, 97, 11292–11296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruening, W.; Giasson, B.; Mushynski, W.; Durham, H.D. Activation of stress-activated MAP protein kinases up-regulates expression of transgenes driven by the cytomegalovirus immediate/early promoter. Nucleic Acids Res. 1998, 26, 486–489. [Google Scholar] [CrossRef] [PubMed]
- Keller, M.J.; Wu, A.W.; Andrews, J.I.; McGonagill, P.W.; Tibesar, E.E.; Meier, J.L. Reversal of Human Cytomegalovirus Major Immediate-Early Enhancer/Promoter Silencing in Quiescently Infected Cells via the Cyclic AMP Signaling Pathway. J. Virol. 2007, 81, 6669–6681. [Google Scholar] [CrossRef] [Green Version]
- Everaert, G.; Van Cauwenberghe, L.; De Rijcke, M.; Koelmans, A.A.; Mees, J.; Vandegehuchte, M.; Janssen, C.R. Risk assessment of microplastics in the ocean: Modelling approach and first conclusions. Environ. Pollut. 2018, 242, 1930–1938. [Google Scholar] [CrossRef]
- Almeida, M.P.d.; Gaylarde, C.; Pompermayer, F.C.; Lima, L.d.S.; Delgado, J.d.F.; Scott, D.; Neves, C.V.; Vieira, K.S.; Baptista Neto, J.A.; Fonseca, E.M. The Complex Dynamics of Microplastic Migration through Different Aquatic Environments: Subsidies for a Better Understanding of Its Environmental Dispersion. Microplastics 2023, 2, 62–77. [Google Scholar] [CrossRef]
- Isinibilir, M.; Eryalçın, K.M.; Kideys, A.E. Effect of Polystyrene Microplastics in Different Diet Combinations on Survival, Growth and Reproduction Rates of the Water Flea (Daphnia magna). Microplastics 2023, 2, 27–38. [Google Scholar] [CrossRef]
- Kirstein, I.V.; Hensel, F.; Gomiero, A.; Iordachescu, L.; Vianello, A.; Wittgren, H.B.; Vollertsen, J. Drinking plastics?—Quantification and qualification of microplastics in drinking water distribution systems by µFTIR and Py-GCMS. Water Res. 2020, 188, 116519. [Google Scholar] [CrossRef]
- Pironti, C.; Ricciardi, M.; Motta, O.; Miele, Y.; Proto, A.; Montano, L. Microplastics in the Environment: Intake through the Food Web, Human Exposure and Toxicological Effects. Toxics 2021, 9, 224. [Google Scholar] [CrossRef]
- Wang, J.; Jensen, U.B.; Jensen, G.V.; Shipovskov, S.; Balakrishnan, V.S.; Otzen, D.; Pedersen, J.S.; Besenbacher, F.; Sutherland, D.S. Soft Interactions at Nanoparticles Alter Protein Function and Conformation in a Size Dependent Manner. Nano Lett. 2011, 11, 4985–4991. [Google Scholar] [CrossRef]
Figure 1.
EMSA of CREB and pCREB used for in vitro transcription. All lanes contain 2 pmol CRE. Lanes 2 and 3 harbor CREB (54 and 108 pmol, respectively) additionally. Lanes 3 and 4 contain pCREB (54 and 108 pmol, respectively) additionally.
Figure 1.
EMSA of CREB and pCREB used for in vitro transcription. All lanes contain 2 pmol CRE. Lanes 2 and 3 harbor CREB (54 and 108 pmol, respectively) additionally. Lanes 3 and 4 contain pCREB (54 and 108 pmol, respectively) additionally.

Figure 2.
TATA-MGFP contains only a TATA-box upstream of MGFP; CRE-MGFP is controlled by three CRE sites and a TATA-box upstream of MGFP; CMV-MGFP harbors five CRE sites and a TATA-box upstream of MGFP.
Figure 2.
TATA-MGFP contains only a TATA-box upstream of MGFP; CRE-MGFP is controlled by three CRE sites and a TATA-box upstream of MGFP; CMV-MGFP harbors five CRE sites and a TATA-box upstream of MGFP.

Figure 3.
Quantification of CREB mediated CRE-MGFP and CMV-MGFP in vitro transcriptions. Transcription increases with CREB and pCREB ranging from 0–108 pmol. Data are presented as means ± standard deviation. Statistical significance is denoted as ns = not significant and p < 0.0001 (****). n = 4 (biological replicates).
Figure 3.
Quantification of CREB mediated CRE-MGFP and CMV-MGFP in vitro transcriptions. Transcription increases with CREB and pCREB ranging from 0–108 pmol. Data are presented as means ± standard deviation. Statistical significance is denoted as ns = not significant and p < 0.0001 (****). n = 4 (biological replicates).

Figure 4.
Quantification of micro- and nano-plastics in vitro transcription with CRE-MGFP and CREB. PS = 100 µg/mL, 1 mm); PMMA = 100 µg/mL, 100 nm). Data are presented as means ± standard deviation. Statistical significance is denoted as ns = not significant and p < 0.0001 (****). n = 4 (biological replicates).
Figure 4.
Quantification of micro- and nano-plastics in vitro transcription with CRE-MGFP and CREB. PS = 100 µg/mL, 1 mm); PMMA = 100 µg/mL, 100 nm). Data are presented as means ± standard deviation. Statistical significance is denoted as ns = not significant and p < 0.0001 (****). n = 4 (biological replicates).

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Pellegrino, A.; Danne, D.; Weigel, C.; Seitz, H. An In Vitro Assay to Quantify Effects of Micro- and Nano-Plastics on Human Gene Transcription. Microplastics 2023, 2, 122-131. https://doi.org/10.3390/microplastics2010009
AMA Style
Pellegrino A, Danne D, Weigel C, Seitz H. An In Vitro Assay to Quantify Effects of Micro- and Nano-Plastics on Human Gene Transcription. Microplastics. 2023; 2(1):122-131. https://doi.org/10.3390/microplastics2010009
Chicago/Turabian StylePellegrino, Antonio, Denise Danne, Christoph Weigel, and Harald Seitz. 2023. "An In Vitro Assay to Quantify Effects of Micro- and Nano-Plastics on Human Gene Transcription" Microplastics 2, no. 1: 122-131. https://doi.org/10.3390/microplastics2010009