
Size- and Stereochemistry-Dependent Transcriptional Bypass of DNA Alkyl Phosphotriester Adducts in Mammalian Cells

1
Environmental Toxicology Graduate Program, University of California Riverside, Riverside, CA 92521-0403, USA
2
Department of Chemistry, University of California Riverside, Riverside, CA 92521-0403, USA
*
Author to whom correspondence should be addressed.
DNA 2022, 2(4), 221-230; https://doi.org/10.3390/dna2040016
Submission received: 10 September 2022
/
Revised: 28 September 2022
/
Accepted: 29 September 2022
/
Published: 5 October 2022
(This article belongs to the Special Issue From Mutation and Repair to Therapeutics)
Abstract
:Environmental, endogenous and therapeutic alkylating agents can react with internucleotide phosphate groups in DNA to yield alkyl phosphotriester (PTE) adducts. Alkyl-PTEs are induced at relatively high frequencies and are persistent in mammalian tissues; however, their biological consequences in mammalian cells have not been examined. Herein, we assessed how alkyl-PTEs with different alkyl group sizes and stereochemical configurations (SP and RP diastereomers of Me and nPr) affect the efficiency and fidelity of transcription in mammalian cells. We found that, while the RP diastereomer of Me- and nPr-PTEs constituted moderate and strong blockages to transcription, respectively, the SP diastereomer of the two lesions did not appreciably perturb transcription efficiency. In addition, none of the four alkyl-PTEs induced mutant transcripts. Furthermore, polymerase η assumed an important role in promoting transcription across the SP-Me-PTE, but not any of other three lesions. Loss of other translesion synthesis (TLS) polymerases tested, including Pol κ, Pol ι, Pol ξ and REV1, did not alter the transcription bypass efficiency or mutation frequency for any of the alkyl-PTE lesions. Together, our study provided important new knowledge about the impact of alkyl-PTE lesions on transcription and expanded the substrate pool of Pol η in transcriptional bypass.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
Exposure to endogenous and exogenous DNA damaging agents can compromise genomic integrity in human cells [1]. Unrepaired DNA lesions may impede DNA replication and transcription, and elicit mutations in these processes, which may contribute to the development of cancer and other human diseases [1]. Alkylation is a major form of DNA damage, and it can arise from metabolism, environmental exposure, and cancer chemotherapy [2,3,4,5].
Understanding the implications of these lesions in the etiology for developing human diseases requires knowledge about how alkylated DNA lesions inhibit DNA replication and transcription, how they induce mutations in these processes, and how they are repaired. Such knowledge may also facilitate the development of better strategies for cancer chemotherapy. In this vein, modulating the activities of translesion synthesis (TLS) and DNA repair is known to affect the anti-tumor activities of cancer chemotherapeutic agents [5,6,7,8,9]. Moreover, combination therapy, where DNA alkylating agents are used in conjunction with small-molecule inhibitors for DNA damage response or repair, is now in clinical trials for cancer treatment [5].
Depending on the nature of the alkylating agents involved, the size of the alkyl groups adducted to nucleobases can vary substantially. For example, some widely prescribed anticancer drugs can elicit methylation and chloroethylation of DNA [3,6] and metabolites of some tobacco-derived N-nitrosamines can lead to the adduction of pyridyloxobutyl (POB) and pyridylhydroxybutyl (PHB) groups with DNA [10].
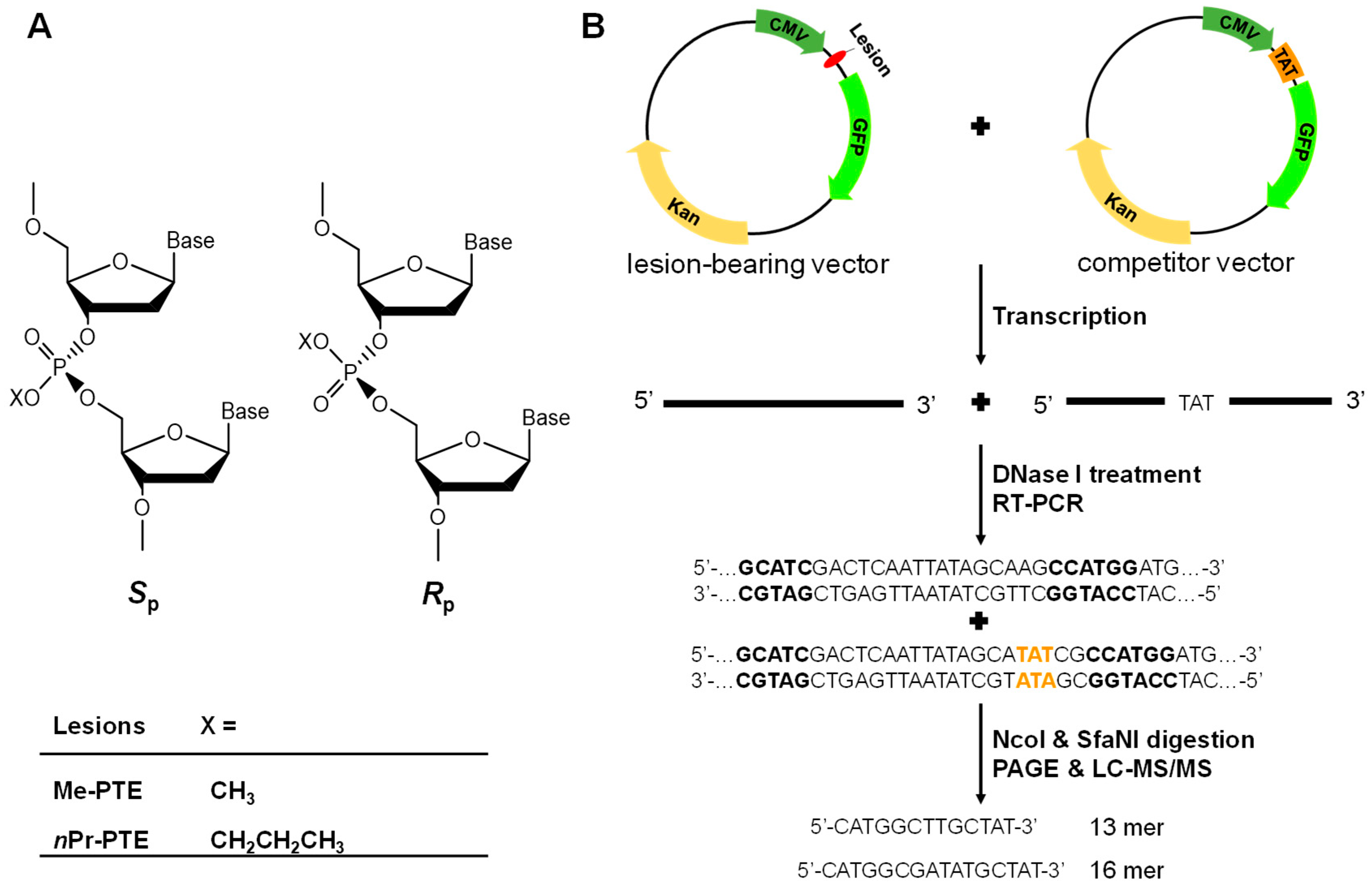
Apart from nucleobases, alkylating agents can attack internucleotide phosphate group in DNA to yield alkyl phosphotriester (alkyl-PTE) lesions [11]. For example, 12–17% and 55–57% of the total alkylation products in duplex DNA treated with N-methyl-N-nitrosourea and N-ethyl-N-nitrosourea are Me- and Et-PTE, respectively [12]. Additionally, PHB-PTE adducts constitute 38–55% and 34–40% of all the measured pyridine-containing DNA lesions in lung and liver tissues, respectively, of rats exposed with nicotine-derived nitrosamine ketone (NNK) in drinking water [13]. Moreover, depending on which of the two non-carbon-bound oxygen atoms on the backbone phosphate is alkylated, alkyl-PTEs can form in the SP or RP configuration (Figure 1A) [11].
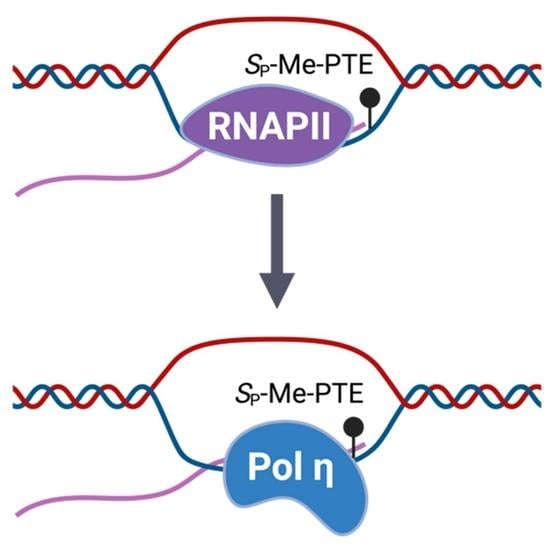
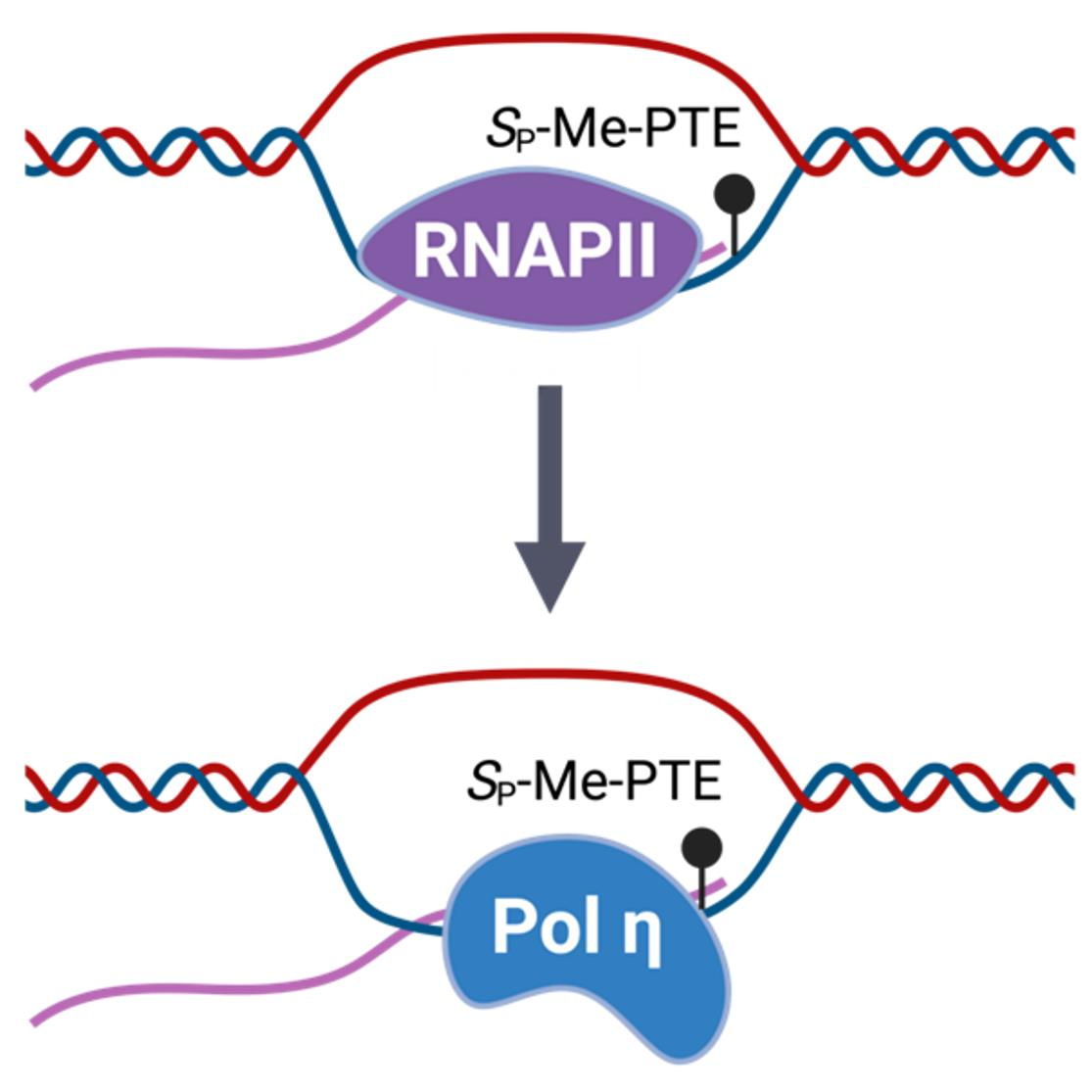
Alkyl-PTE lesions may be more difficult to repair than nucleobase alkylation products. For instance, in mouse liver tissues, Et-PTE lesions exhibited a longer half-life than any nucleobase ethylation products [14]. The poor repair of alkyl-PTE lesions suggests that they are likely encountered by DNA replication and transcription machineries [15]. Little, however, is known about how alkyl-PTE lesions compromise the efficiency and fidelity of DNA replication or transcription in human cells [11]. TLS is an important component of cells’ DNA damage tolerance system [16]. We observed recently that TLS polymerase (Pol) η could promote the transcriptional bypass of the minor-groove N2-alkyl-2′-deoxyguanosine adducts in human cells [17]. It remains unclear whether Pol η would also modulate the transcriptional bypass of alkyl-PTE lesions.
In the present study, we sought to fill in these knowledge gaps by investigating the transcriptional outcomes of alkyl-PTEs in mammalian cells, and the alterations of such outcomes by loss of function of some important TLS polymerases.
2. Experimental Procedures
Except otherwise specified, all chemicals were acquired from Sigma-Aldrich (St. Louis, MO, USA) or Thermo Fisher Scientific (Waltham, MA, USA), and all enzymes were from New England Biolabs (Ipswich, MA, USA). Turbo DNase and M-MLV reverse transcriptase were from Invitrogen (Waltham, MA, USA) and Promega (Madison, WI, USA), respectively. Lesion-free oligodeoxynucleotides (ODNs) used in this study were obtained from Integrated DNA Technologies (Coralville, IA, USA). [γ-32P] ATP was purchased from Perkin Elmer Life Sciences (Waltham, MA, USA).
The 12-mer ODNs carrying a site-specifically inserted Tp(alkyl)T (alkyl = Me or nPr) were freshly synthesized following the same method as reported previously [18]. The HEK293T human embryonic kidney epithelial cells were obtained from ATCC and cultured in Dulbecco’s Modified Eagle’s medium supplemented with 10% fetal bovine serum. HEK293T cells depleted of Pol η, Pol ι, Pol κ, Pol ζ and REV1 were previously generated using CRISPR/Cas9 genome editing [19,20].
2.1. Construction of Transcription Templates
We adhered to previously described procedures [21,22] to prepare the non-replicative double-stranded vectors with an alkyl-PTE being inserted at a specific position. In brief, lesion-free control vector was modified from the non-replicating pTGFP-T7-Hha10 plasmid by EcoRI-NheI digestion, purification and ligation with a 50-mer ODN (5′-CTAGCGGATGCATCGACTCAATTATAGCAAGCCATGGATGACTCGCTGCG-3′) and its complementary strand. The control vectors were then nicked with Nt. BstNBI and annealed with a 25 mer complementary ODN (5′-CATCGACTCAATTATAGCAAGCCAT-3′) in excess to remove the cleaved 25 mer single-stranded ODN. Following purification of the gapped plasmids from the mixture by using a 100 kDa cutoff centrifugal filter, the 5′-phosphorylated 12 mer alkyl-PTE-containing ODN, 5′-ATGGCTp(x)TGCTAT-3′ (x = Me or nPr), and 13 mer lesion-free ODN (5′-AATTGAGTCGATG-3′) were annealed into the gap. T4 DNA ligase was then added to seal the gap. Finally, the fully ligated, supercoiled lesion-containing plasmids were isolated from the ligation mixture by agarose gel electrophoresis and gel extraction.
2.2. Cellular Transcription, RNA Extraction and Amplification
Cellular transcription experiments were carried out as previously reported [21,22]. The lesion-containing plasmids and the lesion-free control plasmid were individually premixed with the competitor plasmid at a molar ratio of 5:1 (lesion/competitor) and 1:1 (control/competitor), respectively, and used as templates for transfection. HEK293T cells and the isogenic TLS polymerase-deficient cells were seeded (1 105) in 24-well plates and cultured at 37 °C in 5% CO2 overnight, followed by transfection with 50 ng of the templates and 450 ng carrier plasmid (self-ligated pGEM-T, Promega) using TransIT-2020 (Mirus Bio, Madison, WI, USA) in accordance with the manufacturer’s instructions. After transfection, the transcripts of the mixed genomes were recovered from cells 24 h later using Total RNA Kit I (Omega Bio-tek, Norcross, GA, USA), and any remaining DNA was eliminated using a DNA-free kit (Ambion, Austin, TX, USA). The relevant transcripts were reverse transcribed and PCR-amplified, as previously described [21].
2.3. Restriction Digestion and Polyacrylamide Gel Electrophoresis (PAGE) Analysis
Prior to PAGE analysis, samples were prepared using restriction digestion with NcoI/SfaNI and post-labeling. In 10 μL of NEB buffer 3.1, 150 ng of the RT-PCR products from each sample were treated with 5 U NcoI and 1 U shrimp alkaline phosphatase (rSAP) at 37 °C for 1 h before being heated at 80 °C for 20 min. The newly released 5′-termini were then radiolabeled by adding 5 U T4 polynucleotide kinase and 1.66 pmol [γ-32P]ATP. The resulting mixture was heated at 75 °C for 20 min and subsequently digested with 2 U SfaNI in 20 μL of NEB buffer 3.1 at 37 °C for 1.5 h. The DNA mixture was resolved using 30% native PAGE (acrylamide/bis-acrylamide = 19:1) and quantified by phosphorimager after the reaction was terminated with 20 μL formamide gel-loading buffer (2) [21]. The relative bypass efficiency (RBE) was calculated from the intensities of the radiolabeled DNA bands using the formula of RBE (%) = (lesion signal/competitor signal)/(control signal/competitor signal) , where the competitor signal served as the internal standard.
2.4. LC-MS/MS Analysis
The transcription products resulting from the alkyl-PTE-containing templates were identified using LC-MS and MS/MS, as previously described [21,22]. RT-PCR products were treated with 50 U NcoI and 20 U rSAP in 250 µL of NEB buffer 3.1 at 37 °C for 2 h, followed by heating at 80 °C for 20 min. SfaNI (50 U) was added to the resultant solution, and allowed to react with amplification products at 37 °C for 2 h before being extracted with phenol, chloroform, and isoamyl alcohol (25:24:1, v/v). To precipitate the DNA, 2.5 volumes of ethanol and 0.1 volume of 3.0 M sodium acetate were added to the collected aqueous phase. The DNA pellet was redissolved in water and used for LC-MS and MS/MS analysis. An LTQ linear ion trap mass spectrometer (Thermo Fisher Scientific) was set up in the negative-ion mode for monitoring the fragmentations of the [M-3H]3− ions of the 13-mer ODNs, 5′-pCATGGCMNGCTAT-3′, where “MN” designates TA, TT, TC, or TG.
3. Results
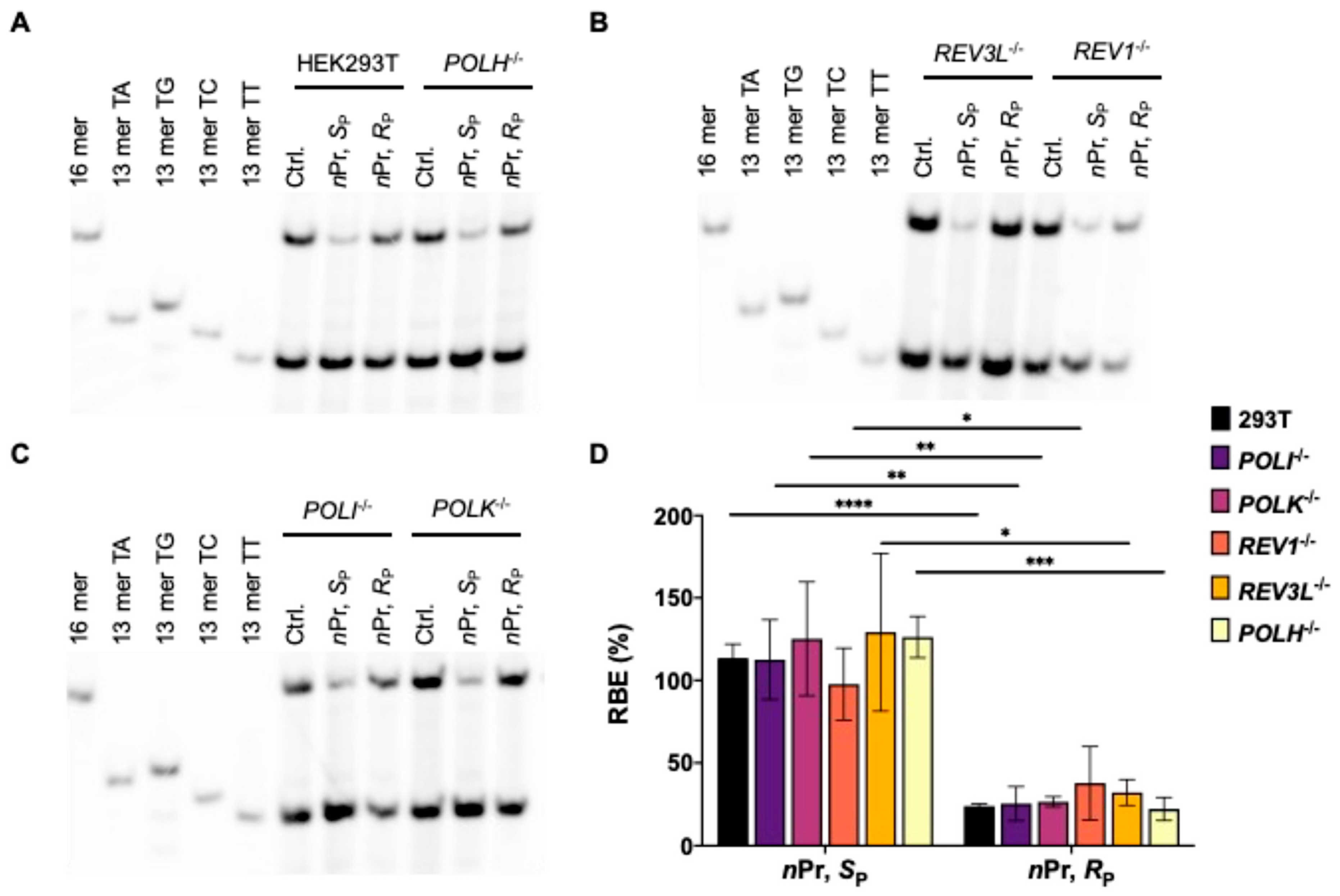
Most human cells are not continuously proliferating or replicating but rather quiescent or slowly replicating in nature; therefore, maintaining faithful and efficient transcription is pivotal for normal cell function [23,24,25]. Recently, our laboratory developed a competitive transcription and adduct bypass (CTAB) assay for investigating transcriptional perturbations conferred by DNA lesions in vitro and in cells [20]. By using this assay (Figure 1), we investigated the impact of two pairs of alkyl-PTEs, i.e., the SP and RP diastereomers of T(Me)T and T(nPr)T (Figure 1), on the efficiency and fidelity of transcription in mammalian cells. In this method, we prepared non-replicative double-stranded vectors harboring a single site-specifically incorporated alkyl-PTE as well as the corresponding non-lesion control and competitor vectors. Relative to the lesion-harboring or control plasmid, the competitor plasmid contains three additional nucleotides (Figure 1B) and serves as an internal reference for quantifying the extent to which a lesion in a double-stranded plasmid impedes cellular transcription. The alkyl-PTE lesions were placed downstream of the transcription start site of the cytomegalovirus (CMV) promoter. This design enables the examination about the effects of DNA lesions on transcription elongation mediated by mammalian RNA polymerase II (RNAPII).
In the CTAB assay (Figure 1B), the lesion-containing or undamaged control plasmids are pre-mixed individually with the competitor vector at specific molar ratios and used as templates for cellular transcription. The resulting transcripts are isolated and incubated with RNase-free DNase I to eliminate residual contaminating DNA template. The ensuing runoff transcripts are reverse transcribed to produce cDNA, which is then PCR amplified. The PCR amplicons are subsequently digested with two restriction enzymes, and the digestion products are subjected to PAGE and LC-MS/MS analyses [21]. The degree through which a DNA adduct impedes transcription, represented as RBE, is determined from the initial genome ratios employed for the transfection and the ratio of the amounts of transcription products formed from the damage-containing genome (i.e., the 13mer products shown in Figure 2) over that from the competitor genome (i.e., the 16mer product, Figure 2), where we also take into account the corresponding ratio obtained for the control damage-free genome over the competitor genome.
3.1. Effects of Alkyl-PTE Lesions on Transcription in Mammalian Cells
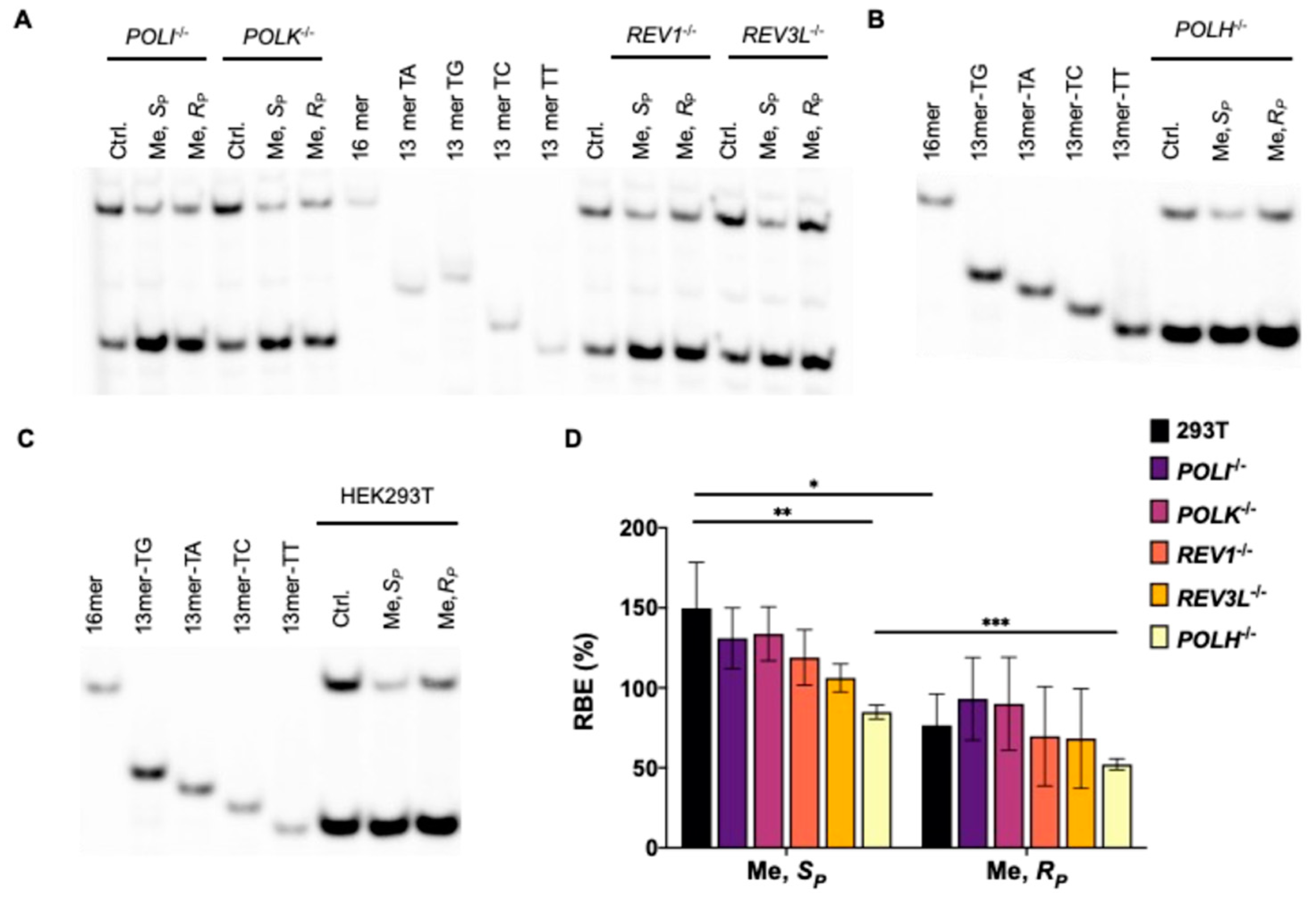
LC-MS/MS and PAGE analyses for the restriction fragments of RT-PCR products showed that the addition of alkyl groups to the phosphate backbone did not perturb transcription fidelity, as manifested by the absence of mutagenic transcription products for the PTE adducts examined (Figure 2, Figure 3, Figure 4 and Figure 5). The efficiencies of transcription, however, were differentially affected by the stereochemical configurations of the alkyl groups. In HEK293T cells, while the SP diastereomers of the Me- and nPr-PTE lesions are well tolerated, the RP diastereomer of the two lesions constitute moderate and strong impediments to transcription elongation, respectively (Figure 2 and Figure 3).
3.2. Pol η Promotes the Transcription of SP-Me-Alkyl-PTE Lesions in Mammalian Cells
DNA polymerase (Pol) η is a Y-family TLS polymerase recently shown to support the transcriptional bypass of N2-alkyl-2′-deoxyguanosine (N2-alkyl-dG) lesions [17]. To examine whether Pol η or other TLS polymerases can modulate the transcription across the alkyl-PTE lesions, we performed the aforementioned CTAB assay in TLS polymerase-knockout cell lines, which were previously generated through CRISPR/Cas9 and single colony isolation from HEK293T cells [19,20]. We observed that genetic ablation of POLH in HEK293T cells conferred a substantial diminution in RBE for SP-Me-PTE, whereas the corresponding change in RBE for RP-Me-PTE was very subtle (Figure 2). However, loss of Pol η did not influence the transcriptional efficiency of nPr-PTEs lesions (Figure 3). Likewise, genetic depletion of Pol ι, Pol κ, Pol ζ or REV1 did not affect appreciably the efficiencies in transcriptional bypass of any of the alkyl-PTE lesions (Figure 2 and Figure 3).
4. Discussion
We systematically investigated, for the first time, how alkyl-PTE lesions compromise the efficiencies and fidelities of transcription in mammalian cells. We found that transcription fidelity is not perturbed by alkyl-PTEs as transcription over alkyl-PTEs is error-free. Previously, our laboratory showed that replication across RP-Me-PTE and both diastereomers of nBu-PTEs is error-free in Escherichia coli cells, whereas replicative bypass of the SP-Me-PTE formed at XT sequence context (X = A, T, C, G), which is a cognitive substrate for Ada protein, is error-prone [26,27]. There is, however, no mammalian ortholog of Ada with a methylphosphotriester-DNA methyltransferase activity [28]. We observed that the transcript yields for the alkyl-PTE lesions are affected by the size and stereochemistry of the alkyl groups. The RP diastereomers Me- and nPr-PTE impose moderate and strong blockages on transcription, respectively; the SP-nPr diastereomers of the two lesions are, however, well tolerated (Figure 2 and Figure 3). It was reported that many of the small non-bulky DNA adducts, including thymidine glycol, abasic site, O6-methylguanine and 8-oxo-7,8-dihydro-2′-deoxyguanosine (8-oxo-dG), did not block RNA polymerase. [29,30,31,32,33,34] On the contrary, the strong helix-distorting bulky adducts can completely hinder RNAP II from forward translocation [35,36].
Our observation of the pivotal effects of stereochemical configuration of alkyl-PTE lesions on transcriptional bypass efficiencies in human cells is reminiscent of the influence of stereochemical configuration of the alkyl-PTEs on DNA replication in E. coli cells, where the RP diastereomers were replicated at markedly lower efficiencies than the SP counterparts [18,26,27]. These results suggest that the alkyl-PTE lesions in the SP configuration, with the alkyl group pointing perpendicular out from the phosphate backbone [11], are in favor of interacting with DNA and RNA polymerases, whereas the lesions in the RP configuration, with the alkyl group projecting into the major groove of DNA double helix [11], hinder damage bypass mediated by both DNA and RNA polymerases. It will be important to assess, by employing molecular modeling and structural biology tools, the molecular basis underlying the effects of the size and stereochemistry of the alkyl-PTE lesions on transcriptional bypass in the future.
Alkyl-PTEs are known to be poorly repaired in mammalian tissues; hence, these lesions may be encountered by DNA replication and transcription machineries. Our result suggested that Pol η may be able to assist transcriptional bypass of the SP-Me-PTE lesion. This result parallels our recent finding that Pol η promotes the transcriptional bypass of N2-alkyl-dG lesions in human cells [17]. Among all the TLS polymerases, Pol η is especially versatile. Deficiency in this polymerase is known to contribute to the variant form of xeroderma pigmentosum (XPV) [37], and this is due to Pol η’s role in efficient and accurate bypass of the UV-induced cyclobutane pyrimidine dimer (CPD) during replication [38]. In addition, Pol η is able to robustly insert and elongate with the correct rNTP opposite CPD-containing template, [39,40] and the polymerase is responsible for inserting consecutive ribonucleotides into genomic DNA [40,41,42]. Moreover, Saccharomyces cerevisiae Pol η extends the RNA primer with rNTPs an order of magnitude more efficiently than inserting rNTPs into DNA [43]. Moreover, Pol η’s large active site can accommodate RNA/DNA, DNA/DNA or DNA/RNA duplex [40]. It is therefore tempting to speculate that Pol η may assist RNAPII in transcription across small unrepaired PTE lesion. Although our results cannot directly exclude the possibility that Pol η may participate in repair, it was previously demonstrated that Pol η’s recruitment to UV-induced damage sites outside of S phase was independent of nucleotide excision repair in human cells [44] Another specialized DNA polymerase, Pol β, can incorporate rCTP opposite 8-oxo-dG [45] and it will be interesting to explore whether other polymerases, including Pol β, also contribute to transcriptional bypass of alkyl-PTE lesions.
Author Contributions
Conceptualization, Y.W.; methodology, Y.T.; investigation, Y.T.; resources, J.W., G.C., L.L. and H.D.; data curation, Y.T.; writing—original draft preparation, Y.T.; writing—review and editing, Y.W.; visualization, Y.T.; supervision, Y.W.; funding acquisition, Y.W. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by National Institutes of Health, grant number R01 CA236204.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Friedberg, E.C.; Walker, G.C.; Siede, W.; Wood, R.D.; Schultz, R.A.; Ellenberger, T. DNA Repair and Mutagenesis; ASM Press: Washington, DC, USA, 2006. [Google Scholar]
- Singer, B.; Grunberger, D. Molecular Biology of Mutagens and Carcinogens; Plenum Press: New York, NY, USA; London, UK, 1983. [Google Scholar]
- Fu, D.; Calvo, J.A.; Samson, L.D. Balancing Repair and Tolerance of DNA Damage Caused by Alkylating Agents. Nat. Rev. Cancer 2012, 12, 104–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sedgwick, B.; Bates, P.A.; Paik, J.; Jacobs, S.C.; Lindahl, T. Repair of Alkylated DNA: Recent Advances. DNA Repair Amst. 2007, 6, 429–442. [Google Scholar] [CrossRef]
- Helleday, T.; Petermann, E.; Lundin, C.; Hodgson, B.; Sharma, R.A. DNA Repair Pathways as Targets for Cancer Therapy. Nat. Rev. Cancer 2008, 8, 193–204. [Google Scholar] [CrossRef]
- Gerson, S.L. MGMT: Its Role in Cancer Aetiology and Cancer Therapeutics. Nat. Rev. Cancer 2004, 4, 296–307. [Google Scholar] [CrossRef]
- Albertella, M.R.; Green, C.M.; Lehmann, A.R.; O’Connor, M.J. A Role for Polymerase Eta in the Cellular Tolerance to Cisplatin-Induced Damage. Cancer Res. 2005, 65, 9799–9806. [Google Scholar] [CrossRef] [Green Version]
- Doles, J.; Oliver, T.G.; Cameron, E.R.; Hsu, G.; Jacks, T.; Walker, G.C.; Hemann, M.T. Suppression of Rev3, the Catalytic Subunit of Polz, Sensitizes Drug-Resistant Lung Tumors to Chemotherapy. Proc. Natl. Acad. Sci. USA 2010, 107, 20786–20791. [Google Scholar] [CrossRef] [Green Version]
- Xie, K.; Doles, J.; Hemann, M.T.; Walker, G.C. Error-Prone Translesion Synthesis Mediates Acquired Chemoresistance. Proc. Natl. Acad. Sci. USA 2010, 107, 20792–20797. [Google Scholar] [CrossRef] [Green Version]
- Ma, B.; Zarth, A.T.; Carlson, E.S.; Villalta, P.W.; Upadhyaya, P.; Stepanov, I.; Hecht, S.S. Methyl DNA Phosphate Adduct Formation in Rats Treated Chronically with 4-(Methylnitrosamino)-1-(3-Pyridyl)-1-Butanone and Enantiomers of Its Metabolite 4-(Methylnitrosamino)-1-(3-Pyridyl)-1-Butanol. Chem. Res. Toxicol. 2018, 31, 48–57. [Google Scholar] [CrossRef]
- Jones, G.D.; Le Pla, R.C.; Farmer, P.B. Phosphotriester Adducts (PTEs): DNA’s Overlooked Lesion. Mutagenesis 2010, 25, 3–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beranek, D.T. Distribution of Methyl and Ethyl Adducts Following Alkylation with Monofunctional Alkylating Agents. Mutat. Res. 1990, 231, 11–30. [Google Scholar] [PubMed]
- Ma, B.; Zarth, A.T.; Carlson, E.S.; Villalta, P.W.; Upadhyaya, P.; Stepanov, I.; Hecht, S.S. Identification of More than One Hundred Structurally Unique DNA-Phosphate Adducts Formed during Rat Lung Carcinogenesis by the Tobacco-Specific Nitrosamine 4-(Methylnitrosamino)-1-(3-Pyridyl)-1-Butanone. Carcinogenesis 2017, 39, 232–241. [Google Scholar] [CrossRef] [Green Version]
- Den Engelse, L.; De Graaf, A.; De Brij, R.J.; Menkveld, G.J. O2- and O4-Ethylthymine and the Ethylphosphotriester DTp(Et)DT Are Highly Persistent DNA Modifications in Slowly Dividing Tissues of the Ethylnitrosourea-Treated Rat. Carcinogenesis 1987, 8, 751–757. [Google Scholar] [CrossRef]
- Shooter, K.V.; Slade, T.A. The Stability of Methyl and Ethyl Phosphotriesters in DNA in Vivo. Chem. Biol. Interact. 1977, 19, 353–361. [Google Scholar] [CrossRef]
- Friedberg, E.C.; Aguilera, A.; Gellert, M.; Hanawalt, P.C.; Hays, J.B.; Lehmann, A.R.; Lindahl, T.; Lowndes, N.; Sarasin, A.; Wood, R.D. DNA Repair: From Molecular Mechanism to Human Disease. DNA Repair Amst. 2006, 5, 986–996. [Google Scholar] [CrossRef]
- Tan, Y.; Guo, S.; Wu, J.; Du, H.; Li, L.; You, C.; Wang, Y. DNA Polymerase η Promotes the Transcriptional Bypass of N2-Alkyl-2′-Deoxyguanosine Adducts in Human Cells. J. Am. Chem. Soc. 2021, 143, 16197–16205. [Google Scholar] [CrossRef]
- Wu, J.; Wang, P.; Wang, Y. Cytotoxic and Mutagenic Properties of Alkyl Phosphotriester Lesions in Escherichia Coli Cells. Nucleic Acids Res. 2018, 46, 4013–4021. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Li, L.; Wang, P.; You, C.; Williams, N.L.; Wang, Y. Translesion Synthesis of O4-Alkylthymidine Lesions in Human Cells. Nucleic Acids Res. 2016, 44, 9256–9265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, J.; Du, H.; Li, L.; Price, N.E.; Liu, X.; Wang, Y. The Impact of Minor-Groove N2-Alkyl-2′-Deoxyguanosine Lesions on DNA Replication in Human Cells. ACS Chem. Biol. 2019, 14, 1708–1716. [Google Scholar] [CrossRef]
- You, C.; Wang, Y. Quantitative Measurement of Transcriptional Inhibition and Mutagenesis Induced by Site-Specifically Incorporated DNA Lesions in Vitro and in Vivo. Nat. Protoc. 2015, 10, 1389–1406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, Y.; You, C.; Park, J.; Kim, H.S.; Guo, S.; Schärer, O.D.; Wang, Y. Transcriptional Perturbations of 2,6-Diaminopurine and 2-Aminopurine. ACS Chem. Biol. 2022, 17, 1672–1676. [Google Scholar] [CrossRef]
- Bregeon, D.; Doetsch, P.W. Transcriptional Mutagenesis: Causes and Involvement in Tumour Development. Nat. Rev. Cancer 2011, 11, 218–227. [Google Scholar] [CrossRef] [Green Version]
- Saxowsky, T.T.; Doetsch, P.W. RNA Polymerase Encounters with DNA Damage: Transcription-Coupled Repair or Transcriptional Mutagenesis? Chem. Rev. 2006, 106, 474–488. [Google Scholar] [CrossRef] [PubMed]
- Morreall, J.F.; Petrova, L.; Doetsch, P.W. Transcriptional Mutagenesis and Its Potential Roles in the Etiology of Cancer and Bacterial Antibiotic Resistance. J. Cell. Physiol. 2013, 228, 2257–2261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, J.; Wang, Y. Replication of Pyridyloxobutyl Phosphotriester Lesions in Cells. Chem. Res. Toxicol. 2020, 33, 308–311. [Google Scholar] [CrossRef]
- Wu, J.; Yuan, J.; Price, N.E.; Wang, Y. Ada Protein- and Sequence Context-Dependent Mutagenesis of Alkyl Phosphotriester Lesions in Escherichia Coli Cells. J. Biol. Chem. 2020, 295, 8775–8783. [Google Scholar] [CrossRef] [PubMed]
- Koike, G.; Maki, H.; Takeya, H.; Hayakawa, H.; Sekiguchi, M. Purification, Structure, and Biochemical Properties of Human O6-Methylguanine-DNA Methyltransferase. J. Biol. Chem. 1990, 265, 14754–14762. [Google Scholar] [CrossRef]
- Viswanathan, A.; Doetsch, P.W. Effects of Nonbulky DNA Base Damages on Escherichia Coli RNA Polymerase-Mediated Elongation and Promoter Clearance. J. Biol. Chem. 1998, 273, 21276–21281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saxowsky, T.T.; Meadows, K.L.; Klungland, A.; Doetsch, P.W. 8-Oxoguanine-Mediated Transcriptional Mutagenesis Causes Ras Activation in Mammalian Cells. Proc. Natl. Acad. Sci. USA 2008, 105, 18877–18882. [Google Scholar] [CrossRef] [Green Version]
- Lans, H.; Hoeijmakers, J.H.J.; Vermeulen, W.; Marteijn, J.A. The DNA Damage Response to Transcription Stress. Nat. Rev. Mol. Cell Biol. 2019, 20, 766–784. [Google Scholar] [CrossRef]
- Tornaletti, S.; Maeda, L.S.; Lloyd, D.R.; Reines, D.; Hanawalt, P.C. Effect of Thymine Glycol on Transcription Elongation by T7 RNA Polymerase and Mammalian RNA Polymerase II. J. Biol. Chem. 2001, 276, 45367–45371. [Google Scholar] [CrossRef]
- Charlet-Berguerand, N.; Feuerhahn, S.; Kong, S.E.; Ziserman, H.; Conaway, J.W.; Conaway, R.; Egly, J.M. RNA Polymerase II Bypass of Oxidative DNA Damage Is Regulated by Transcription Elongation Factors. EMBO J. 2006, 25, 5481–5491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clauson, C.L.; Oestreich, K.J.; Austin, J.W.; Doetsch, P.W. Abasic Sites and Strand Breaks in DNA Cause Transcriptional Mutagenesis in Escherichia Coli. Proc. Natl. Acad. Sci. USA 2010, 107, 3657–3662. [Google Scholar] [CrossRef] [Green Version]
- Tornaletti, S.; Reines, D.; Hanawalt, P.C. Structural Characterization of RNA Polymerase II Complexes Arrested by a Cyclobutane Pyrimidine Dimer in the Transcribed Strand of Template DNA. J. Biol. Chem. 1999, 274, 24124–24130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Todd, R.C.; Lippard, S.J. Inhibition of Transcription by Platinum Antitumor Compounds. Metallomics 2009, 1, 280–291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuasa, M.; Masutani, C.; Eki, T.; Hanaoka, F. Genomic Structure, Chromosomal Localization and Identification of Mutations in the Xeroderma Pigmentosum Variant (XPV) Gene. Oncogene 2000, 19, 4721–4728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biertümpfel, C.; Zhao, Y.; Kondo, Y.; Ramón-Maiques, S.; Gregory, M.; Lee, J.Y.; Masutani, C.; Lehmann, A.R.; Hanaoka, F.; Yang, W. Structure and Mechanism of Human DNA Polymerase η. Nature 2010, 465, 1044–1048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, Y.; Egli, M.; Guengerich, F.P. Mechanism of Ribonucleotide Incorporation by Human DNA Polymerase η. J. Biol. Chem. 2016, 291, 3747–3756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, Y.; Egli, M.; Guengerich, F.P. Human DNA Polymerase η Accommodates RNA for Strand Extension. J. Biol. Chem. 2017, 292, 18044–18051. [Google Scholar] [CrossRef] [Green Version]
- Mentegari, E.; Crespan, E.; Bavagnoli, L.; Kissova, M.; Bertoletti, F.; Sabbioneda, S.; Imhof, R.; Sturla, S.J.; Nilforoushan, A.; Hübscher, U.; et al. Ribonucleotide Incorporation by Human DNA Polymerase η Impacts Translesion Synthesis and RNase H2 Activity. Nucleic Acids Res. 2017, 45, 2600–2614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meroni, A.; Nava, G.M.; Bianco, E.; Grasso, L.; Galati, E.; Bosio, M.C.; Delmastro, D.; Muzi-Falconi, M.; Lazzaro, F. RNase H Activities Counteract a Toxic Effect of Polymerase η in Cells Replicating with Depleted DNTP Pools. Nucleic Acids Res. 2019, 47, 4612–4623. [Google Scholar] [CrossRef]
- Gali, V.K.; Balint, E.; Serbyn, N.; Frittmann, O.; Stutz, F.; Unk, I. Translesion Synthesis DNA Polymerase η Exhibits a Specific RNA Extension Activity and a Transcription-Associated Function. Sci. Rep. 2017, 7, 13055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soria, G.; Belluscio, L.; van Cappellen, W.A.; Kanaar, R.; Essers, J.; Gottifredi, V. DNA Damage Induced Pol η Recruitment Takes Place Independently of the Cell Cycle Phase. Cell Cycle 2009, 8, 3340–3348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crespan, E.; Furrer, A.; Rösinger, M.; Bertoletti, F.; Mentegari, E.; Chiapparini, G.; Imhof, R.; Ziegler, N.; Sturla, S.J.; Hübscher, U.; et al. Impact of Ribonucleotide Incorporation by DNA Polymerases β and λ on Oxidative Base Excision Repair. Nat. Commun. 2016, 7, 10805. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Schematic diagrams depicting the SP and RP diastereomers of Me-phosphotriester (PTE) and nPr-PTE lesions in DNA (A) and the CTAB assay system (B).
Figure 1.
Schematic diagrams depicting the SP and RP diastereomers of Me-phosphotriester (PTE) and nPr-PTE lesions in DNA (A) and the CTAB assay system (B).

Figure 2.
Transcriptional bypass of Me-PTE in HEK293T cells and isogenic cells depleted of TLS polymerases. (A–C) Gel image showing the 16 mer (from the competitor genome) and 13 mer (from the control or lesion-containing genome) digestion products formed from the original template strand, where 13 mer TG/TA/TC/TT designate [5′-32P]-labeled standard ODNs 5′-CATGGCMNGCTAT-3′, with ‘MN’ being TT, TG, TC and TA, respectively. (D) The relative bypass efficiencies (RBE) of the two diastereomers of Me-PTE lesions in HEK293T cells and the isogenic cells depleted of TLS polymerases. The data represent mean ± S.D. (n = 3). *, 0.01 < p < 0.05; **, 0.001 < p < 0.01; ***, 0.0001 < p < 0.001. The p values were calculated by using Sidek’s multiple comparisons test.
Figure 2.
Transcriptional bypass of Me-PTE in HEK293T cells and isogenic cells depleted of TLS polymerases. (A–C) Gel image showing the 16 mer (from the competitor genome) and 13 mer (from the control or lesion-containing genome) digestion products formed from the original template strand, where 13 mer TG/TA/TC/TT designate [5′-32P]-labeled standard ODNs 5′-CATGGCMNGCTAT-3′, with ‘MN’ being TT, TG, TC and TA, respectively. (D) The relative bypass efficiencies (RBE) of the two diastereomers of Me-PTE lesions in HEK293T cells and the isogenic cells depleted of TLS polymerases. The data represent mean ± S.D. (n = 3). *, 0.01 < p < 0.05; **, 0.001 < p < 0.01; ***, 0.0001 < p < 0.001. The p values were calculated by using Sidek’s multiple comparisons test.

Figure 3.
Transcriptional bypass of nPr-PTE in HEK293T cells and isogenic cells depleted of TLS polymerases. (A–C) Gel image showing the 16 mer (from the competitor genome) and 13 mer (from the control or lesion-containing genome) digestion products formed from the original template strand, where 13 mer TG/TA/TC/TT designate [5′-32P]-labeled standard ODNs 5′-CATGGCMNGCTAT-3′, with ‘MN’ being TT, TG, TC and TA, respectively. (D) The RBE values of the two diastereomers of nPr-PTE lesions in HEK293T cells and isogenic cells depleted of TLS polymerases. The data represent the means ± S.D. (n = 3). *, 0.01 < p < 0.05; **, 0.001 < p < 0.01; ***, 0.0001 < p < 0.001; ****, p < 0.0001. The p values were calculated by using Sidek’s multiple comparisons test.
Figure 3.
Transcriptional bypass of nPr-PTE in HEK293T cells and isogenic cells depleted of TLS polymerases. (A–C) Gel image showing the 16 mer (from the competitor genome) and 13 mer (from the control or lesion-containing genome) digestion products formed from the original template strand, where 13 mer TG/TA/TC/TT designate [5′-32P]-labeled standard ODNs 5′-CATGGCMNGCTAT-3′, with ‘MN’ being TT, TG, TC and TA, respectively. (D) The RBE values of the two diastereomers of nPr-PTE lesions in HEK293T cells and isogenic cells depleted of TLS polymerases. The data represent the means ± S.D. (n = 3). *, 0.01 < p < 0.05; **, 0.001 < p < 0.01; ***, 0.0001 < p < 0.001; ****, p < 0.0001. The p values were calculated by using Sidek’s multiple comparisons test.

Figure 4.
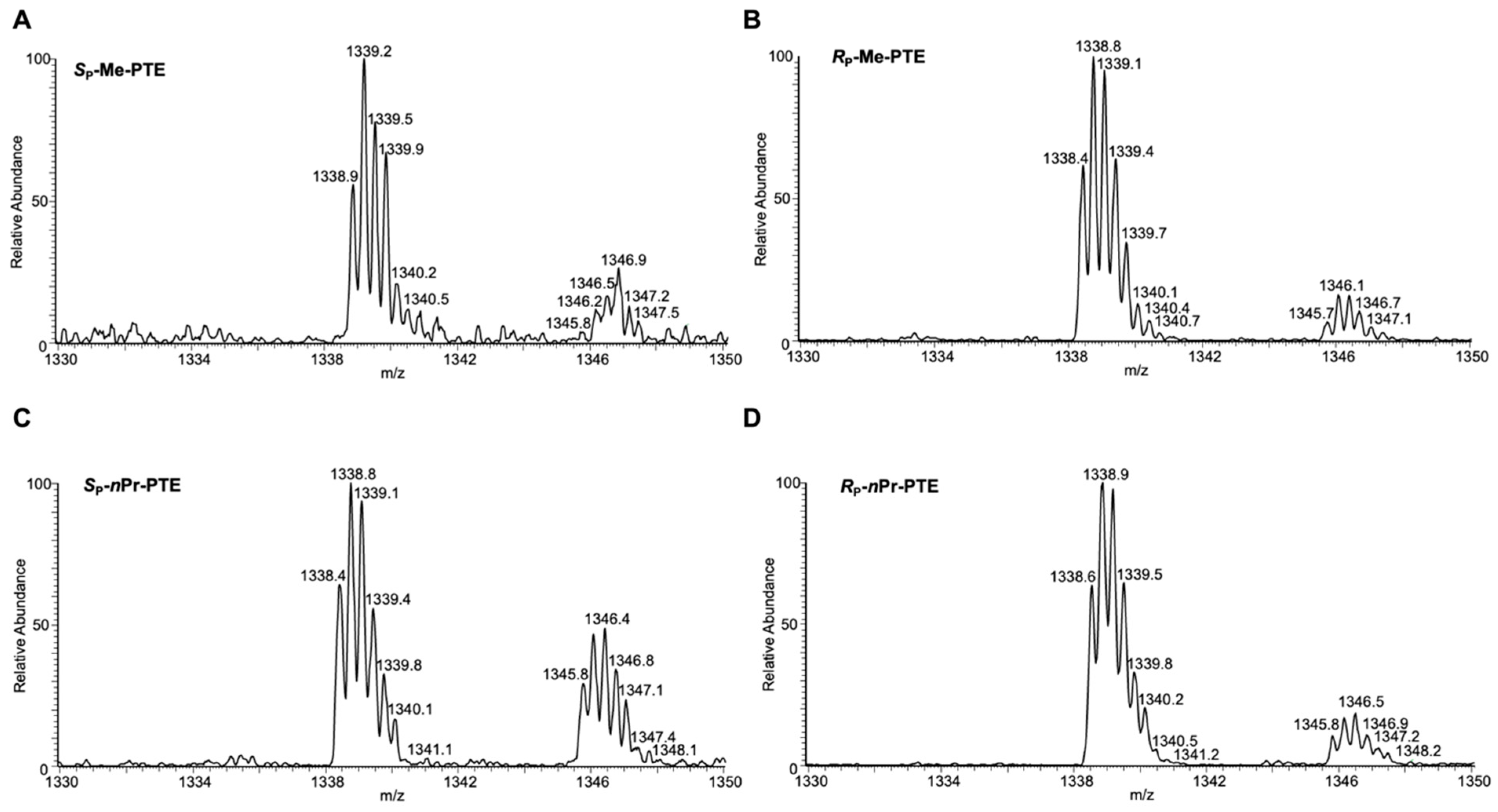
Higher-resolution “zoom-scan” ESI-MS for monitoring the [M-3H]3− ions of the restriction fragments of interest, i.e., 5′-pCATGGCMNGCTAT-3′ (MN = TT, TG, TC and TA) from the transcription of Sp-Me-PTE (A), Rp-Me-PTE (B), Sp-nPr-PTE (C) and Rp-nPr-PTE (D) in HEK293T cells. The isotope cluster at m/z 1339 corresponds to the [M-3H]3− ions of the non-mutagenic product, 5′-pCATGGCTTGCTAT-3′. The isotope cluster at m/z 1346 corresponds to the [M-4H+Na]3− ions of the non-mutagenic product. The calculated m/z values for the putative mutagenic products of TG, TC and TA are 1347, 1333.5 and 1341.5, respectively.
Figure 4.
Higher-resolution “zoom-scan” ESI-MS for monitoring the [M-3H]3− ions of the restriction fragments of interest, i.e., 5′-pCATGGCMNGCTAT-3′ (MN = TT, TG, TC and TA) from the transcription of Sp-Me-PTE (A), Rp-Me-PTE (B), Sp-nPr-PTE (C) and Rp-nPr-PTE (D) in HEK293T cells. The isotope cluster at m/z 1339 corresponds to the [M-3H]3− ions of the non-mutagenic product, 5′-pCATGGCTTGCTAT-3′. The isotope cluster at m/z 1346 corresponds to the [M-4H+Na]3− ions of the non-mutagenic product. The calculated m/z values for the putative mutagenic products of TG, TC and TA are 1347, 1333.5 and 1341.5, respectively.

Figure 5.
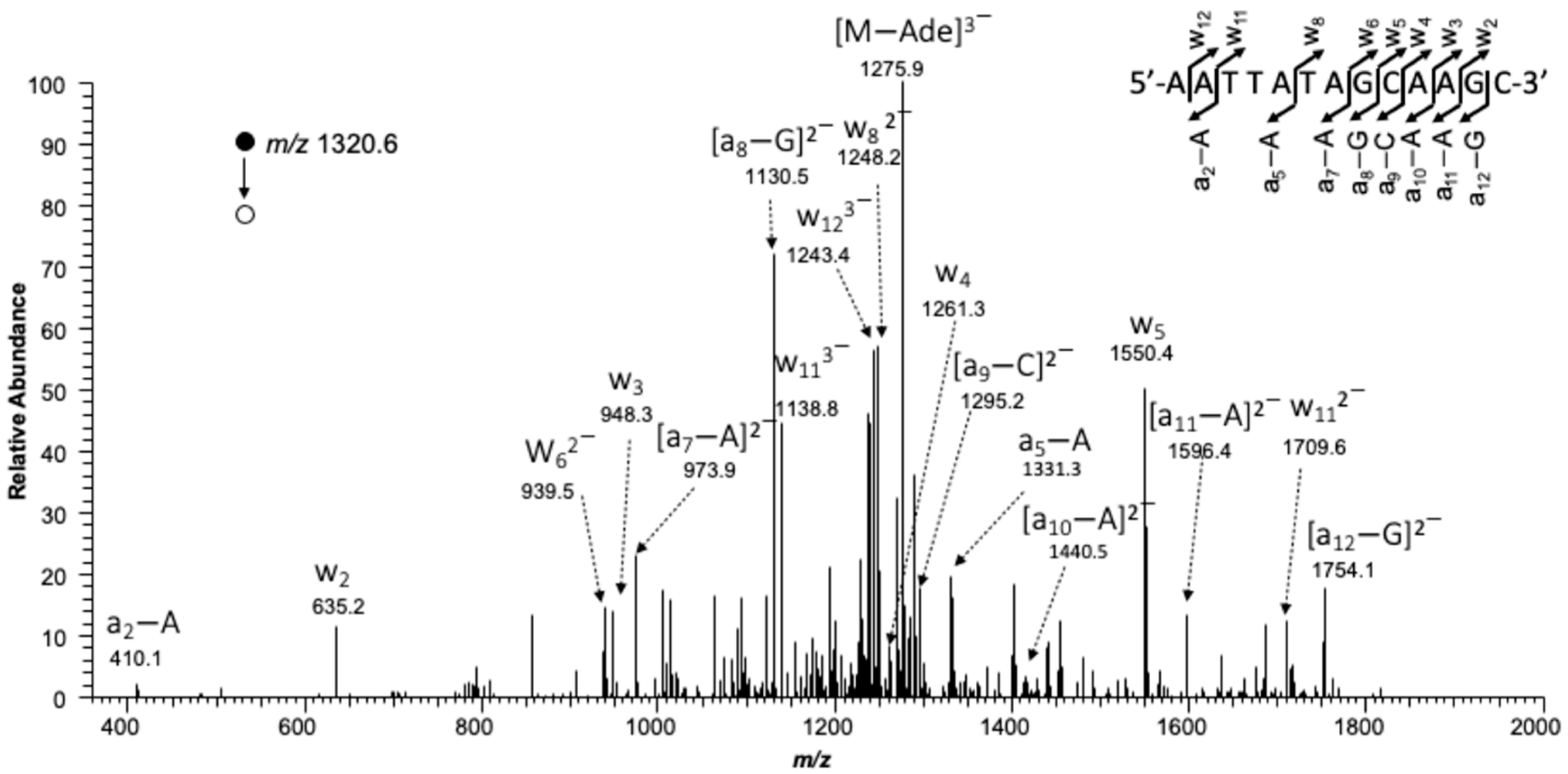
MS/MS for confirming the restriction fragment corresponding to wild-type transcript arising from alkyl-PTE containing substrates.
Figure 5.
MS/MS for confirming the restriction fragment corresponding to wild-type transcript arising from alkyl-PTE containing substrates.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Tan, Y.; Wu, J.; Clabaugh, G.; Li, L.; Du, H.; Wang, Y. Size- and Stereochemistry-Dependent Transcriptional Bypass of DNA Alkyl Phosphotriester Adducts in Mammalian Cells. DNA 2022, 2, 221-230. https://doi.org/10.3390/dna2040016
AMA Style
Tan Y, Wu J, Clabaugh G, Li L, Du H, Wang Y. Size- and Stereochemistry-Dependent Transcriptional Bypass of DNA Alkyl Phosphotriester Adducts in Mammalian Cells. DNA. 2022; 2(4):221-230. https://doi.org/10.3390/dna2040016
Chicago/Turabian StyleTan, Ying, Jiabin Wu, Garrit Clabaugh, Lin Li, Hua Du, and Yinsheng Wang. 2022. "Size- and Stereochemistry-Dependent Transcriptional Bypass of DNA Alkyl Phosphotriester Adducts in Mammalian Cells" DNA 2, no. 4: 221-230. https://doi.org/10.3390/dna2040016