
Origin of the Bunun Indigenous People of Taiwan, a Review of Published Material Using Y-Chromosome and Mitochondrial DNA Gene Systems
Alumni Community, City Campus, University of Technology of Sydney, 15 Broadway, Ultimo, NSW 2007, Australia
†
Retired.
DNA 2022, 2(3), 185-201; https://doi.org/10.3390/dna2030013
Submission received: 3 May 2022
/
Revised: 5 August 2022
/
Accepted: 5 August 2022
/
Published: 1 September 2022
Abstract
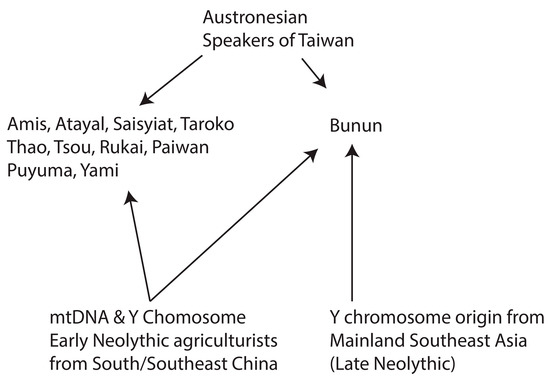
:Anthropological and linguistic studies place the first settlement of Austronesian speaking Taiwanese (AN_Tw) in the mid-Holocene era. However, geneticists have revealed exclusive diversity among the Bunun indigenous people, implying that their ancestral origin needs further study. The mitochondrial DNA (mtDNA) polymorphism of the Bunun shows a homogeneous relationship with other AN_Tw. However, the Y-chromosome polymorphism shows two major haplogroups, O1a2-M50 (60.7%), also seen to a lesser extent among the Northern AN_Tw, and O1b1a1a1a1a1-M88 (37.5%), scarce among all other AN and non-AN groups in Taiwan, but prevailing in Southeast Asia (SEA) and Mainland Southeast Asia (MSEA). While the present-day mtDNA profile of the Bunun typifies the long-term demographic standard for all AN_Tw since the Neolithic era, their Y-chromosome profile suggests an arrival of male settlers in the last two to three millennia from SEA or MSEA, who mixed exclusively with the Bunun indigenous people.
1. Introduction
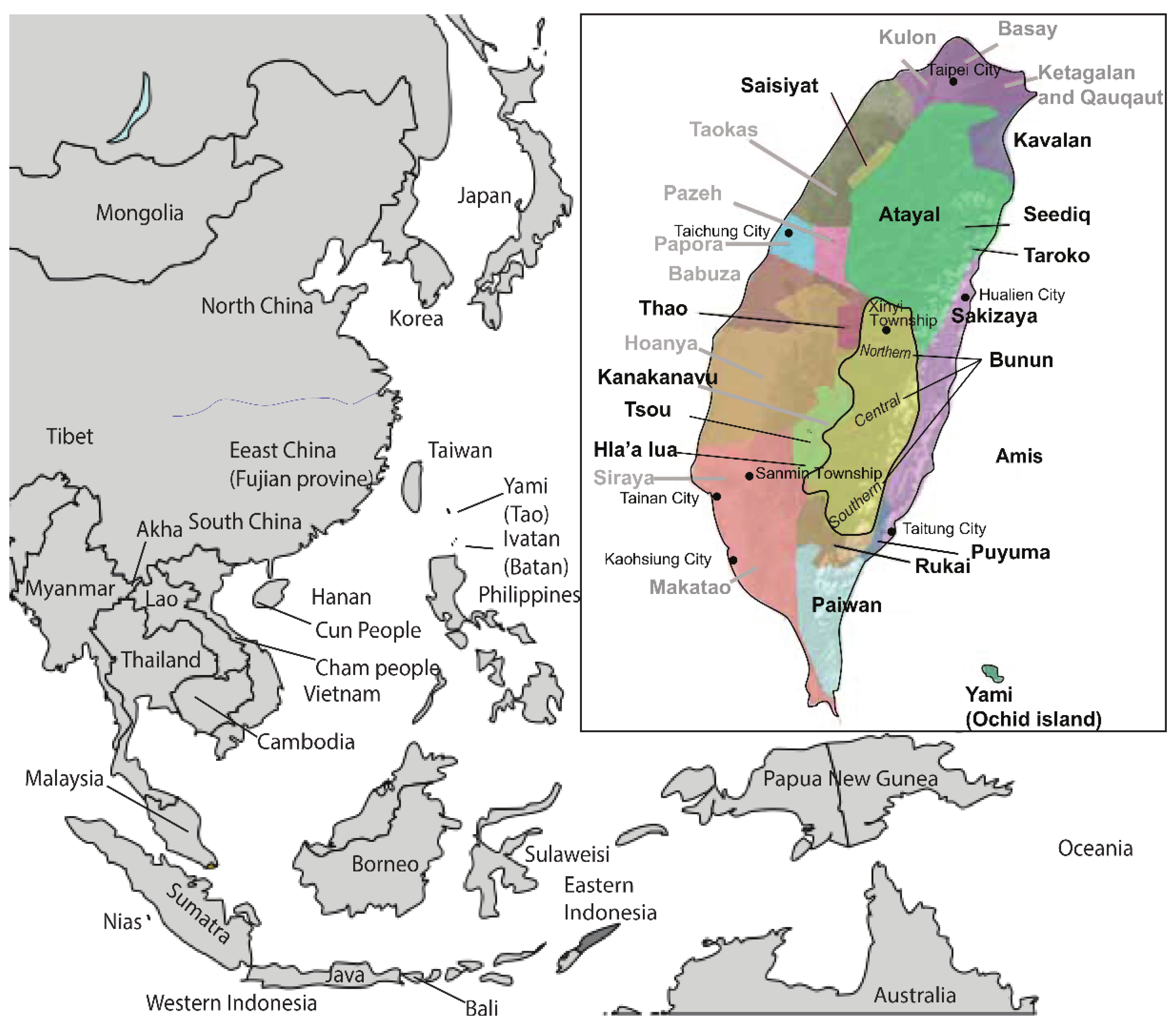
Officially, the Taiwan Austronesian speaking indigenous people (AN_Tw) comprise 16 tribes, who constitute 2.2% of the Taiwan population (approximately 640,000 AN_Tw among 23 million non-Austronesian speaking Taiwanese (NAN_Tw) [1]. In general, the AN_Tw spread in small family units who are widely dispersed on and off the East coast of Taiwan (i.e., Amis, Puyuma, and Yami) and throughout the central mountain ranges (Saisiyat, Atayal, Taroko, Seediq, Bunun, Thao, Tsou, Rukai, and Paiwan). The most recent officially recognized AN_Tw groups are the Hla’alua, Kanakanavu, Kavalan, and the Sakizaya (Figure 1).
To this day, the Babuza, Basay, Hoanya, Ketagalan, Luilang, Makatao, Pazeh/Kaxabu, Papora, Qauqaut, Siraya, Taokas, and Trobiawan groups are not recognized as indigenous people by the central government. They are often referred to as the Taiwan Western Plain tribes or more formally as the Pingpu. Together, the Pingpu represent 0.5% of the Taiwan population and speak Mandarin, since their original languages are extinct.
Ethnologists and archaeologists have distinguished several ancient indigenous cultures in Taiwan. They generally propose that these cultures trace their origin with the Ta-Pen-Kang culture (TPK) in the mid-Holocene period (7000 to 5000 kya) [2,3,4]. However, the discovery of Paleolithic artifacts of a tool industry at the “Changbin culture” site (28 kya) near Taitung in Southeast Taiwan, and ancient human remains (bones and teeth of the Tso-Chen Man) lead archaeologists to propose that Taiwan entered human history in the Paleolithic era and that these people were later replaced by settlers of the Ta-Pen-Kang culture [5,6].
Furthermore, linguistic studies trace all AN_Tw languages back to a common ancestral language, the proto-Austronesian language (PAN), whose filiations are shared by extant groups of Mainland Southeast Asia (MSEA), Peninsular Asia (Malaysia), Polynesia, and Madagascar [7].
The Bunun, with approximately 51,000 members represent approximately 12% of the indigenous population (or 0.23% of Taiwan population), and are the fourth-largest indigenous group in Taiwan [1,8]. In terms of family, kinship and, descent principles, the Bunun are classified, along the Tsou and Saisiyat, as a patrilineal clannish society [9]. Similar to most AN_Tw, they live in family groups, often in mono-ethnic villages that are widely dispersed in the central mountain range. In the past, their staple food was foxtail millet, and females were responsible for gathering and preparing food. On the other hand, males were very individualist, responsible for a slash and burn agriculture, hunting, and are known for having been head-hunter and fierce warriors [10].
The beginning of Taiwan’s recorded history started with the arrival of the Dutch, in 1624 who established a colonial regime to expand their deerskin trade, and the Spanish later who established a base in Northern Taiwan [11,12]. Along with these new settlers was the entrance of missionaries, who translated the Bible into Bunun language and indigenized the Christian conceptual universe. By 1940, many AN_Tw were converted to Christianity. The Bunun lost many traits of their cultural heritage, and as a result, revived present-day cultural features have become poorly supported and their genuineness debatable.
Under Japanese rules, from 1895 to 1945, for security reasons against the slash and burn leading to deforestation, 62% of Bunun people were forcibly resettled in villages of the Western lowlands. Traditional hunting was also prohibited, and despite Bunun Chief Raho Ari who embarked on guerrilla warfare against the Japanese authorities, rice cultivation was imposed as the preferred alternative [10]. In October 1945, the Chinese Nationalist Kuomintang entered Taiwan and encouraged the use of standard Mandarin. This generated drastic changes in their culture. The family set up, that the Bunun were accustomed to, ended and led to a mode of living in individual units. However, the Bunun people endlessly pursue the revival of their cultural heritage. They are best known for the ear-shooting ritual that recreates their ancient practice for young man passage to adulthood, and for the “Pasibutbut”, a singing prayer using an uninterrupted polyphonic song performed by eight singers still performed during the annual Millet Harvest Festival [13]. Linguistic factors are well characterized and well identifiable from other groups. Presently, the Bunun have five distinct communities whose extant dialects have been divided into three main branches [14]: The Northern Bunun (Takituduh and Takibaka), the Central Bunun (Takbanuaz and Takivatan also spoken in Nantou and Hualien counties), and the dominant Southern Bunun dialect (Isbukun). Although many Bunun under the age of 30 today are monolingual Mandarin speakers [15], the revival of their strong ethnic heritage by a large number of Bunun individuals is using Isbukun as language re-establishment [9].
Genetic studies aiming at characterizing the genetic heritage of the Taiwanese started in 1994 with Umetsu et al. [16]. The authors analyzed the polymorphisms of complement components I and C1R in nine Aboriginal Taiwanese populations, and for the first time, suggested long-term genetic and geographic isolation. Within a short time, other genetic studies within the Taiwanese Han and indigenous populations followed. The ABO and Rhesus blood groups [17], the genetic polymorphisms of the promoter region of dopamine D2 receptor and dopamine transporter [18], and a comprehensive study of six Alu insertions [19], contributed to the genetic profile of the Han and Indigenous Taiwanese.
Furthermore, a study using the Histocompatibility Leucocyte Antigens (HLA-A, -B and -DRB1), a highly polymorphic diploid gene system on chromosome 6, showed that Bunun was a member of the genetic cluster composed uniquely of AN_Tw, which separated clearly from NAN_Tw and other east Asian groups [20,21,22].
Other studies, using the uni-parental gene system [22,23,24,25,26] showed that, except for the moderate prevalence among the Bunun of mtDNA haplogroups B4b1a2, B5a2a2b, F4b1, and E1a1a, the Bunun appeared as a group maternally closer to the Northern AN_Tw. Conversely, the low Y-chromosome diversity and the high prevalence of Y-chromosome haplogroup O1a2-M50 and O1b1a1a1a1a1-M8 make the Bunun a paternally distinct outlier to all other Taiwanese groups. However, these characteristics have been attributed to founder effect, bottleneck, and long isolation. Most studies divided Taiwan into two groups, the Han Taiwanese and the Taiwan Indigenous people, and rarely [22,27,28] concentrated on tracing the genetic heritage of a single group. As a result, the genetic heritage of Bunun is still believed to be identical to the other indigenous groups in Taiwan and remains poorly characterized.
The analysis proposed here will combine evidences from two uni-parental gene systems, the non-recombining Y-chromosome (NRY) and the mitochondrial DNA (mtDNA) to compare the genetic structure of the Bunun with other Formosan speaking people, and other groups of East Asia and Island Southeast Asia (ISEA). While the mtDNA findings support previous HLA findings, the extant paternal profile suggests that the Bunun males likely reached Taiwan later in the mid-Neolithic separately from other Austronesian speaking groups.
2. Results and Discussion
2.1. MtDNA Genetic Diversity and Structure
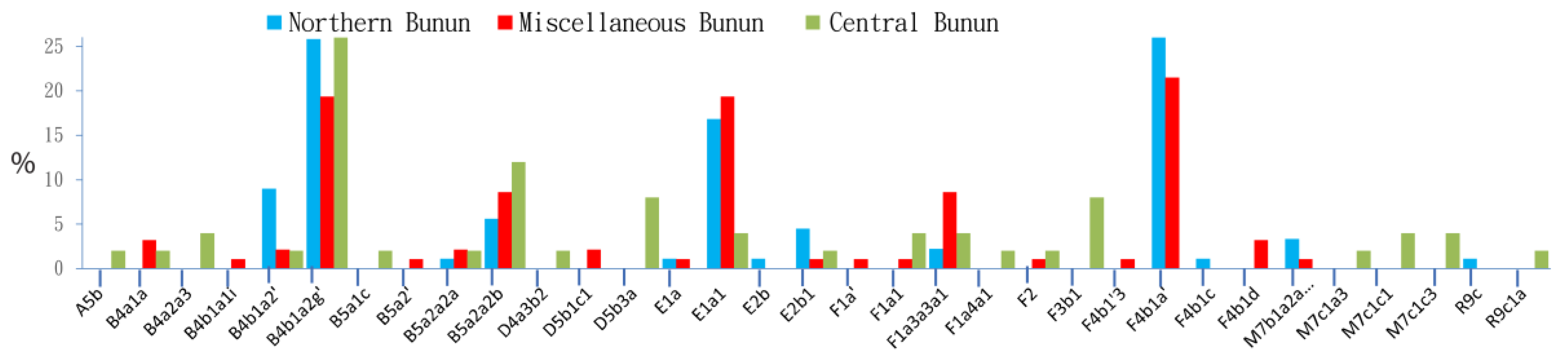
For this analysis, 235 Northern Bunun partial mtDNA sequences comprising segments of the HVS-I and coding regions were compared to 4423 mtDNA sequences from population groups of SEA, MSEA, ISEA, and Taiwan (Table 1, Supplementary Text S1). Using haplogroup assignments according to mtDNA Haplotree-Family Tree 17 [29], 699 different haplogroups were found. Compared to other ethnic groups of East Asia, SEA, and MSEA, the mtDNA polymorphism (the number of haplogroups) seen in each of the three Bunun groups (Northern, Central, and Miscellaneous) appeared to be reduced, but was in range with other indigenous groups of Taiwan. The Miscellaneous Bunun, a self-declared group collected in Taitung city [30], showed 20 haplogroups (Supplementary Text S1 and Table S1) from which only six were exclusive to AN_Tw, and fourteen were of Han origin. However, the table shows that 60.36% are shared with AN_Tw and 10.1% are shared with Han. This suggests that the remaining 20.54% is exclusive to the miscellaneous group or represents a mixture with a group other than the putative parents. This pattern is similar for the Northern Bunun, whereas the Central Bunun appears to be the closest group to the AN_Tw putative parental population.
Pairwise comparisons (Figure 2) between the Northern Bunun and the Miscellaneous Bunun group did not show significant differences (p > 0.3). However, these two groups showed striking differences from the Central Bunun group (p < 0.03) lacking mtDNA subtypes of haplogroup F4b1 and showed a lower prevalence of haplogroup E. On the contrary, haplogroups B5a1c, D5b3a, F3b1, and M7c1 were not seen in the Northern and Miscellaneous Bunun groups, indicating that the genetic history of the Northern and Miscellaneous groups may be different from the Central Bunun.
The genetic diversity of mtDNA haplogroups in the Northern Bunun (h = 0.99) was high, but in range with other indigenous groups in Taiwan (0.989 to 1.00) (Supplementary Table S1). Similarly, the nucleotide diversities between groups were not significantly different (π ~ 0.044) (data not shown).
The three Bunun groups exhibited multimodal mismatch distribution patterns (Supplementary Figure S1) with the mean number of nucleotide-pairwise-differences (MNND) between 12.5 and 14 as well as significant “sum-of-square differences” (SSD, p ≤ 0.05) and raggedness values (p < 0.01). These results indicate a long stationary maternal population history [51]. Furthermore, the Tajima D values of the three Bunun groups were not significant (Supplementary Figure S1d), suggesting the groups were close to or under the neutrality expectation. However, a small Tajima D difference was observed between the Northern Bunun (0.3; p = 0.7) and the Central Bunun (−1.28; p = 0.086). This suggests a minor expansion event of the Central Bunun or possibly gene flow from neighboring groups. This observation is also supported in the Mismatch Distribution Analysis by the presence of two short peaks (left in Supplementary Figure S1C), possibly due to the same sampling size or the result of the admixture. This is not surprising since the Central Bunun group [33], similar to the miscellaneous group [38], were self-declared Bunun indigenous individuals (collected in the south of Taiwan in the Kaohsiung and Taitung counties, respectively).
2.1.1. MtDNA Polymorphism
When using only major mtDNA haplogroups, the AN_Tw can be divided into three homogeneous groups, Northern, Central, and Southern group; however, advances in the sequencing techniques have allowed for the determination of discrete differences within the groups [33,49,52]. Some of these differences can be seen throughout ISEA, sometimes at higher frequency, suggesting drift, bottleneck, and/or long isolation, and reflecting the complexity encountered when linking genetics, languages, and demography throughout ISEA.
In this analysis, the Northern Bunun profile is limited to sub-lineages of four mtDNA para-haplogroups, namely B4b1, B5a2, F4b1, and E1a1a (Figure 2, Supplementary Table S1), that represent 80% of the Northern Bunun genetic structure, a common characteristic of the Northern and Central AN_Tw. Interestingly, these haplogroups do not represent clades that participated in the “out-of-Taiwan” [53].
The most common subtype of B4b1 in Taiwan, B4b1a2g, is seen in the Northern Bunun and Thao groups (24% and 27%, respectively) and has an estimated expansion age of 4.2 ± 3.5 kya [20]. Excluding B4b1a2a, which is principally seen in SEA and MSEA, all other B4b1 subtypes in Taiwan are subclades of B4b1a2. They prevail in the Northern tribes (Saisiyat, Atayal, and Taroko) and groups of the central mountain range, Bunun, Tsou, and Thao [22,33]. They find their origin in SEA and north MSEA, likely along the Mekong river since they are absent from Vietnam and the Cham Austronesian speaking groups of Vietnam [22,33,45,46,54]. Haplogroup B4b1a2g is the most common mtDNA B subtype in Taiwan. It prevails in the Northern Bunun and Thao groups (25% and 27%, respectively). It has an estimated expansion age of 4.2 ± 3.5 kya and most likely expanded exclusively in Taiwan [22].
Haplogroup B5a is common in South and Southeast China. Subtypes of B5a are seen in the three Bunun groups and the Tsou (Supplementary Table S1). They are primarily represented by B5a2a2b, at a lower frequency by B5a2a2a, and are scarce out of the central mountain range. The type seen among Bunun is B5a2a2b1 and likely evolved in Taiwan among the Bunun (the Tsou) 4.3 ± 2.2 kya.
Haplogroup F4b1 is determined by the presence of nucleotide position np 10,097 and 513. It is a common characteristic of the Northern AN_Tw (Supplementary Table S1). With an expansion estimate of 4.3 kya (CI 1.3–7.3 kya), it is found to be a daughter haplogroup of F4b, which is common in Southeast Asia, and represents a mid-Holocene dispersal from this region in Taiwan [22,55].
Haplogroup E1a1a1 is a descendent of haplogroup E1. Commonly seen in the Saisiyat, the Tsou, and the Bunun tribes, its ancestral lineages trace back to haplogroup pre-E1, first described in the Liangdao man [33,47]. All E-lineages are branches of haplogroup M9 seen along the east coast of China and Southeast Asia [33,47]. Since no E-lineages have been reported in Mainland Asia, haplogroup E likely evolved in Island Southeast Asia. With an average founder age of 6.25 ± 1.6 kya, haplogroup E1a1a1 relates to the early dispersal of the Austronesian people to Taiwan.
Finally, lineages B4b1a2, B5a2a2b, and F4b1 prevail in Northern Bunun. Their molecular variation was used to estimate the time of gene flow events in that group. Convergence estimates showed 4.2 kya (CI 0.6–7.8 kya), 4.3 kya (CI 1.3–7.3 kya), and 4.3 kya (CI 1.2–7.5 kya), respectively. While their confidence interval encompasses the time of the Austronesian expansion, it is also likely that these lineages represent later gene flow events of the past four millennia.
Further mtDNA haplogroups with considerably lower frequencies were identified. These were R9c, M7b1a2a, and F4b1c in Northern Bunun, A5a, B4a2a3, B5a1c, D5b3a, F3b1, and M7c1c in Central Bunun, and D5b1c and F4b1a in Miscellaneous Bunun. Except for A5a, a fingerprint of Han admixture, these haplogroups were all associated with the profile of Taiwan indigenous people. Their presence elsewhere was limited to the Philippines and Western Indonesia (Supplementary Table S1). Their low frequencies were likely the result of drift.
2.1.2. Principal Component Analysis
As expected from the pairwise comparison (Figure 2), the principal component (Figure 3) characterizes the Northern Bunun, the Thao, and the Tsou as outgroups. Moreover, the Principal Component Analysis (PCA) plot separates the Northern indigenous groups of Taiwan (Atayal, Taroko, and Saisiyat) seen in the lower right from the Bunun, the Thao, and the Tsou. Furthermore, the plot separates the Southern indigenous groups in the left lower pane from MSEA, the Philippines, Indonesia, and the Taiwan Han top seen on the top of the lower left quadrant.
2.2. Non-Recombining Y-Chromosome
2.2.1. Y-SNP
Y-SNP typing revealed only two Non-Recombining Y-Chromosome (NRY) haplogroups among 56 Northern Bunun males (Table 2). Almost 61% belonged to haplogroup O1a2-M50 and 37.5% to haplogroup O1b1a1a1a1a1-M88 (Supplementary Table S1). The frequency of the last haplogroup is the highest seen in Taiwan and Western ISEA. One single individual was bearing haplogroup p-M45. Another individual bearing the same haplogroup was seen among the Amis. This presence of P-M45 in Taiwan is interesting since it was associated with the Negrito groups of south Luzon in the Philippines (Aeta from Santa Juliana and Agta from Umiray) [56], and its ancestry has been traced back to the upper Paleolithic [57].
Haplogroup O1a2-M50 seen in the Northern Bunun is a subclade of O1a-M119 [58]. It was reported in Cambodia [59], and a few Daic-speaking groups of Southeast Asia [32,60]. Its coalescence is estimated to be 15.8 kya (CI 17.5–14.1 kya) [61,62], but its origin in SEA is not well defined. The expansion of O1a2-M50 likely took place in the South of Taiwan, where it prevails at approximately 16.7%. Its presence in any insular groups out of Taiwan represents an out-of-Taiwan signature. From Taiwan, O1a2-M50 reached the admiralties islands (17%). In the south, it expanded to the Philippines, Borneo, Java, the South Nias Island off the coast of Sumatra (60% to 80%), Malaysia, and finally Madagascar [60,63,64,65,66].
Haplogroup O1b1a1a1a1a1-M88/M111 (previously named O2a1-M88/M111) (Supplementary Table S1) is a subclade of O1b1a1a-M95 (formerly referred to as O2a-M95), which is common in South China and MSEA. According to Waas and Urasin [61,62], O1b1a1a1a1a1-M88/M111 formed 8.6 kya, and occurs at high frequency in SEA [67] and MSEA, particularly in Northern Thailand among Htin and Lawa groups, Lao, North Cambodia, and the De’ang group of the Yunnan province of China [58,68,69]. Except for the Northern Bunun (37.5%), O1b1a1a1a1a1-M88 is scarcely seen elsewhere in Taiwan. Similarly, it has a low frequency in the Philippines and Western Indonesia (<4%) [25,56,70]. On the other hand, the concurrent occurrence of O1b1a1a1a1a1-M88 and O1a2-M50 has been reported in some Tai-Kadai and Mon-Khmer speaking groups of MSEA [32,71], in Borneo [72], in Madagascar [63,64], but rarely elsewhere.
2.2.2. Y-STR Haplotype Networks
The Y-STR haplotype networks of O1a2-M50 and O1b1a1a1a1a1-M88 were constructed to evaluate their early dispersal (Figure 4). However, only the Northern Bunun NRY data were available. All Bunun O1a2-M50 haplotypes clumped within shallow clusters on the outskirt of the network away from the ancestral core. Genetic studies have shown [60] that Taiwan is the most likely region of expansion of O1a2-M50 and it is now generally accepted by linguists, archaeologists, and geneticists [2,60,73], as well as O1a2-M50 is a male signature of the Austronesian expansion. Here, it appears that the absence of NRY haplogroup O1a1-P203 and the high prevalence of O1a2-M50 in Bunun are likely the result of drift. Similarly, the exclusive occurrence of haplogroup O1b1a1a1a1a1-M88 in the Northern Bunun (in purple on the right network of Figure 4) showed a shallow structure, supporting a paternal founding or drift event and isolation. Moreover, we notice that the haplogroup pair (O1a2-M50 and O1b1a1a1a1a1-M88) is seen along the Taiwan-Philippines-Borneo-Sumatra pathway, and can be traced further toward Madagascar and the Comoros islands [63,64,74,75]. While this distribution may suggest that O1a2-M50 and O1b1a1a1a1a1-M88 dispersed as a pair from Taiwan, the low Bunun maternal contribution to ISEA and the exclusive presence of O1b1a1a1a1a1-M88 in the Northern Bunun suggest that O1b1a1a1a1a1-M88 reached Taiwan directly from SEA and that the dispersal of O1a2-M50 from Taiwan and O1b1a1a1a1a1-M88 coming from MSEA merged in Indonesia.
The Y-STR variation of haplogroups O1a2-M50 and O1b1a1a1a1a1-M88 was then calculated based on the seven most used Y-STR loci in the NRY scientific literature. Corroborating the ISSOG dating [61,62] O1a2-M50 and O1b1a1a1a1a1-M88 coalesce at approximately ~11.8 + 4.1 kya and 16.7 + 2.7 kya, respectively, suggested a late Paleolithic Southeast Asian at the beginning of their expansions [59,76]. Furthermore, estimation of the Y-STR variation exclusive to the Northern Bunun provided a TMRCA of 2.4 ± 1.6 kya for O1a2-M50 and 4.1 ± 3.1 kya for O1b1a1a1a1a1-M88. These results contrast with the SSOG TMRCAs obtained for the whole of Asia Oceania using the software Yfull, version 8.04.00 (i.e., 5.1 and 5.6 kya, respectively) [61,62,77], and suggest a separate migration from MSEA and later expansion in Taiwan among the Bunun Indigenous people.
2.2.3. Principal Component Analysis from NRY Haplogroups
The two dimensions of the principal component analysis (PCA) using NRY haplogroup frequencies represent 55.1% of the polymorphism variation (Figure 5). It separated the Northern and Southern AN_Tw groups (red dots) from each other. The southern AN_Tw group shows more affinity with Mainland Southeast Asia (yellow squares) and suggests admixture with Taiwan plain dwellers Pazeh and Siraya groups (light blue dots). The Northern Bunun groups are equidistant from MSEA and Western Indonesia [25,32].
2.2.4. TreeMix
Finally, a Maximum-Likelihood phylogram was constructed using the TreeMix package [78] to infer mtDNA and NRY gene-flow events (Figure 6), which contributed to the genetic history of Bunun.
In concordance with the previous study [79], the mtDNA phylogram (left on Figure 6) inferred gene flow to the Northern and Central tribes (Saisiyat, Taroko, and Bunun), most likely associated with the E1a clade from the Philippines (Supplementary Table S1). Similarly, gene flow associated with F4b1a was seen between Saisiyat, Atayal, Taroko, Thao, and Bunun. Finally, the mtDNA phylogram showed a series of relationships restricted to the Central AN_Tw, Thao, Tsou, and Bunun, likely associated with the high prevalence of subtypes of B4b1a2 in these groups. These results indicate gene flow between Central and Northern AN_Tw and support the findings of the PCA (Figure 3).
Figure 6 also shows results for the NRY-STR dataset (right). Taiwan samples were analyzed along with ISEA, MSEA, Oceania, and Madagascar populations [63,64]. For clarity, the arrows in Figure 6 only show the gene flow corresponding to migration weight, which is greater than 0.5. Most of the gene flow associated with Bunun finds its origin in MSEA/SEA, here represented by the heterogeneous ethnic groups of Lao [81] and primarily indicating the origin of haplogroup O1b1a1a1a1a1-M88. The coalescence age estimate of O1b1a1a1a1a1-M88 in Bunun was ~2.5 kya (95% CI 0–5 kya) (Figure 4). Furthermore, the highest occurrence of haplogroup O1a2-M50 in Taiwan is found in Bunun (60%). One must note that O1a2-M50 is also seen among the non-Austronesian speakers of Taiwan (TwH) and shows signs of local expansion.
Comparisons of the Bunun mtDNA and Y-chromosome diversity to other ethnic groups in Taiwan show that women moved between the groups. Conversely, the almost exclusive NRY profile of the Bunun suggests that 37.5% of the men had a phylogeographic history distinct from other AN_Tw. Most Taiwan indigenous groups are patrilineal. Patrilocality might have been the principal social element that maintained the bearers of O1b1a1a1a1a1-M88 within the Bunun community until the present day. Furthermore, genetic variation in the Bunun shows high within-group diversity for the mtDNA (Supplementary Table S1) and a low Y-chromosome diversity (Table 2) with large between-group distances [25], which is in range with a patrilocal residence expected pattern. Finally, the mtDNA gene flow detected by TreeMix (Figure 6) was principally restricted to the gene flow between Taiwan tribes, whereas the Y-chromosome gene flow covered remote geographic regions. This suggests that the male dispersal pattern in the past covered longer geographical distances compared to the female dispersal pattern.
2.3. Conclusions
The Bunun indigenous people hold significant cultural, anthropological, historical, and linguistic diversity, which differentiates them from other groups in Taiwan. While previous HLA and mitochondrial DNA studies posited the Bunun as a group distinct to the Southern Taiwan indigenous groups, their Y-Chromosome profile showed a mixture of ancestral origin. First, haplogroup O1a2-M50, commonly seen among other AN_Tw and Southeast Asia, showed evidence of expansion exclusive to the Bunun (at 1.6 to 2.4 kya). Second, haplogroup O1b1a1a1a1a1-M88 (O2a1a-M88), frequently seen among Daic and Hmong-Mien populations [32,82,83] of Southeast Asia and Mainland Southeast Asia, was primarily seen in the Bunun group and scarcely elsewhere in Taiwan. This suggests a mid-Neolithic (1.6 to 4.1 kya) intrusive migration event of men from Mainland Southeast Asia (Figure 4) into the Bunun groups. This migration event contributed to the modern genetic pool of the Bunun. In this case, it is likely that the strong cultural practices of the Bunun, such as animism, their strict sexual division of labor, hunting, cultivation, language, and social prestige were placed aside, even replaced by those of the newcomers. However, it had a significant motivating impact on the Bunun of today to revive their initial Austronesian heritage. Moreover, this uni-parental genetic review allowed for the determination of the dual origin of Bunun people.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/dna2030013/s1, Supplementary Figure S1: Mitochondrial Mismatch Distribution Analysis (MMDA) of three Bunun groups. Left: (a) Northern Bunun, (b) Miscellaneous Bunun, (c) Central Bunun. (d) SSD: Sum of Squared Deviation test using Arlequin 3.5.2.2. Groups a, b and c exhibited a multimodal mismatch distribution pattern and an expected distribution (red line) that did not fit the sudden population expansion model (grey line) as it is expected with Tajima D p values > 0.5 and all SSD p values p ≤ 0.05, Supplementary Table S1: Polymorphisms of mtDNA exclusive to the Northern Bunun and comparison with other populations, Supplementary Text S1: Materials and Methods. References [22,24,25,27,29,31,33,34,37,38,45,48,50,51,56,63,64,66,68,75,78,80,84,85,86,87,88,89,90,91,92,93] are cited in the supplementary materials.
Funding
This research received no funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
This study is a Review of published material; the data presented are available from articles described in Table 1.
Acknowledgments
The author would like to thank all the authors listed in Table 1, who made their material available through publication. The author is also grateful to Charmaine Greenwell for her kind proofreading of the manuscript.
Conflicts of Interest
The author declares no conflict of interest.
References
- MOI. Department of Statistics, Ministry of the Interior, Monthly Bulletin of Interior, Taiwan 2016 Taipei, Taiwan. Available online: https://www.moi.gov.tw (accessed on 17 July 2022).
- Bellwood, P. The Origin and Dispersals and Dispersal of Agricultural Communities in Southest Asia. In Southeast Asia: From Prehistory to History; Glover, I., Bellwood, P., Eds.; Routledge: Abingdon, UK, 2004; pp. 21–40. [Google Scholar]
- Liu, L.; Wang, J.; Levin, M.J.; Sinnott-Amstrong, N.; Zhao, H.; Zhao, Y.; Shao, J.; Di, N.; Zhang, T. The origins of specialized pottery and diverse alcohol fermentation techniques in Early Neolithic China. Proc. Natl. Acad. Sci. USA. 2019, 116, 12767–12774. [Google Scholar] [CrossRef]
- Tsang, C.-H. Five Thousand Years of Taiwan’s Past Brought to Light by Rescue Archaeology. 2015. Available online: http://www.chinesearchaeology.net/en/Special_Events/dierjieshanghailuntan/2015/1222/52517.html (accessed on 7 July 2022).
- Hung, H.; Nguyen, K.D.; Bellwood, P.; Carson, M.T. Coastal Connectivity: Long-Term Trading Networks Across the South China Sea. J. Isl. Coast. Archaeol. 2013, 8, 384–404. [Google Scholar] [CrossRef]
- Liu, Y.C.; Wang, S. Encountering the wider world before the transition to history: Chinese ceramics in proto-historic Taiwan (tenth through sixteenth centuries). In Historical Archaeology of Early Modern Colonialism in Asia-Pacific: The Southwest Pacific and Oceanian Regions; Berrocal, M.C., Tsang, C., Eds.; University Press of Florida: Gainesville, FL, USA, 2017; pp. 270–391. [Google Scholar]
- Ross, M. Proto Austronesian verbal morphology: A reappraisal. In Austronesian Historical Linguistics and Culture History: A Festschrift for Robert Blust; Adelaar, A.P.A., Ed.; Pacific Linguistics: Canberra, Australia, 2009; pp. 295–326. [Google Scholar]
- DGBAS—Directorate General of Budget, Accounting and Statistics, Executive Yuan, R.O.C. National Statistics, (Taiwan). Preliminary Statistical Analysis Report of 2000 Population and Housing Census Archived 12 March 2007 at the Wayback Machine; Excerpted from Table 28: Indigenous Population Distribution in Taiwan-Fukien Area. 2007. Available online: https://eng.dgbas.gov.tw/ (accessed on 7 June 2022).
- Chiang, B. Customary Laws, Kinship and Beyond: A Critical Review of the cultural Anthropological studies of the Austronesian People in Taiwan: In Austronesian Taiwan: Linguistic, History, Ethnology, and Prehistory; Blundel, D., Ed.; The Phoebe Museum of Anthropology: Berkeley, CA, USA, 2002; pp. 201–245. [Google Scholar]
- Huang, Y.-K. The “Great Men” model among the Bunun of Taiwan. In Austronesian Studies Relating to Taiwan; Paul, L.J., Tsang, C., Huang, Y., Ho, D., Tseng, C., Eds.; Academica Sinica: Taipei, Taiwan, 1995; pp. 59–107. [Google Scholar]
- Jacobs, J.B. Taiwan’s Colonial Experiences and the Development of Ethnic Identities: Some Hypotheses; Taiwan in Comparative Perspective, Asia Research Centre, London School of Economics and Political Science: London, UK, 2014; Volume 5, pp. 47–59. ISSN 1752–7732. [Google Scholar]
- Nakamura, T. Household registration of indigenous villages during the Dutch occupation. In Taiwan History during the Dutch Occupation; Wu, T.-M., Wong, K.-I., Shu, S.-Y., Eds.; National Library of Taiwan; Academia Sinica: Taipei, Taiwan, 2002; Volume 11. (In Chinese) [Google Scholar]
- Brown, S.; Savage, P.E.; Ko, A.M.-S.; Stoneking, M.; Ko, Y.-C.; Loo, J.-H.; Trejaut, J.A. Correlations in the population structure of music, genes and language. Proc. Biol. Sci. 2014, 281, 20132072. [Google Scholar] [CrossRef] [PubMed]
- Li, P.J.-K. Bulletin of the Institute of History and Philology. A comparative study of Bunun dialects, Taiwan. Acad. Sin. 1988, 59, 479–508. [Google Scholar]
- De Busser, R.L.J. Towards a Grammar of Takivatan Bunun. Ph.D. Thesis, La Trobe University, Bundoora, Victoria, Australia, 2009. [Google Scholar]
- Umetsu, K.; Yuasa, I.; Suzuki, T.; Sun, C.S.; Pan, I.H.; Ishida, T.; Saitou, N.; Horai, S. Polymorphisms of complement component I and C1R subcomponent of C1 in nine aboriginal Taiwanese populations. Hum. Biol. 1994, 66, 339–348. [Google Scholar]
- Lin, M. Blood groups and transfusion medicine in Taiwan. J. Formos. Med. Assoc. 1997, 96, 933–942. [Google Scholar] [PubMed]
- Chen, W.J.; Chen, C.H.; Huang, J.; Hsu, Y.P.; Seow, S.V.; Chen, C.C.; Cheng, A.T. Genetic polymorphisms of the promoter region of dopamine D2 receptor and dopamine transporter genes and alcoholism among four aboriginal groups and Han Chinese in Taiwan. Psychiatr. Genet. 2002, 11, 187–195. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, H.-M.; Su, C.-W.; Tsai, L.-C.; Huang, N.-E.; Shih, R.T.-P.; Linacre, A. Analysis of the Alu insertion polymorphism in Taiwanese Han population. Forensic Sci. J. 2002, 1, 27–30. [Google Scholar]
- Chu, C.C.; Lin, M.; Nakajima, F.; Lee, H.L.; Chang, S.L.; Juji, T.; Tokunaga, K. Diversity of HLA among Taiwan’s indigenous tribes and the Ivatans in the Philippines. Tissue Antigens 2001, 58, 9–18. [Google Scholar] [CrossRef]
- Lin, M.; Chu, C.C.; Lee, H.L.; Chang, S.L.; Ohashi, J.; Tokunaga, K.; Akaza, T.; Juji, T. Heterogeneity of Taiwan’s indigenous population: Possible relation to prehistoric Mongoloid dispersals. Tissue Antigens 2000, 55, 1–9. [Google Scholar] [CrossRef]
- Trejaut, J.A.; Muyard, F.; Lai, Y.-H.; Chen, L.-R.; Chen, Z.-S.; Loo, J.-H.; Huang, J.-Y.; Lin, M. Genetic diversity of the Thao people of Taiwan using Y-chromosome, mitochondrial DNA and HLA gene systems. BMC Evol. Biol. 2019, 19, 64. [Google Scholar] [CrossRef]
- Shinoda, K.-I. MtDNA Analysis of Bunun Remains Stored in the National Taiwan University. Anthropol. Sci. 2008, 116, 154–160. [Google Scholar] [CrossRef]
- Tajima, A.; Sun, C.-S.; Pan, I.-H.; Ishida, T.; Saitou, N.; Horai, S. Mitochondrial DNA polymorphisms in nine aboriginal groups of Taiwan: Implications for the population history of aboriginal Taiwanese. Hum. Genet. 2003, 113, 24–33. [Google Scholar] [CrossRef] [PubMed]
- Trejaut, J.A.; Poloni, E.S.; Yen, J.-C.; Lai, Y.-H.; Loo, J.-H.; Lee, C.-L.; He, C.-L.; Lin, M. Taiwan Y-chromosomal DNA variation and its relationship with Island Southeast Asia. BMC Genet. 2014, 15, 77. [Google Scholar] [CrossRef]
- Huang, J.-Y.; Trejaut, J.A.; Lee, C.-L.; Wang, T.-Y.; Loo, J.-H.; Chen, Z.S.; Chen, L.R.; Liu, K.H.; Liu, Y.C.; Hu, C.H.; et al. Mitochondrial DNA Sequencing of Middle Neolithic Human Remains of Ling-Ding Site II: Implication for the Social Structure and the Origin of Northeast Coast Taiwaneses. J. Phylogenetics Evol. Biol. 2018, 6, 2. [Google Scholar] [CrossRef]
- Chen, L.-R.; Trejaut, J.A.; Lai, Y.-H.; Chen, Z.-S.; Huang, J.-Y.; Lin, M.; Loo, J.-H. Mitochondrial DNA Polymorphisms of the Saisiyat Indigenous Group of Taiwan. Search for a Negrito Signature. Edelweiss J. Biomed. Res. Rev. 2019, 12, 12–18. [Google Scholar] [CrossRef]
- Chen, Z.-S.; Trejaut, J.; Loo, J.-H.; Lai, Y.-H.; Huang, J.-Y.; Lin, M. Mitochondrial DNA Diversity of the Nangan Islanders Living in the Mazu Archipelago of the Taiwan Strait. Edelweiss J. Biomed. Res. 2021, 3, 25–27. [Google Scholar]
- Van Oven, M.; Kayser, M. Updated comprehensive phylogenetic tree of global human mitochondrial DNA variation. Hum. Mutat. 2009, 30, E386–E394. [Google Scholar] [CrossRef]
- Loo, J.-H.; Trejaut, J.A.; Yen, J.-C.; Chen, Z.-S.; Ng, W.-M.; Huang, C.-Y.; Hsu, K.-N.; Hung, K.-H.; Hsiao, Y.; Wei, Y.-H.; et al. Mitochondrial DNA association study of type 2 diabetes with or without ischemic stroke in Taiwan. BMC Res. Notes 2014, 7, 223. [Google Scholar] [CrossRef]
- Trejaut, J.A.; Kivisild, T.; Loo, J.H.; Lee, C.L.; He, C.L.; Hsu, C.J.; Li, Z.Y.; Lin, M. Traces of archaic mitochondrial lineages persist in Austronesian-speaking Formosan populations. PLoS Biol. 2005, 3, e247. [Google Scholar] [CrossRef]
- Li, H.; Wen, B.; Chen, S.J.; Su, B.; Pramoonjago, P.; Liu, Y.; Pan, S.; Qin, Z.; Liu, W.; Cheng, X.; et al. Paternal genetic affinity between Western Austronesians and Daic populations. BMC Evol. Biol. 2008, 8, 146. [Google Scholar] [CrossRef] [PubMed]
- Ko, A.M.; Chen, C.Y.; Fu, Q.; Delfin, F.; Li, M.; Chiu, H.L.; Stoneking, M.; Ko, Y.C. Early Austronesians: Into and out of Taiwan. Am. J. Hum. Genet. 2014, 94, 426–436. [Google Scholar] [CrossRef] [PubMed]
- Melton, T.; Clifford, S.; Martinson, J.; Batzer, M.; Stoneking, M. Genetic evidence for the proto-Austronesian homeland in Asia: mtDNA and nuclear DNA variation in Taiwanese aboriginal tribes. Am. J. Hum. Genet. 1998, 63, 1807–1823. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.-C.; Chen, M.-Y.; Chao, C.-H.; Pu, C.-E. Study on the genetic polymorphisms of Y chromosomal DNA short tandem repeat loci applied to analyzing the relative affinities among ethnic groups in Taiwan. Forensic Sci. Int. Genet. Supplement. Series 2013, 4, e69–e70. [Google Scholar] [CrossRef]
- Xue, Y.; Zerjal, T.; Bao, W.; Zhu, S.; Shu, Q.; Xu, J.; Du, R.; Fu, S.; Li, P.; Hurles, M.E.; et al. Male demography in East Asia: A north-south contrast in human population expansion times. Genetics. 2006, 172, 2431–2439. [Google Scholar] [CrossRef]
- Luo, X.Q.; Du, P.X.; Wang, L.X.; Zhou, B.Y.; Li, Y.C.; Zheng, H.X.; Wei, L.H.; Liu, J.J.; Sun, C.; Meng, H.L.; et al. Uniparental Genetic Analyses Reveal the Major Origin of Fujian Tanka from Ancient Indigenous Daic Populations. Hum. Biol. 2020, 91, 257–277. [Google Scholar] [CrossRef]
- Loo, J.H.; Trejaut, J.A.; Yen, J.C.; Chen, Z.S.; Lee, C.L.; Lin, M. Genetic affinities between the Yami tribe people of Orchid Island and the Philippine Islanders of the Batanes archipelago. BMC Genet. 2011, 12, 21. [Google Scholar] [CrossRef]
- Peng, M.S.; He, J.D.; Liu, H.X.; Zhang, Y.P. Tracing the legacy of the early Hainan Islanders—A perspective from mitochondrial DNA. BMC Evol. Biol. 2011, 11, 46. [Google Scholar] [CrossRef]
- Kong, Q.P.; Yao, Y.G.; Liu, M.; Shen, S.P.; Chen, C.; Zhu, C.L.; Palanichamy, M.G.; Zhang, Y.P. Mitochondrial DNA sequence polymorphisms of five ethnic populations from northern China. Hum. Genet. 2003, 113, 391–405. [Google Scholar] [CrossRef]
- Fairley, S.; Lowy-Gallego, E.; Perry, E.; Flicek, P. The International Genome Sample Resource (IGSR) collection of open human genomic variation resources. Nucleic Acids Research. 2019, 48, D941–D947. [Google Scholar] [CrossRef]
- Liu, J.; Wang, L.D.; Sun, Y.B.; Li, E.M.; Xu, L.Y.; Zhang, Y.P.; Yao, Y.G.; Kong, Q.P. Deciphering the signature of selective constraints on cancerous mitochondrial genome. Mol. Biol. Evol. 2012, 29, 1255–1261. [Google Scholar] [CrossRef]
- Kong, Q.-P.; Yao, Y.-G.; Sun, C.; Bandelt, H.-J.; Zhu, C.-L.; Zhang, Y.-P. Phylogeny of East Asian mitochondrial DNA lineages inferred from complete sequences. Am. J. Hum. Genet. 2003, 73, 671–676. [Google Scholar] [CrossRef] [PubMed]
- Delfin, F.; Min-Shan Ko, A.; Li, M.; Gunnarsdóttir, E.D.; Tabbada, K.A.; Salvador, J.M.; Calacal, G.C.; Sagum, M.S.; Datar, F.A.; Padilla, S.G.; et al. Complete mtDNA genomes of Filipino ethnolinguistic groups: A melting pot of recent and ancient lineages in the Asia-Pacific region. Eur. J. Hum. Genet. 2014, 22, 228–237. [Google Scholar] [CrossRef]
- Duong, N.T.; Macholdt, E.; Ton, N.D.; Arias, L.; Schröder, R.; Van Phong, N.; Thi Bich Thuy, V.; Hai Ha, N.; Thi Thu Hue, H.; Thi Xuan, N.; et al. Complete human mtDNA genome sequences from Vietnam and the phylogeography of Mainland Southeast Asia. Sci. Rep. 2018, 8, 11651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kutanan, W.; Kampuansai, J.; Brunelli, A.; Ghirotto, S.; Pittayaporn, P.; Ruangchai, S.; Schröder, R.; Macholdt, E.; Srikummool, M.; Kangwanpong, D.; et al. New insights from Thailand into the maternal genetic history of Mainland Southeast Asia. Eur. J. Hum. Genet. 2018, 26, 898–911. [Google Scholar] [CrossRef] [PubMed]
- Soares, P.; Rito, T.; Trejaut, J.; Mormina, M.; Hill, C.; Tinkler-Hundal, E.; Braid, M.; Clarke, D.J.; Loo, J.H.; Thomson, N.; et al. Ancient voyaging and Polynesian origins. Am. J. Hum. Genet. 2011, 88, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Hill, C.; Soares, P.; Mormina, M.; Macaulay, V.; Meehan, W.; Blackburn, J.; Clarke, D.; Raja, J.M.; Ismail, P.; Bulbeck, D.; et al. Phylogeography and Ethnogenesis of Aboriginal Southeast Asians. Mol. Biol. Evol. 2006, 23, 2480–2491. [Google Scholar] [CrossRef]
- Gunnarsdóttir, E.D.; Nandineni, M.R.; Li, M.; Myles, S.; Gil, D.; Pakendorf, B.; Stoneking, M. Larger mitochondrial DNA than Y-chromosome differences between matrilocal and patrilocal groups from Sumatra. Nat. Commun. 2011, 2, 228. [Google Scholar] [CrossRef]
- Jinam, T.A.; Hong, L.C.; Phipps, M.E.; Stoneking, M.; Ameen, M.; Edo, J.; HUGO Pan-Asian SNP Consortium; Saitou, N. Evolutionary history of continental southeast Asians: "early train" hypothesis based on genetic analysis of mitochondrial and autosomal DNA data. Mol. Biol. Evol. 2012, 29, 3513–3527. [Google Scholar] [CrossRef]
- Tajima, F. Statistical method for testing the neutral mutation hypothesis by DNA polymorphism. Genetics 1989, 123, 585–595. [Google Scholar] [CrossRef]
- Duggan, A.T.; Evans, B.; Friedlaender, F.R.; Friedlaender, J.S.; Koki, G.; Merriwether, D.A.; Kayser, M.; Stoneking, M. Maternal history of Oceania from complete mtDNA genomes: Contrasting ancient diversity with recent homogenization due to the Austronesian expansion. Am. J. Hum. Genet. 2014, 94, 721–733. [Google Scholar] [CrossRef]
- Soares, P.A.; Trejaut, J.A.; Rito, T.; Cavadas, B.; Hill, C.; Eng, K.K.; Mormina, M.; Brandão, A.; Fraser, R.M.; Wang, T.Y.; et al. Resolving the ancestry of Austronesian-speaking populations. Hum. Genet. 2016, 135, 309–326. [Google Scholar] [CrossRef]
- Zhang, X.; Qi, X.; Yang, Z.; Serey, B.; Sovannary, T.; Bunnath, L.; Aun, H.S.; Samnom, H.; Zhang, H.; Lin, Q.; et al. Analysis of mitochondrial genome diversity identifies new and ancient maternal lineages in Cambodian aborigines. Nat. Commun. 2013, 4, 2599. [Google Scholar] [CrossRef] [PubMed]
- Brandão, A.; Eng, K.K.; Rito, T.; Cavadas, B.; Bulbeck, D.; Gandini, F.; Pala, M.; Mormina, M.; Hudson, B.; White, J.; et al. Quantifying the legacy of the Chinese Neolithic on the maternal genetic heritage of Taiwan and Island Southeast Asia. Hum. Genet. 2016, 135, 363–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delfin, F.; Salvador, J.M.; Calacal, G.C.; Perdigon, H.B.; Tabbada, K.A.; Villamor, L.P.; Halos, S.C.; Gunnarsdóttir, E.; Myles, S.; Hughes, D.A.; et al. The Y-chromosome landscape of the Philippines: Extensive heterogeneity and varying genetic affinities of Negrito and non-Negrito groups. Eur. J. Hum. Genet. 2010, 19, 224–230. [Google Scholar] [CrossRef] [PubMed]
- Heyer, E.; Georges, M.; Pachner, M.; Endicott, P. Genetic diversity of four Filipino negrito populations from Luzon: Comparison of male and female effective population sizes and differential integration of immigrants into Aeta and Agta communities. Hum. Biol. 2013, 85, 189–208. [Google Scholar] [CrossRef]
- Su, B.; Jin, L.; Underhill, P.; Martinson, J.; Saha, N.; McGarvey, S.T.; Shriver, M.D.; Chu, J.; Oefner, P.; Chakraborty, R.; et al. Polynesian origins: Insights from the Y chromosome. Proc. Natl. Acad. Sci. USA 2000, 97, 8225–8228. [Google Scholar] [CrossRef]
- Zhong, H.; Shi, H.; Qi, X.B.; Duan, Z.Y.; Tan, P.P.; Jin, L.; Su, B.; Ma, R.Z. Extended Y chromosome investigation suggests postglacial migrations of modern humans into East Asia via the northern route. Mol. Biol. Evol. 2011, 28, 717–727. [Google Scholar] [CrossRef]
- Kayser, M.; Choi, Y.; van Oven, M.; Mona, S.; Brauer, S.; Trent, R.J.; Suarkia, D.; Schiefenhövel, W.; Stoneking, M. The impact of the Austronesian expansion: Evidence from mtDNA and Y chromosome diversity in the Admiralty Islands of Melanesia. Mol. Biol. Evol. 2008, 25, 1362–1374. [Google Scholar] [CrossRef]
- Urasin, V. YTREE V8.04. Available online: https://www.yfull.com/arch-8.04/tree/ (accessed on 7 July 2022).
- Waas, M.; Yacobi, D.; Kull, L.; Urasin, V.; Magoon, G.; Penninx, W.; Brown, A.; Nogueiro, I. Haplogroup J-Z640-Genetic Insight into the Levantine Bronze Age. J. Phylogenetics Evol. Biol. 2019, 7, 209. [Google Scholar]
- Razafindrazaka, H. Le Peuplement Humain de Madagascar: Anthropologie Génétique de Trois Groupes Traditionnels; Université Toulouse III-Paul Sabatier: Toulouse, France, 2010. (In French) [Google Scholar]
- Razafindrazaka, H.; Ricaut, F.X.; Cox, M.P.; Mormina, M.; Dugoujon, J.M.; Randriamarolaza, L.P.; Guitard, E.; Tonasso, L.; Ludes, B.; Crubézy, E. Complete mitochondrial DNA sequences provide new insights into the Polynesian motif and the peopling of Madagascar. Eur. J. Hum. Genet. 2010, 18, 575–581. [Google Scholar] [CrossRef] [PubMed]
- Tofanelli, S.; Ferri, G.; Bulayeva, K.; Caciagli, L.; Onofri, V.; Taglioli, L.; Bulayev, O.; Boschi, I.; Alù, M.; Berti, A.; et al. J1-M267 Y lineage marks climate-driven pre-historical human displacements. Eur. J. Hum. Genet. 2009, 17, 1520–1524. [Google Scholar] [CrossRef] [PubMed]
- van Oven, M.; Hammerle, J.M.; van Schoor, M.; Kushnick, G.; Pennekamp, P.; Zega, I.; Lao, O.; Brown, L.; Kennerknecht, I.; Kayser, M. Unexpected island effects at an extreme: Reduced Y chromosome and mitochondrial DNA diversity in Nias. Mol. Biol. Evol. 2011, 28, 1349–1361. [Google Scholar] [CrossRef] [PubMed]
- He, J.D.; Peng, M.S.; Quang, H.H.; Dang, K.P.; Trieu, A.V.; Wu, S.F.; Jin, J.Q.; Murphy, R.W.; Yao, Y.G.; Zhang, Y.P. Patrilineal perspective on the Austronesian diffusion in Mainland Southeast Asia. PLoS ONE 2012, 7, e36437. [Google Scholar] [CrossRef]
- Cai, X.; Qin, Z.; Wen, B.; Xu, S.; Wang, Y.; Lu, Y.; Wei, L.; Wang, C.; Li, S.; Huang, X.; et al. Human migration through bottlenecks from Southeast Asia into East Asia during Last Glacial Maximum revealed by Y chromosomes. PLoS ONE 2011, 6, e24282. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Kampuansai, J.; Qi, X.; Yan, S.; Yang, Z.; Serey, B.; Sovannary, T.; Bunnath, L.; Aun, H.S.; Samnom, H.; et al. An Updated Phylogeny of the Human Y-Chromosome Lineage O2a-M95 with Novel SNPs. PLoS ONE 2014, 9, e101020. [Google Scholar] [CrossRef]
- Dalton, R. Neanderthal DNA yields to genome foray. Nature 2006, 441, 260–261. [Google Scholar] [CrossRef]
- Brunelli, A.; Kampuansai, J.; Seielstad, M.; Lomthaisong, K.; Kangwanpong, D.; Ghirotto, S.; Kutanan, W. Y chromosomal evidence on the origin of northern Thai people. PLoS ONE 2017, 12, e0181935. [Google Scholar]
- Simonson, T.S.; Xing, J.; Barrett, R.; Jerah, E.; Loa, P.; Zhang, Y.; Watkins, W.S.; Witherspoon, D.J.; Huff, C.D.; Woodward, S.; et al. Ancestry of the Iban is predominantly Southeast Asian: Genetic evidence from autosomal, mitochondrial, and Y chromosomes. PLoS ONE 2011, 6, e16338. [Google Scholar] [CrossRef]
- Blust, R. Subgrouping, circularity and extinction: Some issues in Austronesian comparative linguistics. Symp. Ser. Inst. Linguist. Acad. Sinica 1999, 1, 31–94. [Google Scholar]
- Msaidie, S.; Ducourneau, A.; Boetsch, G.; Longepied, G.; Papa, K.; Allibert, C.; Yahaya, A.A.; Chiaroni, J.; Mitchell, M.J. Genetic diversity on the Comoros Islands shows early seafaring as major determinant of human biocultural evolution in the Western Indian Ocean. Eur. J. Hum. Genet. 2011, 19, 89–94. [Google Scholar] [CrossRef] [PubMed]
- Tofanelli, S.; Bertoncini, S.; Castri, L.; Luiselli, D.; Calafell, F.; Donati, G.; Paoli, G. On the origins and admixture of Malagasy: New evidence from high-resolution analyses of paternal and maternal lineages. Mol. Biol. Evol. 2009, 26, 2109–2124. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.C.; Li, H. Inferring human history in East Asia from Y chromosomes. Investig. Genet. 2013, 4, 11. [Google Scholar] [CrossRef] [PubMed]
- ISSOG. International Society of Genetic Genealogy. Y-DNA Haplogroup Tree 2019, Version 15.46. Available online: https://www.isogg.org/tree/ (accessed on 12 March 2020).
- Pickrell, J.K.; Pritchard, J.K. Inference of population splits and mixtures from genome-wide allele frequency data. PLoS Genet. 2012, 8, e1002967. [Google Scholar] [CrossRef]
- Soares, P.; Trejaut, J.A.; Loo, J.H.; Hill, C.; Mormina, M.; Lee, C.L.; Chen, Y.M.; Hudjashov, G.; Forster, P.; Macaulay, V.; et al. Climate change and postglacial human dispersals in southeast Asia. Mol. Biol. Evol. 2008, 25, 1209–1218. [Google Scholar] [CrossRef]
- Tabbada, K.A.; Trejaut, J.; Loo, J.H.; Chen, Y.M.; Lin, M.; Mirazon-Lahr, M.; Kisivild, T.; De Ungria, M.C.A. Philippine mitochondrial DNA diversity: A populated viaduct between Taiwan and Indonesia? Mol. Biol. Evol. 2010, 27, 21–31. [Google Scholar] [CrossRef]
- Kutanan, W.; Kampuansai, J.; Srikummool, M.; Brunelli, A.; Ghirotto, S.; Arias, L.; Macholdt, E.; Hübner, A.; Schröder, R.; Stoneking, M. Contrasting Paternal and Maternal Genetic Histories of Thai and Lao Populations. Mol. Biol. Evol. 2019, 36, 1490–1506. [Google Scholar] [CrossRef]
- Gan, R.J.; Pan, S.L.; Mustavich, L.F.; Qin, Z.D.; Cai, X.Y.; Qian, J.; Liu, C.W.; Peng, J.H.; Li, S.L.; Xu, J.S.; et al. Pinghua population as an exception of Han Chinese’s coherent genetic structure. J. Hum. Genet. 2008, 53, 303–313. [Google Scholar] [CrossRef]
- Li, H. Genetic Structure of Austro-Tai Populations. Ph.D. Dissertation, Fudan University, Shanghai, China, 2005. [Google Scholar]
- Kutanan, W.; Kampuansai, J.; Srikummool, M.; Kangwanpong, D.; Ghirotto, S.; Brunelli, A.; Stoneking, M. Complete mitochondrial genomes of Thai and Lao populations indicate an ancient origin of Austroasiatic groups and demic diffusion in the spread of Tai-Kadai languages. Hum Genet. 2017, 136, 85–98. [Google Scholar] [CrossRef]
- Li, D.; Li, H.; Ou, C.; Lu, Y.; Sun, Y.; Yang, B.; Qin, Z.; Zhou, Z.; Li, S.; Jin, L. Paternal genetic structure of Hainan aborigines isolated at the entrance to East Asia. PLoS ONE 2008, 3, e2168. [Google Scholar] [CrossRef]
- Bekaert, B.; Zainuddin, Z.; Hadi, S.; Goodwin, W. A comparison of mtDNA and Y chromosome diversity in Malay populations. Int. Congr. Ser. 2006, 1288, 252–255. [Google Scholar] [CrossRef]
- Chen, Y.; Chu, J.Y.; Yu, J.K.; Yu, L.; Sun, H.; Lin, K.Q.; Tao, Y.F.; Shi, L.; Huang, X.Q.; Shi, T.L.; et al. Polymorphism of DYS287 on Y chromosome in 28 ethnic populations of China. Zhongguo Yi Xue Ke Xue Yuan Xue Bao 2006, 28, 196–201. (In Chinese) [Google Scholar] [PubMed]
- Weissensteiner, H.; Pacher, D.; Kloss-Brandstätter, A.; Forer, L.; Specht, G.; Bandelt, H.; Kronenberg, F.; Salas, A.; Schönherr, S. HaploGrep 2: Mitochondrial haplogroup classification in the era of high-throughput sequencing. Nucl. Acids Res. 2016, 44, W58–W63. [Google Scholar] [CrossRef]
- Fu, Y.X. Statistical tests of neutrality of mutations against population growth, hitchhiking and background selection. Genetics 1997, 147, 915–925. [Google Scholar] [CrossRef] [PubMed]
- Excoffier, L.; Lischer, H.E. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef] [PubMed]
- Hammer, O.; Harper, D.A.T.; Ryan, P.D. Past: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 9. Available online: http://palaeo-electronica.org/2001_1/past/issue1_01.htm (accessed on 17 July 2022).
- Bandelt, H.J.; Forster, P.; Röhl, A. Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Zhivotovsky, L.A.; Underhill, P.A.; Cinnioğlu, C.; Kayser, M.; Morar, B.; Kivisild, T.; Scozzari, R.; Cruciani, F.; Destro-Bisol, G.; Spedini, G.; et al. The effective mutation rate at Y chromosome short tandem repeats, with application to human population-divergence time. Am. J. Hum. Genet. 2004, 74, 50–61. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Location of the Bunun indigenous people within the central mountain range of Taiwan. The currently dominant Bunun language, Isbukun, is spoken in the south. Takbunuaz and Takivatan dialects are spoken by the Central Bunun, and Takituduh and Takibaka are spoken in the north. In Bold: Location of Taiwan, officially recognized indigenous people (AN_Tw). In Grey: Not officially recognized indigenous groups (also referred to as Pingpu). Reference data of other groups in East and Southeast Asia were obtained from the literature.
Figure 1.
Location of the Bunun indigenous people within the central mountain range of Taiwan. The currently dominant Bunun language, Isbukun, is spoken in the south. Takbunuaz and Takivatan dialects are spoken by the Central Bunun, and Takituduh and Takibaka are spoken in the north. In Bold: Location of Taiwan, officially recognized indigenous people (AN_Tw). In Grey: Not officially recognized indigenous groups (also referred to as Pingpu). Reference data of other groups in East and Southeast Asia were obtained from the literature.

Figure 2.
MtDNA comparison of the three Bunun groups: Northern Bunun (Blue), Miscellaneous Bunun (Orange), and Central Bunun (Grey) [33].
Figure 2.
MtDNA comparison of the three Bunun groups: Northern Bunun (Blue), Miscellaneous Bunun (Orange), and Central Bunun (Grey) [33].

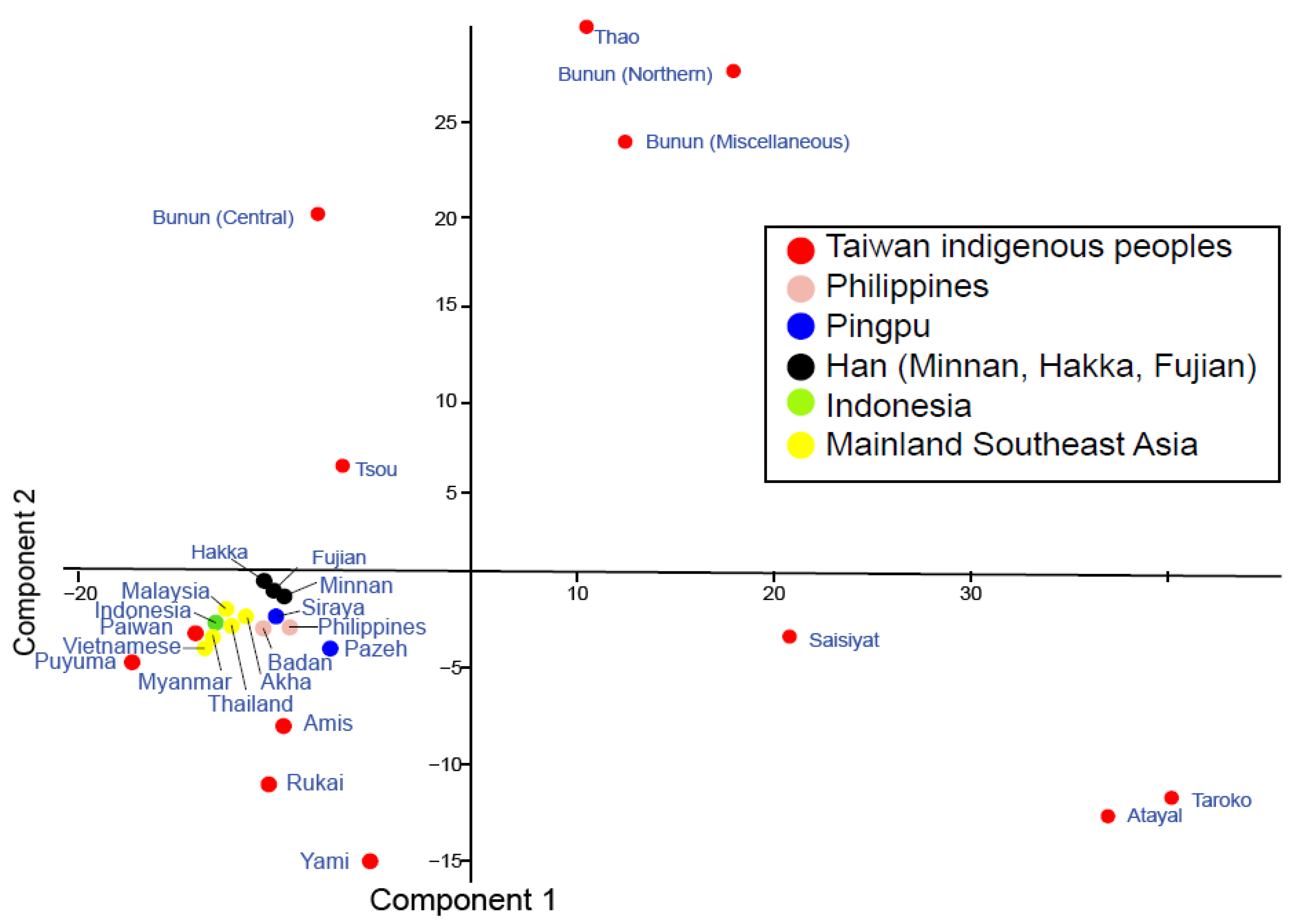
Figure 3.
Principal component analysis (PCA) using mtDNA polymorphism of population from MSEA, ISEA, EA, and Taiwan. The PCA plot separates the Northern and Miscellaneous Bunun from the Central Bunun along Component 1 (upper quadrants) and the Northern and Miscellaneous Bunun (upper right quadrant) from the Central Bunun group (upper left quadrant). MSEA, ISEA, and Han form smaller clusters in the lower left pane showing a closer relationship to the Southern AN_Tw (Puyama, Paiwan, Rukai, Amis, and Yami). Pingpu represents the Taiwan Western plain dwellers Pazeh and Siraya people.
Figure 3.
Principal component analysis (PCA) using mtDNA polymorphism of population from MSEA, ISEA, EA, and Taiwan. The PCA plot separates the Northern and Miscellaneous Bunun from the Central Bunun along Component 1 (upper quadrants) and the Northern and Miscellaneous Bunun (upper right quadrant) from the Central Bunun group (upper left quadrant). MSEA, ISEA, and Han form smaller clusters in the lower left pane showing a closer relationship to the Southern AN_Tw (Puyama, Paiwan, Rukai, Amis, and Yami). Pingpu represents the Taiwan Western plain dwellers Pazeh and Siraya people.

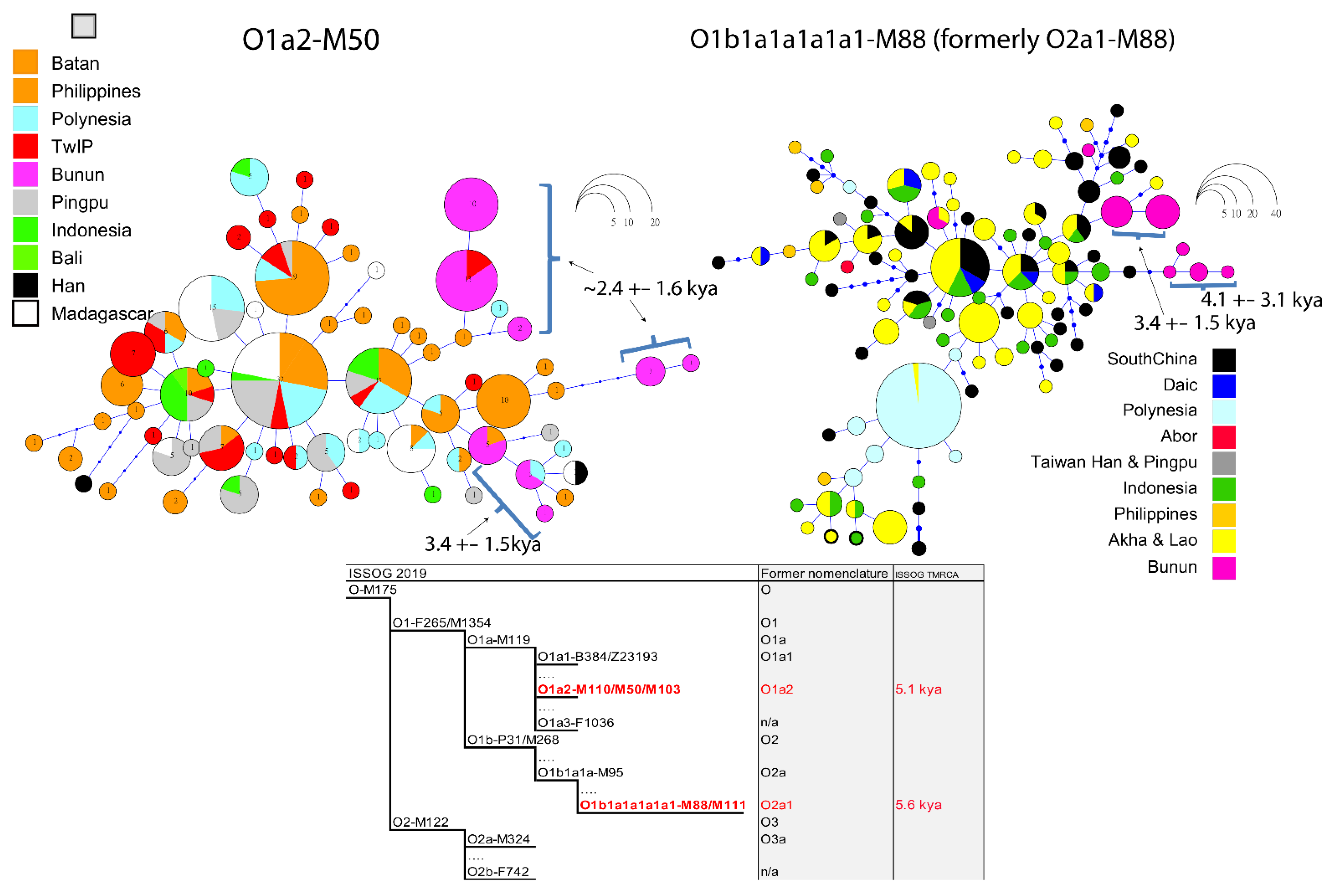
Figure 4.
Median joining network of O1a2-M50 and O1b1a1a1a1a1-M88 NRY haplogroups associated with seven Y-STR. The sizes of circles in the networks represent the number of individuals sharing the same haplotype. The lengths of lines between circles are proportional to the mutation steps. Accolades indicate the Northern Bunun age range in kya. The red prints in the ISSOG phylogram (O1a2-M50 and O1b1a1a1a1a1-M88) correspond to the ISSOG 2019 nomenclature of former O1a2-M50 and O2a1-M88 Y-chromosome haplogroups. Pingpu refers to Western plain dwellers of Taiwan (Pazeh and Siraya). Akha and Lao represent groups of Mainland Southeast Asia. Oceania represents the dataset of Delfin et al. 2011 [42] (e.g., Tonga, Samoa, Fiji).
Figure 4.
Median joining network of O1a2-M50 and O1b1a1a1a1a1-M88 NRY haplogroups associated with seven Y-STR. The sizes of circles in the networks represent the number of individuals sharing the same haplotype. The lengths of lines between circles are proportional to the mutation steps. Accolades indicate the Northern Bunun age range in kya. The red prints in the ISSOG phylogram (O1a2-M50 and O1b1a1a1a1a1-M88) correspond to the ISSOG 2019 nomenclature of former O1a2-M50 and O2a1-M88 Y-chromosome haplogroups. Pingpu refers to Western plain dwellers of Taiwan (Pazeh and Siraya). Akha and Lao represent groups of Mainland Southeast Asia. Oceania represents the dataset of Delfin et al. 2011 [42] (e.g., Tonga, Samoa, Fiji).

Figure 5.
Principal component analysis using NRY haplogroup polymorphism. Note: The Y Chromosome of two Northern Bunun samples [25,32] are shown separately. Reference data for other groups were obtained from the literature (including an East African group used as an out-group [63,64].

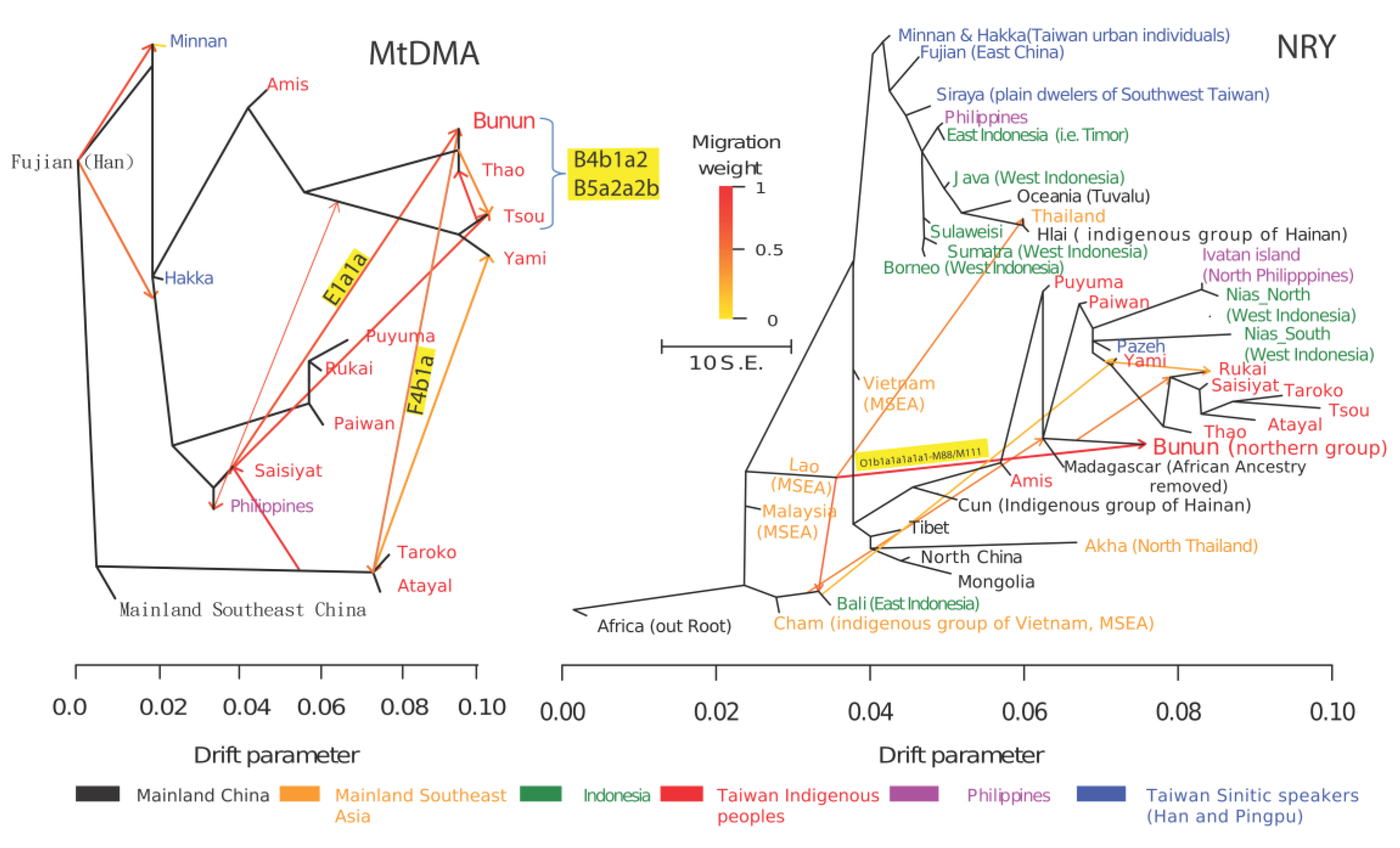
Figure 6.
Inferred sources of gene flow of the Northern Bunun indigenous people. The Maximum-Likelihood tree inferred by TreeMix for Taiwan and SEA populations [78] was constructed using 15 significant gene-flow events. Shaded colored arrows represent directional gene flow across pairs of populations. For clarity, only gene flow with migration weight greater than 0.5 is shown. Left: Gene flow of mtDNA haplogroups. Note: Haplogroups shown on a yellow background (B4b1a2, F4b1a, and E1a1a) are inferences from Supplementary Table S1, and mitochondrial studies [27,80]. Right: Gene flow of NRY-STR for haplogroups O1b1a1a1a1a1-M88. Note: Haplogroups shown on a yellow background are inferences from Table 2 and Y-chromosome studies [25,68]. TreeMix detected gene flow for mtDNA from closer geographic regions compared to the Y-chromosome. This suggests that male dispersal patterns cover longer geographical distances compared to female patterns.
Figure 6.
Inferred sources of gene flow of the Northern Bunun indigenous people. The Maximum-Likelihood tree inferred by TreeMix for Taiwan and SEA populations [78] was constructed using 15 significant gene-flow events. Shaded colored arrows represent directional gene flow across pairs of populations. For clarity, only gene flow with migration weight greater than 0.5 is shown. Left: Gene flow of mtDNA haplogroups. Note: Haplogroups shown on a yellow background (B4b1a2, F4b1a, and E1a1a) are inferences from Supplementary Table S1, and mitochondrial studies [27,80]. Right: Gene flow of NRY-STR for haplogroups O1b1a1a1a1a1-M88. Note: Haplogroups shown on a yellow background are inferences from Table 2 and Y-chromosome studies [25,68]. TreeMix detected gene flow for mtDNA from closer geographic regions compared to the Y-chromosome. This suggests that male dispersal patterns cover longer geographical distances compared to female patterns.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Population samples.
| Country | Population | Size | Languages | Groups | References | ||
|---|---|---|---|---|---|---|---|
| MtDNA | NRY | MtDNA | NRY | ||||
| Taiwan | Bunun (Northern) | 89 | 56 | Austronesian | Northern AN_Tw | [31] | [22,25,32] |
| Taiwan | Bunun (Central) | 50 | n/a | Austronesian | Central AN_Tw | [33] | n/a |
| Taiwan | Bunun (Miscellaneous) | 96 | n/a | Austronesian | Miscellaneous | [24,34] | [35] (STRs only) |
| Taiwan | Hakka | 45 | 34 | Sinitic | TwH/Urban Taiwanese | [33] | [25] |
| Taiwan | Minnan | 50 | 60 | Sinitic | TwH/Urban Taiwanese | [33] | [25,36] |
| China | Fujian and Matsu | 198 | 55 | Sinitic | East Coast China | [28,37] | [25] |
| Taiwan | Saisiyat | 87 | 24 | Austronesian | Northern AN_Tw | [31,33] | [25] |
| Taiwan | Atayal | 157 | 52 | Austronesian | Northern AN_Tw | [31,33] | [25] |
| Taiwan | Amis | 148 | 39 | Austronesian | East Coast Taiwan | [31,33] | [25] |
| Taiwan | Taroko | 54 | 20 | Austronesian | Northern AN_Tw | [22] * | [25] |
| Taiwan | Thao | 30 | 16 | Austronesian | Central AN_Tw | [22,31,33] | [22,25] |
| Taiwan | Tsou | 108 | 41 | Austronesian | Central AN_Tw | [31,33] | [25] |
| Taiwan | Paiwan | 105 | 25 | Austronesian | Southern AN_Tw | [31,33] | [25] |
| Taiwan | Puyuma | 91 | 23 | Austronesian | Southern AN_Tw | [31,33] | [25] |
| Taiwan | Rukai | 99 | 29 | Austronesian | Southern AN_Tw | [31,33] | [25] |
| Taiwan | Yami | 123 | 30 | Austronesian | Southern AN_Tw | [31,38] | [25,38] |
| Northeast China | Beijing, Henan, and Liaoning | 257 | na | Sinitic | NE_China (Han) | [39,40,41,42,43] | [36] |
| South China | South China | 65 | na | Sinitic | Sth_China (Han) | [41,43] | [32,36] |
| Philippines | Indigenous groups | 260 | 121 | Malayo Polynesian | ISEA | [44] | [25] |
| Vietnam | Indigenous groups | 603 | 24 | Austro Asiatic | MSEA | [45] | [25] |
| Thailand | Indigenous groups | 560 | 75 | Kra-Dai | MSEA | [46] | [25] |
| Indonesia (East) | East_Indonesia | 72 | na | Malayo Polynesian | ISEA | [47,48,49] | [25,32] |
| Indonesia (West) | West_Indonesia | 326 | 68 | Malayo Polynesian | ISEA | [47,48,49] | [25] |
| Malaysia | Indigenous groups | 86 | 8 | Austronesian | MSEA | [50] | [25] |
| Total | 4423 | 800 | |||||
AN_Tw: Austronesian speaking Taiwanese (officially recognized Taiwan Indigenous Peoples; *: Incomplete frequency data; TwH: Taiwan Han (Urban Taiwanese: Minnan and Hakka); ISEA: Island Southeast Asia; MSEA: Mainland Southeast Asia (Indochina) (Vietnam, Malaysia, Thailand).
Table 2.
Frequency table of NRY polymorphisms of the Northern Bunun and comparison with other populations.
Table 2.
Frequency table of NRY polymorphisms of the Northern Bunun and comparison with other populations.
| Populations | China | Taiwan | Island Southeast Asia | Mainland Southeast Asia | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Han | Urban Taiwanese | Taiwan Indigenous People (AN_Tw) | Philippines | Indonesia | Total Indonesia (East, West and 36 undefined) | Thailand Total | Vietnam (Hanoy) | |||||||||||||
| Fujian | Hakka | Minnan | Atayal | Taroko | Saisiyat | Thao | Northern Bunun | Tsou | Yami | Amis | Rukai | Paiwan | Puyuma | West Indonesia (Java+Borneo+Sumatra) | East Indonesia(Maluku Ambon) | |||||
| O1a2M50 | 2.94 | 1.92 | 4.17 | 6.25 | 60.71 | 4.88 | 17.95 | 24.14 | 28.00 | 21.74 | 11.64 | 5.73 | 11.11 | 5.28 | ||||||
| O1b1a1a1a1a1-M88 (O2a1aM88) | 2.94 | 1.67 | 37.50 | 3.33 | 3.42 | 3.64 | 3.25 | 19.61 | 25.00 | |||||||||||
| P*M45 | 1.79 | 2.56 | ||||||||||||||||||
| others | 100 | 94.12 | 98.33 | 98.08 | 100 | 95.83 | 93.75 | 0.00 | 95.12 | 96.67 | 79.49 | 75.86 | 72.00 | 78.26 | 84.94 | 90.63 | 88.89 | 91.47 | 80.39 | 75.00 |
| Y-SNP Sample size | 55 | 34 | 60 | 52 | 20 | 24 | 16 | 56 | 41 | 30 | 39 | 29 | 25 | 23 | 146 | 192 | 18 | 246 | 102 | 24 |
| Haplogroup Diversity | 0.86 | 0.92 | 0.90 | 0.18 | 0.10 | 0.24 | 0.24 | 0.50 | 0.19 | 0.65 | 0.69 | 0.48 | 0.73 | 0.72 | 0.90 | 0.86 | 0.88 | 0.87 | 0.89 | 0.91 |
| ±SD | 0.035 | 0.027 | 0.024 | 0.202 | 0.196 | 0.195 | 0.219 | 0.106 | 0.207 | 0.097 | 0.069 | 0.187 | 0.064 | 0.104 | 0.018 | 0.034 | 0.030 | 0.030 | 0.023 | 0.032 |
| Total Number of Haplogroups | 13 | 18 | 16 | 3 | 2 | 4 | 3 | 3 | 3 | 5 | 5 | 3 | 5 | 5 | 26 | 24 | 7 | 27 | 21 | 12 |
AN_Tw: Austronesian speaking Taiwanese.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Trejaut, J.A. Origin of the Bunun Indigenous People of Taiwan, a Review of Published Material Using Y-Chromosome and Mitochondrial DNA Gene Systems. DNA 2022, 2, 185-201. https://doi.org/10.3390/dna2030013
AMA Style
Trejaut JA. Origin of the Bunun Indigenous People of Taiwan, a Review of Published Material Using Y-Chromosome and Mitochondrial DNA Gene Systems. DNA. 2022; 2(3):185-201. https://doi.org/10.3390/dna2030013
Chicago/Turabian StyleTrejaut, Jean A. 2022. "Origin of the Bunun Indigenous People of Taiwan, a Review of Published Material Using Y-Chromosome and Mitochondrial DNA Gene Systems" DNA 2, no. 3: 185-201. https://doi.org/10.3390/dna2030013