
Validation of Adapted Neutralization Assays Developed to Discriminate Anti-Rabies Virus Activity of Two Different Anti-Rabies Virus Monoclonal Antibodies Administered as a Combination

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Validation Experiments
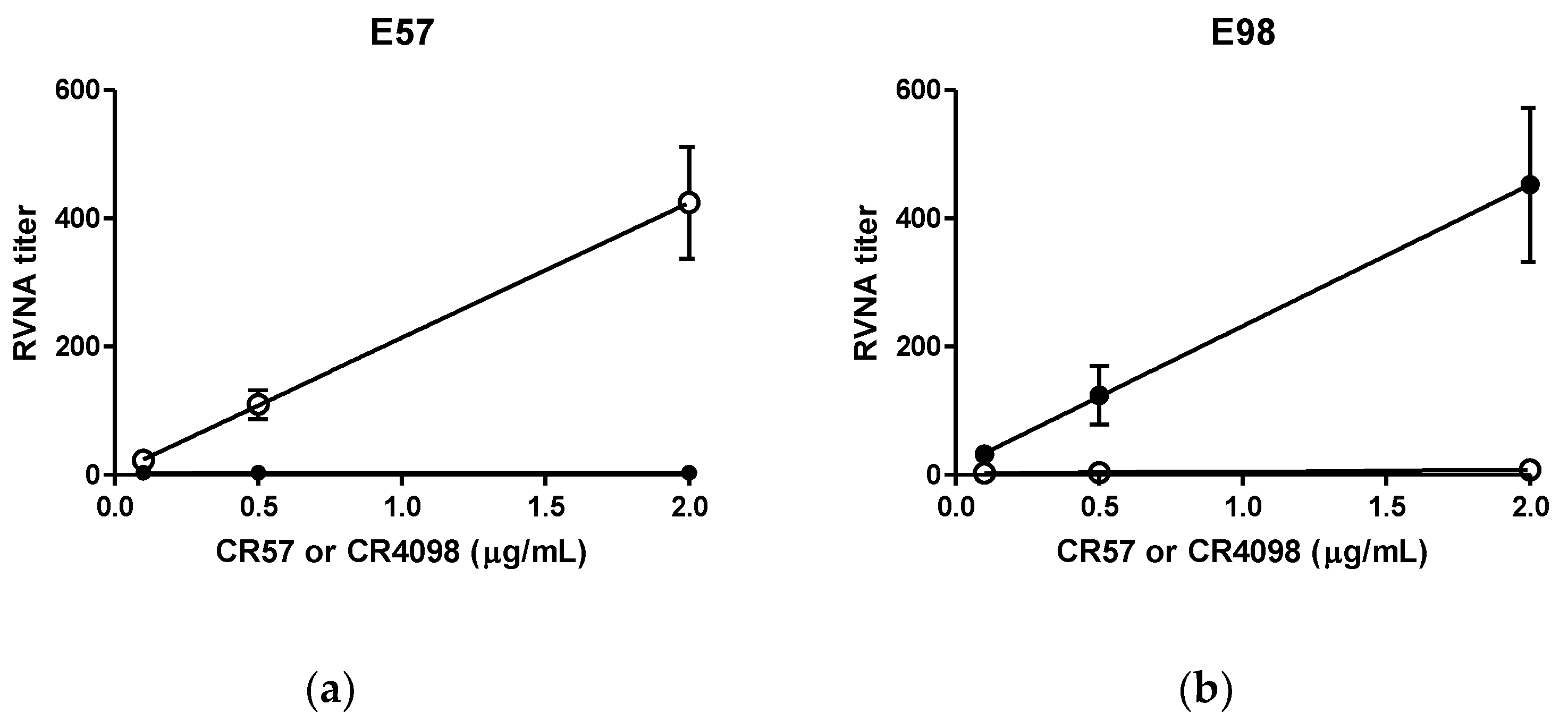
2.1.1. Specificity
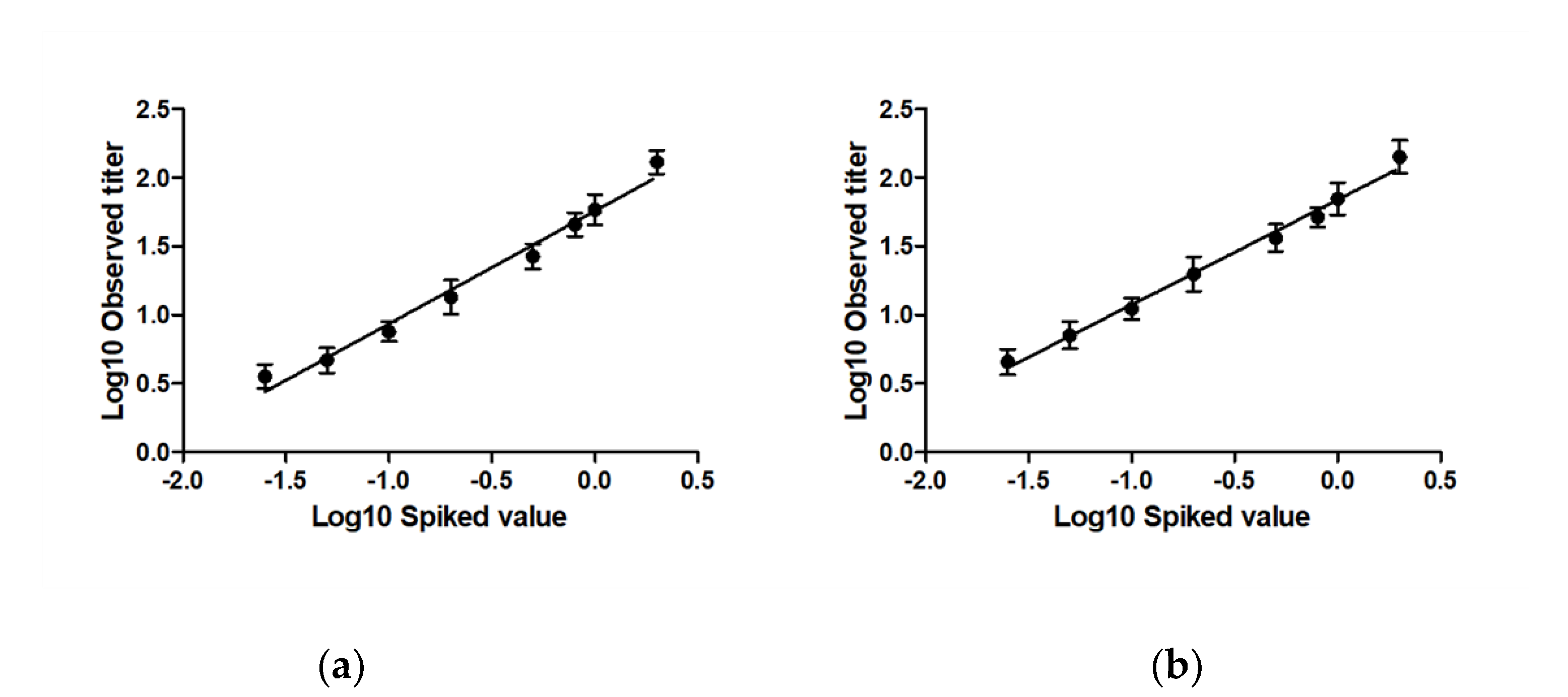
2.1.2. Linearity and Accuracy
2.1.3. Lower Limit of Quantitation (LLOQ) and Lower Limit of Detection (LOD)
2.1.4. Repeatability (Intra-Assay Variation)
2.1.5. Intermediate Precision (Inter-Assay Variation)
2.1.6. Stability
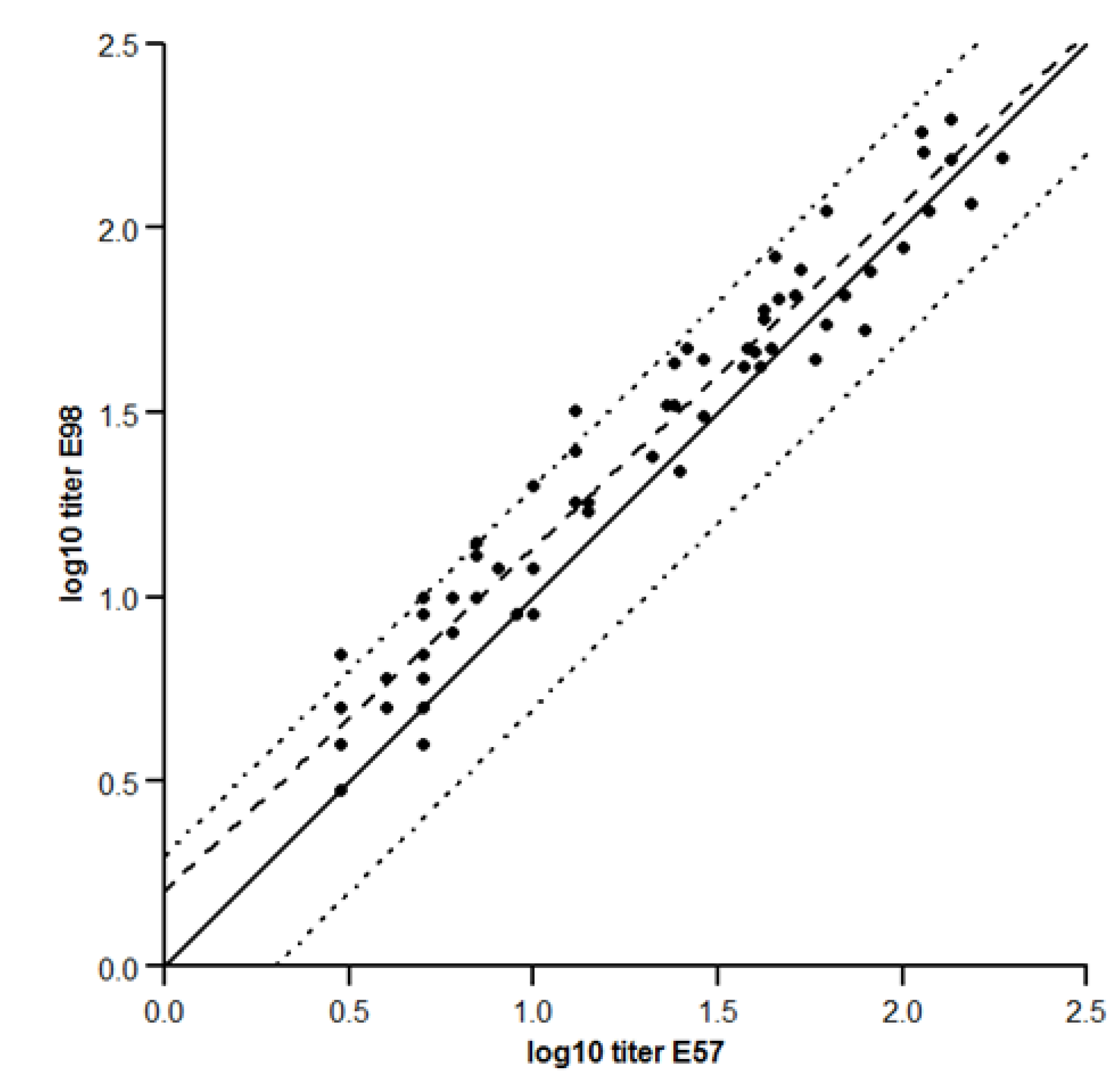
2.1.7. Concordance between the Two Assays
3. Discussion
4. Materials and Methods
- RFFIT protocol
- Validation samples
- Challenge viruses
- Validation experiments
- (1)
- Specificity. Specificity was tested two ways: First, the two MAbs, CR57 and CR4098, were incubated separately with the challenge virus E57 or E98. Criteria were that E57 should be neutralized by CR4098, but not by CR57, whereas E98 should be neutralized by CR57, but not by CR4098. Second, the specificity of the virus in context of CL184 was tested. Hereto inactivated virus was incubated with CL184 to adsorb either CR57 or CR4098. Subsequently the mixture was incubated with live challenge virus and compared with CL184 not adsorbed to the inactivated virus. As a negative control inactivated irrelevant rhabdovirus (Vesicular Stomatitis Virus-Indiana, (VSV-IN)) was used. The ratio of the RVNA titer signal observed using the unabsorbed CL184 and the adsorbed CL184 was calculated. A reduction of ≥4 fold in signal indicates specificity of the assay.
- (2)
- Linearity and Accuracy. To assess linearity, the ICH guidelines on validation of analytical procedures (Q2(R1)) were followed. A regression line was fitted through the observed RVNA titer level data points (i.e., the log10 transformed antibody measured titers as a function of the log10 transformed potency level of CL184) using maximum likelihood method. Linearity was analyzed using a linear regression model and accuracy assessed via analysis of the residuals. The 90% confidence interval at each level should be included within ±0.114 log-units (corresponding with 30% CV on the original scale) from the expected result, (i.e., the regression line). The range where the data follows a linear regression model constitutes the linear range of the assay.
- (3)
- Precision. Repeatability (intra-assay variation) and intermediate precision (the sum of the inter-assay and intra-assay variation) of the assays were determined by assessment of eight CL184 validation samples covering the RVNA range of 0.025 to 2 IU/mL. Guidelines recommend a CV of 20% to 25% for ligand binding assays (LBA), such as enzyme-linked immunosorbent assays (ELISA). However, since bioassays, such as the RFFIT, make use of biological agents such as cells and virus, which are prone to a higher variation, a higher variability is expected, as acknowledged by the World Health Organization [32,33]. A larger CV limit criterion of <30% is therefore accepted. More variation is expected for the LLOQ and ULOQ [21], thus acceptance criteria for the LLOQ is 35% instead of 30%. We accept this higher variability because data in this range are considered relevant in the clinical trials in which, at early time points, relative low levels of passively administered RIG are expected. RVNA titer data were first log10 transformed to achieve a normal distribution. The repeatability and intermediate precision of the RVNA titer values was estimated in a nested error variance components model by analysis of variance (ANOVA) with experiment as random term. To express precision in percent CV, the formulawas used to translate the standard deviation (σ) on a log10 scale to a percent CV, which can be compared with the acceptance criteria.
- (4)
- Stability. Stability was assessed as described in the EMA and FDA guidelines on bioanalytical method validation. The stability items tested reflect the routine handling of clinical samples. During all stability experiments, 3 RVNA spike levels (0.1, 0.5 and 2.0 IU/mL) were tested. During each stability test, a total of 7 aliquots per spike level were stressed and compared to the same number of non-stressed comparator samples. Freeze thaw testing was performed as follows: 3 sets of 7 aliquots were taken from the −80 °C freezer and allowed to thaw at room temperature (RT) for 4 h. Next, the samples were placed back and allowed to freeze for at least 24 h. Subsequently, all samples were taken from the freezer and allowed to thaw at RT for 4 h (freeze/thaw cycle 1). One set was tested in the RFFIT assays, and the remaining 2 sets were placed back in the −80 °C freezer for at least 24 h. This procedure was repeated for a total of 3 cycles. The RVNA titer levels for each stability sample were log10 transformed and compared with the log10 transformed comparator samples by ANOVA, with spiked value as covariate to correct for differences in concentration. Appropriate estimate statements in the ANOVA were used to estimate the mean differences and the corresponding 90% confidence intervals of various stability conditions to the comparator. As a criterion, no significant differences of more 30% on original scale between stability samples and comparator samples as determined by ANOVA were allowed.
- (5)
- Concordance between the two assays was performed using an orthogonal regression model [34], assuming the error on both measurements was the same, as suggested in the precision section.
- Calculations
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Both, L.; Banyard, A.C.; van Dolleweerd, C.; Wright, E.; Ma, J.K.-C.; Fooks, A.R. Monoclonal antibodies for prophylactic and therapeutic use against viral infections. Vaccine 2013, 31, 1553–1559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marasco, W.A.; Sui, J. The growth and potential of human antiviral monoclonal antibody therapeutics. Nat. Biotechnol. 2007, 25, 1421–1434. [Google Scholar] [CrossRef] [PubMed]
- Ekiert, D.C.; Bhabha, G.; Elsliger, M.-A.; Friesen, R.H.E.; Jongeneelen, M.; Throsby, M.; Goudsmit, J.; Wilson, I.A. Antibody recognition of a highly conserved influenza virus epitope. Science 2009, 324, 246–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ekiert, D.C.; Friesen, R.H.; Bhabha, G.; Kwaks, T.; Jongeneelen, M.; Yu, W.; Ophorst, C.; Cox, F.; Korse, H.J.; Brandenburg, B.; et al. A highly conserved neutralizing epitope on group 2 influenza A viruses. Science 2011, 333, 843–850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friesen, R.H.; Lee, P.S.; Stoop, E.J.; Hoffman, R.M.; Ekiert, D.C.; Bhabha, G.; Yu, W.; Juraszek, J.; Koudstaal, W.; Jongeneelen, M.; et al. A common solution to group 2 influenza virus neutralization. Proc. Natl. Acad. Sci. USA 2014, 111, 445–450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kajihara, M.; Marzi, A.; Nakayama, E.; Noda, T.; Kuroda, M.; Manzoor, R.; Matsuno, K.; Feldmann, H.; Yoshida, R.; Kawaoka, Y.; et al. Inhibition of Marburg virus budding by nonneutralizing antibodies to the envelope glycoprotein. J. Virol. 2012, 86, 13467–13474. [Google Scholar] [CrossRef] [Green Version]
- Qiu, X.; Audet, J.; Wong, G.; Fernando, L.; Bello, A.; Pillet, S.; Alimonti, J.B.; Kobinger, G.P. Sustained protection against Ebola virus infection following treatment of infected nonhuman primates with ZMAb. Sci. Rep. 2013, 3, 3365. [Google Scholar] [CrossRef] [Green Version]
- Ruprecht, R.M. Passive immunization with human neutralizing monoclonal antibodies against HIV-1 in macaque models: Experimental approaches. Methods Mol. Biol. 2009, 525, 559–566. [Google Scholar]
- Llorens, X.S.; Castaño, E.; Null, D.; Steichen, J.; Sánchez, P.J.; Ramilo, O.; Top, F.H.; Connor, E. Safety and pharmacokinetics of an intramuscular humanized monoclonal antibody to respiratory syncytial virus in premature infants and infants with bronchopulmonary dysplasia. The MEDI-493 Study Group. Pediatr. Infect. Dis. J. 1998, 17, 787–791. [Google Scholar] [CrossRef]
- Behring, E.A.; Kitasato, S. Ueber das zustandekommen der diptherie-immunität und der tetanus-immunität bei thieren. Deutch. Med. Woch. 1890, 49, 1113–1114. [Google Scholar]
- Casadevall, A.; Dadachova, E.; Pirofski, L.-A. Passive antibody therapy for infectious diseases. Nat. Rev. Microbiol. 2004, 2, 695–703. [Google Scholar] [CrossRef] [PubMed]
- Kluge, T. Gamma-Globulin in the Prevention on Viral Hepatitis: A Study of the Effect of Medium-Size Doses. Acta Med. Scand. 1963, 174, 469–477. [Google Scholar] [CrossRef] [PubMed]
- Hammond, D. CytoGam infusions at home. J. Infus. Nurs. 1999, 22, 331–335. [Google Scholar]
- Oertel, M.D. RespiGam: An RSV immune globulin. Pediatr. Nurs. 1996, 22, 525–528. [Google Scholar] [PubMed]
- Hampson, K.; Coudeville, L.; Lembo, T.; Sambo, M.; Kieffer, A.; Attlan, M.; Barrat, J.; Blanton, J.D.; Briggs, D.J.; Cleaveland, S.; et al. Estimating the global burden of endemic canine rabies. PLoS Negl. Trop. Dis. 2015, 9, e0003709. [Google Scholar]
- WHO. Rabies Vaccines: WHO Position Paper—April 2018; Abela Ridder, B., Ed.; World Health Organization: Geneva, Switzerland, 2018; pp. 201–220. [Google Scholar]
- De Kruif, J.; Bakker, A.B.; Marissen, W.E.; Kramer, R.A.; Throsby, M.; Rupprecht, C.E.; Goudsmit, J. A human monoclonal antibody cocktail as a novel component of rabies postexposure prophylaxis. Annu. Rev. Med. 2007, 58, 359–368. [Google Scholar] [CrossRef]
- Bakker, A.B.H.; Marissen, W.E.; Kramer, R.A.; Rice, A.B.; Weldon, W.C.; Niezgoda, M.; Hanlon, C.A.; Thijsse, S.; Backus, H.H.J.; de Kruif, J.; et al. Novel human monoclonal antibody combination effectively neutralizing natural rabies virus variants and individual in vitro escape mutants. J. Virol. 2005, 79, 9062–9068. [Google Scholar] [CrossRef] [Green Version]
- Bakker, A.; Python, C.; Kissling, C.; Pandya, P.; Marissen, W.; Brink, M.; Lagerwerf, F.; Worst, S.; van Corven, E.; Kostense, S.; et al. First administration to humans of a monoclonal antibody cocktail against rabies virus: Safety, tolerability, and neutralizing activity. Vaccine 2008, 26, 5922–5927. [Google Scholar] [CrossRef] [Green Version]
- Smith, J.S.; Yager, P.A.; Baer, G.M. A rapid reproducible test for determining rabies neutralizing antibody. Bull. World Health Organ. 1973, 48, 535–541. [Google Scholar]
- Kostense, S.; Moore, S.; Companjen, A.; Bakker, A.B.H.; Marissen, W.E.; von Eyben, R.; Weverling, G.J.; Hanlon, C.; Goudsmit, J. Validation of the rapid fluorescent focus inhibition test for rabies virus-neutralizing antibodies in clinical samples. Antimicrob. Agents Chemother. 2012, 56, 3524–3530. [Google Scholar] [CrossRef] [Green Version]
- Marissen, W.E.; Kramer, R.A.; Rice, A.; Weldon, W.C.; Niezgoda, M.; Faber, M.; Slootstra, J.W.; Meloen, R.H.; der Horst, M.C.-V.; Visser, T.J.; et al. Novel rabies virus-neutralizing epitope recognized by human monoclonal antibody: Fine mapping and escape mutant analysis. J. Virol. 2005, 79, 4672–4678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- FDA. Guidance for Industry, Bioanalytical Method Validation; FDA: Silver Spring, MD, USA, 2001.
- EMA. Guideline on Bioanalytical Method Validation, in EMEA/CHMP/EWP/192217/2009; EMA: Amsterdam, The Netherlands, 2012. [Google Scholar]
- ICH. Validation of Analytical Procedures: Text and Methodology Q2(R1); ICH: Geneva, Switzerland, 2005. [Google Scholar]
- Hendriks, J.; Stals, C.; Versteilen, A.; Mommaas, B.; Verhoeven, M.; Tirion, F.; ter Haak, M.; Ribbens, W.; Bosch, M.; Trommel, M.; et al. Stability studies of binding and functional anti-vaccine antibodies. Bioanalysis 2014, 6, 1385–1393. [Google Scholar] [CrossRef] [PubMed]
- Michaut, L.; Laurent, N.; Kentsch, K.; Spindeldreher, S.; Deckert-Salva, F. Stability of anti-immunotherapeutic antibodies in frozen human serum samples. Bioanalysis 2014, 6, 1395–1407. [Google Scholar] [CrossRef] [PubMed]
- Pihl, S.; Michaut, L.; Hendriks, J.; Loebbert, R.; Ryding, J.; Nemansky, M.; Vermet, L.; Companjen, A. EBF recommendation for stability testing of anti-drug antibodies; lessons learned from anti-vaccine antibody stability studies. Bioanalysis 2014, 6, 1409–1413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WHO. WHO Expert Consultation on Rabies. In WHO Technical Report Series; No. 982; WHO: Geneva, Switzerland, 2013. [Google Scholar]
- Smith, J.S.; Yager, P.A.; Baer, G.M. A rapid fluorescent focus inhibition test (RFFIT) for determining rabies virus-neutralizing antibody. In Laboratory Techniques in Rabies; Meslin, F.X., Kaplan, M.M., Koprowski, H., Eds.; World Health Organization: Geneva, Switzerland, 1996; pp. 181–192. [Google Scholar]
- Habel, K. Habel test for potency. In Laboratory Techniques in Rabies; Meslin, F.X., Kaplan, M.M., Koprowski, H., Eds.; World Health Organization: Geneva, Switzerland, 1996; pp. 369–373. [Google Scholar]
- WHO. WHO Expert Committee on Biological Standardization, Sixty-First Report; World Health Organization: Geneva, Switzerland, 2013. [Google Scholar]
- Chaloner-Larsson, G.; Anderson, R.; Egan, A. A WHO Guide to Goo Manufacturing Practice (GMP) Requirements; Part: Validation; World Health Organization: Geneva, Switzerland, 1997. [Google Scholar]
- Tan, C.Y.; Iglewicz, B. Measurement-methods comparisions and linear statistical relationship. Technometrics 1999, 41, 192–201. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Subject | Experiment Number | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | |||||||||
| Day | 1 | 5 | 2 | 6 | 2 | 6 | 3 | 7 | 3 | 7 | 4 | 8 | 4 | 8 | 1 | 5 |
| Virus | E57 | E98 | E98 | E57 | E57 | E98 | E98 | E57 | E57 | E98 | E98 | E57 | E57 | E98 | E98 | E57 |
| Operator | 1 | 2 | 1 | 2 | 1 | 2 | 1 | 2 | ||||||||
| CP# | early | late | late | early | early | late | late | early | ||||||||
| Data collection for: | Repeatability Intermediate precision Accuracy/ Linearity LOD/LLOQ Bench top stability | Repeatability Intermediate precision Accuracy/ Linearity LOD/LLOQ Stability at −20 °C | Repeatability Intermediate precision Accuracy/ Linearity LOD/LLOQ | Repeatability Intermediate precision Accuracy/ Linearity LOD/LLOQ Specificity | Repeatability Intermediate precision Accuracy/ Linearity LOD/LLOQ Freeze/Thaw stability | Repeatability Intermediate precision Accuracy/ Linearity LOD/LLOQ | Repeatability Intermediate precision Accuracy/ Linearity LOD/LLOQ | Repeatability Intermediate precision Accuracy/ Linearity LOD/LLOQ | ||||||||
| Spike (IU/mL) | E57 Virus | E98 Virus | ||||||
|---|---|---|---|---|---|---|---|---|
| Pred. | Obs. | L90%CI | U90%CI | Pred. | Obs. | L90%CI | U90%CI | |
| 0.025 | 2.8 | 3.5 | 3.2 | 3.8 | 4.2 | 4.7 | 4.2 | 5.2 |
| 0.05 | 4.9 | 4.8 | 4.4 | 5.2 | 7.1 | 7.0 | 6.3 | 7.9 |
| 0.1 | 8.6 | 7.5 | 7.0 | 8.1 | 11.9 | 11.2 | 10.3 | 12.2 |
| 0.2 | 15.0 | 13.2 | 12.1 | 14.4 | 19.9 | 19.2 | 17.3 | 21.3 |
| 0.5 | 31.3 | 27.5 | 25.2 | 30.1 | 39.2 | 35.6 | 32.2 | 39.3 |
| 0.8 | 45.7 | 44.4 | 41.7 | 47.3 | 55.7 | 52.6 | 48.9 | 56.6 |
| 1.0 | 54.6 | 57.1 | 51.8 | 62.9 | 65.7 | 67.5 | 61.0 | 74.7 |
| 2.0 | 95.4 | 113.8 | 103.4 | 125.2 | 110.0 | 124.4 | 111.1 | 139.3 |
| Spike (IU/mL) | E57 Virus | E98 Virus | ||||
|---|---|---|---|---|---|---|
| Mean RVNAT | %CV R | %CV IP | Mean RVNAT | %CV R | %CV IP | |
| Overall | 18.4 | 13.2 | 20.5 | 24.0 | 13.5 | 24.6 |
| 0.025 | 3.5 | 14.4 | 19.7 | 4.7 | 14.0 | 24.4 |
| 0.05 | 4.8 | 13.0 | 19.5 | 7.0 | 18.6 | 27.7 |
| 0.1 | 7.5 | 17.2 | 17.9 | 11.2 | 12.0 | 20.9 |
| 0.2 | 13.2 | 11.4 | 20.9 | 19.2 | 11.8 | 26.2 |
| 0.5 | 27.5 | 8.6 | 22.1 | 35.6 | 11.6 | 24.9 |
| 0.8 | 44.4 | 8.2 | 15.6 | 52.6 | 8.1 | 18.1 |
| 1.0 | 57.1 | 11.5 | 24.1 | 67.5 | 10.6 | 25.3 |
| 2.0 | 113.8 | 13.1 | 23.7 | 124.4 | 17.7 | 28.0 |
| Virus Type | Stability Item | Mean RVNAT CS | Mean RVNAT TS | Ratio TS/CS | Lower 90% CI Ratio | Upper 90% CI Ratio | Validation Status |
|---|---|---|---|---|---|---|---|
| E57 | Bench top | 49.38 | 49.54 | 100.3% | 94.1% | 107.0% | P |
| −20 °C | 59.44 | 58.17 | 97.9% | 91.6% | 104.7% | P | |
| F/T 1 | 59.44 | 56.24 | 94.6% | 88.6% | 101.1% | P | |
| F/T 2 | 59.44 | 59.89 | 100.8% | 94.3% | 107.7% | P | |
| F/T 3 | 34.78 | 35.10 | 100.9% | 96.4% | 105.7% | P | |
| E98 | Bench top | 46.52 | 48.64 | 104.6% | 98.9% | 110.5% | P |
| −20 °C | 54.01 | 59.30 | 109.8% | 103.2% | 116.8% | P | |
| F/T 1 | 54.01 | 58.97 | 109.2% | 102.6% | 116.2% | P | |
| F/T 2 | 54.01 | 59.04 | 109.3% | 102.8% | 116.3% | P | |
| F/T 3 | 38.03 | 39.56 | 104.0% | 99.3% | 109.2% | P |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Companjen, A.; Moore, S.M.; Boulanger, B.; Kostense, S.; Marissen, W.E. Validation of Adapted Neutralization Assays Developed to Discriminate Anti-Rabies Virus Activity of Two Different Anti-Rabies Virus Monoclonal Antibodies Administered as a Combination. Biologics 2023, 3, 11-22. https://doi.org/10.3390/biologics3010002
Companjen A, Moore SM, Boulanger B, Kostense S, Marissen WE. Validation of Adapted Neutralization Assays Developed to Discriminate Anti-Rabies Virus Activity of Two Different Anti-Rabies Virus Monoclonal Antibodies Administered as a Combination. Biologics. 2023; 3(1):11-22. https://doi.org/10.3390/biologics3010002
Chicago/Turabian StyleCompanjen, Arjen, Susan M. Moore, Bruno Boulanger, Stefan Kostense, and Wilfred E. Marissen. 2023. "Validation of Adapted Neutralization Assays Developed to Discriminate Anti-Rabies Virus Activity of Two Different Anti-Rabies Virus Monoclonal Antibodies Administered as a Combination" Biologics 3, no. 1: 11-22. https://doi.org/10.3390/biologics3010002