Xeno-miRs and Circulating miRNAs as Novel Biomarkers in Certain Diseases




1. miRNAs
2. Dietary xeno-miRNAs
3. miRNAs and Their Functions and Potential Uses in Diseases
4. The Effects of Diet on miRNAs
5. Conclusions and Recommendations
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Wang, Y.; Stricker, H.M.; Gou, D.; Liu, L. MicroRNA: Past and present. Front. Biosci. Landmark 2007, 12, 2316–2329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, L.; Jiang, F. MicroRNA (miRNA) profiling. Cancer Gene Profiling 2016, 1381, 151–161. [Google Scholar]
- Chuang, J.C.; Jones, P. Epigenetics and microRNAs. Pediatr. Res. 2007, 61, 24–29. [Google Scholar] [CrossRef] [PubMed]
- Hammond, S.M. An overview of microRNAs. Adv. Drug Deliv. Rev. 2015, 87, 3–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wahid, F.; Shehzad, A.; Khan, T.; Kim, Y.Y. MicroRNAs: Synthesis, mechanism, function, and recent clinical trials. Biochim. Biophys. Acta BBA Mol. Cell Res. 2010, 1803, 1231–1243. [Google Scholar] [CrossRef] [Green Version]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of microRNA biogenesis, mechanisms of actions, and circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef] [Green Version]
- Yang, B.-f.; Ai, J. MicroRNA transport: A new way in cell communication. J. Cell. Physiol. 2013, 228, 1713–1719. [Google Scholar]
- Mohr, A.M.; Mott, J. Overview of microRNA biology. Semin. Liver Dis. 2015, 35, 3–11. [Google Scholar]
- Becker, N.; Lockwood, C. Pre-analytical variables in miRNA analysis. Clin. Biochem. 2013, 46, 861–868. [Google Scholar] [CrossRef]
- Huang, Y.; Shen, X.J.; Zou, Q.; Wang, S.P.; Tang, S.M.; Zhang, G.Z. Biological functions of microRNAs: A review. J. Physiol. Biochem. 2011, 67, 129–139. [Google Scholar] [CrossRef]
- Fan, Y.; Habib, M.; Xia, J. Xeno-miRNet: A comprehensive database and analytics platform to explore xeno-miRNAs and their potential targets. PeerJ 2018, 6, e5650. [Google Scholar] [CrossRef]
- Anfossi, S.; Babayan, A.; Pantel, K.; Calin, G.A. Clinical utility of circulating non-coding RNAs—An update. Nat. Rev. Clin. Oncol. 2018, 15, 541–563. [Google Scholar] [CrossRef]
- Díez-Sainz, E.; Lorente-Cebrián, S.; Aranaz, P.; Riezu-Boj, J.I.; Martínez, J.A.; Milagro, F.I. Potential Mechanisms Linking Food-Derived MicroRNAs, Gut Microbiota and Intestinal Barrier Functions in the Context of Nutrition and Human Health. Front. Nutr. 2021, 8, 586564. [Google Scholar] [CrossRef]
- Fabris, L.; Calin, G. Circulating free xeno-microRNAs—The new kids on the block. Mol. Oncol. 2016, 10, 503–508. [Google Scholar] [CrossRef]
- Tingö, L.; Ahlberg, E.; Johansson, L.; Pedersen, S.A.; Chawla, K.; Sætrom, P.; Cione, E.; Simpson, M.R. Non-coding RNAs in human breast milk: A systematic review. Front. Immunol. 2021, 12, 3522. [Google Scholar] [CrossRef]
- Stephen, B.J.; Pareek, N.; Saeed, M.; Kausar, M.A.; Rahman, S.; Datta, M. Xeno-miRNA in maternal-infant immune crosstalk: An aid to disease alleviation. Front. Immunol. 2020, 11, 404. [Google Scholar] [CrossRef] [Green Version]
- Zhu, W.-J.; Liu, Y.; Cao, Y.N.; Peng, L.X.; Yan, Z.Y.; Zhao, G. Insights into Health-Promoting Effects of Plant MicroRNAs: A Review. J. Agric. Food Chem. 2021, 69, 14372–14386. [Google Scholar] [CrossRef]
- Lukasik, A.; Zielenkiewicz, P. Plant MicroRNAs—Novel Players in Natural Medicine? Int. J. Mol. Sci. 2017, 18, 9. [Google Scholar] [CrossRef] [Green Version]
- Wagner, A.E.; Piegholdt, S.; Ferraro, M.; Pallauf, K.; Rimbach, G. Food derived microRNAs. Food Funct. 2015, 6, 714–718. [Google Scholar] [CrossRef] [Green Version]
- Link, J.; Thon, C.; Schanze, D.; Steponaitiene, R.; Kupcinskas, J.; Zenker, M.; Canbay, A.; Malfertheiner, P.; Link, A. Food-Derived Xeno-microRNAs: Influence of Diet and Detectability in Gastrointestinal Tract—Proof-of-Principle Study. Mol. Nutr. Food Res. 2019, 63, 1800076. [Google Scholar] [CrossRef]
- Zhou, M.; Kara, H.; Dai, Y.; Mou, L.; Cooper, D.K.C.; Wu, C.; Cai, Z. Circulating Organ-Specific MicroRNAs Serve as Biomarkers in Organ-Specific Diseases: Implications for Organ Allo- and Xeno-Transplantation. Int. J. Mol. Sci. 2016, 17, 1232. [Google Scholar] [CrossRef]
- Turchinovich, A.; Samatov, T.R.; Tonevitsky, A.G.; Burwinkel, B. Circulating miRNAs: Cell–cell communication function? Front. Genet. 2013, 4, 119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reid, G.; Kirschner, M.; van Zandwijk, N. Circulating microRNAs: Association with disease and potential use as biomarkers. Crit. Rev. Oncol. Hematol. 2011, 80, 193–208. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.; Soh, J.; Ying, J. A microarray platform for detecting disease-specific circulating miRNA in human serum. Biosens. Bioelectron. 2016, 75, 238–246. [Google Scholar] [CrossRef]
- Wei, C.; Henderson, H.; Spradley, C.; Li, L.; Kim, I.K.; Kumar, S.; Hong, N.; Arroliga, A.C.; Gupta, S. Circulating miRNAs as Potential Marker for Pulmonary Hypertension. PLoS ONE 2013, 8, e64396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldberg, L.; Tirosh-Wagner, T.; Vardi, A.; Abbas, H.; Pillar, N.; Shomron, N.; Nevo-Caspi, Y.; Paret, G. Circulating MicroRNAs: A Potential Biomarker for Cardiac Damage, Inflammatory Response, and Left Ventricular Function Recovery in Pediatric Viral Myocarditis. J. Cardiovasc. Transl. Res. 2018, 11, 319–328. [Google Scholar] [CrossRef]
- Jaeger, A.; Zollinger, L.; Saely, C.H.; Muendlein, A.; Evangelakos, I.; Nasias, D.; Charizopoulou, N.; Schofield, J.D.; Othman, A.; Soran, H.; et al. Circulating microRNAs -192 and -194 are associated with the presence and incidence of diabetes mellitus. Sci. Rep. 2018, 8, 14274. [Google Scholar] [CrossRef] [Green Version]
- Thomou, T.; Mori, M.A.; Dreyfuss, J.M.; Konishi, M.; Sakaguchi, M.; Wolfrum, C.; Rao, T.N.; Winnay, J.N.; Garcia-Martin, R.; Grinspoon, S.K.; et al. Adipose-derived circulating miRNAs regulate gene expression in other tissues. Nature 2017, 542, 450–455. [Google Scholar] [CrossRef] [Green Version]
- Rottiers, V.; Näär, A. MicroRNAs in metabolism and metabolic disorders. Nat. Rev. Mol. Cell Biol. 2012, 13, 239–250. [Google Scholar] [CrossRef] [Green Version]
- Shi, S.B.; Wang, M.; Tian, J.; Li, R.; Chang, C.X.; Qi, J.L. MicroRNA 25, microRNA 145, and microRNA 210 as biomarkers for predicting the efficacy of maintenance treatment with pemetrexed in lung adenocarcinoma patients who are negative for epidermal growth factor receptor mutations or anaplastic lymphoma kinase translocations. Transl. Res. 2016, 170, 1–7. [Google Scholar]
- Fan, B.; Shen, C.; Wu, M.; Zhao, J.; Guo, Q.; Luo, Y. miR-17-92 cluster is connected with disease progression and oxaliplatin/capecitabine chemotherapy efficacy in advanced gastric cancer patients: A preliminary study. Medicine 2018, 97, e12007. [Google Scholar] [CrossRef]
- Srivastava, A.; Goldberger, H.; Dimtchev, A.; Ramalinga, M.; Chijioke, J.; Marian, C.; Oermann, E.K.; Uhm, S.; Kim, J.S.; Chen, L.N.; et al. MicroRNA profiling in prostate cancer—The diagnostic potential of urinary miR-205 and miR-214. PLoS ONE 2013, 8, e76994. [Google Scholar] [CrossRef] [Green Version]
- Jiménez-Lucena, R.; Camargo, A.; Alcalá-Diaz, J.F.; Romero-Baldonado, C.; Luque, R.M.; Ommen, B.V.; Delgado-Lista, J.; Ordovás, J.M.; Pérez-Martínez, P.; Rangel-Zúñiga, O.A.; et al. A plasma circulating miRNAs profile predicts type 2 diabetes mellitus and prediabetes: From the CORDIOPREV study. Exp. Mol. Med. 2018, 50, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Corral-Fernández, N.E.; Salgado-Bustamante, M.; Martínez-Leija, M.E.; Cortez-Espinosa, N.; García-Hernández, M.H.; Reynaga-Hernández, E.; Quezada-Calvillo, R.; Portales-Pérez, D.P. Dysregulated miR-155 expression in peripheral blood mononuclear cells from patients with type 2 diabetes. Exp. Clin. Endocrinol. Diabetes 2013, 121, 347–353. [Google Scholar] [CrossRef]
- Mensà, E.; Giuliani, A.; Matacchione, G.; Gurău, F.; Bonfigli, A.R.; Romagnoli, F.; Luca, M.D.; Sabbatinelli, J.; Olivieri, F. Circulating miR-146a in healthy aging and type 2 diabetes: Age- and gender-specific trajectories. Mech. Ageing Dev. 2019, 180, 1–10. [Google Scholar] [CrossRef]
- Cortez-Dias, N.; Costa, M.C.; Carrilho-Ferreira, P.; Silva, D.; Jorge, C.; Calisto, C.; Pessoa, T.; Martins, S.R.; Sousa, J.C.D.; Silva, P.C.D.; et al. Circulating miR-122-5p/miR-133b Ratio Is a Specific Early Prognostic Biomarker in Acute Myocardial Infarction. Circ. J. 2016, 80, 2183–2191. [Google Scholar] [CrossRef] [Green Version]
- Vegter, E.L.; Schmitter, D.; Hagemeijer, Y.; Ovchinnikova, E.S.; Harst, P.V.D.; Teerlink, J.R.; O’Connor, C.M.; Metra, M.; Davison, B.A.; Bloomfield, D.; et al. Use of biomarkers to establish potential role and function of circulating microRNAs in acute heart failure. Int. J. Cardiol. 2016, 224, 231–239. [Google Scholar] [CrossRef] [Green Version]
- Tabuchi, T.; Satoh, M.; Itoh, T.; Nakamura, M. MicroRNA-34a regulates the longevity-associated protein SIRT1 in coronary artery disease: Effect of statins on SIRT1 and microRNA-34a expression. Clin. Sci. 2012, 123, 161–171. [Google Scholar] [CrossRef]
- Shen, Z.; Tang, W.; Guo, J.; Sun, S. miR-483-5p plays a protective role in chronic obstructive pulmonary disease. Int. J. Mol. Med. 2017, 40, 193–200. [Google Scholar] [CrossRef] [Green Version]
- Zhao, T.; Zheng, Y.L.; Hao, D.Z.; Jin, X.; Luo, Q.Z.; Guo, Y.T.; Li, D.X.; Xi, W.; Xu, Y.; Chen, Y.S.; et al. Blood circRNAs as biomarkers for the diagnosis of community-acquired pneumonia. J. Cell Biochem. 2019, 120, 16483–16494. [Google Scholar] [CrossRef]
- Chen, Y.J.; Zhu, J.M.; Wu, H.; Fan, J.; Zhou, J.; Hu, J.; Yu, Q.; Liu, T.T.; Yang, L.; Wu, C.L.; et al. Circulating microRNAs as a Fingerprint for Liver Cirrhosis. PLoS ONE 2013, 8, e66577. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Fu, T.; Guo, Z.; Zhang, Y.; Zhang, L.; Su, H.; Long, Y.; Ji, Z.; Yan, Y.; Shao, Z. Serum miR-375 Levels Are Closely Related to Disease Progression from HBV Infection to HBV-Related Hepatocellular Carcinoma. Biomed. Res. Int. 2020, 2020, 5819385. [Google Scholar] [CrossRef] [PubMed]
- Derkow, K.; Rössling, R.; Schipke, C.; Krüger, C.; Bauer, J.; Fähling, M.; Stroux, A.; Schott, E.; Ruprecht, K.; Peters, O.; et al. Distinct expression of the neurotoxic microRNA family let-7 in the cerebrospinal fluid of patients with Alzheimer’s disease. PLoS ONE 2018, 13, e0200602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grossi, I.; Radeghieri, A.; Paolini, L.; Porrini, V.; Pilotto, A.; Padovani, A.; Marengoni, A.; Barbon, A.; Bellucci, A.; Pizzi, M.; et al. MicroRNA-34a-5p expression in the plasma and in its extracellular vesicle fractions in subjects with Parkinson’s disease: An exploratory study. Int. J. Mol. Med. 2021, 47, 533–546. [Google Scholar] [CrossRef] [PubMed]
- Farroni, C.; Marasco, E.; Marcellini, V.; Giorda, E.; Valentini, D.; Petrini, S.; D’Oria, V.; Pezzullo, V.; Cascioli, S.; Scarsella, M.; et al. Dysregulated miR-155 and miR-125b Are Related to Impaired B-cell Responses in Down Syndrome. Front. Immunol. 2018, 9, 2683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, J.; Kong, M.; Ye, Y.; Hong, S.; Cheng, L.; Jiang, L. Serum miR-206 and other muscle-specific microRNAs as non-invasive biomarkers for Duchenne muscular dystrophy. J. Neurochem. 2014, 129, 877–883. [Google Scholar] [CrossRef]
- Ishida, M.; Selaru, F. miRNA-Based Therapeutic Strategies. Curr. Pathobiol. Rep. 2013, 1, 63–70. [Google Scholar] [CrossRef]
- Zempleni, J.; Baier, S.R.; Howard, K.M.; Cui, J. Gene regulation by dietary microRNAs. Can. J. Physiol. Pharmacol. 2015, 93, 1097–1102. [Google Scholar] [CrossRef] [Green Version]
- Fu, X.; Dong, B.; Tian, Y.; Lefebvre, P.; Meng, Z.; Wang, X.; Pattou, F.; Han, W.; Wang, X.; Lou, F.; et al. MicroRNA-26a regulates insulin sensitivity and metabolism of glucose and lipids. J. Clin. Invest. 2015, 125, 2497–2509. [Google Scholar] [CrossRef] [Green Version]
- Baier, S.R.; Nguyen, C.; Xie, F.; Wood, J.R.; Zempleni, J. MicroRNAs are absorbed in biologically meaningful amounts from nutritionally relevant doses of cow milk and affect gene expression in peripheral blood mononuclear cells, HEK-293 kidney cell cultures, and mouse livers. J. Nutr. 2014, 144, 1495–1500. [Google Scholar] [CrossRef] [Green Version]
- Díez-Ricote, L.; Ruiz-Valderrey, P.; Micó, V.; Blanco-Rojo, R.; Tomé-Carneiro, J.; Dávalos, A.; Ordovás, J.M.; Daimiel, L. Trimethylamine n-Oxide (TMAO) Modulates the Expression of Cardiovascular Disease-Related microRNAs and Their Targets. Int. J. Mol. Sci. 2021, 22, 11145. [Google Scholar] [CrossRef]
- Gil-Zamorano, J.; Tomé-Carneiro, J.; Hazas, M.C.L.D.L.; Pozo-Acebo, L.D.; Crespo, M.C.; Gómez-Coronado, D.; Chapado, L.A.; Herrera, E.; Latasa, M.J.; Ruiz-Roso, M.B.; et al. Intestinal miRNAs regulated in response to dietary lipids. Sci. Rep. 2020, 10, 18921. [Google Scholar] [CrossRef] [PubMed]
- Dambal, S.; Giangreco, A.A.; Acosta, A.M.; Fairchild, A.; Richards, Z.; Deaton, R.; Wagner, D.; Vieth, R.; Gann, P.H.; Kajdacsy-Balla, A.; et al. microRNAs and DICER1 are regulated by 1,25-dihydroxyvitamin D in prostate stroma. J. Steroid Biochem. Mol. Biol. 2017, 167, 192–202. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, H.D.; Kim, M.-S. The role of mixed B vitamin intakes on cognitive performance: Modeling, genes and miRNAs involved. J. Psychiatr. Res. 2022, 152, 38–56. [Google Scholar] [CrossRef]
- Vertuani, S.; Angusti, A.; Manfredini, S. The Antioxidants and Pro-Antioxidants Network: An Overview. Curr. Pharm. Des. 2004, 10, 1677–1694. [Google Scholar] [CrossRef] [Green Version]
- Alehagen, U.; Johansson, P.; Aaseth, J.; Alexander, J.; Wågsäter, D. Significant changes in circulating microRNA by dietary supplementation of selenium and coenzyme Q10 in healthy elderly males. A subgroup analysis of a prospective randomized double-blind placebo-controlled trial among elderly Swedish citizens. PLoS ONE 2017, 12, e0174880. [Google Scholar] [CrossRef] [Green Version]
- Tian, F.Y.; Kennedy, E.M.; Hermetz, K.; Burt, A.; Everson, T.M.; Punshon, T.; Jackson, B.P.; Hao, K.; Chen, J.; Karagas, M.R.; et al. Selenium-associated differentially expressed microRNAs and their targeted mRNAs across the placental genome in two U.S. birth cohorts. Epigenetics 2022, 17, 1234–1245. [Google Scholar] [CrossRef]
- Kiełczykowska, M.; Kocot, J.; Paździor, M.; Musik, I. Selenium—A fascinating antioxidant of protective properties. Adv. Clin. Exp. Med. 2018, 27, 245–255. [Google Scholar] [CrossRef]
- Hargreaves, I.; Heaton, R.; Mantle, D. Disorders of Human Coenzyme Q10 Metabolism: An Overview. Int. J. Mol. Sci. 2020, 21, 6695. [Google Scholar] [CrossRef]
- Bonakdar, R.A.; Guarneri, E. Coenzyme Q10. Am. Fam. Physician 2005, 72, 1065–1070. [Google Scholar]
- Mihelich, B.L.; Khramtsova, E.A.; Arva, N.; Vaishnav, A.; Johnson, D.N.; Giangreco, A.A.; Martens-Uzunova, E.; Bagasra, O.; Kajdacsy-Balla, A.; Nonn, L. miR-183-96-182 cluster is overexpressed in prostate tissue and regulates zinc homeostasis in prostate cells. J. Biol. Chem. 2011, 286, 44503–44511. [Google Scholar] [CrossRef] [Green Version]
- Shi, C.Y.; Kingston, E.R.; Kleaveland, B.; Lin, D.H.; Stubna, M.W.; Bartel, D.P. The ZSWIM8 ubiquitin ligase mediates target-directed microRNA degradation. Science 2020, 370, eabc9359. [Google Scholar] [CrossRef] [PubMed]


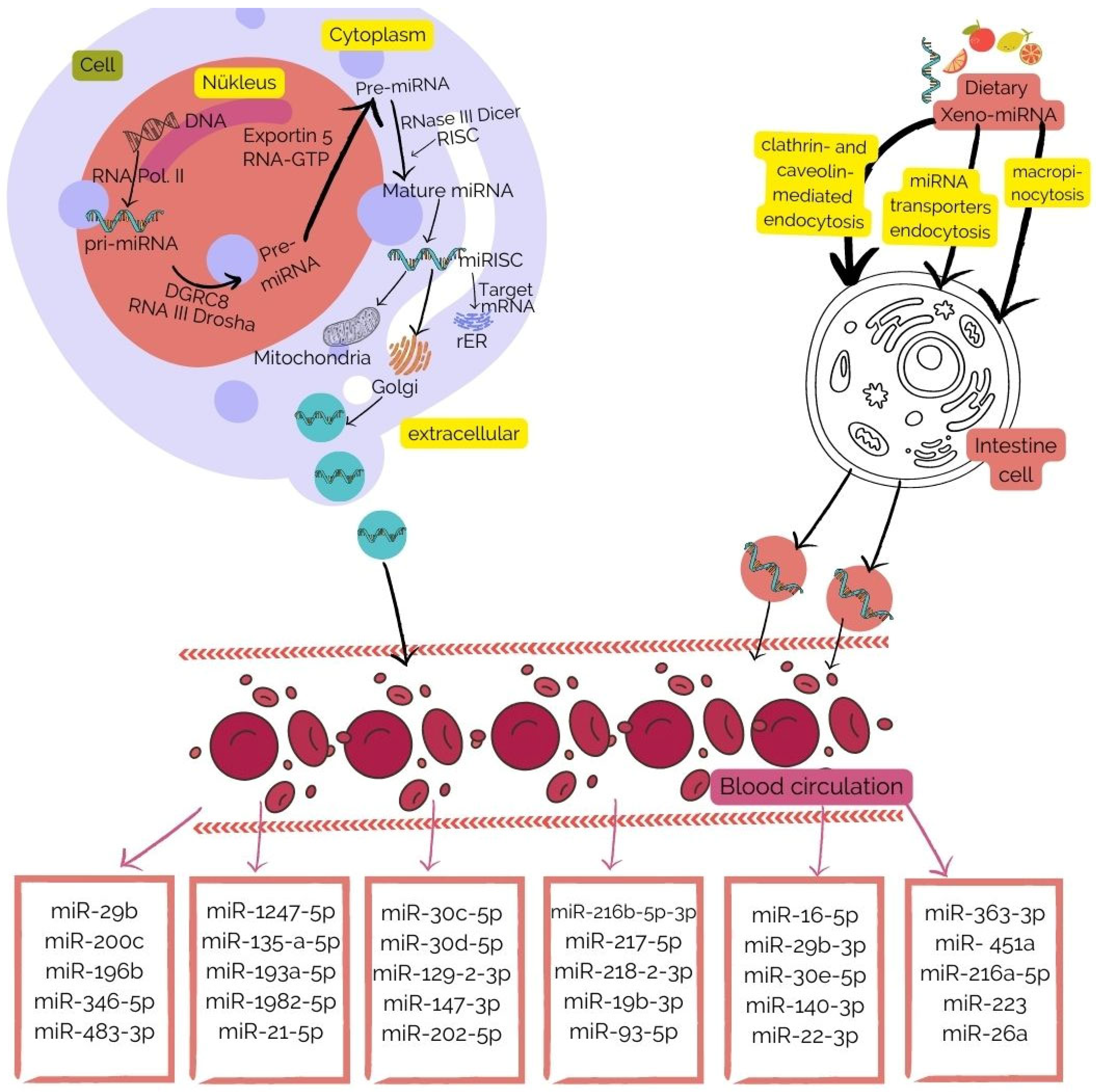{kind=link}
| Disease | Type of miRNA | Function | Up/Down-Regulate | Reference |
|---|---|---|---|---|
| Stage IIIb or IV lung adenocarcinoma | miR-25, miR-145 and miR-210 | Biomarkers in predicting the effectiveness of pemetrexed treatment | - | [30] |
| Stomach cancer | miR-17-92 | Markers in the prognosis of advanced stomach cancer and the effectiveness of chemotherapy | Plasma miR-17-92 in advanced stomach cancer ↑, ↓ after chemotherapy Plasma miR-17-92 in chemo-resistant patients ↔ | [31] |
| Prostate cancer | miR-205 miR-214 | Diagnostic biomarkers | Tissue miR-205 and miR-214 ↓ | [32] |
| Type 2 diabetes mellitus | miR-150 miR-30a-5p miR-15a miR-375 | Diagnostic marker of prediabetes | miR-150 and miR-30a-5p ↑ miR-15a and miR-375 ↓ | [33] |
| Type 2 diabetes mellitus | miR-155 | Marker of pathogenesis and metabolic control | miR-155 ↓ | [34] |
| Aging and Type 2 diabetes mellitus | miR-146a | Marker utilized through the detection of circulating miRNA | miR-146a with aging ↓ miR-146a with aging in T2DM patients ↓ miR-146a in T2DM patients treated with metformin ↑ | [35] |
| Acute myocardial infarction | miR-122-5p/133b ratio | Prognostic biomarker | - | [36] |
| Acute heart failure | miR-16-5p miR-106a-5p miR-223-3p miR-652-3p miR-199a-3p miR-18a-5p | Relationship with biochemical pathways in heart diseases (CRP, creatinine, growth differential factor soluble ST-2, procalcitonin, galectin-3) | - | [37] |
| Coronary artery disease | miR-34a | Expression in endothelial progenitor cells | miR-34a ↑ | [38] |
| Chronic obstructive pulmonary disease (COPD) | miR-489-5p | Early detection biomarker and therapy tool | miR-489-5p ↓ | [39] |
| Pneumonia | hsa_circ_0018429 hsa_circ_0026579 hsa_circ_0125357 hsa_circ_0099188 | Sensitive and specific biomarker for diagnosis | - | [40] |
| Liver cirrhosis | miR-106b miR-181b | Sensitive and specific clinical diagnostic biomarker at early stages | - | [41] |
| Hepatitis B Virus (HBV) | miR-375 | Early predictive marker in the prognosis of hepatocellular carcinoma due to HBV-associated hepatitis or cirrhosis | Serum miR-375 ↓ | [42] |
| Alzheimer’s disease | let-7b let-7e | Indicator of neuropathological pathways | Cerebrospinal fluid let-7b and let-7e ↑ | [43] |
| Parkinson’s disease | miR-34a-5p | Diagnostic biomarker | miR-34a-5p in small extracellular vesicles free of exogenous protein contaminants ↑ | [44] |
| Down syndrome | miR-155 miR-125b | Down syndrome treatment | miR-155 and miR-125b in tonsillar β cells ↑ Plasma miR-125b ↑ | [45] |
| Duchenne muscular dystrophy | miR-206 miR-1 miR-133 | Monitoring of prognosis and alternative non-invasive biomarker | Serum miR-1 and miR-206 ↑ | [46] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deveci, G.; Capasso, R.; Ağagündüz, D. Xeno-miRs and Circulating miRNAs as Novel Biomarkers in Certain Diseases. Biologics 2023, 3, 1-10. https://doi.org/10.3390/biologics3010001
Deveci G, Capasso R, Ağagündüz D. Xeno-miRs and Circulating miRNAs as Novel Biomarkers in Certain Diseases. Biologics. 2023; 3(1):1-10. https://doi.org/10.3390/biologics3010001
Chicago/Turabian StyleDeveci, Gülsüm, Raffaele Capasso, and Duygu Ağagündüz. 2023. "Xeno-miRs and Circulating miRNAs as Novel Biomarkers in Certain Diseases" Biologics 3, no. 1: 1-10. https://doi.org/10.3390/biologics3010001