
Oleaginous Red Yeasts: Concomitant Producers of Triacylglycerides and Carotenoids

{kind=link}
{kind=link}
{kind=link}
Definition
:1. Introduction
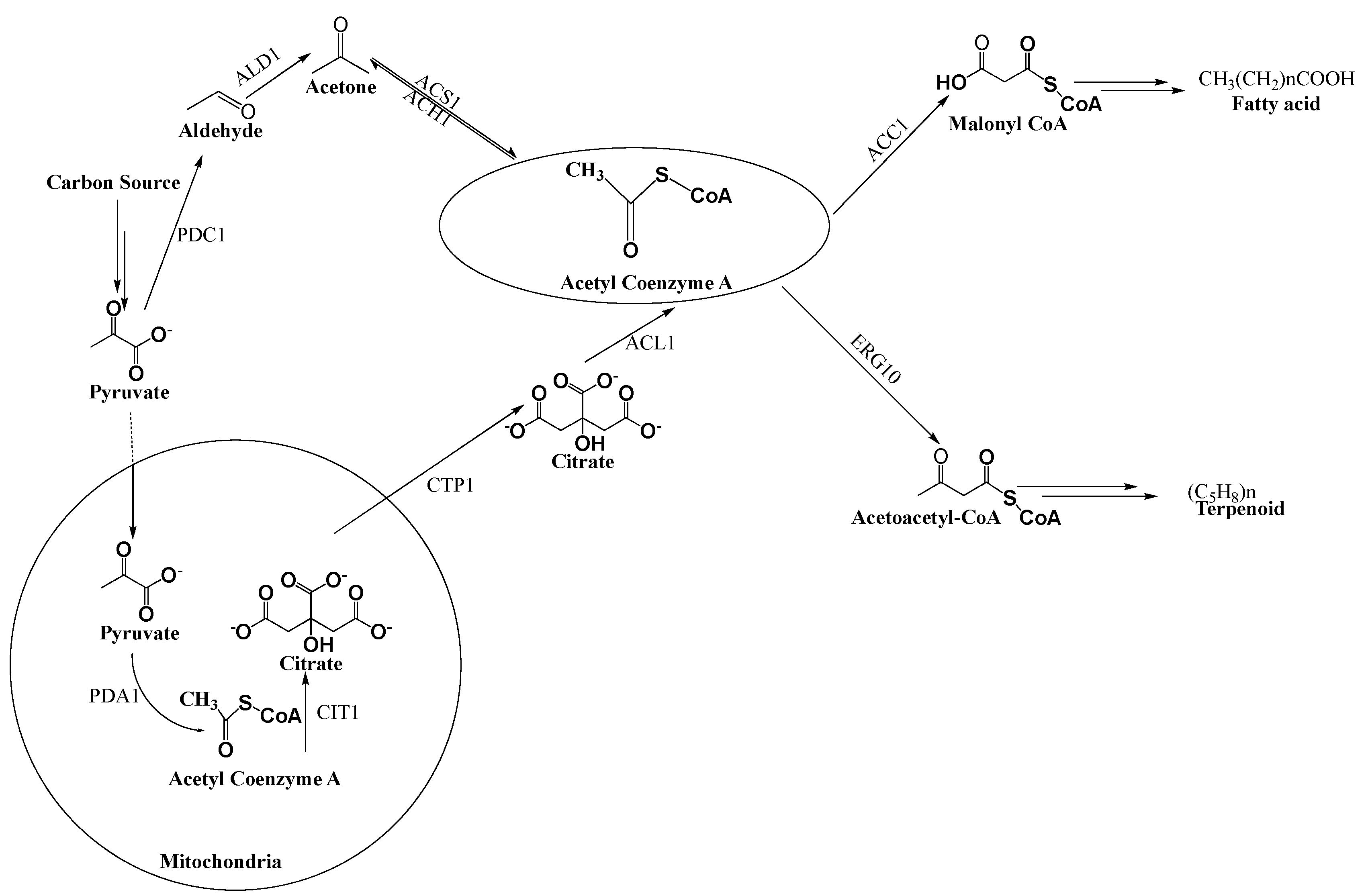
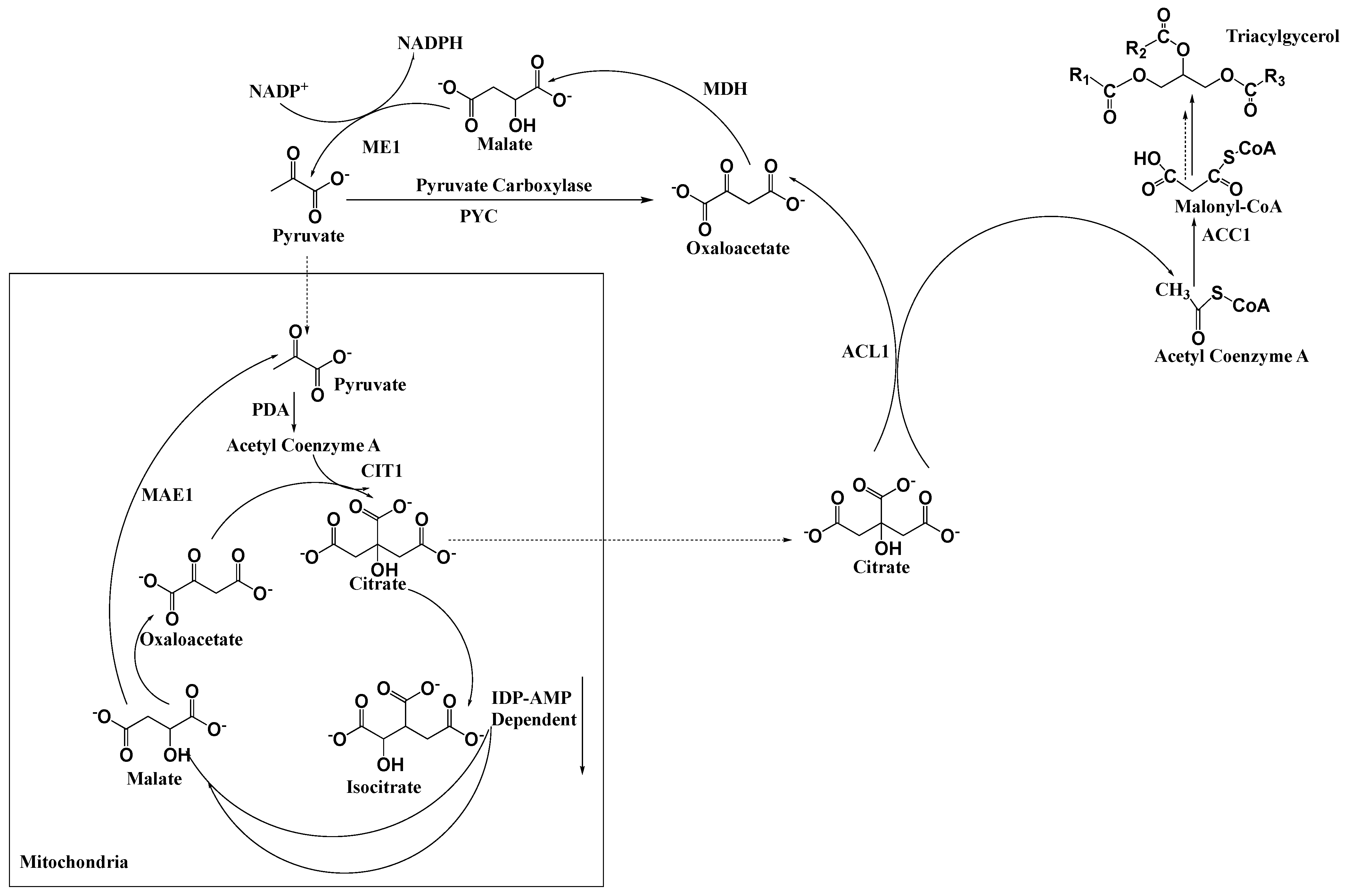
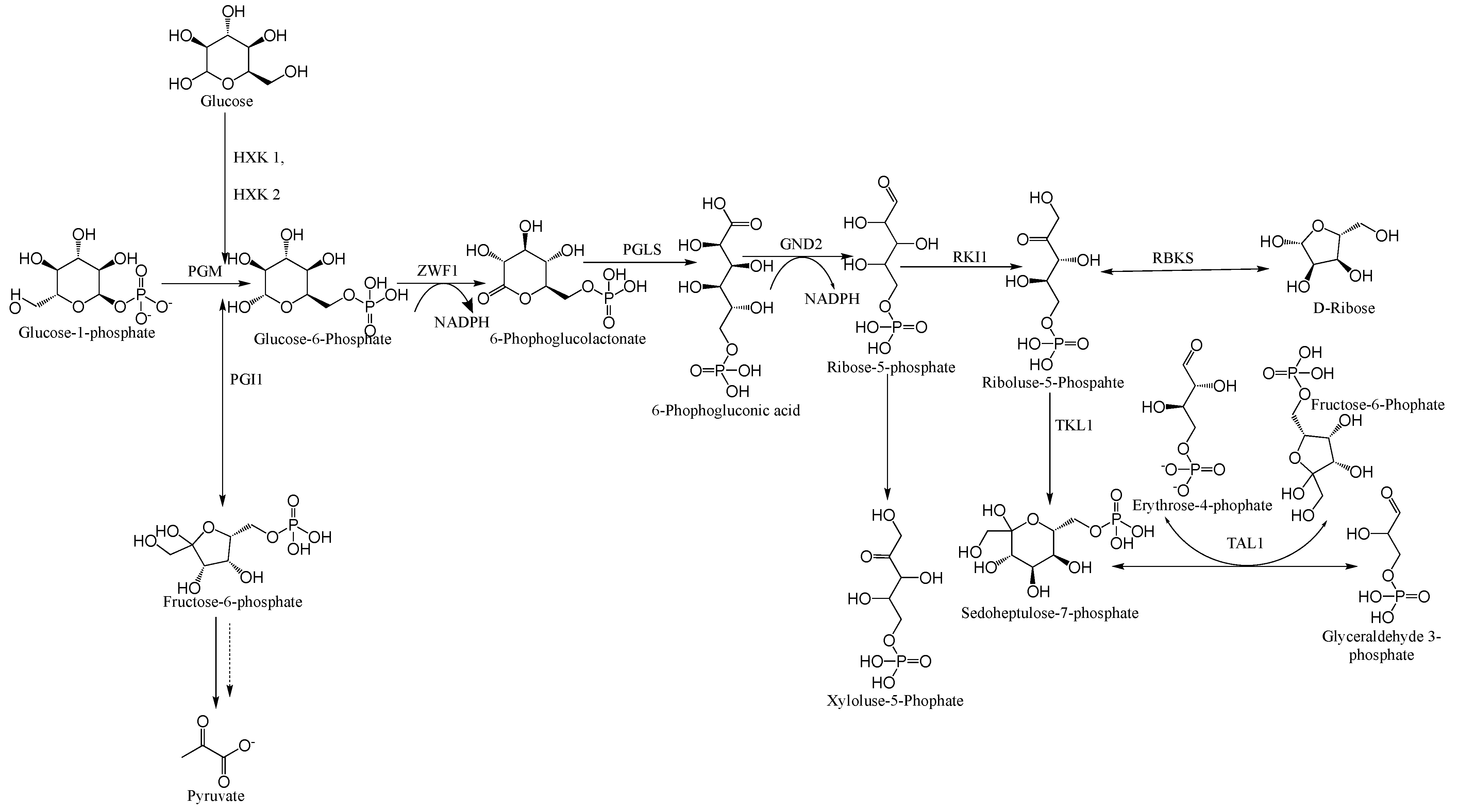
2. Generation of Acetyl-CoA for the Biosynthesis of Fatty Acids and Carotenoids
2.1. Carotenoid Biosynthesis
2.2. De Novo Fatty Acid Biosynthesis
3. Genetic Manipulations and Its Effects on the Fatty Acid and Carotenoid Biosynthesis
3.1. Genomic and Transcriptomic Investigations of Oleaginicity
3.2. Proteomics Approach
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Beopoulos, A.; Nicaud, J.-M.; Gaillardin, C. An Overview of Lipid Metabolism in Yeasts and Its Impact on Biotechnological Processes. Appl. Microbiol. Biotechnol. 2011, 90, 1193–1206. [Google Scholar] [CrossRef]
- Donot, F.; Fontana, A.; Baccou, J.C.; Strub, C.; Schorr-Galindo, S. Single Cell Oils (SCOs) from Oleaginous Yeasts and Moulds: Production and Genetics. Biomass Bioenergy 2014, 68, 135–150. [Google Scholar] [CrossRef]
- Cohen, Z.; Ratledge, C. Single Cell Oils; Elsevier: Amsterdam, The Netherlands, 2015; pp. 6–25. [Google Scholar]
- Buzzini, P.; Innocenti, M.; Turchetti, B.; Libkind, D.; van Broock, M.; Mulinacci, N. Carotenoid Profiles of Yeasts Belonging to the Genera Rhodotorula, Rhodosporidium, Sporobolomyces, and Sporidiobolus. Can. J. Microbiol. 2007, 53, 1024–1031. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhao, Z.; Bai, F. High-Density Cultivation of Oleaginous Yeast Rhodosporidium toruloides Y4 in Fed-Batch Culture. Enzym. Microb. Technol. 2007, 41, 312–317. [Google Scholar] [CrossRef]
- Papanikolaou, S. Oleaginous Yeasts: Biochemical Events Related with Lipid Synthesis and Potential Biotechnological Applications. Ferment. Technol. 2012, 1, e103. [Google Scholar] [CrossRef]
- Munch, G.; Sestric, R.; Sparling, R.; Levin, D.B.; Cicek, N. Lipid Production in the Under-Characterized Oleaginous Yeasts, Rhodosporidium Babjevae and Rhodosporidium Diobovatum, from Biodiesel-Derived Waste Glycerol. Bioresour. Technol. 2015, 185, 49–55. [Google Scholar] [CrossRef]
- Fakankun, I.; Mirzaie, M.; Levin, D.B. Impact of Culture Conditions on Neutral Lipid Production by Oleaginous Yeast. In Microbial Lipid Production; Springer: New York, NY, USA, 2019; pp. 311–325. [Google Scholar]
- Zhu, C.; Chen, B.; Fang, B. Pretreatment of Raw Glycerol with Activated Carbon for 1,3-Propanediol Production by Clostridium Butyricum. Eng. Life Sci. 2013, 13, 376–384. [Google Scholar] [CrossRef]
- Nielsen, J. Synthetic Biology for Engineering Acetyl Coenzyme a Metabolism in Yeast. mBio 2014, 5, e02153-14. [Google Scholar] [CrossRef]
- van Rossum, H.M.; Kozak, B.U.; Pronk, J.T.; van Maris, A.J.A. Engineering Cytosolic Acetyl-Coenzyme a Supply in Saccharomyces Cerevisiae: Pathway Stoichiometry, Free-Energy Conservation and Redox-Cofactor Balancing. Metab. Eng. 2016, 36, 99–115. [Google Scholar] [CrossRef]
- Ratledge, C. Regulation of Lipid Accumulation in Oleaginous Micro-Organisms. Biochem. Soc. 2002, 30, 1047–1050. [Google Scholar] [CrossRef]
- Rodriguez, S.; Denby, C.M.; Van Vu, T.; Baidoo, E.E.K.; Wang, G.; Keasling, J.D. ATP Citrate Lyase Mediated Cytosolic Acetyl-CoA Biosynthesis Increases Mevalonate Production in Saccharomyces Cerevisiae. Microb. Cell Factories 2016, 15, 48. [Google Scholar] [CrossRef]
- Nasirian, N.; Mirzaie, M.; Cicek, N.; Levin, D.B. Lipid and Carotenoid Synthesis by Rhodosporidium Diobovatum, Grown on Glucose versus Glycerol, and Its Biodiesel Properties. Can. J. Microbiol. 2018, 64, 277–289. [Google Scholar] [CrossRef] [PubMed]
- Enshaeieh, M.; Abdoli, A.; Nahvi, I.; Madani, M. Bioconversion of Different Carbon Sources into Microbial Oil and Biodiesel Using Oleaginous Yeasts. J. Biol. Today’s World 2012, 1, 82–92. [Google Scholar] [CrossRef]
- Xu, J.; Liu, D. Exploitation of Genus Rhodosporidium for Microbial Lipid Production. World J. Microbiol. Biotechnol. 2017, 33, 54. [Google Scholar] [CrossRef]
- Park, Y.-K.; Nicaud, J.-M.; Ledesma-Amaro, R. The Engineering Potential of Rhodosporidium Toruloides as a Workhorse for Biotechnological Applications. Trends Biotechnol. 2018, 36, 304–317. [Google Scholar] [CrossRef] [PubMed]
- Vorapreeda, T.; Thammarongtham, C.; Cheevadhanarak, S.; Laoteng, K. Alternative Routes of Acetyl-CoA Synthesis Identified by Comparative Genomic Analysis: Involvement in the Lipid Production of Oleaginous Yeast and Fungi. Microbiology 2012, 158, 217–228. [Google Scholar] [CrossRef]
- Goodwin, T.W. Biosynthesis of Carotenoids: An Overview. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1993; pp. 330–340. [Google Scholar]
- Sandmann, G. Carotenoid Biosynthesis and Biotechnological Application. Arch. Biochem. Biophys. 2001, 385, 4–12. [Google Scholar] [CrossRef]
- Hayman, E.P.; Yokoyama, H.; Chichester, C.O.; Simpson, K.L. Carotenoid Biosynthesis in Rhodotorula Glutinis. J. Bacteriol. 1974, 120, 1339–1343. [Google Scholar] [CrossRef]
- Frengova, G.I.; Beshkova, D.M. Carotenoids from Rhodotorula and Phaffia: Yeasts of Biotechnological Importance. J. Ind. Microbiol. Biotechnol. 2008, 36, 163–180. [Google Scholar] [CrossRef]
- Andrewes, A.G.; Phaff, H.J.; Starr, M.P. Carotenoids of Phaffia Rhodozyma, a Red-Pigmented Fermenting Yeast. Phytochemistry 1976, 15, 1003–1007. [Google Scholar] [CrossRef]
- Aksu, Z.; Eren, A.T. Carotenoids Production by the Yeast Rhodotorula Mucilaginosa: Use of Agricultural Wastes as a Carbon Source. Process Biochem. 2005, 40, 2985–2991. [Google Scholar] [CrossRef]
- Aksu, Z.; Eren, A.T. Production of Carotenoids by the Isolated Yeast of Rhodotorula Glutinis. Biochem. Eng. J. 2007, 35, 107–113. [Google Scholar] [CrossRef]
- Saenge, C.; Cheirsilp, B.; Suksaroge, T.T.; Bourtoom, T. Potential Use of Oleaginous Red Yeast Rhodotorula Glutinis for the Bioconversion of Crude Glycerol from Biodiesel Plant to Lipids and Carotenoids. Process Biochem. 2011, 46, 210–218. [Google Scholar] [CrossRef]
- Tang, W.; Wang, Y.; Zhang, J.; Cai, Y.; He, Z. Biosynthetic Pathway of Carotenoids in Rhodotorula and Strategies for Enhanced Their Production. J. Microbiol. Biotechnol. 2019, 29, 507–517. [Google Scholar] [CrossRef]
- Pinheiro, M.J.; Bonturi, N.; Belouah, I.; Miranda, E.A.; Lahtvee, P.-J. Xylose Metabolism and the Effect of Oxidative Stress on Lipid and Carotenoid Production in Rhodotorula Toruloides: Insights for Future Biorefinery. Front. Bioeng. Biotechnol. 2020, 8, 1008. [Google Scholar] [CrossRef] [PubMed]
- Newell, S.Y.; Hunter, I.L. Rhodosporidium Diobovatum Sp. N., the Perfect Form of an Asporogenous Yeast (Rhodotorula Sp.)1. J. Bacteriol. 1970, 104, 503–508. [Google Scholar] [CrossRef]
- Lyman, M.; Urbin, S.; Strout, C.; Rubinfeld, B. The Oleaginous Red Yeast Rodotorula/Rhodosporidium: A Factory for Industrial Bioproducts. In Yeast in Biotechnology; Basso, T.P., Ed.; Intechopen: London, UK, 2019. [Google Scholar]
- Schneider, T.; Graeff-Hönninger, S.; French, W.T.; Hernandez, R.; Merkt, N.; Claupein, W.; Hetrick, M.; Pham, P. Lipid and Carotenoid Production by Oleaginous Red Yeast Rhodotorula Glutinis Cultivated on Brewery Effluents. Energy 2013, 61, 34–43. [Google Scholar] [CrossRef]
- Zhang, J.; Li, Q.; Liu, J.; Lu, Y.; Wang, Y.; Wang, Y. Astaxanthin Overproduction and Proteomic Analysis of Phaffia Rhodozyma under the Oxidative Stress Induced by TiO2. Bioresour. Technol. 2020, 311, 123525. [Google Scholar] [CrossRef]
- Han, M.; Xu, Z.; Du, C.; Qian, H.; Zhang, W.-G. Effects of Nitrogen on the Lipid and Carotenoid Accumulation of Oleaginous Yeast Sporidiobolus Pararoseus. Bioprocess Biosyst. Eng. 2016, 39, 1425–1433. [Google Scholar] [CrossRef]
- Ghilardi, C.; Sanmartin Negrete, P.; Carelli, A.A.; Borroni, V. Evaluation of olive mill waste as substrate for carotenoid production by Rhodotorula mucilaginosa. Bioresour. Bioprocess. 2020, 7, 1–11. [Google Scholar] [CrossRef]
- Tang, W.; Wang, Y.; Cai, Y.; Liu, S.; Zhang, J.; He, Z. Genome Sequence of a Marine Carotenoid Producing Yeast Rhodotorula Mucilaginosa CYJ03. J. Ocean. Univ. China 2020, 19, 466–472. [Google Scholar] [CrossRef]
- Gan, H.M.; Thomas, B.N.; Cavanaugh, N.T.; Morales, G.H.; Mayers, A.N.; Savka, M.A.; Hudson, A.O. Whole Genome Sequencing of Rhodotorula Mucilaginosa Isolated from the Chewing Stick (Distemonanthus Benthamianus): Insights into Rhodotorula Phylogeny, Mitogenome Dynamics and Carotenoid Biosynthesis. PeerJ 2017, 5, e4030. [Google Scholar] [CrossRef]
- Verdoes, J.C.; Sandmann, G.; Visser, H.; Diaz, M.; van Mossel, M.; van Ooyen, A.J.J. Metabolic Engineering of the Carotenoid Biosynthetic Pathway in the Yeast Xanthophyllomyces Dendrorhous (Phaffia Rhodozyma). Appl. Environ. Microbiol. 2003, 69, 3728–3738. [Google Scholar] [CrossRef] [PubMed]
- Pi, H.-W.; Anandharaj, M.; Kao, Y.-Y.; Lin, Y.-J.; Chang, J.-J.; Li, W.-H. Engineering the Oleaginous Red Yeast Rhodotorula Glutinis for Simultaneous β-Carotene and Cellulase Production. Sci. Rep. 2018, 8, 10850. [Google Scholar] [CrossRef]
- Gong, G.; Zhang, X.; Tan, T. Simultaneously Enhanced Intracellular Lipogenesis and β-Carotene Biosynthesis of Rhodotorula Glutinis by Light Exposure with Sodium Acetate as the Substrate. Bioresour. Technol. 2020, 295, 122274. [Google Scholar] [CrossRef]
- Pham, K.D.; Shida, Y.; Miyata, A.; Takamizawa, T.; Suzuki, Y.; Ara, S.; Yamazaki, H.; Masaki, K.; Mori, K.; Aburatani, S.; et al. Effect of Light on Carotenoid and Lipid Production in the Oleaginous Yeast Rhodosporidium Toruloides. Biosci. Biotechnol. Biochem. 2020, 84, 1501–1512. [Google Scholar] [CrossRef] [PubMed]
- Evans, C.T.; Ratledge, C. Effect of Nitrogen Source on Lipid Accumulation in Oleaginous Yeasts. Microbiology 1984, 130, 1693–1704. [Google Scholar] [CrossRef]
- Lopes, H.J.S.; Bonturi, N.; Kerkhoven, E.J.; Miranda, E.A.; Lahtvee, P.-J. C/N Ratio and Carbon Source-Dependent Lipid Production Profiling in Rhodotorula Toruloides. Appl. Microbiol. Biotechnol. 2020, 104, 2639–2649. [Google Scholar] [CrossRef]
- Papanikolaou, S.; Aggelis, G. Lipids of Oleaginous Yeasts. Part I: Biochemistry of Single Cell Oil Production. Eur. J. Lipid Sci. Technol. 2011, 113, 1031–1051. [Google Scholar] [CrossRef]
- Sitepu, I.R.; Sestric, R.; Ignatia, L.; Levin, D.; German, J.B.; Gillies, L.A.; Almada, L.A.G.; Boundy-Mills, K.L. Manipulation of Culture Conditions Alters Lipid Content and Fatty Acid Profiles of a Wide Variety of Known and New Oleaginous Yeast Species. Bioresour. Technol. 2013, 144, 360–369. [Google Scholar] [CrossRef]
- Granger, L.-M.; Perlot, P.; Goma, G.; Pareilleux, A. Effect of Various Nutrient Limitations on Fatty Acid Production by Rhodotorula Glutinis. Appl. Microbiol. Biotechnol. 1993, 38, 784–789. [Google Scholar] [CrossRef]
- Ratledge, C. Microbial Lipids: Commercial Realities or Academic Curiosities. In Industrial Applications of Single Cell Oil; Kyle, D.J., Ed.; American Oil Chemists’ Society: Champaign, IL, USA, 1992; pp. 1–15. [Google Scholar]
- Fakankun, I.; Fristensky, B.; Levin, D.B. Genome Sequence Analysis of the Oleaginous Yeast, Rhodotorula Diobovata, and Comparison of the Carotenogenic and Oleaginous Pathway Genes and Gene Products with Other Oleaginous Yeasts. J. Fungi 2021, 7, 320. [Google Scholar] [CrossRef]
- Shen, Q.; Chen, Y.; Jin, D.; Lin, H.; Wang, Q.; Zhao, Y.-H. Comparative Genome Analysis of the Oleaginous Yeast Trichosporon Fermentans Reveals Its Potential Applications in Lipid Accumulation. Microbiol. Res. 2016, 192, 203–210. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Zhang, S.; Liu, H.; Shen, H.; Lin, X.; Yang, F.; Zhou, Y.J.; Jin, G.; Ye, M.; Zou, H.; et al. A Multi-Omic Map of the Lipid-Producing Yeast Rhodosporidium Toruloides. Nat. Commun. 2012, 3, 1112. [Google Scholar] [CrossRef] [PubMed]
- Fabiszewska, A.; Misiukiewicz-Stępień, P.; Paplińska-Goryca, M.; Zieniuk, B.; Białecka-Florjańczyk, E. An Insight into Storage Lipid Synthesis by Yarrowia Lipolytica Yeast Relating to Lipid and Sugar Substrates Metabolism. Biomolecules 2019, 9, 685. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhang, L.; Chen, H.; Chen, Y.Q.; Ratledge, C.; Song, Y.; Chen, W. Regulatory Properties of Malic Enzyme in the Oleaginous Yeast, Yarrowia Lipolytica, and Its Non-Involvement in Lipid Accumulation. Biotechnol. Lett. 2013, 35, 2091–2098. [Google Scholar] [CrossRef]
- Ratledge, C. The Role of Malic Enzyme as the Provider of NADPH in Oleaginous Microorganisms: A Reappraisal and Unsolved Problems. Biotechnol. Lett. 2014, 36, 1557–1568. [Google Scholar] [CrossRef]
- Chen, H.; Hao, G.; Wang, L.; Wang, H.; Gu, Z.; Liu, L.; Zhang, H.; Chen, W.; Chen, Y.Q. Identification of a Critical Determinant That Enables Efficient Fatty Acid Synthesis in Oleaginous Fungi. Sci. Rep. 2015, 5, 11247. [Google Scholar] [CrossRef]
- Dulermo, T.; Lazar, Z.; Dulermo, R.; Rakicka, M.; Haddouche, R.; Nicaud, J.-M. Analysis of ATP-Citrate Lyase and Malic Enzyme Mutants of Yarrowia Lipolytica Points out the Importance of Mannitol Metabolism in Fatty Acid Synthesis. Biochim. Biophys. Acta (BBA) Mol. Cell Biol. Lipids 2015, 1851, 1107–1117. [Google Scholar] [CrossRef]
- Zhang, S.; Skerker, J.M.; Rutter, C.D.; Maurer, M.J.; Arkin, A.P.; Rao, C.V. Engineering Rhodosporidium Toruloides for Increased Lipid Production. Biotechnol. Bioeng. 2015, 113, 1056–1066. [Google Scholar] [CrossRef]
- Sun, T.; Hayakawa, K.; Bateman, K.S.; Fraser, M.E. Identification of the Citrate-Binding Site of Human ATP-Citrate Lyase Using X-Ray Crystallography. J. Biol. Chem. 2010, 285, 27418–27428. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Ito, M.; Skerker, J.M.; Arkin, A.P.; Rao, C.V. Metabolic Engineering of the Oleaginous Yeast Rhodosporidium Toruloides IFO0880 for Lipid Overproduction during High-Density Fermentation. Appl. Microbiol. Biotechnol. 2016, 100, 9393–9405. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, S.; Zhu, Z.; Shen, H.; Lin, X.; Jin, X.; Jiao, X.; Zhao, Z.K. Systems Analysis of Phosphate-Limitation-Induced Lipid Accumulation by the Oleaginous Yeast Rhodosporidium Toruloides. Biotechnol. Biofuels 2018, 11, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Koh CM, J.; Yap, S.A.; Du, M.; Hlaing, M.M.; Ji, L. Identification of Novel Genes in the Carotenogenic and Oleaginous Yeast Rhodotorula Toruloides through Genome-Wide Insertional Mutagenesis. BMC Microbiol. 2018, 18, 14. [Google Scholar] [CrossRef]
- Jiao, X.; Lyu, L.; Zhang, Y.; Huang, Q.; Zhou, R.; Wang, S.; Wang, S.; Zhang, S.; Zhao, Z.K. Reduction of Lipid-Accumulation of Oleaginous Yeast Rhodosporidium Toruloides through CRISPR/Cas9-Mediated Inactivation of Lipid Droplet Structural Proteins. FEMS Microbiol. Lett. 2021, 368, fnab111. [Google Scholar] [CrossRef]
- Tran, T.N.; Tran, Q.-V.; Huynh, H.T.; Hoang, N.-S.; Nguyen, H.C.; Ngo, D.-N. Astaxanthin Production by Newly Isolated Rhodosporidium Toruloides: Optimization of Medium Compositions by Response Surface Methodology. Not. Bot. Horti Agrobot. Cluj-Napoca 2018, 47, 320–327. [Google Scholar] [CrossRef]
- Tran, T.N.; Ngo, D.-H.; Tran, Q.T.; Nguyen, H.C.; Su, C.-H.; Ngo, D.-N. Enhancing Astaxanthin Biosynthesis by Rhodosporidium Toruloides Mutants and Optimization of Medium Compositions Using Response Surface Methodology. Processes 2020, 8, 497. [Google Scholar] [CrossRef]
- Chaturvedi, S.; Gupta, A.K.; Bhattacharya, A.; Dutta, T.; Nain, L.; Khare, S.K. Overexpression and Repression of Key Rate-Limiting Enzymes (Acetyl CoA Carboxylase and HMG Reductase) to Enhance Fatty Acid Production from Rhodotorula Mucilaginosa. J. Basic Microbiol. 2020, 61, 4–14. [Google Scholar] [CrossRef]
- Ruenwai, R.; Cheevadhanarak, S.; Laoteng, K. Overexpression of Acetyl-CoA Carboxylase Gene of Mucor Rouxii Enhanced Fatty Acid Content in Hansenula Polymorpha. Mol. Biotechnol. 2009, 42, 327–332. [Google Scholar] [CrossRef]
- Meng, X.; Yang, J.; Cao, Y.; Li, L.; Jiang, X.; Xu, X.; Liu, W.; Xian, M.; Zhang, Y. Increasing Fatty Acid Production in E. Coli by Simulating the Lipid Accumulation of Oleaginous Microorganisms. J. Ind. Microbiol. Biotechnol. 2010, 38, 919–925. [Google Scholar] [CrossRef]
- Davis, M.S.; Solbiati, J.; Cronan, J.E. Overproduction of Acetyl-CoA Carboxylase Activity Increases the Rate of Fatty Acid Biosynthesis in Escherichia Coli. J. Biol. Chem. 2000, 275, 28593–28598. [Google Scholar] [CrossRef] [PubMed]
- Kourist, R.; Bracharz, F.; Lorenzen, J.; Kracht, O.N.; Chovatia, M.; Daum, C.; Deshpande, S.; Lipzen, A.; Nolan, M.; Ohm, R.A.; et al. Genomics and Transcriptomics Analyses of the Oil-Accumulating Basidiomycete Yeast Trichosporon Oleaginosus: Insights into Substrate Utilization and Alternative Evolutionary Trajectories of Fungal Mating Systems. mBio 2015, 6, e00918-15. [Google Scholar] [CrossRef] [PubMed]
- Sestric, R.; Spicer, V.V.; Krokhin, O.; Sparling, R.B.; Levin, D. Analysis of the Yarrowia Lipolytica Proteome Reveals Subtle Variations in Expression Levels between Lipogenic and Non-Lipogenic Conditions. FEMS Yeast Res. 2021, 21, foab007. [Google Scholar] [CrossRef] [PubMed]
- Fakankun, I.; Spicer, V.; Levin, D.B. Proteomic Analyses of the Oleaginous and Carotenogenic Yeast Rhodotorula Diobovata across Growth Phases under Nitrogen- and Oxygen-Limited Conditions. J. Biotechnol. 2021, 332, 11–19. [Google Scholar] [CrossRef]
- Gao, S.; Han, L.; Zhu, L.; Ge, M.; Yang, S.; Jiang, Y.; Chen, D. One-Step Integration of Multiple Genes into the Oleaginous Yeast Yarrowia Lipolytica. Biotechnol. Lett. 2014, 36, 2523–2528. [Google Scholar] [CrossRef]
- Lin, X.; Wang, Y.; Zhang, S.; Zhu, Z.; Zhou, Y.J.; Yang, F.; Sun, W.; Wang, X.; Zhao, Z.K. Functional Integration of Multiple Genes into the Genome of the Oleaginous Yeast Rhodosporidium Toruloides. FEMS Yeast Res. 2014, 14, 547–555. [Google Scholar] [CrossRef]
- Morin, N.; Cescut, J.; Beopoulos, A.; Lelandais, G.; Le Berre, V.; Uribelarrea, J.-L.; Molina-Jouve, C.; Nicaud, J.-M. Transcriptomic Analyses during the Transition from Biomass Production to Lipid Accumulation in the Oleaginous Yeast Yarrowia Lipolytica. PLoS ONE 2011, 6, e27966. [Google Scholar] [CrossRef]
- Qian, X.; Ba, Y.; Zhuang, Q.; Zhong, G. RNA-Seq Technology and Its Application in Fish Transcriptomics. OMICS J. Integr. Biol. 2014, 18, 98–110. [Google Scholar] [CrossRef]
- Martin, D.B.; Nelson, P.S. From Genomics to Proteomics: Techniques and Applications in Cancer Research. Trends Cell Biol. 2001, 11, S60–S65. [Google Scholar] [CrossRef]
- Dove, A. Proteomics: Translating Genomics into Products? Nat. Biotechnol. 1999, 17, 233–236. [Google Scholar] [CrossRef]
- Aebersold, R.; Mann, M. Mass Spectrometry-Based Proteomics. Nature 2003, 422, 198–207. [Google Scholar] [CrossRef] [PubMed]
- Resing, K.A.; Ahn, N.G. Proteomics Strategies for Protein Identification. FEBS Lett. 2004, 579, 885–889. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Feng, H.; Lee, J.; Ning Chen, W. Comparative Proteomics Profile of Lipid-Cumulating Oleaginous Yeast: An ITRAQ-Coupled 2-D LC-MS/MS Analysis. PLoS ONE 2013, 8, e85532. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Zhao, X.; Wang, F.; Li, Y.; Jiang, X.; Ye, M.; Zhao, Z.K.; Zou, H. Comparative Proteomic Analysis of Rhodosporidium Toruloides during Lipid Accumulation. Yeast 2009, 26, 553–566. [Google Scholar] [CrossRef]
- Liu, H.; Zhao, X.; Wang, F.; Jiang, X.; Zhang, S.; Ye, M.; Zhao, Z.K.; Zou, H. The Proteome Analysis of Oleaginous Yeast Lipomyces Starkeyi. FEMS Yeast Res. 2010, 11, 42–51. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fakankun, I.; Levin, D.B. Oleaginous Red Yeasts: Concomitant Producers of Triacylglycerides and Carotenoids. Encyclopedia 2023, 3, 490-500. https://doi.org/10.3390/encyclopedia3020034
Fakankun I, Levin DB. Oleaginous Red Yeasts: Concomitant Producers of Triacylglycerides and Carotenoids. Encyclopedia. 2023; 3(2):490-500. https://doi.org/10.3390/encyclopedia3020034
Chicago/Turabian StyleFakankun, Irene, and David B. Levin. 2023. "Oleaginous Red Yeasts: Concomitant Producers of Triacylglycerides and Carotenoids" Encyclopedia 3, no. 2: 490-500. https://doi.org/10.3390/encyclopedia3020034