
Optimization of Probiotic Lactobacilli Production for In-Feed Supplementation to Feedlot Cattle

Abstract
:1. Introduction
2. Material and Methods
2.1. Probiotic Bacteria Used in This Study
2.2. Optimization of Culture Media
2.3. Carbon Sources
2.4. Nitrogen Sources
2.5. Optimization of Growth Conditions for Biomass Production
2.5.1. Evaluation of Temperature and pH in MRS Medium
2.5.2. Biomass Production in a Laboratory Bioreactor
2.6. Preservation of the Probiotic Biomass
2.7. Maintenance of the Probiotic Properties of the Lactobacilli
2.8. Statistics
3. Results
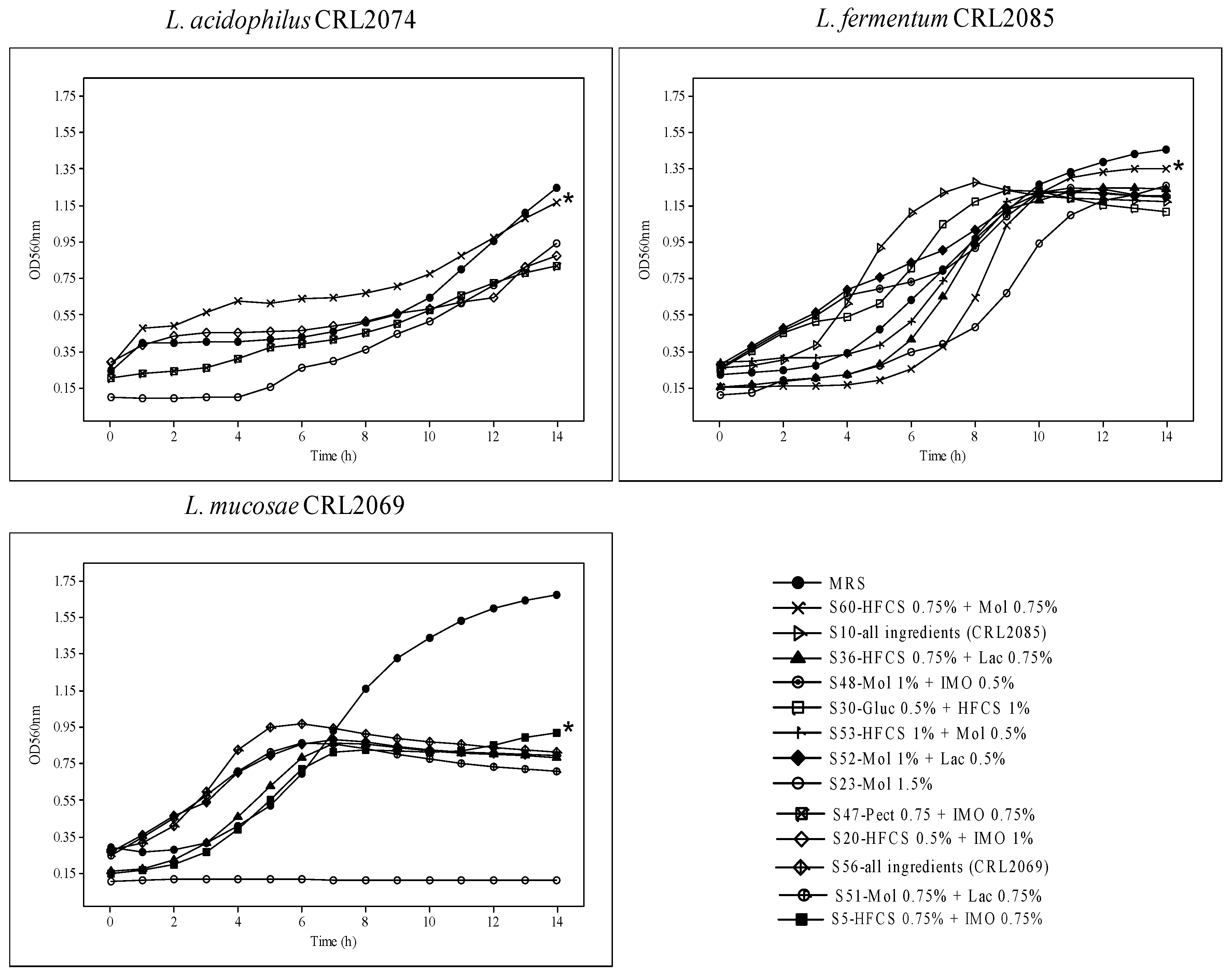
3.1. Selection of Carbon and Nitrogen Sources
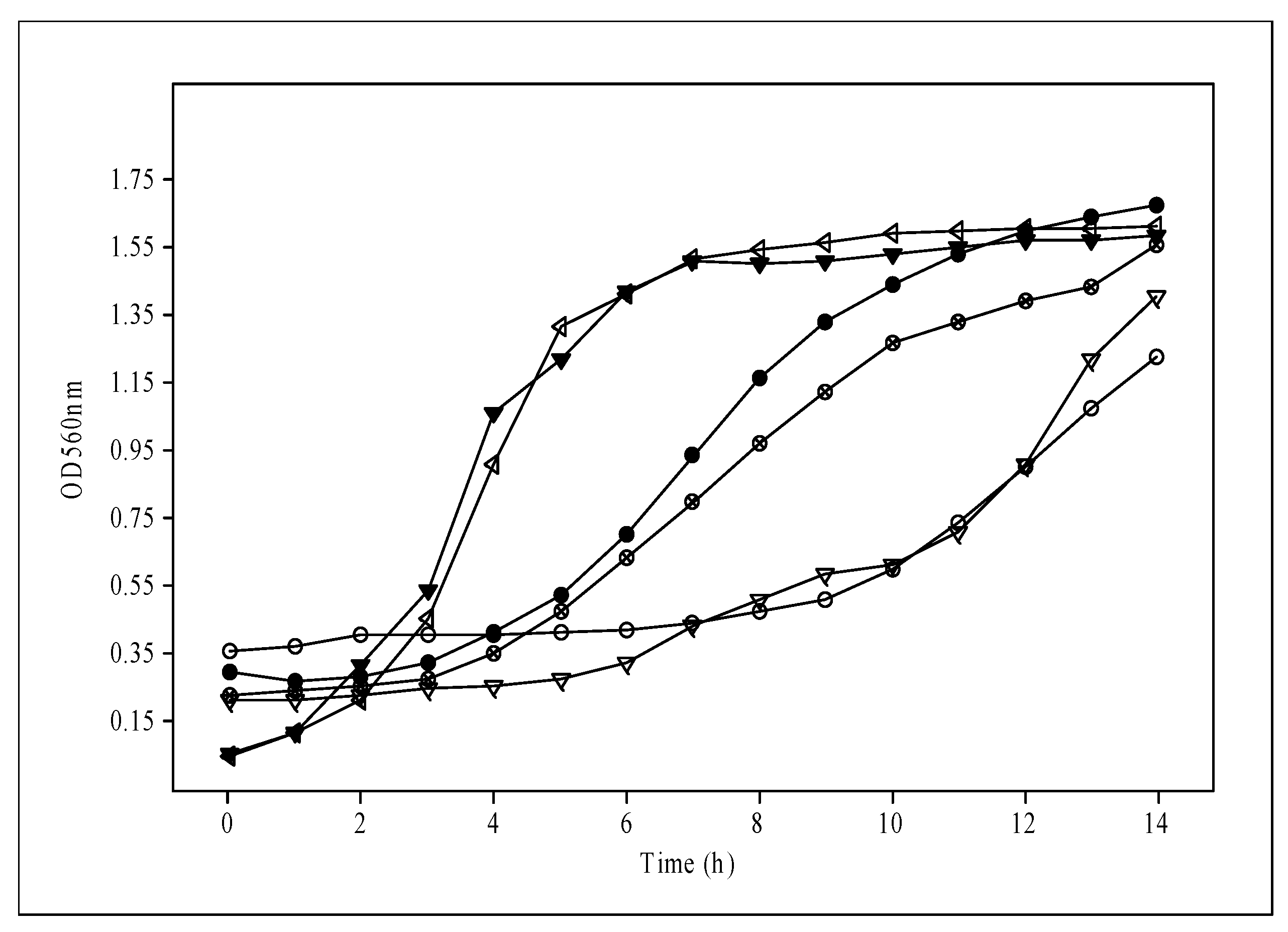
3.2. pH and Incubation Temperature Impact on Biomass Production for Probiotic Lactobacilli
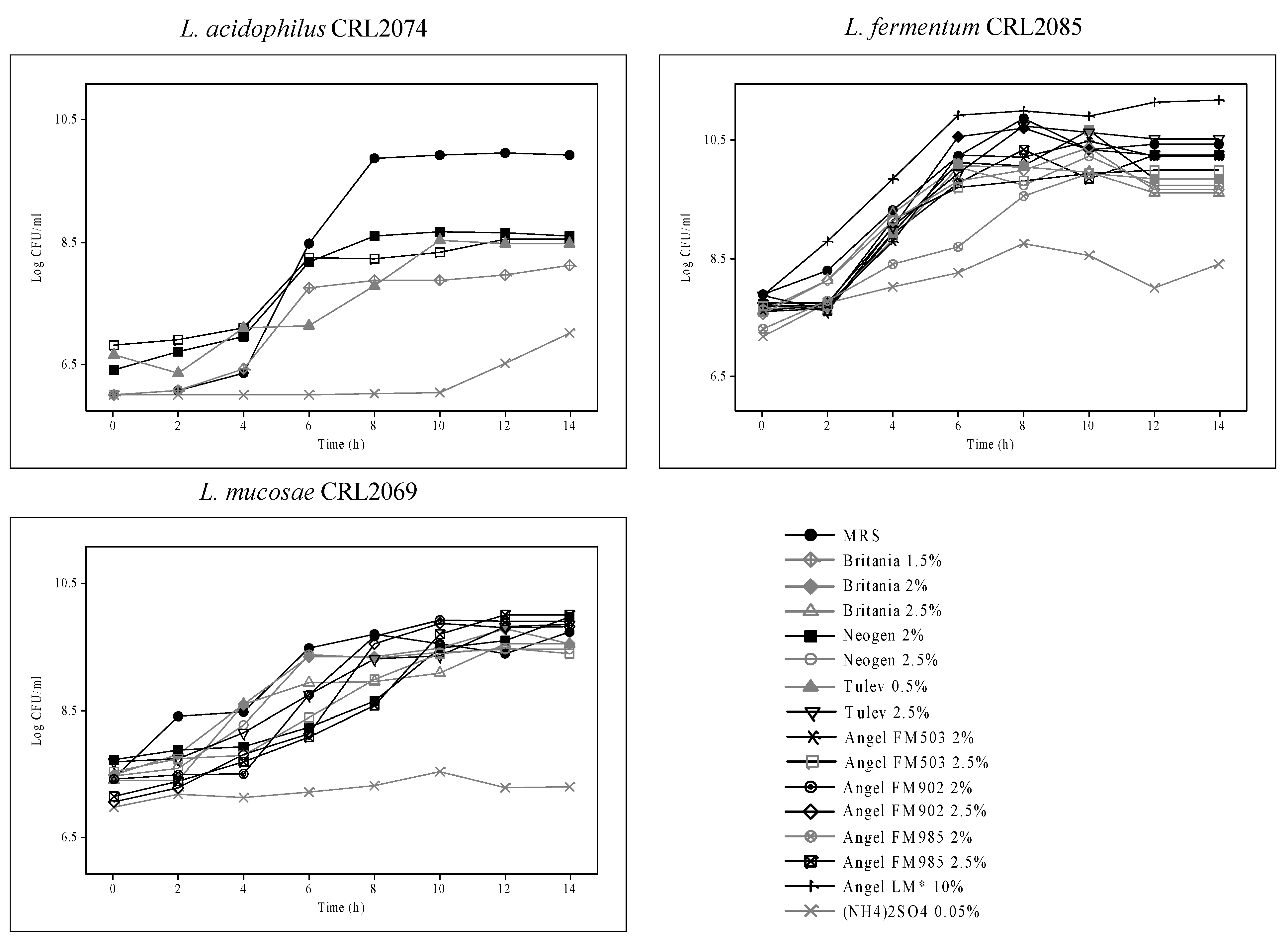
3.3. Biomass Preservation
3.4. Maintenance of Probiotic Properties after Freeze Drying
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Adjei-Fremah, S.; Ekwemalor, K.; Worku, M.; Ibrahim, S. Probiotics and Ruminant Health. In Probiotics. Current Knowledge and Future Prospects; Enany, S., Ed.; IntechOpen: London, UK, 2018; Chapter 8; pp. 133–150. [Google Scholar] [CrossRef] [Green Version]
- Alayande, K.A.; Aiyegoro, O.A.; Ateba, C.N. Probiotics in animal husbandry: Applicability and associated risk factors. Sustainability 2020, 12, 1087. [Google Scholar] [CrossRef] [Green Version]
- Zamojska, D.; Nowak, A.; Nowak, I.; Macierzyňska-Piotrowska, E. Probiotics and Postbiotics as substitutes of antibiotics in farm animals: A Review. Animals 2021, 11, 3431. [Google Scholar] [CrossRef] [PubMed]
- Salminen, S.; Collado, M.C.; Endo, A.; Hill, C.; Lebeer, S.; Quigley, E.M.M.; Sanders, M.E.; Shamir, R.; Swann, J.R.; Szajewska, H.; et al. The International Scientific Association of Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of postbiotics. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 649–667. [Google Scholar] [CrossRef] [PubMed]
- Anee, I.J.; Alam, S.; Begum, R.A.; Md Shahjahan, R.; Khandaker, A.M. The role of probiotics on animal health and nutrition. J. Basic Appl. Zool. 2021, 82, 52. [Google Scholar] [CrossRef]
- Kober, A.K.M.H.; Riaz Rajoka, M.S.; Mehwish, H.M.; Villena, J.; Kitazawa, H. Immunomodulation potential of probiotics: A novel strategy for improving livestock health, immunity, and productivity. Microorganisms 2022, 10, 388. [Google Scholar] [CrossRef] [PubMed]
- Bajagay, Y.S.; Klieve, A.V.; Dart, P.J.; Bryden, W.L. Probiotics in Animal Nutrition–Production, Impact and Regulation. In FAO Animal Production and Health Paper; FAO: Rome, Italy, 2016. [Google Scholar]
- Cangiano, L.R.; Yohe, T.T.; Steele, M.A.; Renaud, D.L. Strategic use of microbial-based probiotics and prebiotics in dairy calf rearing. Appl. Anim. Sci. 2020, 36, 630–651. [Google Scholar] [CrossRef]
- Mansilla, F.I.; Aristimuño Ficoseco, C.; Miranda, M.H.; Puglisi, E.; Nader-Macías, M.E.F.; Vignolo, G.M.; Fontana, C.A. Administration of probiotic lactic acid bacteria to modulate fecal microbiome in feedlot cattle, Sci. Rep. 2022, 12, 12957. [Google Scholar] [CrossRef]
- De Mann, J.C.; Rogosa, M.; Sharpe, E.M. A medium for the cultivation of lactobacilli. J. Appl. Bacteriol. 1960, 23, 130–135. [Google Scholar] [CrossRef]
- Chiang, M.L.; Chen, H.C.; Chen, K.N.; Lin, Y.C.; Lin, Y.T.; Chen, M.J. Optimizing production of two potential probiotic lactobacilli strains isolated from piglet feces as feed additives for weaned piglets. Asian-Australas. J. Anim. Sci. 2015, 28, 1163–1170. [Google Scholar] [CrossRef] [Green Version]
- Hwang, C.F.; Chang, J.H.; Houng, J.Y.; Tsai, C.C.; Lin, C.K.; Tsen, H.Y. Optimization of medium composition for improving biomass production of Lactobacillus plantarum Pi06 using the Taguchi Array design and the Box-Behnken method. Biotechnol. Bioprocess Eng. 2012, 17, 827–834. [Google Scholar] [CrossRef]
- Manzoor, A.; Qazi, J.I.; Haq, I.; Mukhtar, H.; Rasool, A. Significantly enhanced biomass production of a novel bio-therapeutic strain Lactobacillus plantarum (AS-14) by developing low cost media cultivation strategy. J. Biol. Eng. 2017, 11, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chatterjee, E.; Manuel, S.G.A.; Hassan, S.H. Effect of fruit pectin on growth of lactic acid bacteria. J. Prob. Health 2016, 4, 2. [Google Scholar] [CrossRef]
- Coghetto, C.C.; Bettker Vasconcelos, C.; Brusch Brinques, G.; Záchia Ayubet, M.A. Lactobacillus plantarum BL011 cultivation in industrial isolated soybean protein acid residue. Braz. J. Microbiol. 2016, 47, 941–948. [Google Scholar] [CrossRef] [Green Version]
- Śliżewska, K.; Chlebicz-Wójcik, A. Growth kinetics of probiotic Lactobacillus strains in the alternative, cost-efficient semi-solid fermentation medium. Biology 2020, 9, 423. [Google Scholar] [CrossRef] [PubMed]
- Vinderola, G.; Champagne, C.P.; Desfossès-Foucailt, E. The Production of Lactic Acid Bacteria Starters and Probiotics. An Industrial Perspective. In Lactic Acid Bacteria. Microbioloical and Functional Aspects; Vinderola, G., Ouwehand, A.C., Salminen, S., von Wright, A., Eds.; CRC Press, Taylor & Francis Group: Boca Raton, FL, USA, 2019; Chapter 20; pp. 317–336. [Google Scholar]
- Carvalho, A.S.; Silva, J.; Ho, P.; Teixeira, P.; Malcata, F.X.; Gibbs, P. Relevant factors for the preparation of freeze-dried lactic acid bacteria. Int. Dairy J. 2004, 14, 835–847. [Google Scholar] [CrossRef]
- Mansilla, F.I.; Miranda, M.H.; Uezen, J.D.; Maldonado, N.C.; D’Urso Villar, M.A.; Merino, L.A.; Vignolo, G.M.; Nader-Macias, M.E.F. Effect of probiotic lactobacilli supplementation on growth parameters, blood profile, productive performance, and fecal microbiology in feedlot cattle. Res. Vet. Sci. 2023, 155, 76–87. [Google Scholar] [CrossRef]
- Mansilla, F.I.; Miranda, M.H.; Aristimuño Ficoseco, C.; Obregozo, M.; D’Urso Villar, M.; Nader-Macias, M.E.F.; Vignolo, G.M. Use of probiotic lactobacilli as alternative to monensin in beef feedlot cattle. Res. Vet. Sci. 2023; in press. [Google Scholar]
- Aristimuño Ficoseco, C.; Mansilla, F.I.; Maldonado, N.C.; Miranda, H.; Nader-Macias, M.E.F.; Vignolo, G.M. Safety and growth optimization of lactic acid bacteria isolated from feedlot cattle for probiotic formula design. Front. Microbiol. 2018, 9, 2220. [Google Scholar] [CrossRef]
- Maldonado, N.C.; Aristimuño Ficoseco, C.; Mansilla, F.I.; Melián, C.; Hébert, E.M.; Vignolo, G.M.; Nader-Macías, M.E.F. Identification, characterization and selection of autochthonous lactic acid bacteria as probiotic for feedlot cattle. Livest. Sci. 2018, 212, 99–110. [Google Scholar] [CrossRef]
- Buruk Şahin, Y.; Aktar Demirtaş, E.; Burnak, N. Mixture design: A review of recent applications in the food industry. Pamukkale Univ. J. Eng. Sci. 2016, 22, 297–304. [Google Scholar] [CrossRef]
- Heenan, C.N.; Adams, M.C.; Hosken, R.W.; Fleet, G.H. Growth medium for culturing probiotic bacteria for applications in vegetarian food products. LWT-Food Sci. Technol. 2002, 35, 171–176. [Google Scholar] [CrossRef]
- Palmonari, A.; Cavallini, D.; Sniffen, C.J.; Fernandes, L.; Holder, P.; Fagioli, L.; Fusaro, I.; Biagi, G.; Formigoni, A.; Mammi, L. Characterization of molasses chemical composition. J. Dairy Sci. 2020, 103, 6244–6249. [Google Scholar] [CrossRef]
- Sen, K.Y.; Hazwan Hussin, M.; Baidurah, S. Biosynthesis of poly(3-hydroxybutyrate) (PHB) by Cupriavidus necator from various pretreated molasses as carbon source. Biocatal. Agric. Biotechnol. 2019, 17, 51–59. [Google Scholar] [CrossRef]
- Parker, K.; Salas, M.; Nwosu, V.C. High fructose corn syrup: Production, uses and public health concerns. Biotechnol. Mol. Biol. Rev. 2010, 5, 71–78. [Google Scholar]
- Srinivas, D.; Mital, B.K.; Garg, K. Utilization of sugars by Lactobacillus acidophilus strains. Int. J. Food Microbiol. 1990, 10, 51–57. [Google Scholar] [CrossRef]
- Verce, M.; De Vuyst, L.; Weckx, S. Comparative genomics of Lactobacillus fermentum suggests a free-living lifestyle of this lactic acid bacterial species. Food Microbiol. 2020, 89, 103448. [Google Scholar] [CrossRef]
- Jia, Y.; Yang, B.; Ross, P.; Stanton, C.; Zhang, H.; Zhao, J.; Chenet, W. Comparative genomics analysis of Lactobacillus mucosae from different niches. Genes 2020, 11, 95. [Google Scholar] [CrossRef] [Green Version]
- Krzywonos, M.; Eberhard, T. High density process to cultivate Lactobacillus plantarum biomass using wheat stillage and sugar beet molasses. Electron. J. Biotechnol. 2011, 14, 6. [Google Scholar] [CrossRef] [Green Version]
- Saha, B.C. A low-cost medium for mannitol production by Lactobacillus intermedius NRRL B-3693. Appl. Microbiol. Biotechnol. 2006, 72, 676–680. [Google Scholar] [CrossRef]
- Tari, C.; Genckal, H.; Tokatlı, F. Optimization of a growth medium using a statistical approach for the production of an alkaline protease from a newly isolated Bacillus sp. L21. Process Biochem. 2006, 41, 659–665. [Google Scholar] [CrossRef] [Green Version]
- KiBeom, L.; Kee, K.S.; Jun, J.C. A low-cost Lactobacillus salivarius L29 growth medium containing molasses and corn steep liquor allows the attainment of high levels of cell mass and lactic acid production. Afr. J. Biotechnol. 2013, 12, 2013–2018. [Google Scholar] [CrossRef] [Green Version]
- Goderska, K.; Nowak, J.; Czarnecki, Z. Comparison of the growth of Lactobacillus acidophilus and Bifidobacterium bifidum species in media supplemented with selected saccharides including prebiotics. Acta Sci. Pol. Technol. Aliment. 2008, 7, 5–20. [Google Scholar]
- Gänzle, M.; Follador, R. Metabolism of oligosaccharides and starch in lactobacilli: A Review. Front. Microbiol. 2012, 3, 340. [Google Scholar] [CrossRef] [Green Version]
- Yeo, S.; Shin, H.; Lee, H.W.; Hong, D.; Park, H.; Holzapfel, W.; Kim, E.B.; Huh, C.S. Determination of optimized growth medium and cryoprotective additives to enhance the growth and survival of Lactobacillus salivarius. J. Microbiol. Biotechnol. 2018, 28, 718–731. [Google Scholar] [CrossRef] [PubMed]
- Saarela, M.H.; Alakomi, H.L.; Puhakka, A.; Mättö, J. Effect of the fermentation pH on the storage stability of Lactobacillus rhamnosus preparations and suitability of in vitro analyses of cell physiological functions to predict it. J. Appl. Microbiol. 2009, 106, 1204–1212. [Google Scholar] [CrossRef]
- Sashihara, T.; Sueki, N.; Furuichi, K.; Ikegami, S. Effect of growth conditions of Lactobacillus gasseri OLL2809 on the immunostimulatory activity for production of interleukin-12 (p70) by murine splenocytes. Int. J. Food Microbiol. 2007, 120, 274–281. [Google Scholar] [CrossRef] [PubMed]
- Corcoran, B.M.; Stanton, C.; Fitzgerald, G.F.; Ross, R.P. Survival of probiotic lactobacilli in acidic environments is enhanced in the presence of metabolizable sugars. Appl. Environ. Microbiol. 2005, 71, 3060–3067. [Google Scholar] [CrossRef] [Green Version]
- Choi, G.H.; Lee, N.K.; Paik, H.D. Optimization of medium composition for biomass production of Lactobacillus plantarum 200655 using response surface methodology. J. Microbiol. Biotechnol. 2021, 31, 717–725. [Google Scholar] [CrossRef]
- Selvamani, S.; Dailin, D.J.; Rostom, M.; Malek, R.A.; Gupta, V.K.; El-Enshasy, H.A. Optimizing medium components to enhance high cell mass production of biotherapeutic strain Lactobacillus reuteri DSM 20016T by statistical method. J. Sci. Ind. Res. 2020, 79, 798–803. [Google Scholar]
- Lavari, L.; Ianniello, R.; Paez, R.; Zotta, T.; Cuatrin, A.L.; Reinheimer, J.A.; Parente, E.; Vinderola, C.G. Growth of Lactobacillus rhamnosus 64 in whey permeate and study of the effect of mild stresses on survival to spray drying. LWT-Food Sci. Technol. 2015, 63, 322–330. [Google Scholar] [CrossRef]
- Bagad, M.; Pande, R.; Dubey, V.; Ghosh, A.S. Survivability of freeze-dried probiotic Pediococcus pentosaceus strains GS4, GS17 and Lactobacillus gasseri (ATCC 19992) during storage with commonly used pharmaceutical excipients within a period of 120 days. Asian Pac. J. Trop. Biomed. 2017, 7, 921–929. [Google Scholar] [CrossRef]
- Pyar, H.; Peh, K.K. Cost effectiveness of cryoprotective agents and modified De Man Rogosa Sharpe medium on growth of Lactobacillus acidophilus. Pak. J. Biol. Sci. 2014, 17, 462–471. [Google Scholar] [CrossRef]
- Duong, T.; Barrangou, R.; Russell, W.M.; Klaenhammer, T.R. Characterization of the tre locus and analysis of trehalose cryoprotection in Lactobacillus acidophilus NCFM. Appl. Env. Microbiol. 2006, 72, 1218–1225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jalali, M.; Abedi, D.; Varshosaz, J.; Najjarzadeh, M.; Mirlohi, M.; Tavakoli, N. Stability evaluation of freeze-dried Lactobacillus paracasei subsp. tolerance and Lactobacillus delbrueckii subsp. bulgaricus in oral capsules. Res. Pharm. Sci. 2012, 7, 31–36. [Google Scholar] [PubMed]
- Stefanello, R.F.; Nabeshima, E.H.; Iamanaka, T.; Ludwig, A.; Martins Fries, L.L.; Olivier Bernardi, A.; Venturini Copetti, M. Survival and stability of Lactobacillus fermentum and Wickerhamomyces anomalus strains upon lyophilisation with different cryoprotectant agents. Food Res. Int. 2019, 115, 90–94. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Tian, F.; Liu, X.; Zhao, J.; Zhang, H.; Chen, W. Effects of cryoprotectants on viability of Lactobacillus reuteri CICC6226. Appl. Microbiol. Biotechnol. 2011, 92, 609–616. [Google Scholar] [CrossRef]
- Savedboworn, W.; Teawsomboonkit, K.; Surichay, S.; Riansa-ngawong, W.; Rittisak, S.; Charoen, R.; Phattayakorn, K. Impact of protectants on the storage stability of freeze-dried probiotic Lactobacillus plantarum. Food Sci. Biotechnol. 2019, 28, 795–805. [Google Scholar] [CrossRef]
- Montel Mendoza, G.; Pasteris, S.; Otero, M.C.; Nader-Macías, M.E.F. Survival and beneficial properties of lactic acid bacteria from raniculture subjected to freeze-drying and storage. J. Appl. Microbiol. 2014, 116, 157–166. [Google Scholar] [CrossRef]
- JuárezTomás, M.S.; De Gregorio, P.R.; LecceseTerraf, M.C.; Nader-Macías, M.E. Encapsulation and subsequent freeze-drying of Lactobacillus reuteri CRL 1324 for its potential inclusion in vaginal probiotic formulations. Eur J Pharm Sci. 2015, 15, 87–95. [Google Scholar] [CrossRef] [Green Version]
- Miranda, M.H.; AristimuñoFicoseco, M.E.; Nader-Macias, M.E.F. Safety, environmental and technological characterization of beneficial autochthonous lactic bacteria, and their vaginal administration to pregnant cows for the design of homologous multi-strain probiotic formulas. Braz. J. Microbiol. 2021, 52, 2455–2473. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Run Order (Samples) | Glu % | HFCS % | Mol % | Pect % | Lac % | IMO % | Run Order (Samples) | Glu % | HFCS % | Mol % | Pect % | Lac % | IMO % |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| S1 | 0 | 0.5 | 0 | 1.0 | 0 | 0 | S31 | 0 | 0 | 0.75 | 0.75 | 0 | 0 |
| S2 | 0 | 0 | 0.5 | 1.0 | 0 | 0 | S32 | 0 | 0 | 0 | 0 | 0 | 1.5 |
| S3 | 0 | 0 | 0 | 0.5 | 0 | 1.0 | S33 | 0.75 | 0.75 | 0 | 0 | 0 | 0 |
| S4 | 0 | 0 | 0 | 0 | 1.5 | 0 | S34 | 0.25 | 0.25 | 0.25 | 0.25 | 0.25 | 0.25 |
| S5 | 0 | 0.75 | 0 | 0 | 0 | 0.75 | S35 | 0 | 0.5 | 0 | 0 | 1.0 | 0 |
| S6 | 0.25 | 0.25 | 0.25 | 0.25 | 0.25 | 0.25 | S36 | 0 | 0.75 | 0 | 0 | 0.75 | 0 |
| S7 | 0 | 0 | 0 | 1.5 | 0 | 0 | S37 | 0.25 | 0.25 | 0.25 | 0.25 | 0.25 | 0.25 |
| S8 | 0 | 0 | 0 | 0 | 0.75 | 0.75 | S38 | 1.0 | 0 | 0 | 0 | 0.5 | 0 |
| S9 | 0 | 1.0 | 0 | 0.5 | 0 | 0 | S39 | 1.0 | 0 | 0.5 | 0 | 0 | 0 |
| S10 | 0.25 | 0.25 | 0.25 | 0.25 | 0.25 | 0.25 | S40 | 0.5 | 0 | 0 | 0 | 0 | 1.0 |
| S11 | 0 | 0 | 0 | 0.75 | 0.75 | 0 | S41 | 0.5 | 1.0 | 0 | 0 | 0 | 0 |
| S12 | 1.5 | 0 | 0 | 0 | 0 | 0 | S42 | 0 | 0 | 1.0 | 0.5 | 0 | 0 |
| S13 | 0 | 1.5 | 0 | 0 | 0 | 0 | S43 | 0.25 | 0.25 | 0.25 | 0.25 | 0.25 | 0.25 |
| S14 | 0.5 | 0 | 0 | 1 | 0 | 0 | S44 | 0 | 0.5 | 1.0 | 0 | 0 | 0 |
| S15 | 0.75 | 0 | 0 | 0 | 0.75 | 0 | S45 | 0.5 | 0 | 0 | 0 | 1.0 | 0 |
| S16 | 0 | 0 | 0 | 0 | 0.5 | 1.0 | S46 | 0 | 0 | 0 | 1.0 | 0.5 | 0 |
| S17 | 0 | 0 | 0 | 0.5 | 1.0 | 0 | S47 | 0 | 0 | 0 | 0.75 | 0 | 0.75 |
| S18 | 0.25 | 0.25 | 0.25 | 0.25 | 0.25 | 0.25 | S48 | 0 | 0 | 1.0 | 0 | 0 | 0.5 |
| S19 | 0.25 | 0.25 | 0.25 | 0.25 | 0.25 | 0.25 | S49 | 0 | 1.0 | 0 | 0 | 0.5 | 0 |
| S20 | 0 | 0.5 | 0 | 0 | 0 | 1.0 | S50 | 0 | 0.75 | 0 | 0.75 | 0 | 0 |
| S21 | 0.75 | 0 | 0.75 | 0 | 0 | 0 | S51 | 0 | 0 | 0.75 | 0 | 0.75 | 0 |
| S22 | 0.25 | 0.25 | 0.25 | 0.25 | 0.25 | 0.25 | S52 | 0 | 0 | 1.0 | 0 | 0.5 | 0 |
| S23 | 0 | 0 | 1.5 | 0 | 0 | 0 | S53 | 0 | 1.0 | 0.5 | 0 | 0 | 0 |
| S24 | 0 | 1.0 | 0 | 0 | 0 | 0.5 | S54 | 0 | 0 | 0.75 | 0 | 0 | 0.75 |
| S25 | 1.0 | 0 | 0 | 0 | 0 | 0.5 | S55 | 1.0 | 0 | 0 | 0.5 | 0 | 0 |
| S26 | 0 | 0 | 0.5 | 0 | 1.0 | 0 | S56 | 0.25 | 0.25 | 0.25 | 0.25 | 0.25 | 0.25 |
| S27 | 0.75 | 0 | 0 | 0.75 | 0 | 0 | S57 | 0 | 0 | 0.5 | 0 | 0 | 1.0 |
| S28 | 0 | 0 | 0 | 1.0 | 0 | 0.5 | S58 | 0 | 0 | 0 | 0 | 1.0 | 0.5 |
| S29 | 0.5 | 0 | 1.0 | 0 | 0 | 0 | S59 | 0.75 | 0 | 0 | 0 | 0 | 0.75 |
| S30 | 0.5 | 1.0 | 0 | 0 | 0 | 0 | S60 | 0 | 0.75 | 0.75 | 0 | 0 | 0 |
| Probiotic Strains | Culture Media (Table 1) | Growth Parameters | ||
|---|---|---|---|---|
| Max OD560nm | t (h) * | μmax (h−1) | ||
| L. acidophilus CRL2074 | MRS | 1.24 ± 0.03 | 14 | 0.196 ± 0.02 |
| S60-0.75% HFCS + 0.75% Mol * | 1.17 ± 0.01 | 14 | 0.135 ± 0.01 | |
| S23-1.5% Mol | 0.90 ± 0.02 | 14 | 0.140 ± 0.01 | |
| S47-0.75% Pect + 0.75% IMO | 0.81 ± 0.04 | 14 | 0.056 ± 0.02 | |
| S20-0.5% HFCS + 1% IMO | 0.78 ± 0.01 | 14 | 0.047 ± 0.01 | |
| 3% Mol | 1.40 ± 0.01 | 14 | 0.160 ± 0.02 | |
| L. fermentum CRL2085 | MRS | 1.46 ± 0.01 | 14 | 0.260 ± 0.03 |
| S60-0.75% HFCS + 0.75% Mol * | 1.32 ± 0.02 | 13 | 0.480 ± 0.01 | |
| S10-0.25% all ingredients | 1.25 ± 0.03 | 8 | 0.400 ± 0.02 | |
| S23-1.5% Mol | 1.25 ± 0.01 | 14 | 0.334 ± 0.01 | |
| S36-0.75% HFCS + 75% Lac | 1.24 ± 0.01 | 12 | 0.457 ± 0.03 | |
| S48-1% Mol + 0.5% IMO | 1.24 ± 0.03 | 11 | 0.171 ± 0.03 | |
| S30-0.5% Gluc + 1% HFCS | 1.24 ± 0.03 | 9 | 0.256 ± 0.02 | |
| S52-1% Mol + 0.5% Lac | 1.24 ± 0.03 | 10 | 0.119 ± 0.02 | |
| S53-1% HFCS + 0.5% Mol | 1.20 ± 0.03 | 10 | 0.234 ± 0.01 | |
| 3% Mol | 1.60 ± 0.01 | 10 | 0.425 ± 0.01 | |
| L. mucosae CRL2069 | MRS | 1.68 ± 0.03 | 14 | 0.253 ± 0.02 |
| S56-0.25% all ingredients | 0.97 ± 0.02 | 6 | 0.315 ± 0.03 | |
| S5-0.75% HFCS + 0.75% IMO * | 0.93 ± 0.03 | 14 | 0.310 ± 0.01 | |
| S51-0.75 Mol + 0.75 Lac | 0.86 ± 0.01 | 7 | 0.231 ± 0.02 | |
| S52-1% Mol + 0.5% Lac | 0.86 ± 0.02 | 7 | 0.157 ± 0.02 | |
| S36-0.75 HFCS + 0.75% Lac | 0.78 ± 0.01 | 7 | 0.346 ± 0.05 | |
| 3% Mol | 1.56 ± 0.02 | 12 | 0.495 ± 0.01 | |
| 30 °C | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Incubation Conditions Probiotic Strains Time (h) | Static | Agitation | ||||||||||||||||
| CRL2074 | CRL2085 | CRL2069 | CRL2074 | CRL2085 | CRL2069 | CRL2074 | CRL2085 | CRL2069 | CRL2074 | CRL2085 | CRL2069 | |||||||
| Free pH | Controlled pH | Free pH | Controlled pH | |||||||||||||||
| DO560nm | pH | DO560nm | pH | DO560nm | pH | DO560nm | DO560nm | pH | DO560nm | pH | DO560nm | pH | DO560nm | |||||
| 0 | 0.001 | 6.50 | 0.001 | 6.50 | 0.001 | 6.50 | 0.001 | 0.001 | 0.001 | 0.001 | 6.50 | 0.001 | 6.50 | 0.001 | 6.70 | 0.001 | 0.001 | 0.001 |
| 3 | 0.149 | 6.45 | 0.062 | 6.48 | 0.127 | 6.58 | 0.091 | 0.119 | 0.124 | 0.225 | 6.48 | 0.081 | 6.45 | 0.175 | 6.48 | 0.186 | 0.060 | 0.195 |
| 6 | 0.101 | 6.04 | 0.298 | 6.08 | 0.353 | 5.99 | 0.171 | 0.128 | 0.326 | 0.770 | 5.46 | 0.223 | 6.32 | 0.793 | 5.29 | 0.768 | 0.207 | 0.854 |
| 9 | 0.695 | 5.40 | 0.628 | 5.63 | 0.835 | 5.14 | 0.840 | 0.757 | 0.950 | 1.095 | 4.74 | 0.816 | 5.61 | 1.225 | 4.58 | 0.920 | 0.758 | 0.875 |
| 24 | 1.298 | 4.26 | 1.280 | 4.69 | 1.410 | 4.12 | 1.322 | 1.193 | 1.178 | 1.109 | 4.65 | 1.149 | 4.94 | 1.351 | 4.22 | 1.005 | 0.907 | 1.146 |
| Biomass (g/L) | 12.8 | 8.0 | 12.4 | 10.0 | 8.6 | 9.8 | 8.2 | 7.2 | 8.4 | 9.0 | 7.4 | 7.9 | ||||||
| 37 °C | ||||||||||||||||||
| Incubation Conditions/ Probiotic Strains Time (h) | Static | Agitation | ||||||||||||||||
| CRL2074 | CRL2085 | CRL2069 | CRL2074 | CRL2085 | CRL2069 | CRL2074 | CRL2085 | CRL2069 | CRL2074 | CRL2085 | CRL2069 | |||||||
| Free pH | Controlled pH | Free pH | Controlled pH | |||||||||||||||
| DO560nm | pH | DO560nm | pH | DO560nm | pH | DO560nm | DO560nm | pH | DO560nm | pH | DO560nm | pH | DO560nm | |||||
| 0 | 0.001 | 6.50 | 0.001 | 6.50 | 0.001 | 6.50 | 0.001 | 0.001 | 0.001 | 0.001 | 6.50 | 0.001 | 6.50 | 0.001 | 6.50 | 0.001 | 0.001 | 0.001 |
| 3 | 0.057 | 6.40 | 0.037 | 6.38 | 0.079 | 6.47 | 0.030 | 0.033 | 0.045 | 0.081 | 6.47 | 0.054 | 6.48 | 0.041 | 6.37 | 0.036 | 0.030 | 0.095 |
| 6 | 0.115 | 6.00 | 0.140 | 6.10 | 0.181 | 6.20 | 0.116 | 0.071 | 0.159 | 0.192 | 6.10 | 0.104 | 6.20 | 0.125 | 6.10 | 0.106 | 0.087 | 0.116 |
| 9 | 0.501 | 5.70 | 0.340 | 5.40 | 0.831 | 5.50 | 0.386 | 0.277 | 0.602 | 0.582 | 5.70 | 0.328 | 5.75 | 0.483 | 5.80 | 0.319 | 0.415 | 0.466 |
| 24 | 1.367 | 4.10 | 1.3207 | 4.70 | 1.424 | 4.40 | 1.347 | 1.239 | 1.347 | 0.825 | 4.80 | 0.956 | 4.60 | 0.903 | 5.20 | 0.876 | 0.803 | 0.656 |
| Biomass (g/L) | 12.9 | 9.4 | 13.0 | 11.8 | 9.4 | 12.4 | 8.8 | 9.8 | 9.6 | 9.8 | 8.0 | 9.0 | ||||||
| Probiotic Strain | Growth Conditions | Incubation Time (h) | ||||
|---|---|---|---|---|---|---|
| 0 | 6 | 12 | 24 | |||
| L. acidophilus CRL2074 | Free pH | CFU/mL | 2.1 × 107 | 2.9 × 107 | 1.4 × 108 | 1.0 × 109 |
| pH | 6.5 | 5.5 | 4.4 | 3.6 | ||
| Biomass (g/L) | 12.25 | |||||
| Controlled pH | CFU/mL | 1.2 × 107 | 7.5 × 107 | 1.4 × 108 | 1.3 × 108 | |
| Biomass (g/L) | 10.00 | |||||
| L. fermentum CRL2085 | Free pH | CFU/mL | 1.8 × 107 | 2.65 × 109 | 8.0 × 109 | 1.5 × 1010 |
| pH | 6.5 | 4.5 | 4.3 | 4.4 | ||
| Biomass (g/L) | 11.50 | |||||
| Controlled pH | CFU/mL | 1.5 × 107 | 2.3 × 109 | 1.9 × 1010 | 8.65 × 1010 | |
| Biomass (g/L) | 12.95 | |||||
| L. mucosae CRL2069 | Free pH | CFU/mL | 4.3 × 107 | 1.6 × 1010 | 1.5 × 1010 | 1.2 × 1010 |
| pH | 6.5 | 4.2 | 3.4 | 3.8 | ||
| Biomass (g/L) | 12.40 | |||||
| Controlled pH | CFU/mL | 3.4 × 107 | 5.0 × 109 | 5.5 × 1010 | 2.3 × 1010 | |
| Biomass (g/L) | 18.20 | |||||
| Analyzed Conditions | Viability (log CFU/g) after Freeze Drying | ||||||
|---|---|---|---|---|---|---|---|
| Cryoprotectants a | |||||||
| Fruct | Sucr | Treh | MSG | WPC | WPC + Treh | MIX | |
| L. acidophilus CRL2074 | |||||||
| Free pH | <4 | <4 | 5.32 | 8.74 | 6.44 | 4.70 | 6.84 |
| Stability (30 days) | 8.41 | ||||||
| % Survival b | 96% | ||||||
| L. fermentum CRL2085 | |||||||
| Controlled pH | 5.74 | 5.94 | 6.80 | 5.47 | 3.89 | 5.08 | 9.96 |
| Stability (30 days) | 8.78 | ||||||
| % Survival | 88% | ||||||
| L. mucosae CRL2069 | |||||||
| Controlled pH | 4.73 | 4.11 | 6.77 | 9.17 | 6.83 | 9.99 | 6.49 |
| Stability (30 days) | 9.95 | ||||||
| Probiotic Strain | Culture Medium | Surface Properties (%) | ||
|---|---|---|---|---|
| Auto-Aggregation | Hydrophobicity | |||
| Xylene | Toluene | |||
| L. acidophilus CRL2074 | MRS | 21.6 (33) * | 8.0 (75) | 1.6 (73) |
| Mgl + 3% Mol | 51.5 | 5.7 | 1.4 | |
| L. fermentum CRL2085 | MRS | 83.3 (13) | 3.0 (0) | 1.0 (3) |
| Mgl + 3% Mol | 91.3 | 10.0 | 8.5 | |
| L. mucosae CRL20 | MRS | 80.0 (82) | 14.9 (73) | 5.3 (0) |
| Mgl + 3% Mol | 65.0 | 13.1 | 11.1 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aristimuño Ficoseco, C.; Mansilla, F.I.; Vignolo, G.M.; Nader-Macías, M.E.F. Optimization of Probiotic Lactobacilli Production for In-Feed Supplementation to Feedlot Cattle. Appl. Microbiol. 2023, 3, 339-357. https://doi.org/10.3390/applmicrobiol3020024
Aristimuño Ficoseco C, Mansilla FI, Vignolo GM, Nader-Macías MEF. Optimization of Probiotic Lactobacilli Production for In-Feed Supplementation to Feedlot Cattle. Applied Microbiology. 2023; 3(2):339-357. https://doi.org/10.3390/applmicrobiol3020024
Chicago/Turabian StyleAristimuño Ficoseco, Cecilia, Flavia I. Mansilla, Graciela M. Vignolo, and María E. Fátima Nader-Macías. 2023. "Optimization of Probiotic Lactobacilli Production for In-Feed Supplementation to Feedlot Cattle" Applied Microbiology 3, no. 2: 339-357. https://doi.org/10.3390/applmicrobiol3020024