
The Effects of Atrazine, Diuron, Fluazifop-P-butyl, Haloxyfop-P-methyl, and Pendimethalin on Soil Microbial Activity and Diversity
 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Herbicide Degradation over Time
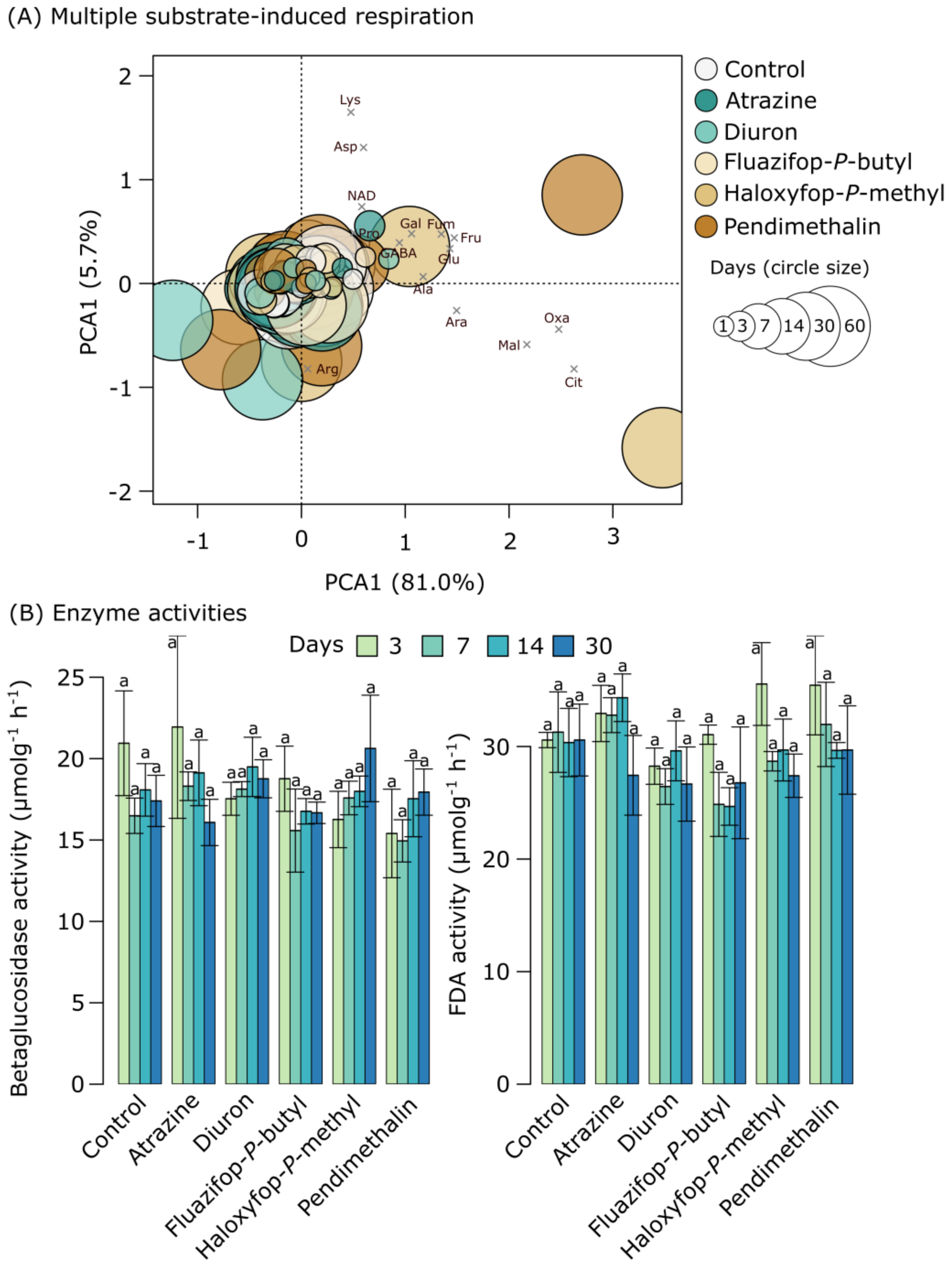
2.2. Soil Microbial Activity
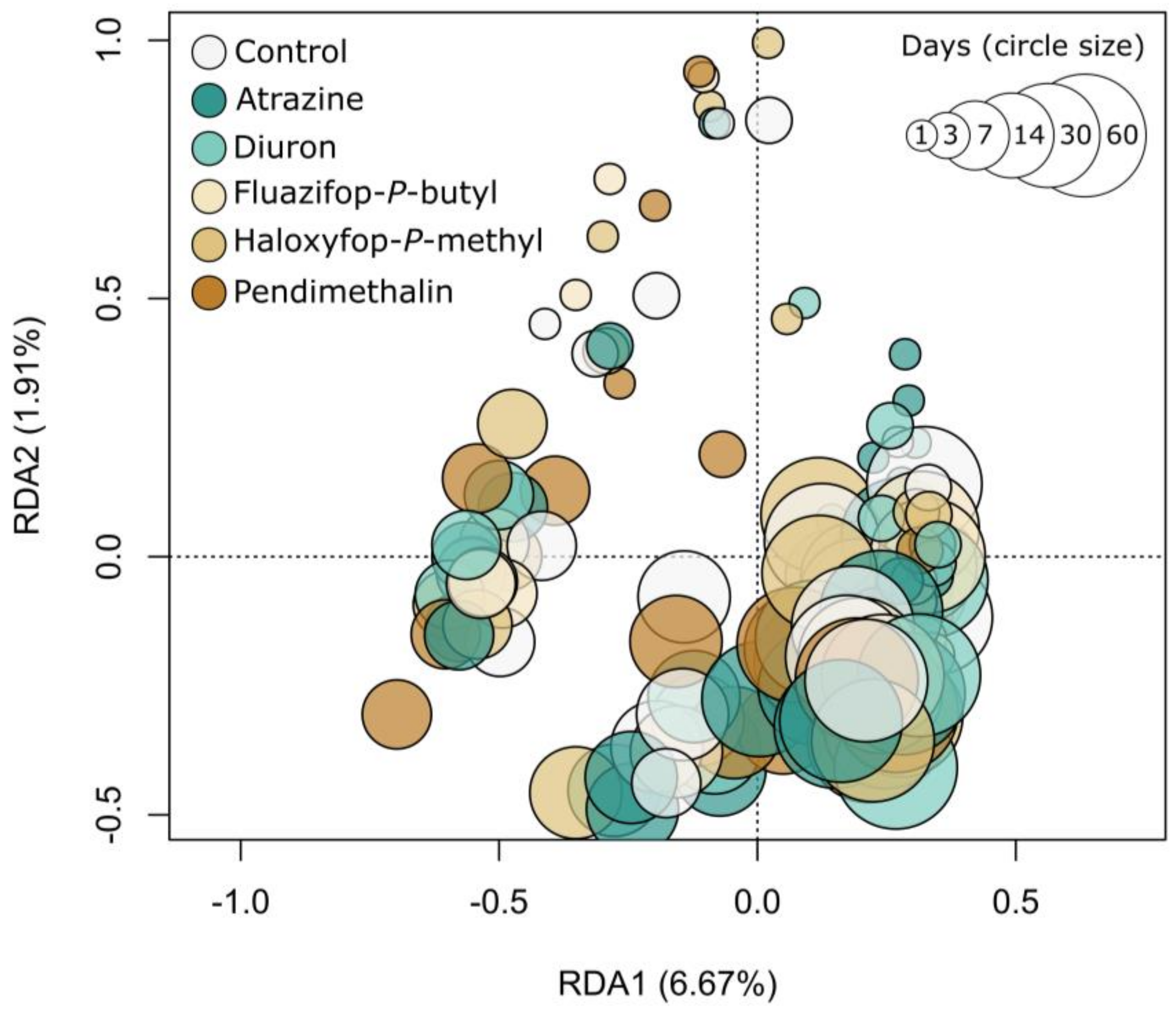
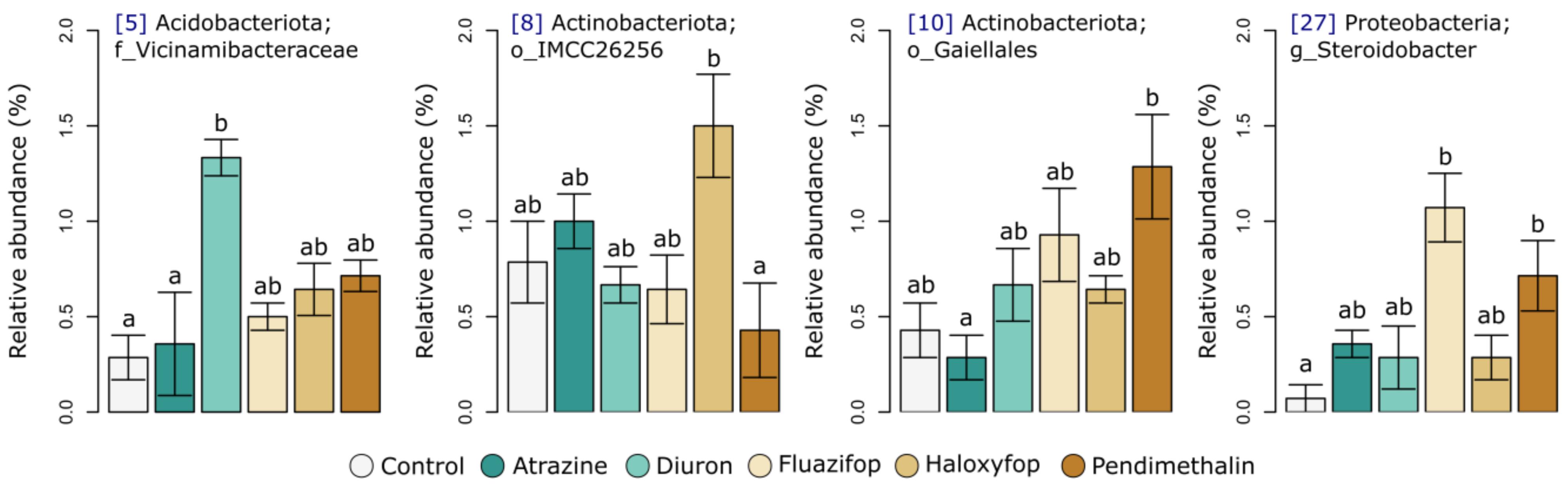
2.3. Soil Microbial Diversity
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Experimental Design and Soil Sampling
5.2. Liquid Chromatography–Mass Spectrometry–Mass Spectrometry (LC-MSMS)
5.3. Enzyme Assays
5.4. MicroResp
5.5. Microbial Diversity
5.6. Processing of Sequence Data
5.7. Statistical Analyses
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Imfeld, G.; Vuilleumier, S. Measuring the effects of pesticides on bacterial communities in soil: A critical review. Eur. J. Soil Biol. 2012, 49, 22–30. [Google Scholar] [CrossRef]
- Jacobsen, C.S.; Hjelmsø, M.H. Agricultural soils, pesticides and microbial diversity. Curr. Opin. Biotechnol. 2014, 27, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Ruuskanen, S.; Fuchs, B.; Nissinen, R.; Puigbo, P.; Rainio, M.; Saikkonen, K. Ecosystem consequences of herbicides: The role of microbiome. Trends Ecol. Evol. 2023, 38, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Dennis, P.G.; Kukulies, T.; Forstner, C.; Orton, T.G.; Pattison, A.B. The effects of glyphosate, glufosinate, paraquat and paraquat-diquat on soil microbial activity and bacterial, archaeal and nematode diversity. Sci. Rep. 2018, 8, 2119. [Google Scholar] [CrossRef] [Green Version]
- Tyler, H.L.; Khalid, S.; Jackson, C.R.; Moore, M.T. Determining potential for microbial atrazine degradation in agricultural drainage ditches. J. Environ. Qual. 2013, 42, 828. [Google Scholar] [CrossRef] [Green Version]
- Mahía, J.; González-Prieto, S.J.; Martín, A.; Bååth, E.; Díaz-Raviña, M. Biochemical properties and microbial community structure of five different soils after atrazine addition. Biol. Fertil. Soils. 2011, 47, 577–589. [Google Scholar] [CrossRef]
- Marchand, A.L.; Piutti, S.; Lagacherie, B.; Soulas, G. Atrazine mineralization in bulk soil and maize rhizosphere. Biol. Fertil. Soils. 2002, 35, 288–292. [Google Scholar] [CrossRef]
- Yale, R.L.; Sapp, M.; Sinclair, C.J.; Moir, J.W.B. Microbial changes linked to the accelerated degradation of the herbicide atrazine in a range of temperate soils. Environ. Sci. Pollut. Res. 2017, 24, 7359–7374. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, A.F.T.; Wang, P.; Staley, C.; Aparecida Silva Moretto, J.; Miguel Altarugio, L.; Chagas Campanharo, S.; Guedes Stehling, E.; Jay Sadowsky, M. Impact of atrazine exposure on the microbial community structure in a Brazilian tropical latosol soil. Microbes Environ. 2020, 35, ME19143. [Google Scholar] [CrossRef]
- Briceño, G.; Jorquera, M.A.; Demanet, R.; Mora, M.L.; Durán, N.; Palma, G. Effect of cow slurry amendment on atrazine dissipation and bacterial community structure in an agricultural Andisol. Sci. Total Environ. 2010, 408, 2833–2839. [Google Scholar] [CrossRef]
- Percich, J.A.; Lockwood, J.L. Interaction of atrazine with soil microorganisms: Population changes and accumulation. Can. J. Microbiol. 1978, 24, 1145–1152. [Google Scholar] [CrossRef]
- Tejada, M.; Morillo, E.; Gómez, I.; Madrid, F.; Undabeytia, T. Effect of controlled release formulations of diuron and alachlor herbicides on the biochemical activity of agricultural soils. J. Hazard Mater. 2017, 322, 334–347. [Google Scholar] [CrossRef] [Green Version]
- El Fantroussi, S.; Verschuere, L.; Verstraete, W.; Top, E.M. Effect of phenylurea herbicides on soil microbial communities estimated by analysis of 16S rRNA gene fingerprints and community-level physiological profiles. Appl. Environ. Microbiol. 1999, 65, 982–988. [Google Scholar] [CrossRef] [Green Version]
- Angly, F.E.; Pantos, O.; Morgan, T.C.; Rich, V.; Tonin, H.; Bourne, D.G.; Mercurio, P.; Negri, A.P.; Tyson, G.W. Diuron tolerance and potential degradation by pelagic microbiomes in the Great Barrier Reef lagoon. PeerJ 2016, 4, e1758. [Google Scholar] [CrossRef] [Green Version]
- Darine, T.; Alaeddine, C.; Fethi, B.; Ridha, M. Fluazifop-P-butyl (herbicide) affects richness and structure of soil bacterial communities. Soil Biol. Biochem. 2015, 81, 89–97. [Google Scholar] [CrossRef]
- Santos, J.B.; Jakelaitis, A.; Silva, A.A.; Costa, M.D.; Manabe, A.; Silva, M.C.S. Action of two herbicides on the microbial activity of soil cultivated with common bean (Phaseolus vulgaris) in conventional-till and no-till systems. Weed Res. 2006, 46, 284–289. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Mallek, A.Y.; Abdel-Kader, M.I.A.; Omar, S.A. Effect of the herbicide fluazifop-butyl on fungal populations and activity in soil. Water Air Soil Pollut. 1996, 86, 151–157. [Google Scholar] [CrossRef]
- Tu, C.M. Effect of three newer pesticides on microbial and enzymatic activities in soil. Bull. Environ. Contam. Toxicol. 2004, 49, 120–128. [Google Scholar] [CrossRef]
- Liang, Q.; Yan, Z.; Li, X. Influence of the herbicide haloxyfop-R-methyl on bacterial diversity in rhizosphere soil of Spartina alterniflora. Ecotoxicol. Environ. Saf. 2020, 194, 110366. [Google Scholar] [CrossRef]
- Kocčárek, M.; Artikov, H.; Vorříšek, K.; Borůvka, L. Pendimethalin degradation in soil and its interaction with soil microorganisms. Soil Water Res. 2016, 11, 213–219. [Google Scholar] [CrossRef]
- Singh, B.K.; Walker, A.; Wright, D.J. Persistence of chlorpyrifos, fenamiphos, chlorothalonil, and pendimethalin in soil and their effects on soil microbial characteristics. Bull. Environ. Contam. Toxicol. 2002, 69, 181–188. [Google Scholar] [CrossRef] [PubMed]
- Nayak, B.S.; Prusty, J.C.; Monhanty, S.K. Effect of herbicides on bacteria, fungi and actinomycetes in sesame (Sesamum indicum) soil. Ind. J. Agric. Sci. 1994, 64, 888–890. [Google Scholar]
- Chikoye, D.; Abaidoo, R.; Fontem, L.A. Response of weeds and soil microorganisms to imazaquin and pendimethalin in cowpea and soybean. Crop Prot. 2014, 65, 168–172. [Google Scholar] [CrossRef]
- Eivazi, F.; Tabatabai, M.A. Glucosidases and galactosidases in soils. Soil Biol. Biochem. 1988, 20, 601–606. [Google Scholar] [CrossRef]
- Schnürer, J.; Rosswall, T. Fluorescein diacetate hydrolysis as a measure of total microbial activity in soil and litter. Appl. Environ. Microbiol. 1982, 43, 1256–1261. [Google Scholar] [CrossRef] [Green Version]
- Campbell, C.D.; Chapman, S.J.; Cameron, C.M.; Davidson, M.S.; Potts, J.M. A rapid microtiter plate method to measure carbon dioxide evolved from carbon substrate amendments so as to determine the physiological profiles of soil microbial communities by using whole soil. Appl. Environ. Microbiol. 2003, 69, 3593–3599. [Google Scholar] [CrossRef] [Green Version]
- Engelbrektson, A.; Kunin, V.; Wrighton, K.C.; Zvenigorodsky, N.; Chen, F.; Ochman, H.; Hugenholtz, P. Experimental factors affecting PCR-based estimates of microbial species richness and evenness. ISME J. 2010, 4, 642–647. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2012, 41, D590–D596. [Google Scholar] [CrossRef]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef]
- Zhang, Z.; Schwartz, S.; Wagner, L.; Miller, W. A greedy algorithm for aligning DNA sequences. J. Comput. Biol. 2000, 7, 203–214. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Herbicide Active | Corresponding Commercial Herbicide | Active Concentration in Commercial Herbicide (g/L or kg) | Upper Limit of Recommended Application Rate for Each Commercial Herbicide (L or kg/ha) | Concentration of Herbicide Active Applied to Soil (ppm) | Predicted Half-Life (days) | Reported Half-Life (days) |
|---|---|---|---|---|---|---|
| Atrazine | Atrazine 900 WG | 900 | 3.30 | 30.28 | 49.9 | 15–100 |
| Diuron | Diuron 900 WG | 900 | 1.90 | 17.43 | 90.8 | 79–108 |
| Fluazifop-P-butyl | Fusilade | 128 | 1.65 | 2.15 | 31.6 | 7–21 |
| Haloxyfop-P-methyl | Verdict | 520 | 0.80 | 4.24 | 66.0 | 55–100 |
| Pendimethalin | Stomp 330 EC | 330 | 12.00 | 40.37 | n/a | 44 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dennis, P.G.; Kukulies, T.; Forstner, C.; Plisson, F.; Eaglesham, G.; Pattison, A.B. The Effects of Atrazine, Diuron, Fluazifop-P-butyl, Haloxyfop-P-methyl, and Pendimethalin on Soil Microbial Activity and Diversity. Appl. Microbiol. 2023, 3, 79-89. https://doi.org/10.3390/applmicrobiol3010007
Dennis PG, Kukulies T, Forstner C, Plisson F, Eaglesham G, Pattison AB. The Effects of Atrazine, Diuron, Fluazifop-P-butyl, Haloxyfop-P-methyl, and Pendimethalin on Soil Microbial Activity and Diversity. Applied Microbiology. 2023; 3(1):79-89. https://doi.org/10.3390/applmicrobiol3010007
Chicago/Turabian StyleDennis, Paul G., Tegan Kukulies, Christian Forstner, Fabien Plisson, Geoff Eaglesham, and Anthony B. Pattison. 2023. "The Effects of Atrazine, Diuron, Fluazifop-P-butyl, Haloxyfop-P-methyl, and Pendimethalin on Soil Microbial Activity and Diversity" Applied Microbiology 3, no. 1: 79-89. https://doi.org/10.3390/applmicrobiol3010007