
Antioxidant Potential Profile of Portuguese Wheat (Bread and Durum) Germplasm
 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Preparation of Plant Material
2.3. Chemicals and Reagents
2.4. Preparation of Wheat Flour Extracts
2.5. Preparation of the Ferulic Acid Standard and Calibration Curve
2.6. Analysis of Free, Bound, and Total Phenolics Using the Folin–Ciocalteu Method
2.7. Determination of Ferulic Acid Content Using HPLC
2.8. Determination of Total Antioxidant Activity
2.8.1. DPPH Radical Scavenging Assay
2.8.2. ABTS Assay
2.9. Reducing Power Method—Fe3+ to Fe2+ Activity Assay
2.10. Statistics
3. Results
3.1. Total Phenolics Content (TPC)
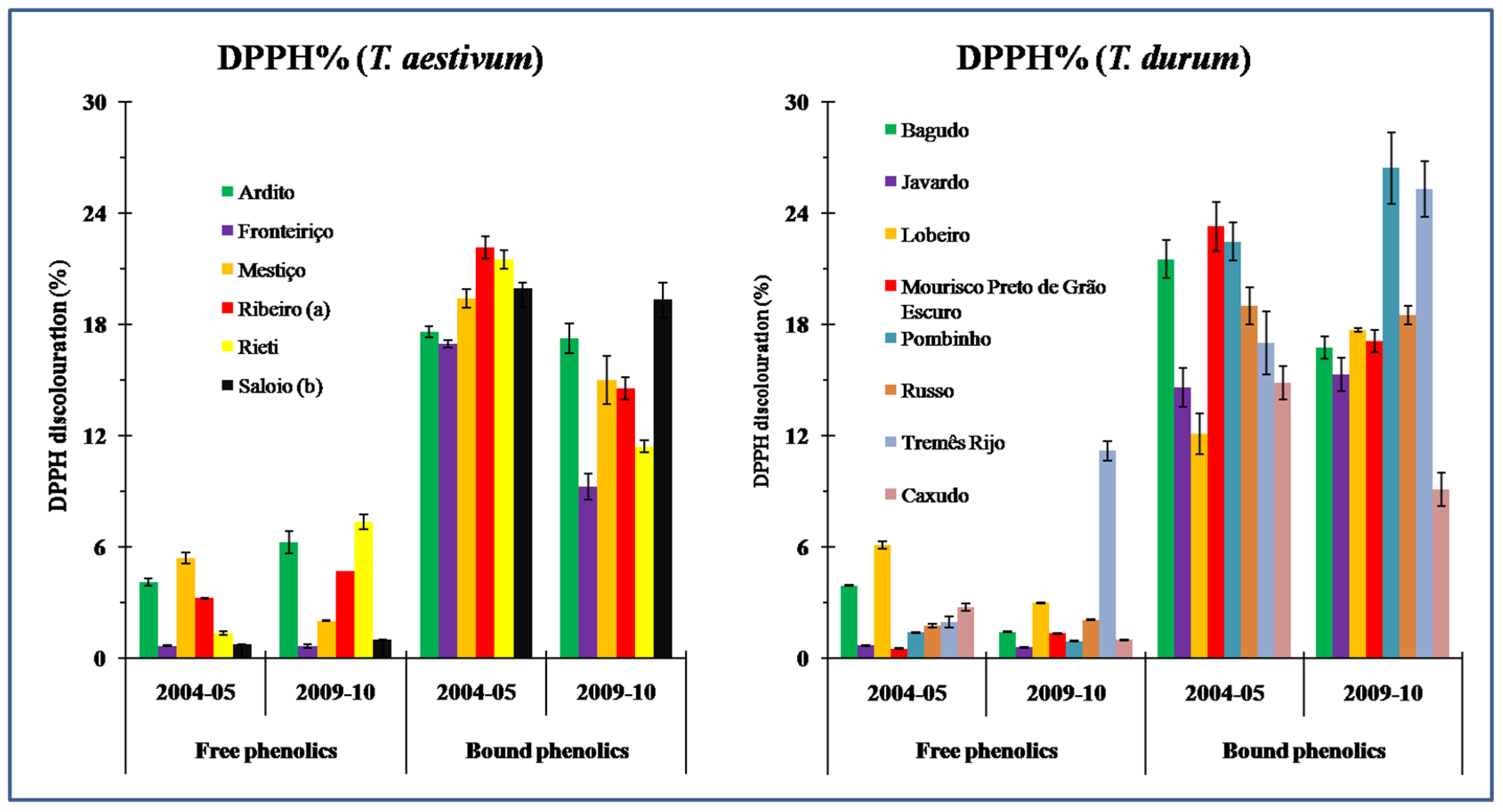
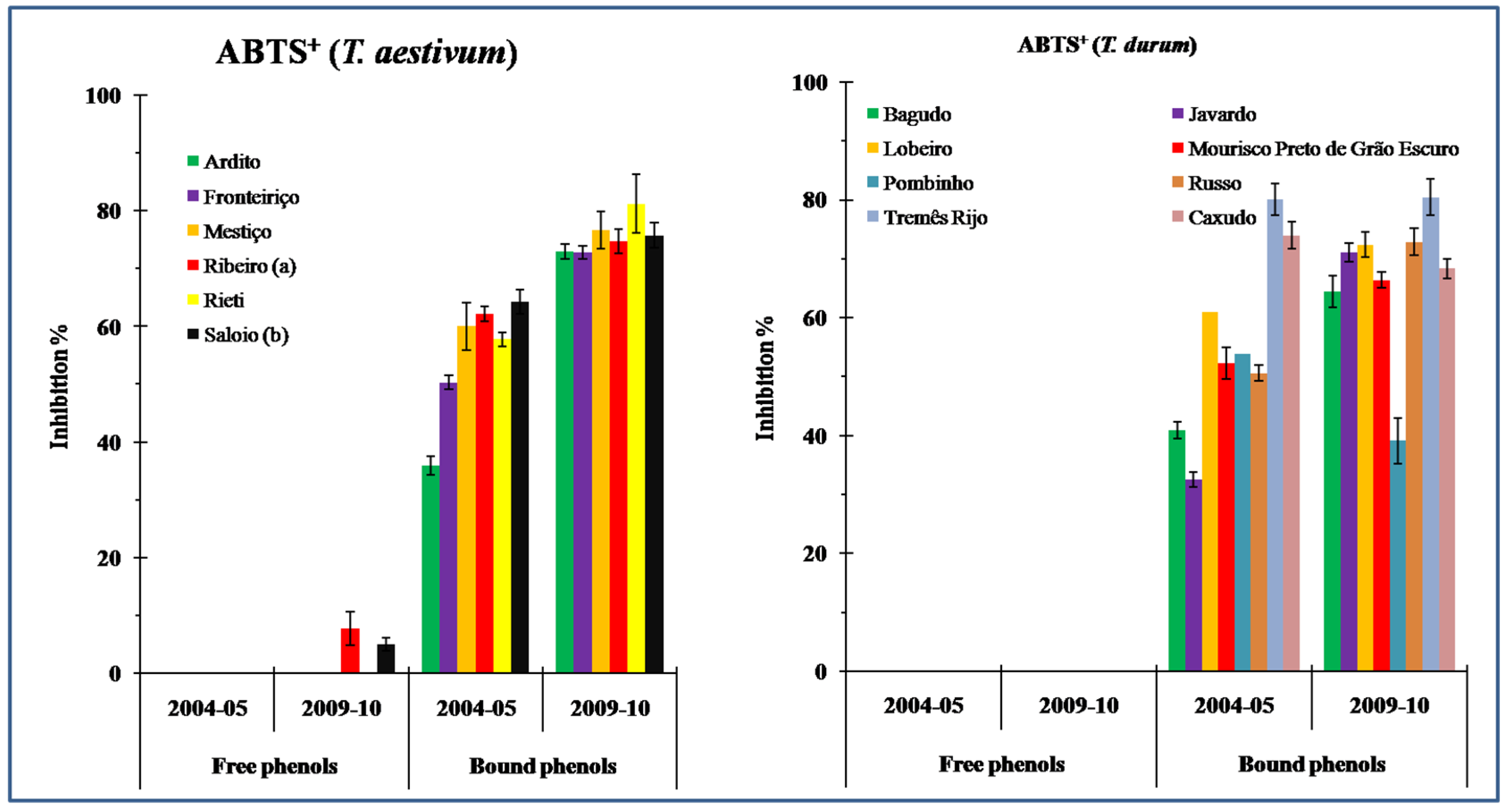
3.2. Antioxidant Capacity (AC)
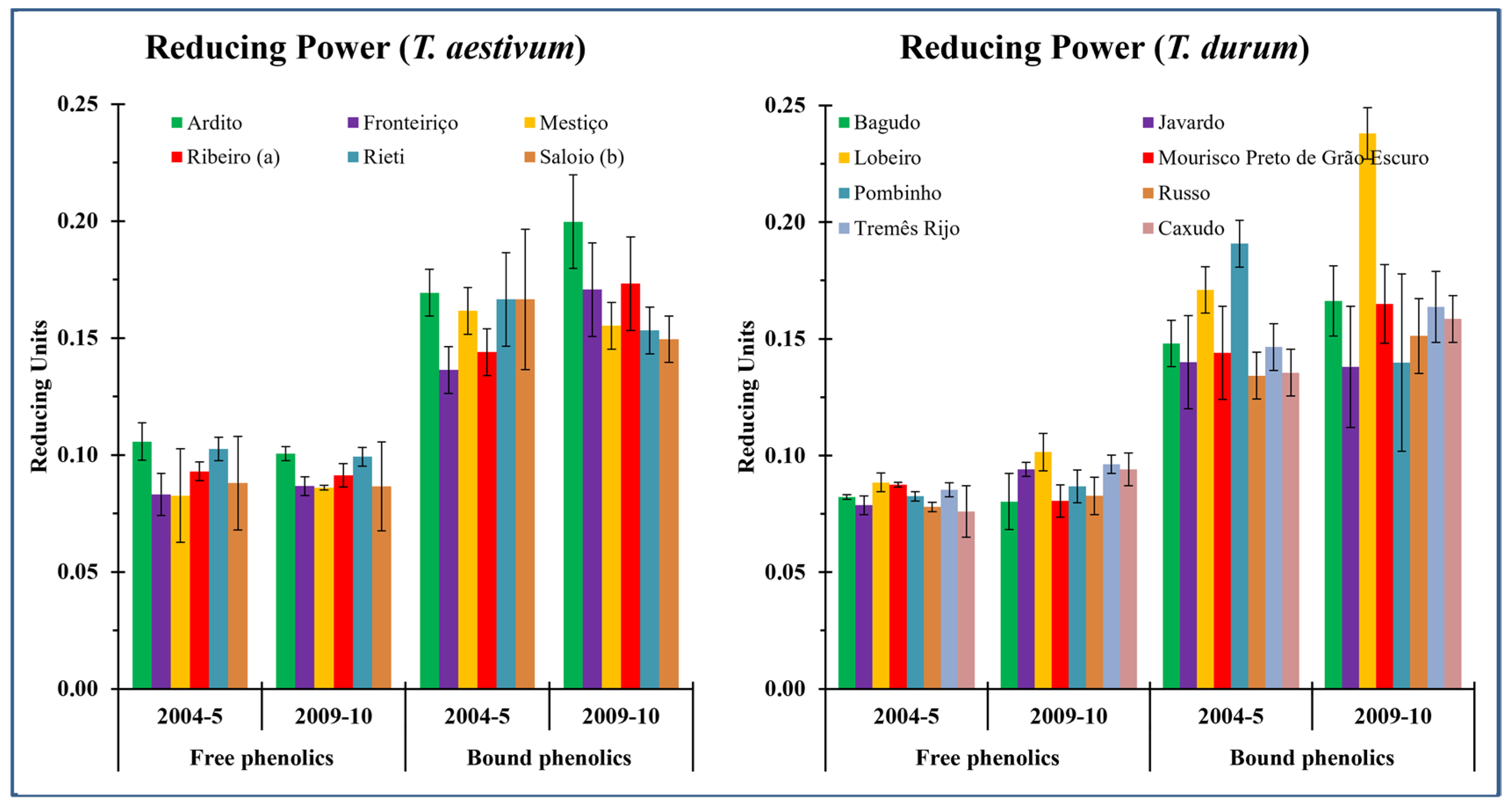
3.3. Reducing Power (RP)
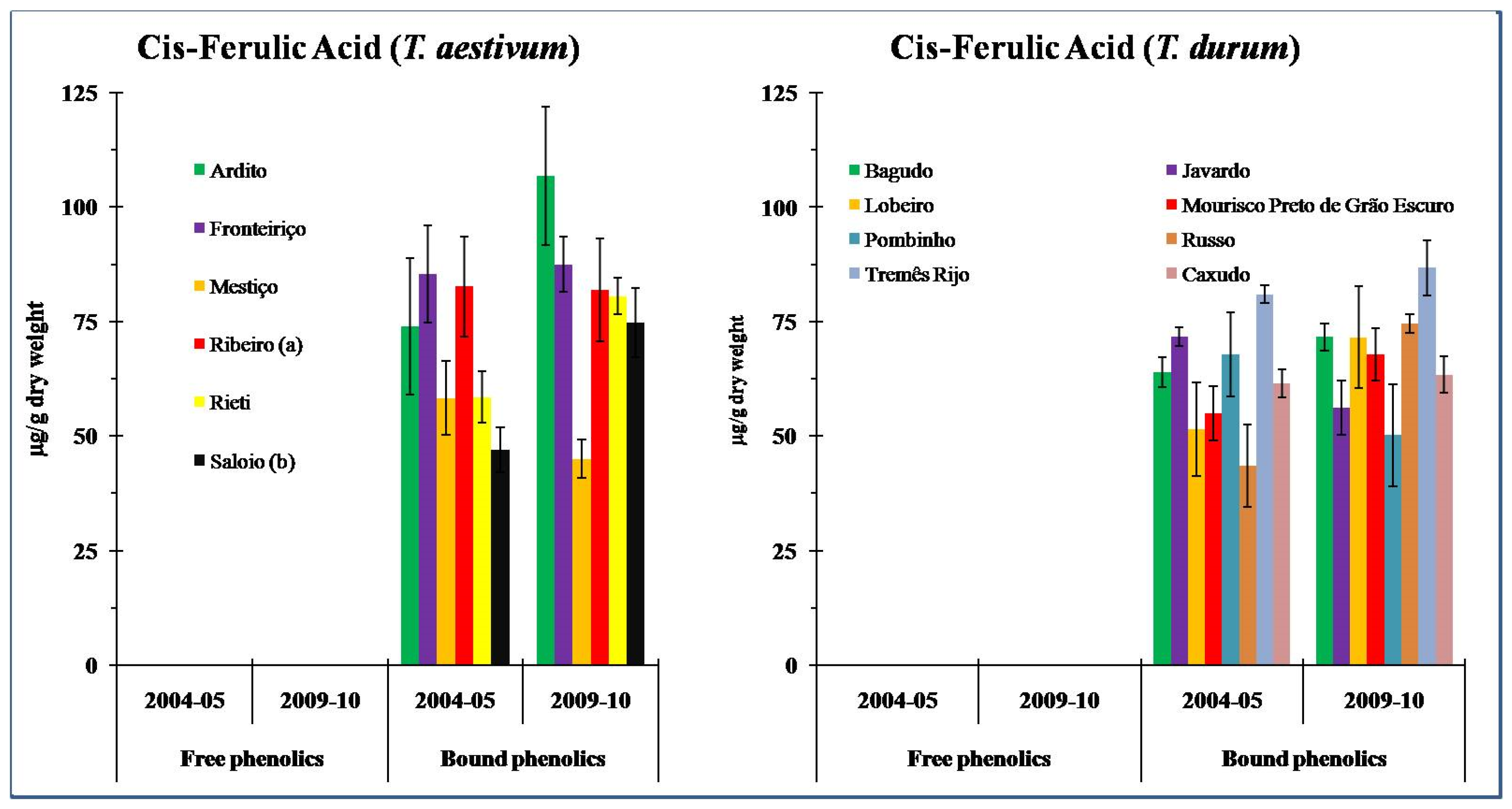
3.4. Ferulic Acid (FA) Content
4. Discussion
4.1. Phenolic Content in Free and Bound Forms in Wheat
4.2. Antioxidant Capacity (AC) in Wheat
4.3. Content and Contribution of Ferulic Acid Isomers in Wheat
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bhattacharyya, A.; Chattopadhyay, R.; Mitra, S.; Crowe, S.E. Oxidative Stress: An Essential Factor in the Pathogenesis of Gastrointestinal Mucosal Diseases. Physiol. Rev. 2014, 94, 329–354. [Google Scholar] [CrossRef] [PubMed]
- Pichersky, E.; Gang, D.R. Genetics and biochemistry of secondary metabolites in plants: An evolutionary perspective. Trends Plant Sci. 2000, 5, 439–445. [Google Scholar] [CrossRef] [PubMed]
- Griendling, K.; Fitzgerald, G.A. Oxidative Stress and Cardiovascular Injury: Part II: Animal and human studies. Circulation 2003, 108, 1912–1916. [Google Scholar] [CrossRef] [PubMed]
- Bandyopadhyay, D.; Chattopadhyay, A.; Ghosh, G.; Datta, A. Oxidative Stress-Induced Ischemic Heart Disease: Protection by Antioxidants. Curr. Med. Chem. 2004, 11, 369–387. [Google Scholar] [CrossRef]
- He, S.; Liu, F.; Xu, L.; Yin, P.; Li, D.; Mei, C.; Jiang, L.; Ma, Y.; Xu, J. Protective Effects of Ferulic Acid against Heat Stress-Induced Intestinal Epithelial Barrier Dysfunction In Vitro and In Vivo. PLoS ONE 2016, 11, e0145236. [Google Scholar] [CrossRef]
- Abraão, A.S.; Fernandes, N.; Silva, A.M.; Domínguez-Perles, R.; Barros, A. Prunus lusitanica L. Fruits as a Novel Source of Bioactive Compounds with Antioxidant Potential: Exploring the Unknown. Antioxidants 2022, 11, 1738. [Google Scholar] [CrossRef]
- Fischer, R.A.; Edmeades, G.O. Breeding and Cereal Yield Progress. Crop. Sci. 2010, 50, S85–S98. [Google Scholar] [CrossRef]
- Gülçin, I. Antioxidant activity of food constituents: An overview. Arch. Toxicol. 2012, 86, 345–391. [Google Scholar] [CrossRef]
- So, Y.B.; Woong, B.J.; Dong, S.K.; Hwa, Y.H.; Yong, W.S. Antioxidant activity and total phenolic compounds in grain extracts of wheat, barley, and oat. Korean J. Crop Sci. 2002, 47, 102–107. [Google Scholar]
- Beta, T.; Nam, S.; Dexter, J.E.; Sapirstein, H.D. Phenolic Content and Antioxidant Activity of Pearled Wheat and Roller-Milled Fractions. Cereal Chem. 2005, 82, 390–393. [Google Scholar] [CrossRef]
- Li, W.; Shan, F.; Sun, S.; Corke, H.; Beta, T. Free Radical Scavenging Properties and Phenolic Content of Chinese Black-Grained Wheat. J. Agric. Food Chem. 2005, 53, 8533–8536. [Google Scholar] [CrossRef] [PubMed]
- Goufo, P.; Trindade, H. Rice antioxidants: Phenolic acids, flavonoids, anthocyanins, proanthocyanidins, tocopherols, tocotrienols, γ-oryzanol, and phytic acid. Food Sci. Nutr. 2014, 2, 75–104. [Google Scholar] [CrossRef] [PubMed]
- Liyana-Pathirana, C.M.; Shahidi, F. Antioxidant Activity of Commercial Soft and Hard Wheat (Triticum aestivum L.) as Affected by Gastric pH Conditions. J. Agric. Food Chem. 2005, 53, 2433–2440. [Google Scholar] [CrossRef] [PubMed]
- Irakli, M.N.; Samanidou, V.F.; Biliaderis, C.; Papadoyannis, I.N. Development and validation of an HPLC-method for determination of free and bound phenolic acids in cereals after solid-phase extraction. Food Chem. 2012, 134, 1624–1632. [Google Scholar] [CrossRef]
- Fincher, G.B. Ferulic acid in barley cell walls: A fluorescence study. J. Inst. Brew. 1976, 82, 347–349. [Google Scholar] [CrossRef]
- Stewart, D.; Robertson, G.W.; Morrison, I.M. Phenolic acids dimers in the cell walls of barley. J. Mass Spectrom. 1994, 23, 71–74. [Google Scholar] [CrossRef]
- Sun, R.; Sun, X.; Wang, S.; Zhu, W.; Wang, X. Ester and ether linkages between hydroxycinnamic acids and lignins from wheat, rice, rye, and barley straws, maize stems, and fast-growing poplar wood. Ind. Crop. Prod. 2002, 15, 179–188. [Google Scholar] [CrossRef]
- van Ruth, S.; Villegas, B.; Akkermans, W.; Rozijn, M.; van der Kamp, H.; Koot, A. Prediction of the identity of fats and oils by their fatty acid, triacylglycerol and volatile compositions using PLS-DA. Food Chem. 2010, 118, 948–955. [Google Scholar] [CrossRef]
- Lv, J.; Lu, Y.; Niu, Y.; Whent, M.; Ramadan, M.F.; Costa, J.; Yu, L. Effect of genotype, environment, and their interaction on phytochemical compositions and antioxidant properties of soft winter wheat flour. Food Chem. 2013, 138, 454–462. [Google Scholar] [CrossRef]
- Avramiuc, M. The influence of viability on phenolic content, conductivity and sugars efflux, and the relation-ships between these indices in wheat seeds. Food Environ. Saf. 2014, 13, 116–121. [Google Scholar]
- Narwal, S.; Thakur, V.; Sheoran, S.; Dahiya, S.; Jaswal, S.; Gupta, R.K. Antioxidant activity and phenolic content of the Indian wheat varieties. J. Plant Biochem. Biotechnol. 2014, 23, 11–17. [Google Scholar] [CrossRef]
- Yiğit, A.; Erekul, O. Antioxidant Activity and Essential Amino acid Content of Bread Wheat (Triticum aestivum L.) Varieties. J. Agric. Sci. 2023, 29, 130–141. [Google Scholar] [CrossRef]
- Garcia-Oliveira, A.L.; Machado, N.; Barros, A.; Carnide, V.; Lima-Brito, J. Genotypic Variation For Carotenoids Content and Chemometric Model Development For Seed Quality Parameters in Wheat. Procedia Environ. Sci. 2015, 29, 162–163. [Google Scholar] [CrossRef]
- de Vasconcelos, M.; Bennett, R.; Castro, C.; Cardoso, P.; Saavedra, M.; Rosa, E. Study of composition, stabilization and processing of wheat germ and maize industrial by-products. Ind. Crop. Prod. 2012, 42, 292–298. [Google Scholar] [CrossRef]
- Huang, D.; Ou, B.; Prior, R.L. The Chemistry behind Antioxidant Capacity Assays. J. Agric. Food Chem. 2005, 53, 1841–1856. [Google Scholar] [CrossRef]
- Chandrasekara, A.; Shahidi, F. Content of Insoluble Bound Phenolics in Millets and Their Contribution to Antioxidant Capacity. J. Agric. Food Chem. 2010, 58, 6706–6714. [Google Scholar] [CrossRef]
- Stratil, P.; Klejdus, B.; Kubáň, V. Determination of Total Content of Phenolic Compounds and Their Antioxidant Activity in VegetablesEvaluation of Spectrophotometric Methods. J. Agric. Food Chem. 2006, 54, 607–616. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Sanchez-Moreno, C.; Larrauri, J.A.; Saura-Calixto, F.A. A procedure to measure the antiradical efficiency of polyphenols. J. Sci. Food Agric. 1998, 76, 270–276. [Google Scholar] [CrossRef]
- Siddhuraju, P.; Becker, K. Antioxidant Properties of Various Solvent Extracts of Total Phenolic Constituents from Three Different Agroclimatic Origins of Drumstick Tree (Moringa oleifera Lam.) Leaves. J. Agric. Food Chem. 2003, 51, 2144–2155. [Google Scholar] [CrossRef]
- Gülçin, I.; Mshvildadze, V.; Gepdiremen, A.; Elias, R. Screening of antiradical and antioxidant activity of monodesmosides and crude extract from Leontice smirnowii tuber. Phytomedicine 2006, 13, 343–351. [Google Scholar] [CrossRef] [PubMed]
- Hinneburg, I.; Dorman, H.D.; Hiltunen, R. Antioxidant activities of extracts from selected culinary herbs and spices. Food Chem. 2006, 97, 122–129. [Google Scholar] [CrossRef]
- Antolovich, M.; Prenzler, P.D.; Patsalides, E.; McDonald, S.; Robards, K. Methods for testing antioxidant activity. Analyst 2001, 127, 183–198. [Google Scholar] [CrossRef] [PubMed]
- McCord, J.M. The evolution of free radicals and oxidative stress. Am. J. Med. 2000, 108, 652–659. [Google Scholar] [CrossRef]
- Shahidi, F.; Naczk, M. Phenolics in Food and Nutraceuticals; CRC Press: Boca Raton, FL, USA, 2004; pp. 1–545. [Google Scholar]
- Menga, V.; Fares, C.; Troccoli, A.; Cattivelli, L.; Baiano, A. Effects of genotype, location and baking on the phenolic content and some antioxidant properties of cereal species. Int. J. Food Sci. Technol. 2010, 45, 7–16. [Google Scholar] [CrossRef]
- Adom, K.K.; Liu, R.H. Antioxidant Activity of Grains. J. Agric. Food Chem. 2002, 50, 6182–6187. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, L.; Yao, Y.; Yan, J.; He, Z. Phenolic acid profiles of Chinese wheat cultivars. J. Cereal Sci. 2012, 56, 629–635. [Google Scholar] [CrossRef]
- Rocchetti, G.; Gregorio, R.P.; Lorenzo, J.M.; Barba, F.J.; Oliveira, P.G.; Prieto, M.A.; Simal-Gandara, J.; Mosele, J.I.; Motilva, M.; Tomas, M.; et al. Functional implications of bound phenolic compounds and phenolics–food interaction: A review. Compr. Rev. Food Sci. Food Saf. 2022, 21, 811–842. [Google Scholar] [CrossRef]
- Jacobs, D.R., Jr.; Meyer, K.A.; Kushi, L.H.; Folsom, A.R. Whole-grain intake may reduce the risk of ischemic heart disease death in postmenopausal women: The Iowa Women’s Health Study. Am. J. Clin. Nutr. 1998, 68, 248–257. [Google Scholar] [CrossRef]
- Maillard, M.-N.; Berset, C. Evolution of Antioxidant Activity during Kilning: Role of Insoluble Bound Phenolic Acids of Barley and Malt. J. Agric. Food Chem. 1995, 43, 1789–1793. [Google Scholar] [CrossRef]
- Revanappa, S.; Salimath, P.V. Phenolic acid profiles and antioxidant activities of different wheat (Triticum aestivum L.) varieties. J. Food Biochem. 2011, 35, 759–775. [Google Scholar] [CrossRef]
- Verma, B.; Hucl, P.; Chibbar, R.N. Phenolic Content and Antioxidant Properties of Bran in 51 Wheat Cultivars. Cereal Chem. 2008, 85, 544–549. [Google Scholar] [CrossRef]
- Mpofu, A.; Sapirstein, H.D.; Beta, T. Genotype and Environmental Variation in Phenolic Content, Phenolic Acid Composition, and Antioxidant Activity of Hard Spring Wheat. J. Agric. Food Chem. 2006, 54, 1265–1270. [Google Scholar] [CrossRef] [PubMed]
- Ma, D.; Wang, C.; Feng, J.; Xu, B. Wheat grain phenolics: A review on composition, bioactivity, and influencing fac-tors. J. Sci. Food Agric. 2021, 101, 6167–6185. [Google Scholar] [CrossRef]
- Abdel-Aal, E.-S.M.; Hucl, P.; Sosulski, F.W.; Graf, R.; Gillott, C.; Pietrzak, L. Screening Spring Wheat for Midge Resistance in Relation to Ferulic Acid Content. J. Agric. Food Chem. 2001, 49, 3559–3566. [Google Scholar] [CrossRef] [PubMed]
- Adom, K.K.; Sorrells, M.E.; Liu, R.H. Phytochemical Profiles and Antioxidant Activity of Wheat Varieties. J. Agric. Food Chem. 2003, 51, 7825–7834. [Google Scholar] [CrossRef]
- Tian, W.; Chen, G.; Gui, Y.; Zhang, G.; Li, Y. Rapid quantification of total phenolics and ferulic acid in whole wheat using UV–Vis spectrophotometry. Food Control 2020, 123, 107691. [Google Scholar] [CrossRef]
- Laddomada, B.; Caretto, S.; Mita, G. Wheat bran phenolic acids: Bioavailability and stability in whole wheat-based foods. Molecules 2015, 20, 15666–15685. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Wheat Species | Crop Season | Free Phenolics | Bound Phenolics | Total Phenolics | |||
|---|---|---|---|---|---|---|---|
| Range | Average | Range | Average | Range | Average | ||
| T. aestivum (n = 46) | 2004-2005 | 58–252 | 122.2 ± 7.8 | 292–1160 | 815.4 ± 25.9 | 378–1253 | 937.6 ± 26.9 |
| 2009-2010 | 200–371 | 268.0 ± 6.3 | 492–1033 | 774.3 ± 16.3 | 741–1261 | 1018.5 ± 28.7 | |
| Combined | 119–286 | 193.4 ± 5.7 | 485–1029 | 797.7 ± 17.1 | 650–1148 | 991.1 ± 16.9 | |
| T. durum (n = 46) | 2004-2005 | 167–430 | 246.7 ± 9.2 | 310–880 | 660.1 ± 19.1 | 668–1230 | 906.8 ± 20.6 |
| 2009-2010 | 93–616 | 304.0 ± 20.9 | 176–613 | 362.7 ± 13.6 | 398–1098 | 666.7 ± 22.7 | |
| Combined | 152–412 | 275.3 ± 10.3 | 389–659 | 511.4 ± 9.3 | 638–966 | 786.7 ± 11.9 | |
| Wheat Species | Crop Season | Ferulic Acid (μg/g Dry Matter) | |||||
|---|---|---|---|---|---|---|---|
| Free Phenolics | Bound Phenolics | Total | |||||
| Range | Average | Range | Average | Range | Average | ||
| T. aestivum (n = 6) | 2004-2005 | 14–47 | 29.7 ± 5.7 | 526–956 | 773.2 ± 93.0 | 573–971 | 803 ± 63.1 |
| 2009-2010 | 23–40 | 28.3 ± 2.6 | 449–1141 | 860.4 ± 150.9 | 478–1181 | 889 ± 95.9 | |
| Combined | 19–42 | 29.0 ± 3.8 | 552–986 | 816.8 ± 72.2 | 580–1027 | 846 ± 71.7 | |
| T. durum (n = 8) | 2004-2005 | 3–35 | 20.4 ± 3.7 | 470–1021 | 671.4 ± 60.0 | 485–1024 | 632 ± 58.4 |
| 2009-2010 | 11–35 | 20.0 ± 3.0 | 509–973 | 748.7 ± 52.1 | 520–997 | 1010 ± 54.0 | |
| Combined | 13–32 | 20.2 ± 2.7 | 619–997 | 710.0 ± 44.3 | 632–1010 | 730 ± 44.3 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garcia-Oliveira, A.L.; Aires, A.; Carvalho, R.; Carnide, V.; Barros, A.; Lima-Brito, J. Antioxidant Potential Profile of Portuguese Wheat (Bread and Durum) Germplasm. Crops 2023, 3, 124-135. https://doi.org/10.3390/crops3020012
Garcia-Oliveira AL, Aires A, Carvalho R, Carnide V, Barros A, Lima-Brito J. Antioxidant Potential Profile of Portuguese Wheat (Bread and Durum) Germplasm. Crops. 2023; 3(2):124-135. https://doi.org/10.3390/crops3020012
Chicago/Turabian StyleGarcia-Oliveira, Ana Luísa, Alfredo Aires, Rosa Carvalho, Valdemar Carnide, Ana Barros, and José Lima-Brito. 2023. "Antioxidant Potential Profile of Portuguese Wheat (Bread and Durum) Germplasm" Crops 3, no. 2: 124-135. https://doi.org/10.3390/crops3020012