
Dynamics of Micronutrient Uptake and Removal by Three Modern Runner Peanut Cultivars

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Site, Climate, and Soil
2.2. Plant Measurements and Analysis
2.3. Pod and Kernel Yields and Nutrient Removal
2.4. Statistical Analysis
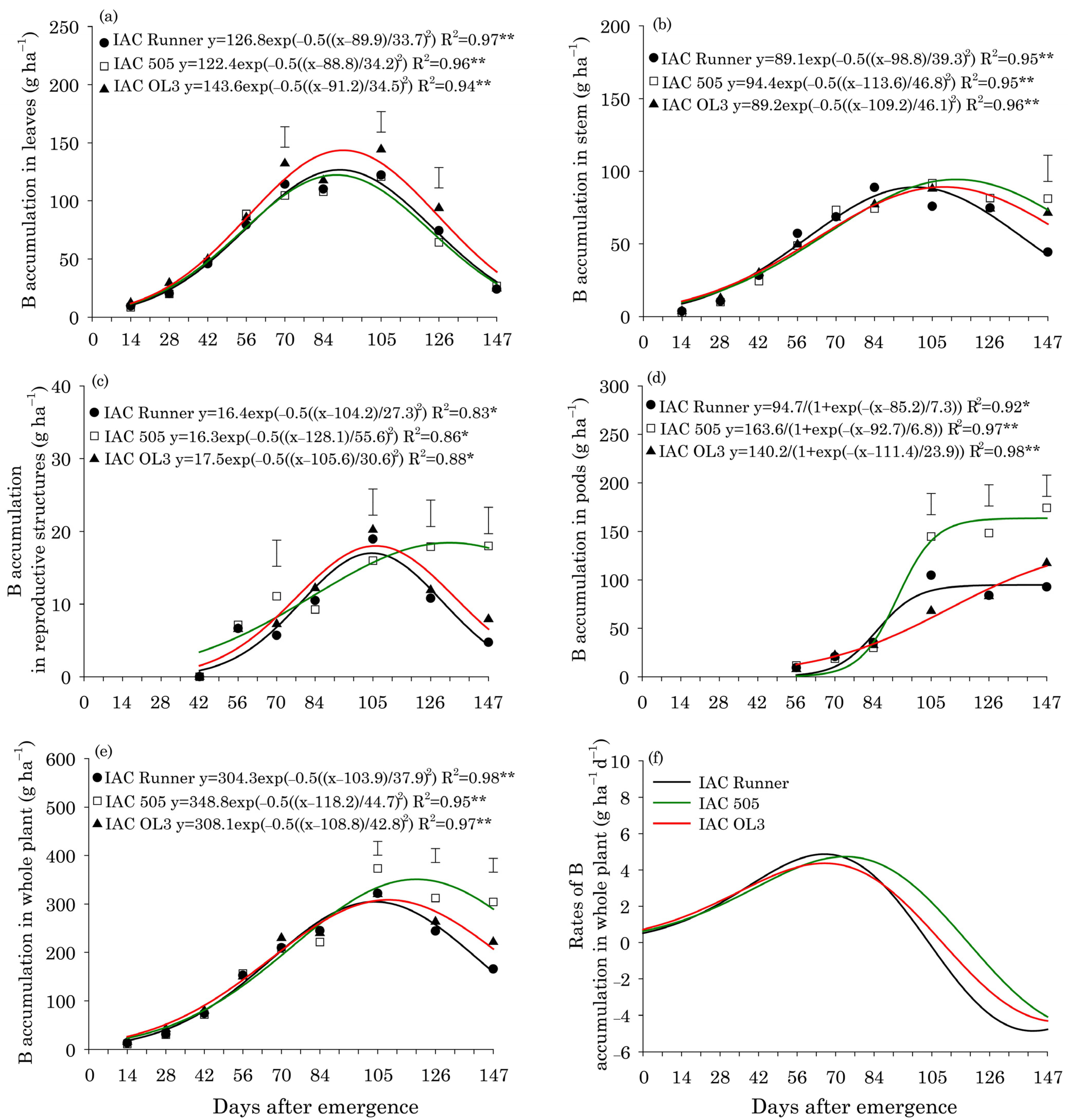
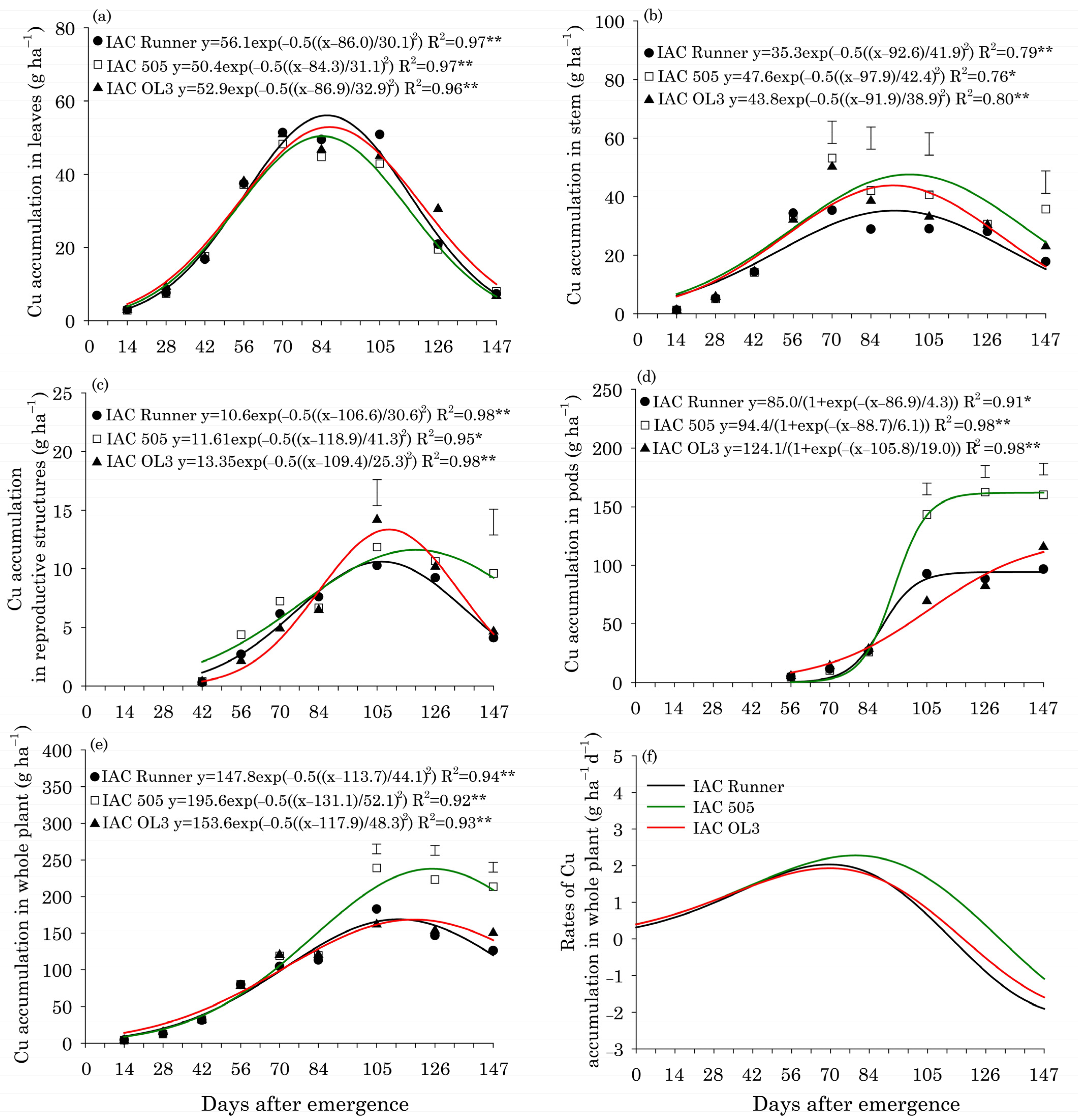
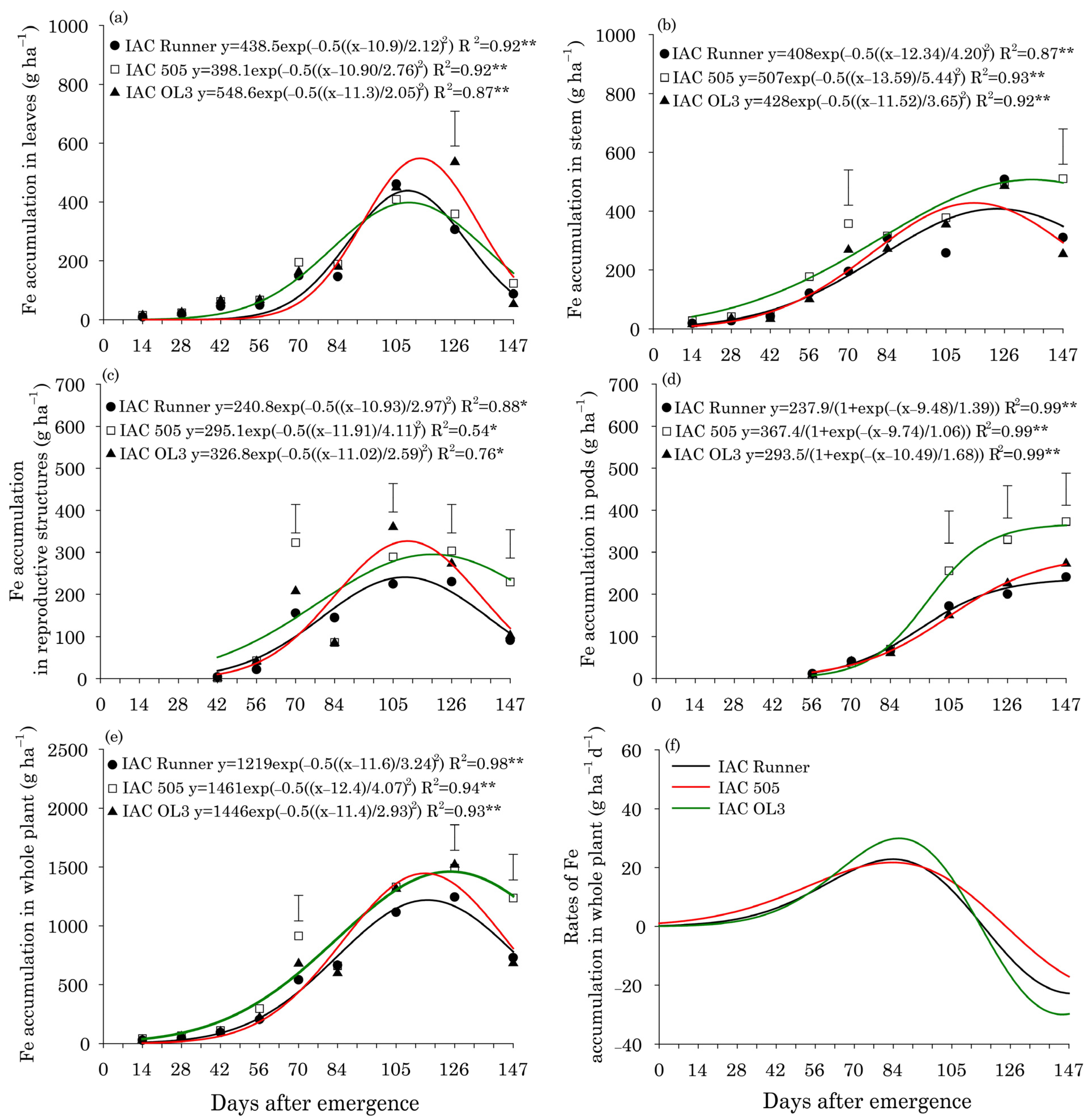
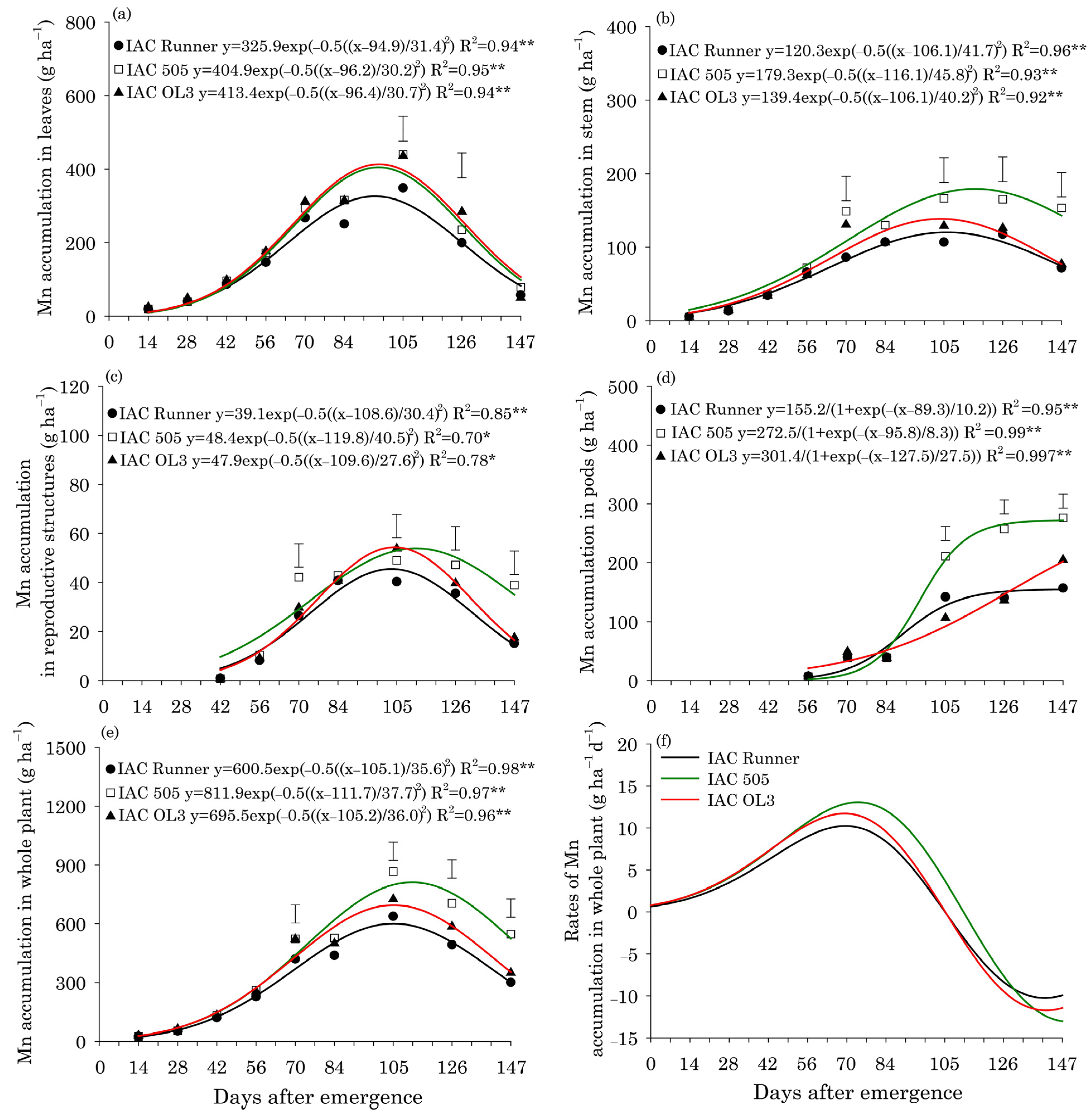
3. Results
Leaf Content and Micronutrient Uptake
4. Discussion
Concentrations and Uptake of Micronutrients
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sharma, C.P. Plant Micronutrients; CRC Press: Boca Raton, FL, USA, 2006; ISBN 0429079427. [Google Scholar]
- Khan, M.N.; Siddiqui, M.H.; Mohammad, F.; Naeem, M.; Khan, M.M.A. Calcium Chloride and Gibberellic Acid Protect Linseed (Linum usitatissimum L.) from NaCl Stress by Inducing Antioxidative Defence System and Osmoprotectant Accumulation. Acta Physiol. Plant. 2010, 32, 121. [Google Scholar] [CrossRef]
- Pawar, P.B.; Khadilkar, J.P.; Kulkarni, M.V.; Melo, J.S. An Approach to Enhance Nutritive Quality of Groundnut (Arachis hypogaea L.) Seed Oil through Endo Mycorrhizal Fertigation. Biocatal. Agric. Biotechnol. 2018, 14, 18–22. [Google Scholar] [CrossRef]
- Xie, M.; Wang, Z.; Xu, X.; Zheng, X.; Liu, H.; Shi, P. Quantitative Estimation of the Nutrient Uptake Requirements of Peanut. Agronomy 2020, 10, 119. [Google Scholar] [CrossRef] [Green Version]
- CONAB. Acompanhamento Da Safra Brasileira-Sétimo Levantamento Safra 2020/21; CONAB: Brasília, Brazil, 2021; Volume 8. [Google Scholar]
- Suassuna, T.; Suassuna, N.; Martins, K.; Matos, R.; Heuert, J.; Bertioli, D.; Leal-Bertioli, S.; Moretzsohn, M. Broadening the Variability for Peanut Breeding with a Wild Species-Derived Induced Allotetraploid. Agronomy 2020, 10, 1917. [Google Scholar] [CrossRef]
- Brown, N.; Branch, W.D.; Johnson, M.; Wallace, J. Genetic Diversity Assessment of Georgia Peanut Cultivars Developed during Ninety Years of Breeding. Plant Genome 2021, 14, e20141. [Google Scholar] [CrossRef]
- Godoy, I.J.; Santos, J.F.; Michelotto, M.D.; Moraes, A.R.A.; Bolonhezi, D.; de Freitas, R.S.; de Carvalho, C.R.L.; Finoto, E.L.; Martins, A.L.M. IAC OL 5-New High Oleic Runner Peanut Cultivar. Crop Breed. Appl. Biotechnol. 2017, 17, 289–292. [Google Scholar] [CrossRef] [Green Version]
- Crusciol, C.A.C.; Soratto, R.P. Nitrogen Supply for Cover Crops and Effects on Peanut Grown in Succession under a No-till System. Agron. J. 2009, 101, 41–46. [Google Scholar] [CrossRef]
- Mishra, A.K.; Shukla, S.K.; Yadav, D.V.; Awasthi, S.K. Iron, Manganese and Sulphur Uptake and Nutrients Availability in Sugarcane Based System in Subtropical India. Sugar Tech. 2014, 16, 300–310. [Google Scholar] [CrossRef]
- Souza, L.G.M.; Parente, T.d.L.; Lazarini, E.; Bossolani, J.W.; Caioni, S. Growth Regulator and Density Population in the Culture of Runner Peanut. Nucleus 2018, 15, 217–225. [Google Scholar] [CrossRef] [Green Version]
- Crusciol, C.A.C.; Ferrari Neto, J.; Mui, T.S.; Franzluebbers, A.J.; Da Costa, C.H.M.; Castro, G.S.A.; Ribeiro, L.C.; Costa, N.R. Rhizobial Inoculation and Molybdenum Fertilization in Peanut Crops Grown in a No Tillage System after 20 Years of Pasture. Rev. Bras. Cienc. Solo 2019, 43, e0170399. [Google Scholar] [CrossRef]
- Gericó, T.G.; Tavanti, R.F.R.; de Oliveira, S.C.; Lourenzani, A.E.B.S.; de Lima, J.P.; Ribeiro, R.P.; dos Santos, L.C.C.; dos Reis, A.R. Bradyrhizobium sp. Enhance Ureide Metabolism Increasing Peanuts Yield. Arch. Microbiol. 2020, 202, 645–656. [Google Scholar] [CrossRef] [PubMed]
- Moretti, L.G.; Crusciol, C.A.C.; Kuramae, E.E.; Bossolani, J.W.; Moreira, A.; Costa, N.R.; Alves, C.J.; Pascoaloto, I.M.; Rondina, A.B.L.; Hungria, M. Effects of Growth-Promoting Bacteria on Soybean Root Activity, Plant Development, and Yield. Agron. J. 2020, 112, 418–428. [Google Scholar] [CrossRef]
- Perrino, E.V.; Wagensommer, R.P. Crop Wild Relatives (Cwr) Priority in Italy: Distribution, Ecology, in Situ and Ex Situ Conservation and Expected Actions. Sustainability 2021, 13, 1682. [Google Scholar] [CrossRef]
- Perrino, E.V.; Valerio, F.; Jallali, S.; Trani, A.; Mezzapesa, G.N. Ecological and Biological Properties of Satureja Cuneifolia Ten. and Thymus Spinulosus Ten.: Two Wild Officinal Species of Conservation Concern in Apulia (Italy). A Preliminary Survey. Plants 2021, 10, 1952. [Google Scholar] [CrossRef]
- Zhao, C.X.; Jia, L.H.; Wang, Y.F.; Wang, M.L.; McGiffen, M.E. Effects of Different Soil Texture on Peanut Growth and Development. Commun. Soil Sci. Plant Anal. 2015, 46, 2249–2257. [Google Scholar] [CrossRef]
- Dhaliwal, S.S.; Naresh, R.K.; Mandal, A.; Singh, R.; Dhaliwal, M.K. Dynamics and Transformations of Micronutrients in Agricultural Soils as Influenced by Organic Matter Build-up: A Review. Environ. Sustain. Indic. 2019, 1–2, 100007. [Google Scholar] [CrossRef]
- Steiner, F.; de Queiroz, L.F.M.; Zuffo, A.M.; da Silva, K.C.; Lima, I.M.d.O. Peanut Response to Co-Inoculation of Bradyrhizobium spp. and Azospirillum brasilense and Molybdenum Application in Sandy Soil of the Brazilian Cerrado. Agron. J. 2021, 113, 623–632. [Google Scholar] [CrossRef]
- Shafaat, H.S.; Rüdiger, O.; Ogata, H.; Lubitz, W. [NiFe] Hydrogenases: A Common Active Site for Hydrogen Metabolism under Diverse Conditions. Biochim. Biophys. Acta Bioenerg. 2013, 1827, 986–1002. [Google Scholar] [CrossRef] [Green Version]
- Moretti, L.G.; Lazarini, E.; Bossolani, J.W.; Parente, T.L.; Caioni, S.; Araujo, R.S.; Hungria, M. Can Additional Inoculations Increase Soybean Nodulation and Grain Yield? Agron. J. 2018, 110, 715–721. [Google Scholar] [CrossRef]
- Moretti, L.G.; Crusciol, C.A.C.; Bossolani, J.W.; Calonego, J.C.; Moreira, A.; Garcia, A.; Momesso, L.; Kuramae, E.E.; Hungria, M. Beneficial Microbial Species and Metabolites Alleviate Soybean Oxidative Damage and Increase Grain Yield during Short Dry Spells. Eur. J. Agron. 2021, 127, 126293. [Google Scholar] [CrossRef]
- Moretti, L.G.; Crusciol, C.A.C.; Bossolani, J.W.; Momesso, L.; Garcia, A.; Kuramae, E.E.; Hungria, M. Bacterial Consortium and Microbial Metabolites Increase Grain Quality and Soybean Yield. J. Soil Sci. Plant Nutr. 2020, 20, 1923–1934. [Google Scholar] [CrossRef]
- Carrega, W.C.; Dos Santos, J.I.; Cesarin, A.E.; Gallardo, G.J.T.; Bacha, A.L.; De Godoy, I.J.; da Alves, P.L.C.A. Physiological responses of peanut genotypes to water deficit. Rev. Bras. Ciências Ambient. 2020, 54, 119–133. [Google Scholar] [CrossRef]
- Bolaños, L.; Brewin, N.J.; Bonilla, I. Effects of Boron on Rhizobium-Legume Cell-Surface Interactions and Nodule Development. Plant Physiol. 1996, 110, 1249–1256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reguera, M.; Espí, A.; Bolaños, L.; Bonilla, I.; Redondo-Nieto, M. Endoreduplication before Cell Differentiation Fails in Boron-Deficient Legume Nodules. Is Boron Involved in Signalling during Cell Cycle Regulation? New Phytol. 2009, 183, 8–12. [Google Scholar] [CrossRef]
- Garcia-Gonzalez, M.; Mateo, P.; Bonilla, I. Boron Protection for O2 Diffusion in Heterocysts of Anabaena sp. PCC 7119. Plant Physiol. 1988, 87, 785–789. [Google Scholar] [CrossRef] [Green Version]
- Pandey, S.N. Role of Micronutrients in Biochemical Responses of Crops under Abiotic Stresses; Springer: Berlin/Heidelberg, Germany, 2020. [Google Scholar]
- Tripathi, D.K.; Singh, S.; Singh, S.; Mishra, S.; Chauhan, D.K.; Dubey, N.K. Micronutrients and Their Diverse Role in Agricultural Crops: Advances and Future Prospective. Acta Physiol. Plant 2015, 37, 139. [Google Scholar] [CrossRef]
- Mantovani, J.P.M.; Calonego, J.C.; Foloni, J.S.S. Adubação Foliar de Boro Em Diferentes Estádios Fenológicos Da Cultura Do Amendoim. Rev. Ceres 2013, 60, 270–278. [Google Scholar] [CrossRef] [Green Version]
- Powell, N.L.; Swann, C.W.; Martens, D.C. Foliar Fertilization of Virginia-Type Peanut with MnEDTA—Crop Grade, Pod Yield, and Value1. Peanut Sci. 1996, 23, 98–103. [Google Scholar] [CrossRef]
- Aboyeji, C.; Dunsin, O.; Adekiya, A.O.; Chinedum, C.; Suleiman, K.O.; Okunlola, F.O.; Aremu, C.O.; Owolabi, I.O.; Olofintoye, T.A.J. Zinc Sulphate and Boron-Based Foliar Fertilizer Effect on Growth, Yield, Minerals, and Heavy Metal Composition of Groundnut (Arachis hypogaea L.) Grown on an Alfisol. Int. J. Agron. 2019, 2019, 5347870. [Google Scholar] [CrossRef] [Green Version]
- Irmak, S.; Cil, A.N.; Yucel, H.; Kaya, Z. Effects of Zinc Application on Yield and Some Yield Components in Peanut (Arachis hypogaea) in the Easthern Mediterranean Region. J. Agric. Sci. (CAS) 2016, 22, 109–116. [Google Scholar] [CrossRef]
- Crusciol, C.A.C.; Portugal, J.R.; Bossolani, J.W.; Moretti, L.G.; Fernandes, A.M.; Garcia, J.L.N.; Garcia, G.L.d.B.; Pilon, C.; Cantarella, H. Dynamics of Macronutrient Uptake and Removal by Modern Peanut Cultivars. Plants 2021, 10, 2167. [Google Scholar] [CrossRef] [PubMed]
- Santos, H.G.; Jacomine, P.T.; Dos Anjos, L.H.C.; de Oliveira, V.A.; Lumbreras, J.F.; Coelho, M.R.; Almeida, J.A.; Araujo Filho, J.C.; Oliveira, J.B.; Cunha, T.J.F. Brazilian Soil Classification System.-Portal Embrapa, 5th ed.; Embrapa Solos: Brasília, Brazil, 2018; ISBN 978-85-7035-817-2. [Google Scholar]
- Soil Survey Staff. Keys to Soil Taxonomy, 12th ed.; USDA-Natural Resources Conservation Service: Washington, DC, USA, 2014. [Google Scholar]
- Unicamp Center of Meteorological and Climatic Research Applied to Agriculture. Botucatu: Municipalities Climate of São Paulo State. Available online: www.cpa.unicamp.br/outras-informacoes/clima_muni_086.html (accessed on 20 June 2020).
- Malavolta, E.; Vitti, G.C.; Oliveira, S. Evaluation of the Nutritional State of Plants: Principles and Applications, 2nd ed.; POTAFOS: Piracicaba, Brazil, 1989. [Google Scholar]
- Ambrosano, E.J.; Tanaka, R.T.; Mascarenhas, H.A.A.; van Raij, B.; Quaggio, J.A.; Cantarella, H. Leguminosas e Oleaginosas. In Recomendações de Adubação e Calagem Para o Estado de São Paulo; van Raij, B., Cantarella, H., Quaggio, J.A., Furlani, A.M.C., Eds.; Instituto Agronômico: Campinas, Brazil, 1997; pp. 187–203. [Google Scholar]
- van Raij, B.; Cantarella, H.; Quaggio, J.A.; Furlani, A.M.C. Recomendações de Adubação e Calagem Para o Estado de São Paulo, 2nd ed.; Fundação IAC Campinas: Campinas, Brazil, 1997. [Google Scholar]
- Feitosa, C.T.; Nogueira, S.S.S.; Gerin, M.A.N.; Rodrigues Filho, F.S.O. Peanut Growth and Nutrient Evaluation. Sci. Agric. 1993, 50, 427–437. [Google Scholar] [CrossRef] [Green Version]
- Fernando, D.R.; Marshall, A.T.; Forster, P.I.; Hoebee, S.E.; Siegele, R. Multiple Metal Accumulation within a Manganese-Specific Genus. Am. J. Bot. 2013, 100, 690–700. [Google Scholar] [CrossRef] [PubMed]
- Fernando, D.R.; Lynch, J.P. Manganese Phytotoxicity: New Light on an Old Problem. Ann. Bot. 2015, 116, 313–319. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, S.B.; Husted, S. The Biochemical Properties of Manganese in Plants. Plants 2019, 8, 381. [Google Scholar] [CrossRef] [Green Version]
- Bottega, E.L.; de Queiroz, D.M.; Pinto, F.d.A.d.C.; de Souza, C.M.A. Spatial Variability of Soil Attributes in No a No-Tillage System with Crop Rotation in the Brazilian Savannah. Rev. Cienc. Agron. 2013, 44, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Dučić, T.; Polle, A. Transport and Detoxification of Manganese and Copper in Plants. Braz. J. Plant Physiol. 2005, 17, 103–112. [Google Scholar] [CrossRef] [Green Version]
- Alejandro, S.; Höller, S.; Meier, B.; Peiter, E. Manganese in Plants: From Acquisition to Subcellular Allocation. Front. Plant Sci. 2020, 11, 300. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
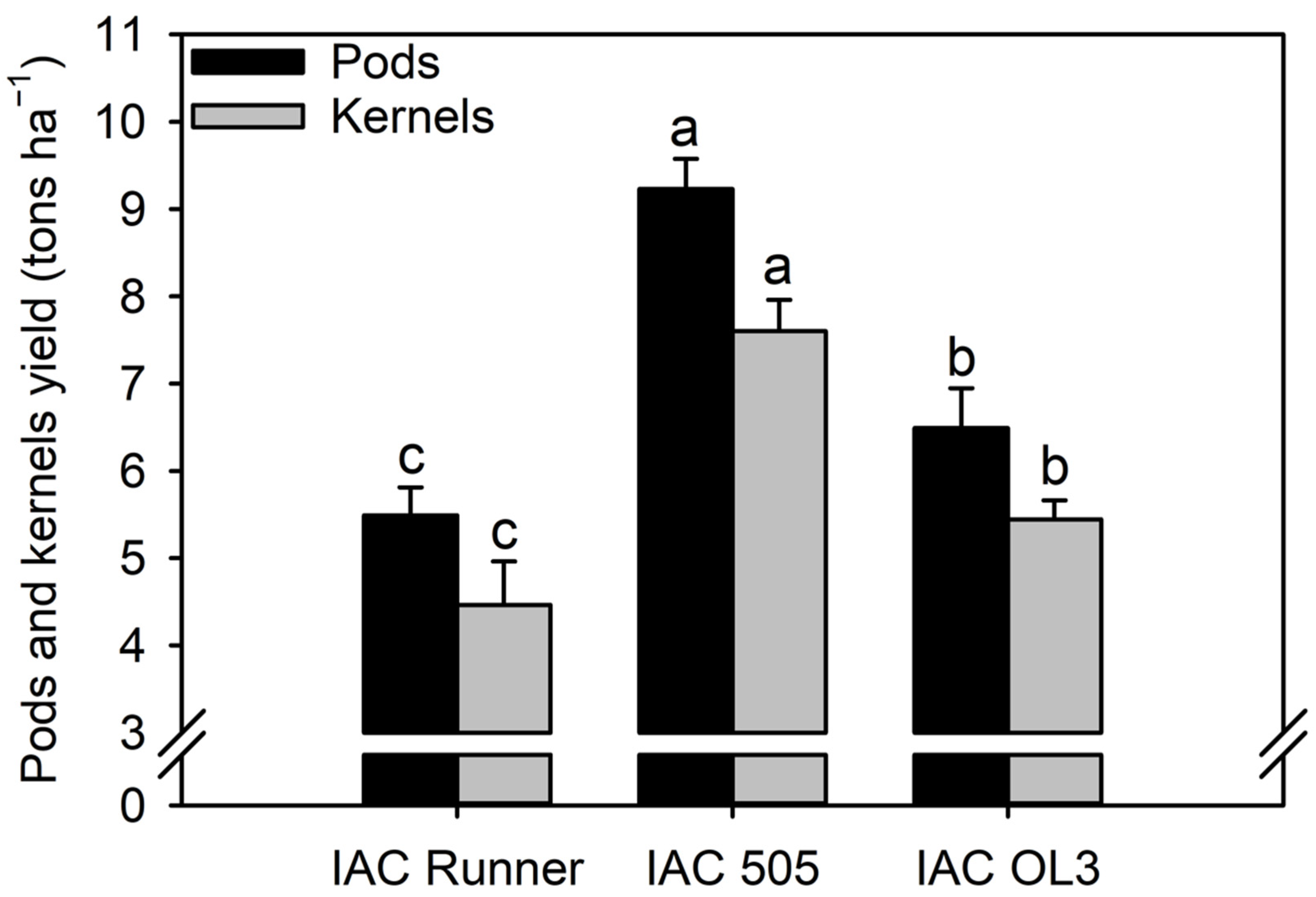{kind=link}
| Cultivars | B | Cu | Fe | Mn | Zn |
|---|---|---|---|---|---|
| mg kg−1 | |||||
| IAC Runner | 52 ± 4.3 a | 20 ± 1.3 a | 193 ± 8.1 b | 95 ± 5.9 b | 45 ± 1.9 a |
| IAC 505 | 60 ± 4.2 a | 19 ± 1.1 ab | 270 ± 7.2 a | 109 ± 7.1 a | 47 ± 1.6 a |
| IAC OL3 | 54 ± 3.8 a | 17 ± 0.9 b | 178 ± 6.9 b | 100 ± 5.4 ab | 47 ± 1.5 a |
| CV% | 12.3 | 6.5 | 7.9 | 5.9 | 13.4 |
| Cultivar | B | Cu | Fe | Mn | Zn |
|---|---|---|---|---|---|
| Micronutrients taken up per Mg of pods produced (g Mg−1) (1) | |||||
| IAC Runner | 55.4 ± 1.8 a | 30.4 ± 0.8 a | 224.1 ± 44.9 a | 105.6 ± 10.2 a | 56.5 ± 4.5 a |
| IAC 505 | 37.8 ± 2.1 c | 25.9 ± 0.5 b | 158.9 ± 38.2 a | 87.8 ± 9.3 a | 48.4 ± 5.2 a |
| IAC OL3 | 47.5 ± 1.7 b | 25.9 ± 0.4 b | 222.6 ± 42.4 a | 104.7 ± 9.8 a | 51.6 ± 3.8 a |
| Micronutrients taken up per Mg of kernels produced (g Mg−1) (1) | |||||
| IAC Runner | 68.1 ± 4.8 a | 37.4 ± 2.3 a | 275.8 ± 40.1 a | 129.9 ± 10.3 a | 69.5 ± 5.3 a |
| IAC 505 | 45.9 ± 5.7 b | 31.4 ± 3.8 ab | 193.0 ± 49.2 a | 106.5 ± 13.1 a | 58.8 ± 6.2 a |
| IAC OL3 | 56.6 ± 5.4 b | 30.9 ± 2.1 b | 265.3 ± 35. 3 a | 124.8 ± 9.8 a | 61.6 ± 4.4 a |
| Micronutrient content in pods (mg kg−1) | |||||
| IAC Runner | 16.8 ± 0.6 b | 17.6 ± 0.5 a | 43.7 ± 0.9 a | 28.7 ± 0.6 b | 31.5 ± 1.2 a |
| IAC 505 | 18.9 ± 0.6 a | 17.3 ± 0.4 a | 40.1 ± 0.5 b | 30.0 ± 0.8 ab | 30.5 ± 0.8 a |
| IAC OL3 | 18.1 ± 0.8 ab | 17.8 ± 0.3 a | 41.9 ± 1.0 a | 31.5 ± 0.7 a | 29.2 ± 1.0 a |
| Micronutrient content in kernels (mg kg−1) | |||||
| IAC Runner | 16.4 ± 1.5 a | 15.6 ± 0.9 a | 4.6 ± 1.1 a | 17.6 ± 0.5 b | 35.4 ± 2.3 a |
| IAC 505 | 19.1 ± 1.3 a | 15.2 ± 1.1 a | 4.0 ± 0.8 a | 19.7 ± 0.7 a | 33.9 ± 1.9 a |
| IAC OL3 | 17.4 ± 0.9 a | 15.4 ± 1.2 a | 3.4 ± 1.2 a | 18.0 ± 0.6 b | 31.6 ± 2.0 a |
| Micronutrient removal by pods per area (g ha−1) | |||||
| IAC Runner | 92.7 ± 4.5 c | 96.7 ± 10.4 b | 240.9 ± 17.8 b | 156.9 ± 15.4 c | 173.5 ± 11.5 b |
| IAC 505 | 174.2 ± 5.1 a | 160.1 ± 9.2 a | 373.2 ± 15.2 a | 276.5 ± 19.1 a | 281.5 ± 17.2 a |
| IAC OL3 | 117.3 ± 5.2 b | 115.7 ±10.1 b | 272.7 ± 12.1 ab | 204.9 ± 10.1 b | 189.6 ± 12.3 b |
| Micronutrient removal by kernels per area (g ha−1) | |||||
| IAC Runner | 73.4 ± 6.9 c | 69.3 ± 5.4 c | 22.4 ± 6.4 a | 77.6 ± 7.8 c | 158.2 ± 7.4 b |
| IAC 505 | 144.5 ± 8.3 a | 115.2 ± 9.8 a | 30.1 ± 6.9 a | 149.4 ± 9.3 a | 257.8 ± 8.5 a |
| IAC OL3 | 94.8 ± 7.3 b | 83.5 ± 6.1 b | 18.5 ± 7.3 a | 97.8 ± 6.4 b | 171.4 ± 9.9 b |
| Micronutrient removal per Mg of pods produced (g Mg−1) | |||||
| IAC Runner | 16.8 ± 2.4 a | 17.6 ± 0.5 a | 43.7 ± 1.4 a | 28.7 ± 0.8 b | 31.5 ± 0.9 a |
| IAC 505 | 18.9 ± 2.1 a | 17.3 ± 0.7 a | 40.1 ± 0.5 b | 29.9 ± 0.4 ab | 30.5 ± 1.0 a |
| IAC OL3 | 18.1 ± 1.8 a | 17.8 ± 0.3 a | 41.9 ± 0.9 a | 31.5 ± 0.5 a | 29.2 ± 1.4 a |
| Micronutrient removal per Mg of kernels produced (g Mg−1) | |||||
| IAC Runner | 16.4 ± 0.5 c | 15.6 ± 0.5 a | 4.6 ± 0.6 a | 17.6 ± 0.5 b | 35.4 ± 2.1 a |
| IAC 505 | 19.1 ± 0.4 a | 15.2 ± 0.6 a | 4.0 ± 0.4 a | 19.7 ± 0.7 a | 33.9 ± 1.5 a |
| IAC OL3 | 17.4 ± 0.4 b | 15.4 ± 0.4 a | 3.4 ± 0.6 a | 18.0 ± 0.6 b | 31.6 ± 1.7 a |
| Relative micronutrient removal by pods (%) (2) | |||||
| IAC Runner | 30.5 | 57.9 | 19.6 | 27.1 | 56.0 |
| IAC 505 | 49.9 | 67.0 | 25.4 | 34.1 | 63.0 |
| IAC OL3 | 38.1 | 68.9 | 18.9 | 30.2 | 56.6 |
| Relative micronutrient removal by kernels (%) (2) | |||||
| IAC Runner | 24.2 | 41.5 | 1.8 | 13.4 | 51.0 |
| IAC 505 | 41.4 | 48.2 | 2.1 | 18.4 | 57.7 |
| IAC OL3 | 30.7 | 49.7 | 1.3 | 14.4 | 51.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Crusciol, C.A.C.; Portugal, J.R.; Bossolani, J.W.; Moretti, L.G.; Fernandes, A.M.; Moreira, A.; Garcia, J.L.N.; Garcia, G.L.d.B.; Pilon, C.; Cantarella, H. Dynamics of Micronutrient Uptake and Removal by Three Modern Runner Peanut Cultivars. Crops 2023, 3, 101-115. https://doi.org/10.3390/crops3020010
Crusciol CAC, Portugal JR, Bossolani JW, Moretti LG, Fernandes AM, Moreira A, Garcia JLN, Garcia GLdB, Pilon C, Cantarella H. Dynamics of Micronutrient Uptake and Removal by Three Modern Runner Peanut Cultivars. Crops. 2023; 3(2):101-115. https://doi.org/10.3390/crops3020010
Chicago/Turabian StyleCrusciol, Carlos Alexandre Costa, José Roberto Portugal, João William Bossolani, Luiz Gustavo Moretti, Adalton Mazetti Fernandes, Adônis Moreira, Jader Luis Nantes Garcia, Gleize Leviski de Brito Garcia, Cristiane Pilon, and Heitor Cantarella. 2023. "Dynamics of Micronutrient Uptake and Removal by Three Modern Runner Peanut Cultivars" Crops 3, no. 2: 101-115. https://doi.org/10.3390/crops3020010