

Characterization of Secondary Metabolites Responsible for the Resistance of Local Tomato Accessions to Whitefly (Bemisia tabaci, Gennadius 1889) Hemiptera in Tanzania

Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Site, Plant Materials and Experimental Design
2.2. Identification of Metabolites Responsible for Whitefly Resistance
3. Statistical Analysis
4. Results
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dube, J.; Ddamulira, G.; Maphosa, M. Tomato breeding in sub-Saharan Africa-Challenges and opportunities: A review. Afr. Crop Sci. J. 2020, 28, 131–140. [Google Scholar]
- Ochilo, W.N.; Nyamasyo, G.N.; Kilalo, D.; Otieno, W.; Otipa, M.; Chege, F.; Karanja, T.; Lingeera, E.K. Practice, Ecological limits and management practices of major arthropod pests of tomato in Kenya. J. Agric. Sci. Pract. 2019, 4, 29–42. [Google Scholar] [CrossRef] [Green Version]
- Ali, A.; Rakha, M.; Shaheen, F.A.; Srinivasan, R. Resistance of certain wild tomato (Solanum spp.) accessions to Helicoverpa armigera (Hübner)(Lepidoptera: Noctuidae) based on choice and no-choice bioassays. Fla. Entomol. 2019, 102, 544–548. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; Sachan, S.; Kumar, S.; Kumar, P. Efficacy of some novel insecticides against whitefly (Bemisia tabaci Gennadius) in Brinjal. J. Entomol. Zool. Stud. 2017, 5, 424–427. [Google Scholar]
- Alam, M.; Islam, M.; Haque, M.; Humayun, R.; Khalequzzaman, K. Bio-rational management of whitefly (Bemisia tabaci) for suppressing tomato yellow leaf curl virus. J. Agril. Res. 2016, 41, 583–597. [Google Scholar] [CrossRef] [Green Version]
- Mutayoba, V.; Ngaruko, D. Assessing tomato farming and marketing among smallholders in high potential agricultural areas of Tanzania. Int. J. Econ. Commer. Manag. 2018, 6, 577–590. [Google Scholar]
- Bala, K.; Sood, A.; Pathania, V.S.; Thakur, S. Effect of plant nutrition in insect pest management: A review. J. Pharmacogn. Phytochem. 2018, 7, 2737–2742. [Google Scholar]
- Vosman, B.; van’t Westende, W.P.; Henken, B.; van Eekelen, H.D.; de Vos, R.C.; Voorrips, R.E. Broad spectrum insect resistance and metabolites in close relatives of the cultivated tomato. Euphytica 2018, 214, 46. [Google Scholar] [CrossRef] [Green Version]
- Sri, N.R.; Jha, S. Whitefly biology and morphometry on tomato plants. J. Entomol. Zool. Stud. 2018, 6, 2079–2081. [Google Scholar]
- Perring, T.M.; Stansly, P.A.; Liu, T.; Smith, H.A.; Andreason, S.A. Whiteflies: Biology, ecology, and management. In Sustainable Management of Arthropod Pests of Tomato; Elsevier: Amsterdam, The Netherlands, 2018; pp. 73–110. [Google Scholar]
- Chand, R.; Jokhan, A.; Kelera, R. Spiralling whitefly and its management practices in the South Pacific. A review. J. Adv. Hortic. Sci. 2019, 33, 123–131. [Google Scholar]
- Wan, F.-H.; Yang, N.-W. Invasion and management of agricultural alien insects in China. Annu. Rev. Entomol. 2016, 61, 77–98. [Google Scholar] [CrossRef] [PubMed]
- Stephenson, R.C.; Coker, C.E.; Posadas, B.C.; Bachman, G.R.; Harkess, R.L.; Adamczyk, J.J.; Knight, P.R.J.H. Economic Effect of Insect Pest Management Strategies on Small-scale Tomato Production in Mississippi. HortTechnology 2020, 30, 64–75. [Google Scholar] [CrossRef] [Green Version]
- Cathrin, P.B.; Ghanim, M. Recent advances on interactions between the whitefly Bemisia tabaci and begomoviruses, with emphasis on Tomato yellow leaf curl virus. In Plant Virus–Host Interaction—Molecular Approaches and Viral Evolution; Gaur, R.K., Paul Khurana, S.M., Sharma, P., Hohn, T., Eds.; Academic Press: Cambridge, MA, USA, 2014; pp. 79–103. [Google Scholar]
- Jamiołkowska, A. Natural compounds as elicitors of plant resistance against diseases and new biocontrol strategies. Agronomy 2020, 10, 173. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Zhang, W.; Che, W.; Qu, C.; Li, F.; Desneux, N.; Luo, C. Lethal and sublethal effects of cyantraniliprole, a new anthranilic diamide insecticide, on Bemisia tabaci (Hemiptera: Aleyrodidae) MED. Crop Prot. 2017, 91, 108–113. [Google Scholar] [CrossRef]
- Laizer, H.C.; Chacha, M.N.; Ndakidemi, P.A. Farmers’ Knowledge, Perceptions and Practices in Managing Weeds and Insect Pests of Common Bean in Northern Tanzania. Sustainability 2019, 11, 4076. [Google Scholar] [CrossRef] [Green Version]
- Raja, N.; Masresha, G. Plant based biopesticides: Safer alternative for organic food production. J. Fertil. Pestic. 2015, 6, e128. [Google Scholar] [CrossRef]
- Amin, M.; Chakma, A.; Alam, M.; Hossain, M.; Ge, F. Screening of tomato varieties against tomato fruit borer and associated plant characters. J. Agric. 2016, 14, 150–161. [Google Scholar] [CrossRef] [Green Version]
- Firdaus, S.; van Heusden, A.W.; Hidayati, N.; Supena, E.D.J.; Mumm, R.; de Vos, R.C.; Visser, R.G.; Vosman, B. Identification and QTL Mapping of Whitefly Resistance Components in SOLANUM Galapagense; Wageningen University: Wageningen, The Netherlands, 2013. [Google Scholar]
- Lima, I.P.; Resende, J.T.; Oliveira, J.R.; Faria, M.V.; Dias, D.M.; Resende, N.C. Selection of tomato genotypes for processing with high zingiberene content, resistant to pests. Hortic. Bras. 2016, 34, 387–391. [Google Scholar] [CrossRef]
- Bleeker, P.M.; Mirabella, R.; Diergaarde, P.J.; VanDoorn, A.; Tissier, A.; Kant, M.R.; Prins, M.; de Vos, M.; Haring, M.A.; Schuurink, R.C. Improved herbivore resistance in cultivated tomato with the sesquiterpene biosynthetic pathway from a wild relative. Plant Biol. 2012, 109, 20124–20129. [Google Scholar] [CrossRef] [Green Version]
- Azwanida, N. A review on the extraction methods use in medicinal plants, principle, strength and limitation. Med. Aromat. Plants 2015, 4, 196. [Google Scholar]
- Yactayo-Chang, J.P.; Tang, H.V.; Mendoza, J.; Christensen, S.A.; Block, A.K.J.A. Plant defense chemicals against insect pests. Agronomy 2020, 10, 1156. [Google Scholar] [CrossRef]
- Konno, K. Plant latex and other exudates as plant defense systems: Roles of various defense chemicals and proteins contained therein. Phytochemistry 2011, 72, 1510–1530. [Google Scholar] [CrossRef] [PubMed]
- D’Addabbo, T.; Laquale, S.; Lovelli, S.; Candido, V.; Avato, P. Biocide plants as a sustainable tool for the control of pests and pathogens in vegetable cropping systems. Ital. J. Agron. 2014, 9, 137–145. [Google Scholar] [CrossRef]
- Kant, M.; Jonckheere, W.; Knegt, B.; Lemos, F.; Liu, J.; Schimmel, B.; Villarroel, C.; Ataide, L.; Dermauw, W.; Glas, J.J.A.o.b. Mechanisms and ecological consequences of plant defence induction and suppression in herbivore communities. Ann. Bot. 2015, 115, 1015–1051. [Google Scholar] [CrossRef] [PubMed]
- Bhonwong, A.; Stout, M.J.; Attajarusit, J.; Tantasawat, P. Defensive role of tomato polyphenol oxidases against cotton bollworm (Helicoverpa armigera) and beet armyworm (Spodoptera exigua). J. Chem. Ecol. 2009, 35, 28–38. [Google Scholar] [CrossRef]
- Caspi, R.; Altman, T.; Billington, R.; Dreher, K.; Foerster, H.; Fulcher, C.A.; Holland, T.A.; Keseler, I.M.; Kothari, A.; Kubo, A. The MetaCyc database of metabolic pathways and enzymes and the BioCyc collection of Pathway/Genome Databases. J. Nucleic Acids Res. 2014, 42, D459–D471. [Google Scholar] [CrossRef] [Green Version]
- Erb, M.; Kliebenstein, D.J. Plant secondary metabolites as defenses, regulators, and primary metabolites: The blurred functional trichotomy. Plant Physiol. 2020, 184, 39–52. [Google Scholar] [CrossRef]
- War, A.R.; Buhroo, A.A.; Hussain, B.; Ahmad, T.; Nair, R.M.; Sharma, H.C. Plant defense and insect adaptation with reference to secondary metabolites. In Co-Evolution of Secondary Metabolites, Reference Series in Phytochemistry; Mérillon, J.M., Ramawat, K., Eds.; Springer: Cham, Switzerland, 2020; pp. 795–822. [Google Scholar]
- Cheng, A.X.; Lou, Y.G.; Mao, Y.B.; Lu, S.; Wang, L.J.; Chen, X.Y. Plant terpenoids: Biosynthesis and ecological functions. J. Integr. Plant Biol. 2007, 49, 179–186. [Google Scholar] [CrossRef]
- Thirumurugan, D.; Cholarajan, A.; Raja, S.; Vijayakumar, R. An Introductory Chapter: Secondary Metabolites; IntechOpen: London, UK, 2018; pp. 1–21. [Google Scholar]
- Falara, V.; Alba, J.M.; Kant, M.R.; Schuurink, R.C.; Pichersky, E. Geranyllinalool synthases in solanaceae and other angiosperms constitute an ancient branch of diterpene synthases involved in the synthesis of defensive compounds. Plant Physiol. 2014, 166, 428–441. [Google Scholar] [CrossRef] [Green Version]
- Mumm, R.; Posthumus, M.A.; Dicke, M. Significance of terpenoids in induced indirect plant defence against herbivorous arthropods. Plant Cell Environ. 2008, 31, 575–585. [Google Scholar] [CrossRef]
- Mbata, G.N.; Payton, M.E. Effect of monoterpenoids on oviposition and mortality of Callosobruchus maculatus (F.) (Coleoptera: Bruchidae) under hermetic conditions. J. Stored Prod. Res. 2013, 53, 43–47. [Google Scholar] [CrossRef]
- Divekar, P.A.; Narayana, S.; Divekar, B.A.; Kumar, R.; Gadratagi, B.G.; Ray, A.; Singh, A.K.; Rani, V.; Singh, V.; Singh, A.K. Plant secondary metabolites as defense tools against herbivores for sustainable crop protection. Int. J. Mol. Sci. 2022, 23, 2690. [Google Scholar] [CrossRef] [PubMed]
- Erb, M.; Meldau, S.; Howe, G.A. Role of phytohormones in insect-specific plant reactions. Trends Plant Sci. 2012, 17, 250–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonaventure, G. Plants recognize herbivorous insects by complex signalling networks. J. Annu. Plant Rev. 2014, 47, 1–36. [Google Scholar]
- González-Klenner, F.J.; Albornoz, M.V.; Ávila-Sákar, G.; Verdugo, J.A. Tomato Defense against Whiteflies under Drought Stress: Non-Additive Effects and Cultivar-Specific Responses. Plants 2022, 11, 1049. [Google Scholar] [CrossRef] [PubMed]
- Glas, J.J.; Schimmel, B.C.; Alba, J.M.; Escobar-Bravo, R.; Schuurink, R.C.; Kant, M.R. Plant glandular trichomes as targets for breeding or engineering of resistance to herbivores. Int. J. Mol. Sci. 2012, 13, 17077–17103. [Google Scholar] [CrossRef] [Green Version]
- Boncan, D.A.T.; Tsang, S.S.; Li, C.; Lee, I.H.; Lam, H.-M.; Chan, T.-F.; Hui, J.H. Terpenes and terpenoids in plants: Interactions with environment and insects. Int. J. Mol. Sci. 2020, 21, 7382. [Google Scholar] [CrossRef]
- Ramírez-Gómez, X.S.; Jiménez-García, S.N.; Campos, V.B.; Campos, M.L.G. Plant metabolites in plant defense against pathogens. In Plant Diseases-Current Threats and Management Trends; Topolovec-Pintarić, S., Ed.; IntechOpen: London, UK, 2019; pp. 49–68. [Google Scholar]
- Züst, T.; Agrawal, A.A. Mechanisms and evolution of plant resistance to aphids. Nat. Plants 2016, 2, 15206. [Google Scholar] [CrossRef]
- Lattanzio, V. Phenolic Compounds: Introduction 50. Nat. Prod. 2013, 1543–1580. [Google Scholar] [CrossRef]
- Schulz, E.; Tohge, T.; Zuther, E.; Fernie, A.; Hincha, D. Flavonoids are determinants of freezing tolerance and cold acclimation in Arabidopsis thaliana. Sci. Rep. 2016, 6, 34027. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Season One (July–October 2021) | Season Two (January–April 2022) | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Assessment from 30 Days Post-Inoculation | Assessment from 60 Days Post-Inoculation | Assessment from 30 Days Post-Inoculation | Assessment from 60 Days Post-Inoculation | |||||||||||
| Accession | AW/L | E/N/L | HD/L | AW/L | E/N/L | HD/L | Y/P | AW/L | E/N/L | HD/L | AW/L | E/N/L | HD/L | Y/P (kg) |
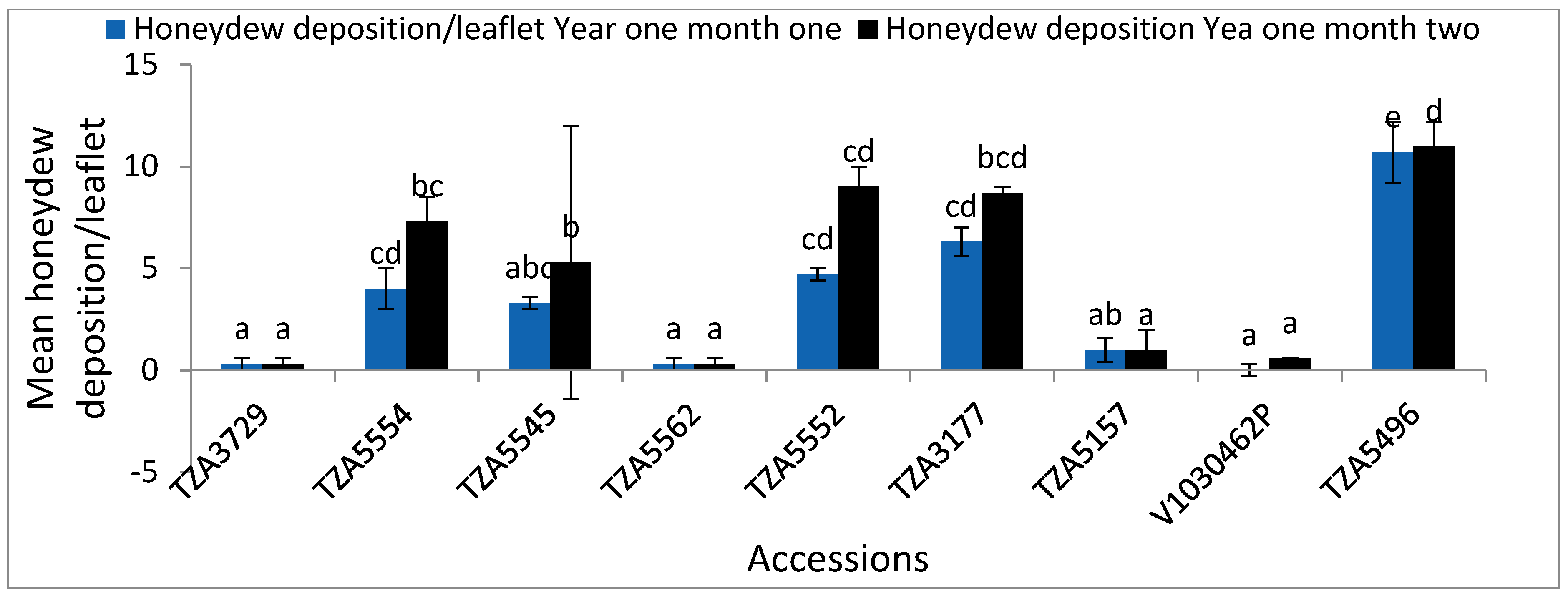
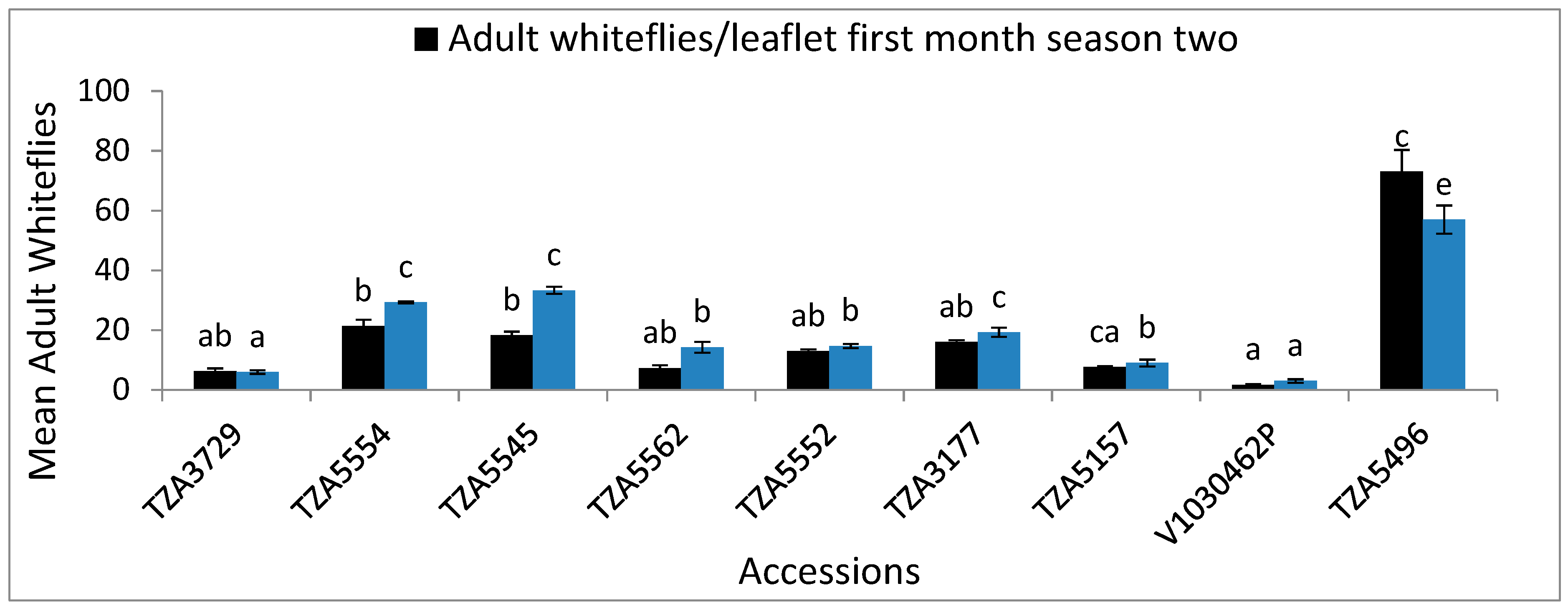
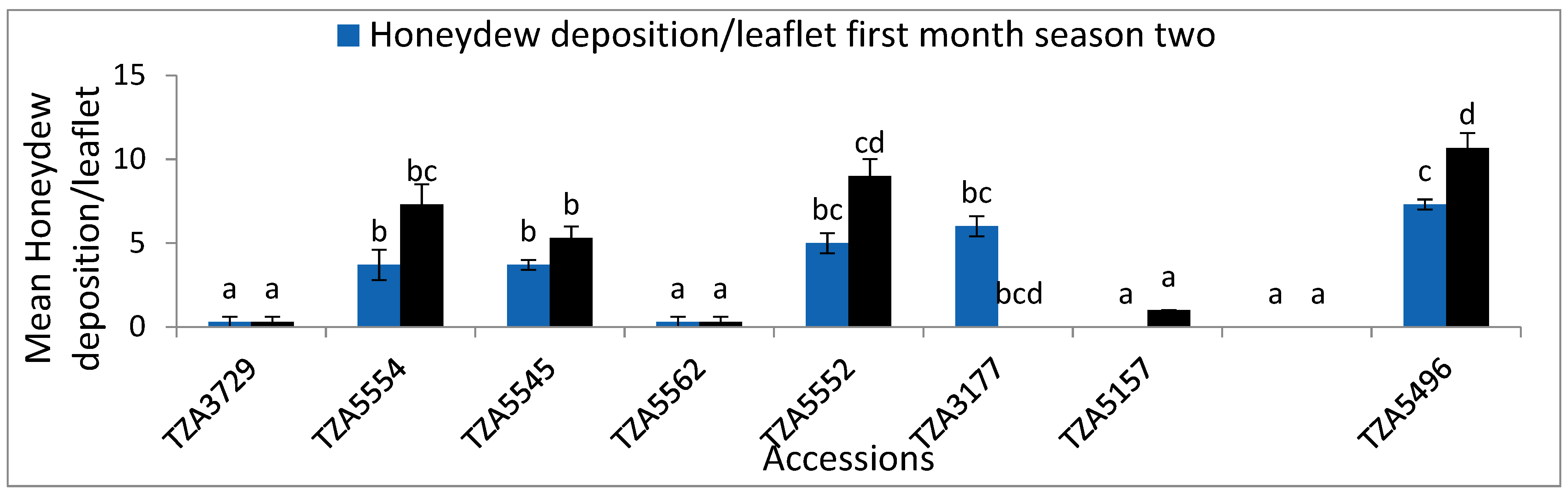
| TZA3729 | 5.3 ± 0.9 a | 2 ± 0.6 a | 0.3 ± 0.3 a | 8.7 ± 2 a | 4.3 ± 0.7 a | 0.3 ± 0.3 a | 1.4 ± 0.1 d | 6.3 ± 0.9 ab | 2 ± 0.58 ab | 0.3 ± 0.3 a | 6. ± 0.6 a | 4 ± 0.6 ab | 0.3 ± 0.3 a | 1.1 ± 0 d |
| TZA5554 | 20 ± 2.5 dcd | 14.7 ± 1.2 bc | 4 ± 1 bcd | 27.7 ± 0.9 c | 16.3 ± 2.6 cde | 7.3 ± 1.2 bc | 0.9 ± 0.0 bc | 21.3 ± 2.2 b | 14.7 ± 1.2 c | 3.7 ± 0.9 b | 29.3 ± 0.3 d | 17.7 ± 3 d | 7.3 ± 1.2 bc | 0.9 ± 0.0 bcd |
| TZA5545 | 24.7 ± 1.9 d | 12.3 ± 3.2 bc | 3.3 ± 0.3 abc | 31.3 ± 1.2 c | 11 ± 2.1 abcd | 5.3 ± 6.7 b | 0.9 ± 0.0 bc | 18.3 ± 1.2 b | 12.3 ± 3.3 c | 3.7 ± 0.3 b | 33.3 ± 1.2 cd | 16.7 ± 2.3 d | 5.3 ± 0.7 b | 0.9 ± 0 bcd |
| TZA5562 | 9 ± 0.6 ab | 9 ± 0.6 ab | 0.3 ± 0.3 a | 15 ± 1.2 ab | 15.3 ± 0.7 bcde | 0.3 ± 0.3 a | 1.2 ± 0.08 d | 7.3 ± 0.9 ab | 9 ± 0.6 abc | 0.3 ± 0.3 a | 14.3 ± 1.8 b | 15.3 ± 0.7 bc | 0.3 ± 0.3 a | 1 ± 0.1 bcd |
| TZA5552 | 12.3 ± 0.7 abc | 18 ± 0.6 bc | 4.7 ± 0.3 cd | 31.3 ± 1.5 c | 17.7 ± 1.8 cde | 9 ± 1 cd | 0.9 ± 0.0 b | 13 ± 0.6 ab | 18 ± 0.6 c | 5 ± 0.6 bc | 14.7 ± 0.7 b | 14.7 ± 0.7 d | 9 ± 1 cd | 0.8 ± 0.1 bc |
| TZA3177 | 15.7 ± 0.7 bc | 21.7 ± 1.5 c | 6.3 ± 0.7 cd | 36 ± 1.2 c | 21.7 ± 3.8 de | 8.7 ± 0.3 bcd | 0.8 ± 0.0 b | 16 ± 0.6 eab | 21.7 ± 1.5 bc | 6 ± 0.6 bc | 19.3 ± 1.5 c | 19.3 ± 1.5 e | 0 bcd | 0.8 ± 0 b |
| TZA5157 | 16.7 ± 3.1 bc | 9.3 ± 0.9 ab | 1 ± 0.6 ab | 18 ± 0.6 cb | 9.3 ± 0.9 abc | 1 a | 1.2 ± 0.0 cd | 7.7 ± 0.3 cab | 9.3 ± 0.8 bc | 0 ± 0.00 a | 9 ± 1.15 b | 9 ± 1.2 bc | 1 ± 0 a | 1.0 ± 0.1 bcd |
| V1030462 | 6. ± 1.2 a | 1 ± 0.0 a | 0.6 ± 0.3 a | 9 ± 1.7 a | 3 ± 0.6 a | 0 a | 1.4 ± 0.0 bcd | 1.7 ± 0.3 a | 1 ± 0.00 a | 0 ± 0.00 a | 3 ± 0.58 a | 3 ± 0.58 a | 0 ± a | 1.1 ± 0.11 cd |
| TZA5496 | 70.3 ± 0 f | 62.3 ± 4.1 d | 10.7 ± 1.5 e | 84 ± 3.5 e | 72.3 ± 4.3 f | 11 ± 1.2 d | 0.1 ± 0.07 a | 73 ± 7.3 c | 62.3 ± 4.0 d | 7.3 ± 0.3 c | 57 ± 4.7 e | 48.7 ± 3.2 f | 10.67 ± 0.9 d | 0.1 ± 0.1 a |
| F-S | 138.4 | 82.1 | 41.1 | 176.3 | 71.2 | 36.2 | 68.2 | 56.1 | 71 | 29.4 | 169.9 | 47 | 42.9 | 33.7 |
| p | ≤0.001 | ≤0.001 | ≤0.001 | ≤0.001 | ≤0.001 | ≤0.001 | ≤0.001 | ≤0.001 | ≤0.001 | ≤0.001 | ≤0.001 | ≤0.001 | ≤0.001 | ≤0.001 |
| Adult Whiteflies | Egg/Nymph | Honeydew Deposition | Yield (Kg) | ||
|---|---|---|---|---|---|
| Adult Whiteflies | Pearson’s r | - | |||
| p-value | - | ||||
| Egg/Nymph | Pearson’s r | 0.928 | - | ||
| p-value | <0 .001 | - | |||
| Honeydew Deposition | Pearson’s r | 0.826 | 0.662 | - | |
| p-value | <0.001 | <0.001 | - | ||
| Yield (Kg) | Pearson’s r | −0.921 | −0.804 | −0.849 | - |
| p-value | <0.001 | <0.001 | <0.001 | - |
| Chemical Information | Accessions | References | |||
|---|---|---|---|---|---|
| Name | Formula | Class | R/T (Min) | ||
| 1,2-15,16-Diepoxyhexadecane | C16H30O2 | 34.769 | TZA5562 | Unknown | |
| 3,7,11,15-Tetramethyl-2-hexadecan-ol | C20H40O | Phytol/Diterpene | 15.034 | V1030462, TZA5562, TZA5157 | Insect repelant [24], Toxic to Insects and herbivore [25] |
| Demeclocycline/Declomycin/Clortetrin | C21H21ClN2O8 | 50.968 | TZA5562 | Unknown | |
| 4aR*,6S*,7S*,7As*,8E,10R*,11R*,11As* | C32H39NO10 | Alkaloid | V1030462, TZA3729, TZA5562, TZA5157 | Affect insect nerve transmission [26] | |
| 2-(5-(5-[Cyano-(9,9-dimethyl-1,4-dioxa-7-aza-spiro [4.4] 3-Pyridinecarboxylic acid,2,7,10-tris(acetyloxy-1,1a,2,3,4,6,7,10,11,11a-decahydro-1,1,3,6,9-pentamethyl-4-oxo-4a,7a-epoxy-5H-cyclopenta[a]cyclopropa[f]cycloundecen-11-yl ester,[1aR-(1aR*2R*,3S*.non-7en-8-yl)-methylene]-3,3-dimethylpyrrolidin-2-ylidenemethyl)-3,3-dimethyl-ᵟ1-pyrrolin-5-ylidenemethyl-4,4,4-trimethylᵟ1-pyrroline-5-carbonitrile] | C32H42N6O2 | Alkaloid | V1030462, TZA3729 | Affect insect physiology and behaviour [27] | |
| 7,8-Epoxylonostan-11-ol,3-acetoxy- | C32H54O4 | Flavonoid/Phenol | 49.417 | TZA5562 | Insecticidal activity [28] Insect pests feeding Deterrents [29] Antifeedant Regulates plant responses to environmental stress [30] |
| Hematoporphyrin ix | C34H38N4O6 | Alkaloid | 25.418 | V1030462, TZA3729 | Affect insect nerve transmission [26] |
| Lycoxanthin | C40H56O | Tetraterpene | 48.073 | V1030462, TZA3729, TZA5562, TZA5157, TZA5496 | toxins to herbivore, feeding deterrents, or oviposition deterrents [31] |
| 4a-Phorbol12,13-didecanoate | C40H64O8 | Tetraterpene | 30.152 | V1030462, TZA3729, TZA5562, TZA5157 | Expression of genes responsible for plant defense upon herbivore attack [32] |
| Rhodopin | C40H58O | Tetraterpene | 28.870 | V1030462, TZA3729, TZA5562 | Mediate manufacturing of compounds in response to stress factors and herbivore attack [33] |
| Rhodoxanthin | C40H50O2 | Tetraterpene | 48.462 | V1030462, TZA3729, TZA5562 | Act against pathogens, herbivores [34] |
| Lycopene/β carotene | C40H56 | Tetraterpene | 28.870 | V1030462, TZA3729, TZA5157 | Repellant to herbivore arthropods [35] |
| Β,.Psi,-Carotene,3′,4,-didehydro-1′,2′-dihydro-1,′2′,-dihydroxy-,(2′R)- | C40H56O2 | Tetraterpene | 31.485 | V1030462, TZA3729, TZA5157 | Act against pathogens, herbivores [34] |
| Psi,psi,-Carotene,1,1′,2,2′,-tetrahydro-1,1′,-dimethoxy- | C42H64O2 | Carotenoid | 43.564 | V1030462, TZA3729, TZA5562, TZA5157 | Unknown |
| L-Lysine,N6-acetyl-N2-[N-[N(N2-acetyl-N,N,N2-trimethyl-Lasparoginyl)-N-methyl-L-phenylalanui]-N-methyl-L-phenylalanyl]-N,1-dimethyl-L-tryptophyl]-2N,N6-dimethyl-methyl ester. | C53H72N8O9 | Alkaloid | 42.17428.870 | V1030462, TZA3729, TZA5562 | Insect toxicity [31] Affect insect cytoskeleton, cell membrane and result into cell leakage and collapse [36] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mrosso, S.E.; Ndakidemi, P.A.; Mbega, E.R. Characterization of Secondary Metabolites Responsible for the Resistance of Local Tomato Accessions to Whitefly (Bemisia tabaci, Gennadius 1889) Hemiptera in Tanzania. Crops 2022, 2, 445-460. https://doi.org/10.3390/crops2040032
Mrosso SE, Ndakidemi PA, Mbega ER. Characterization of Secondary Metabolites Responsible for the Resistance of Local Tomato Accessions to Whitefly (Bemisia tabaci, Gennadius 1889) Hemiptera in Tanzania. Crops. 2022; 2(4):445-460. https://doi.org/10.3390/crops2040032
Chicago/Turabian StyleMrosso, Secilia E., Patrick A. Ndakidemi, and Ernest R. Mbega. 2022. "Characterization of Secondary Metabolites Responsible for the Resistance of Local Tomato Accessions to Whitefly (Bemisia tabaci, Gennadius 1889) Hemiptera in Tanzania" Crops 2, no. 4: 445-460. https://doi.org/10.3390/crops2040032