
Plant Growth and Yield Response to Salinity Stress of Rice Grown under the Application of Different Nitrogen Levels and Bacillus pumilus Strain TUAT-1

Abstract
:1. Introduction
2. Materials and Methods
2.1. Nursery Preparation
2.2. Nursery Seedling Vigor Measurement
2.3. Pot Experiments and Plant Growth Conditions
2.4. Growth and Yield Attributes
2.5. Statistical Analysis
3. Results and Discussions
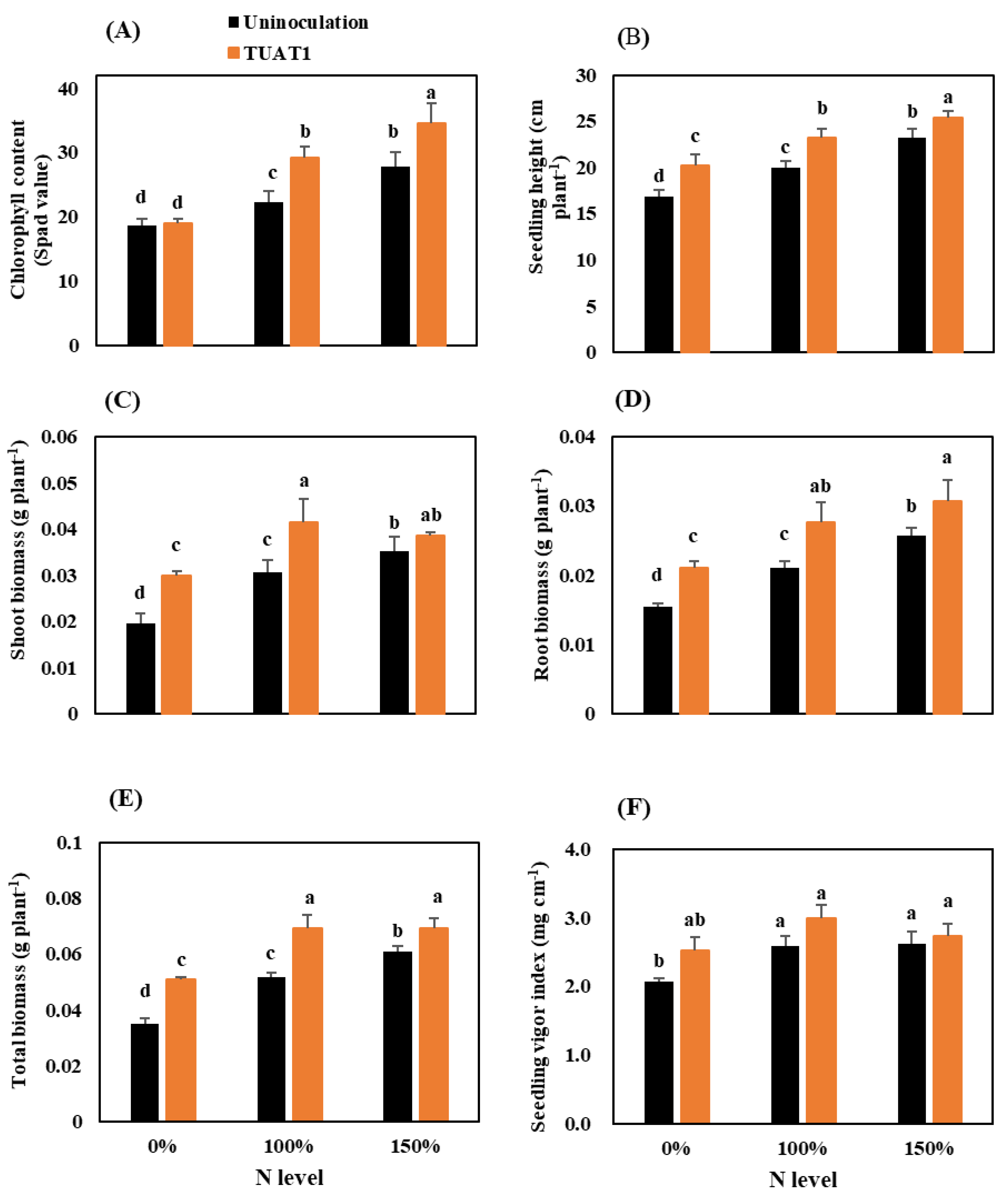
3.1. Effect of N Levels and TUAT-1 on Seedling Vigor
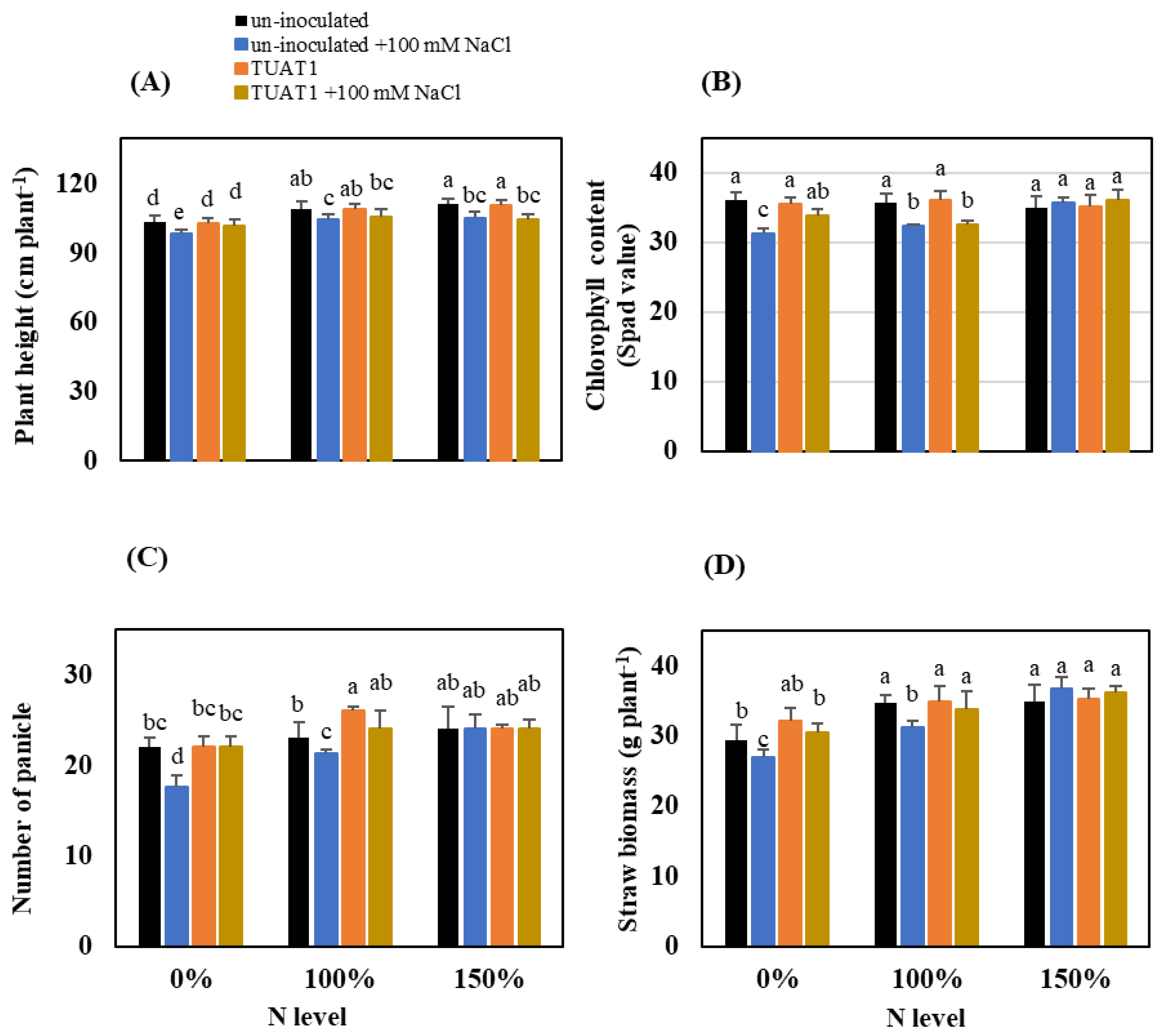
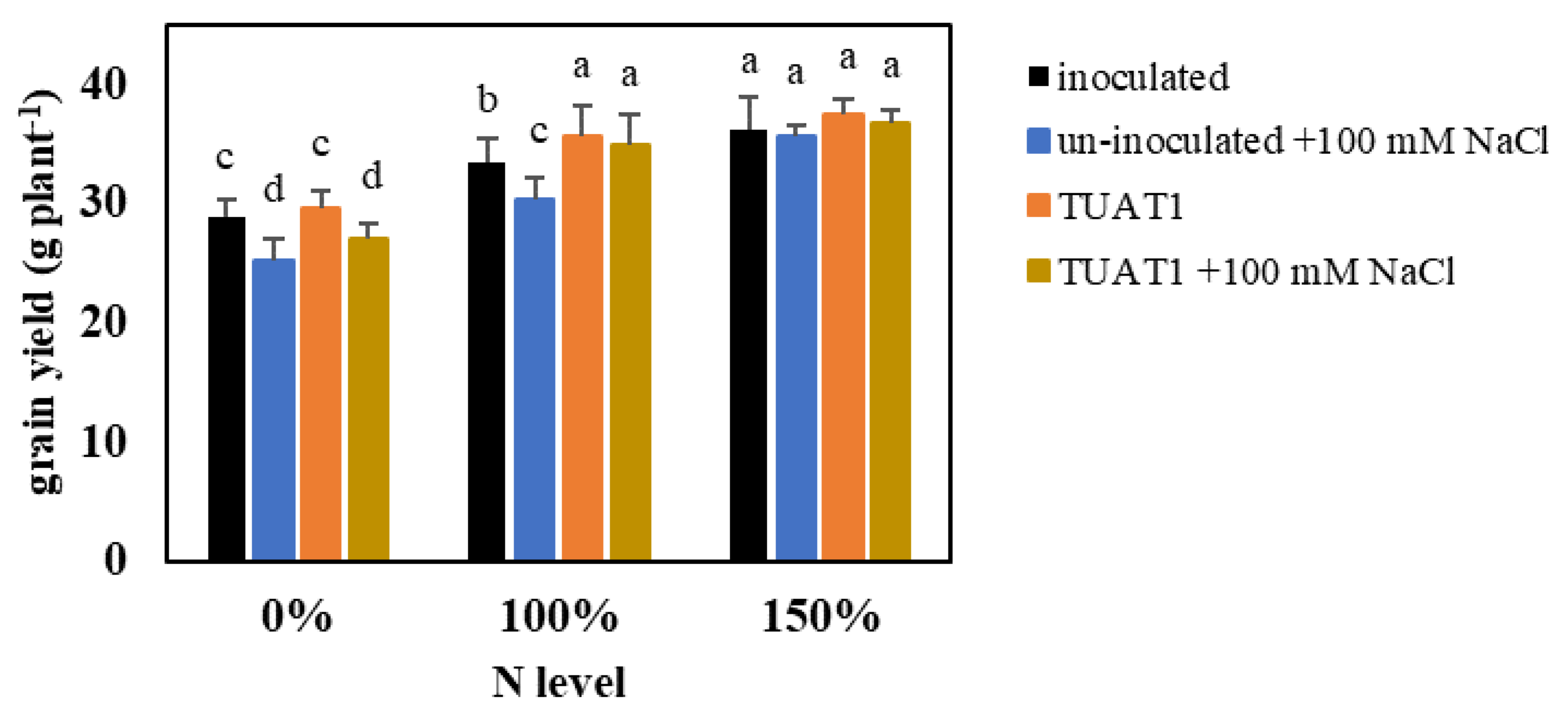
3.2. Rice Growth and Yield under NaCl Stress
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gamal-Eldin, H.; Elbanna, K. Field evidence for the potential of Rhodobacter capsulatus as biofertilizer for flooded rice. Curr. Microbiol. 2011, 62, 391–395. [Google Scholar] [CrossRef] [PubMed]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Bio. 2008, 59, 651–681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Win, K.T.; Oo, A.Z.; Ookawa, T.; Kanekatsu, M.; Hirasawa, T. Changes in hydraulic conductance cause the difference in growth response to short-term salt stress between salt-tolerant and sensitive black gram (Vigna mungo) varieties. J. Plant Physiol. 2016, 193, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Chakrabarti, N.; Mukherji, S. Effect of phytohormone pretreatment on nitrogen metabolism in Vigna radiata under salt stress. Biol. Plant. 2003, 46, 63–66. [Google Scholar] [CrossRef]
- Luque, A.A.; Bingham, F.T. The effect of the osmotic potential and specific ion concentration of the nutrient solution on the uptake and the reduction of nitrate by barley seedlings. Plant Soil 1981, 63, 227–237. [Google Scholar] [CrossRef]
- Palfi, G. A correlation between nitrogen nutrition of rice and asparagines concentration in leaves. Növenyterméles 1963, 12, 157–168. [Google Scholar]
- Cordovilla, M.P.; Ligero, F.; Liuch, C. The effect of salinity on nitrogen fixation and assimilation in Vicia faba. J. Exp. Bot. 1994, 45, 1483–1488. [Google Scholar] [CrossRef]
- Figueira, E.M.A.P.; Calderia, G.C.N. Effect of nitrogen nutrition on salt tolerance of Pisum salivum during vegetative growth. J. Plant Nutri. Soil Sci. 2005, 168, 359–363. [Google Scholar] [CrossRef]
- Papadopoulos, I.; Rendig, V.V. Interactive effects of salinity and nitrogen on growth and yield of tomato plants. Plant Soil 1983, 73, 47–57. [Google Scholar] [CrossRef]
- Ravikovitch, S.; Porath, A. The effect of nutrients on the salt tolerance of crops. Plant Soil 1967, 26, 49–71. [Google Scholar] [CrossRef]
- Mohanty, N.; Rath, A.K.; Nayak, D.N. Reclamation of salt amended soil for enhancement of growth and yield of rice. Oryza 1999, 36, 318–321. [Google Scholar]
- Zhao, B.; Dong, S.; Zhang, J.; Liu, P. Effects of Controlled-Release Fertiliser on Nitrogen Use Efficiency in Summer Maize. PLoS ONE 2013, 8, e70569. [Google Scholar] [CrossRef] [PubMed]
- Dimkpa, C.; Weinand, T.; Ash, F. Plant-rhizobacteria interactions alleviate abiotic stress conditions. Plant Cell Environ. 2009, 32, 1682–1694. [Google Scholar] [CrossRef] [PubMed]
- Arshad, M.; Shahroona, B.; Mahmood, T. Inoculation with Pseudomonas spp. containing ACC-Deaminase partially eliminates the effects of drought stress on growth, yield, and ripening of pea (Pisum sativum L.). Pedosphere 2008, 18, 611–620. [Google Scholar] [CrossRef]
- Marulanda, A.; Azcon, R.; Chaumont, F.; Ruiz-Lozano, J.M.; Aroca, R. Regulation of plasma membrane aquaporins by inoculation with a Bacillus megaterium strain in maize (Zea mays L.) plants under unstressed and salt-stressed conditions. Planta 2010, 232, 533–543. [Google Scholar] [CrossRef]
- Egamberdiyeva, D.; Höflich, G. Influence of growth-promoting bacteria on the growth of wheat in different soils and temperatures. Soil Biol. Biochem. 2003, 35, 973–978. [Google Scholar] [CrossRef]
- Barka, E.A.; Norwak, J.; Clément, C. Enhancement of Chilling Resistance of Inoculated Grapevine Plantlets with a Plant Growth-Promoting Rhizobacterium, Burkholderia phytofirmans Strain. Appl. Environ. Microbiol. 2006, 72, 7246–7252. [Google Scholar] [CrossRef] [Green Version]
- Fu, Q.; Liu, C.; Ding, N.; Lin, Y.; Guo, B. Ameliorative effects of inoculation with the plant growth-promoting rhizobacterium Pseudomonas sp. DW1 on growth of eggplant (Solanum melongena L.) seedlings under salt stress. Agric. Water Manag. 2010, 97, 1994–2000. [Google Scholar] [CrossRef]
- Yao, L.; Wu, Z.; Zheng, Y.; Kaleem, I.; Li, C. Growth promotion and protection against salt stress by Pseudomonas putida Rs-198 on cotton. Eur. J. Soil Biol. 2010, 46, 49–54. [Google Scholar] [CrossRef]
- Rojas-Tapias, D.; Moreno-Galván, A.; Pardo-Díaz, S.; Obando, M.; Rivera, D.; Bonilla, R. Effect of inoculation with plant growth-promoting bacteria (PGPB) on amelioration of saline stress in maize (Zea mays). Appl. Soil Ecol. 2012, 61, 264–272. [Google Scholar] [CrossRef]
- Upadhyay, S.K.; Singh, J.S.; Saxena, A.K.; Singh, D.P. Impact of PGPR inoculation on growth and antioxidant status of wheat under saline conditions. Plant Biol. 2012, 14, 605–611. [Google Scholar] [CrossRef] [PubMed]
- Win, K.T.; Tanaka, F.; Okazaki, K.; Ohwaki, Y. The ACC deaminase expressing endophyte Pseudomonas spp. Enhances NaCl stress tolerance by reducing stress-related ethylene production, resulting in improved growth, photosynthetic performance, and ionic balance in tomato plant. Plant Physiol. Biochem. 2018, 127, 599–607. [Google Scholar] [CrossRef] [PubMed]
- Panwar, M.; Tewari, R.; Nayyar, H. Native halo-tolerant plant growth promoting rhizobacteria Enterococcus and Pantoea sp. improve seed yield of Mungbean (Vigna radiata L.) under soil salinity by reducing sodium uptake and stress injury. Physiol. Molec. Plants. 2016, 22, 445–459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sukweenadhi, J.; Kim, Y.-J.; Choi, E.-S.; Koh, S.-C.; Lee, S.-W.; Kim, Y.-J.; Yang, D.C. Paenibacillus yonginesis DCY84 T induces changes in Arabidopsis thaliana gene expression against aluminum, drought, and salt stress. Microbiol. Res. 2015, 172, 7–15. [Google Scholar] [CrossRef]
- Sapre, S.; Gontia-Mishra, I.; Tiwari, S. Klebsiella sp. confers enhanced tolerance to salinity and plant growth promoting in oat seedlings (Avena sativa). Microbiol. Res. 2018, 206, 25–32. [Google Scholar] [CrossRef]
- Sirajuddin; Khan, A.; Ali, L.; Chaudhary, H.J.; Munis, F.H.; Bano, A.; Masood, S. Bacillus pumilus alleviates boron toxicity in tomato (Lycopersicum esculentum L.) due to enhanced antioxidant enzymatic activity. Sci. Hortic. 2016, 200, 178–185. [Google Scholar] [CrossRef]
- Torii, A. Analysis of Field Factors Resulting Frutuations of Yield and Nutritional Uptakes of Forage Rice Leaf Star with Inoculation of an Endophytic Nitrogen Fixing Bacteria TUAT-1. Master’s Thesis, Graduate School of Agriculture, Tokyo University of Agriculture and Technology, Fuchu City, Tokyo, Japan, March 2012. [Google Scholar]
- Win, K.T.; Oo, A.; Ohkama-Ohtsu, N.; Tadashi, Y. Bacillus Pumilus Strain TUAT-1 and Nitrogen Application in Nursery Phase Promote Growth of Rice Plants under Field Conditions. Agronomy 2018, 8, 216. [Google Scholar] [CrossRef] [Green Version]
- Aung, H.P.; Salem, D.; Oo, A.Z.; Aye, Y.S.; Yokoyama, T.; Suzuki, S.; Sekimoto, H.; Bellingrath-Kimura, S.D. Growth and 137Cs uptake of four Brassica species influenced by inoculation with a plant growth-promoting rhizobacterium Bacillus pumilus in three contaminated farmlands in Fukushima prefecture, Japan. Sci. Total Environ. 2015, 521–522, 261–269. [Google Scholar] [CrossRef]
- Walia, H.; Wilson, C.; Condamine, P.; Liu, X.; Ismail, A.M.; Zeng, L.; Wanamaker, S.I.; Mandal, J.; Xu, J.; Cui, X.; et al. Comparative Transcriptional profiling of two contrasting rice genotypes under salinity stress during vegetative growth stage. Plant Physiol. 2005, 139, 822–835. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.; Hamid, A.; Karim, M. Effect of sodium chloride on germination and seedling characters of different types of rice (Oryza saliva. L.). J. Agron. Crop. Sci. 1997, 179, 163–169. [Google Scholar] [CrossRef]
- Surekha, R.; Mishra, P.B.; Gupta, S.R.; Rathore, A. Reproductive stage tolerance to salinity and alkalinity stresses in rice genotypes. Plant Breed. 2008, 127, 256–261. [Google Scholar]
- Win, K.T.; Okazaki, K.; Ookawa, T.; Yokoyama, T.; Ohwaki, Y. Influence of rice-husk biochar and Bacillus pumilus strain TUAT-1 on yield, biomass production, and nutrient uptake in two forage rice genotypes. PLoS ONE 2019, 14, e0220236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lynch, J.P.; Lynch, A.F.; Jonathan, P. Roots of the second green revolution. Aust. J. Bot. 2007, 55, 493–512. [Google Scholar] [CrossRef]
- Sharma, A.; Shankhdhar, D.; Sharma, A.; Shankhdhar, S.C. Growth promotion of the rice genotypes by PGPRs isolated from rice rhizosphere. J. Soil Sci. Plant Nutr. 2014, 14, 505–517. [Google Scholar] [CrossRef] [Green Version]
- Thorup-Kristensen, K.; Kirkegaard, J. Root system-based limits to agricultural productivity and efficiency: The farming systems context. Ann. Bot. 2016, 118, 573–592. [Google Scholar] [CrossRef]
- Kloepper, J.W.; Zablotowick, R.M.; Tipping, E.M.; Lifshitz, R. Plant growth promotion mediated by bacterial rhizosphere colonizers. In The Rhizosphere and Plant Growth; Keister, D.L., Cregan, P.B., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1991; pp. 315–326. [Google Scholar]
- Song, X.; Zhou, G.; Ma, B.-L.; Wu, W.; Ahmad, I.; Zhu, G.; Yan, W.; Jiao, X. Nitrogen application improved photosynthetic productivity, chlorophyll fluorescence, yield and yield components of two oat genotypes under salinity conditions. Agronomy 2019, 9, 115. [Google Scholar] [CrossRef] [Green Version]
- Flores, P.; Carvajal, M.; Cerda, A.; Martinez, V. Salinity and ammonium/nitrate interactions on tomato plant development, nutrition, and metabolites. J. Plant Nutr. 2012, 4, 1561–1573. [Google Scholar] [CrossRef]
- Razzaque, S.; Haque, T.; Elias, S.M.; Rahman, S.; Biswas, S.; Schwartz, S.; Ismail, A.M.; Walia, H.; Juenger, T.E.; Seraj, Z.I. Reproductive stage physiological and transcriptional responses to salinity stress in reciprocal populations derived from tolerance (Horkuch) and susceptible (IR29) rice. Sci. Rep. 2017, 7, 46138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, G.; Wang, Y.; Shi, X.; Lu, H.; Ren, Z.; Shi, Y.; Jiao, X.; Ibrahim, M.E.H.; Irshad, A.; Zhu, W.; et al. Optimum nitrogen management enhances growth, antioxidant ability and yield performance of rice in saline soil of coastal area of China. Chil. J. Agric. Res. 2020, 80, 629–639. [Google Scholar] [CrossRef]
- Idris, E.E.S.; Iglesias, D.J.; Talon, M.; Borriss, R. Tryptophan-dependent production of Indole 3-Acetic Acid (IAA) affects level of plant growth promotion by Bacillus amyloliquefaciens FZB42. Mol. Plant Microbe Interact. 2007, 20, 619–626. [Google Scholar] [CrossRef] [Green Version]
- Nautiyal, C.S.; Srivastava, S.; Chauhan, P.S.; Seem, K.; Mishra, A.; Sopory, S.K. Plant growth-promoting bacteria Bacillus amyloliquefaciens NBRISN13 modulates gene expression profile of leaf and rhizosphere community in rice during salt stress. Plant Physiol. Biochem. 2013, 66, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Okazaki, S.; Yokoyama, T. Bacillus pumilus DNA, Complete Genome, Strain: TUAT1. Accession: NZ_AP014928, NCBI (29 July 2015); Tokyo University of Agriculture and Technology: Fuchu, Japan, 2015. Available online: https://www.ncbi.nlm.nih.gov/nuccore/NZ_AP014928 (accessed on 12 June 2022).
- Abbas, R.; Rasul, S.; Aslam, K.; Baber, M.; Shahid, M.; Mubeen, F.; Naqqash, T. Halotolerant PGPR: A hope for cultivation of saline soils. J. King Saud. Univ. Sci. 2019, 31, 1195–1201. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| N Levels (%) | Root Dry Weight (g plant−1) | Root Surface Area (cm2 plant−1) | Total Root Length (cm plant−1) | Root Number plant−1 | |
|---|---|---|---|---|---|
| Un-inoculated | N0 | 0.015 bc ± 0.001 | 12.67 d ± 2.7 | 240.28 e ± 32.4 | 9.66 d ± 0.9 |
| N100 | 0.021 b ± 0.001 | 20.74 c ± 3.5 | 340.86 d ± 48.0 | 13.32 bc ± 1.1 | |
| N150 | 0.021 b ± 0.001 | 29.75 b ± 5.3 | 450.84 c ± 66.0 | 14.63 b ± 1.4 | |
| TUAT-1 | N0 | 0.028 ab ± 0.001 | 20.02 c ± 3.6 | 342.41 d ± 47.0 | 11.14 c ± 0.9 |
| N100 | 0.026 ab ± 0.001 | 35.89 ab ± 0.7 | 582.66 b ± 42.4 | 15.61 ab ± 1.2 | |
| N150 | 0.031 a ± 0.003 | 42.61 a ± 0.2 | 753.73 a ± 54 | 16.19 a ± 1.1 | |
| Analysis of variance (p value) | |||||
| N | 0.000 | 0.000 | 0.015 | 0.010 | |
| T | 0.000 | 0.000 | 0.000 | 0.015 | |
| N × T | 0.651 | 0.028 | 0.015 | 0.015 | |
| ANOVA (p Values) | |||||
|---|---|---|---|---|---|
| SPAD Value | Plant Height (cm) | Number of Panicle | Straw Biomass (g plant−1) | Grain Yield (g plant−1) | |
| N | 0.011 | 0.004 | 0.005 | 0.000 | 0.003 |
| T | 0.558 | 0.389 | 0.008 | 0.000 | 0.002 |
| N × T | 0.978 | 0.452 | 0.012 | 0.007 | 0.171 |
| S | 0.001 | 0.066 | 0.048 | 0.137 | 0.004 |
| N × S | 0.005 | 0.67 | 0.245 | 0.829 | 0.006 |
| T × S | 0.154 | 0.254 | 0.376 | 0.001 | 0.214 |
| N × S × T | 0.125 | 0.912 | 0.082 | 0.732 | 0.005 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Win, K.T.; Oo, A.Z.; Yokoyama, T. Plant Growth and Yield Response to Salinity Stress of Rice Grown under the Application of Different Nitrogen Levels and Bacillus pumilus Strain TUAT-1. Crops 2022, 2, 435-444. https://doi.org/10.3390/crops2040031
Win KT, Oo AZ, Yokoyama T. Plant Growth and Yield Response to Salinity Stress of Rice Grown under the Application of Different Nitrogen Levels and Bacillus pumilus Strain TUAT-1. Crops. 2022; 2(4):435-444. https://doi.org/10.3390/crops2040031
Chicago/Turabian StyleWin, Khin Thuzar, Aung Zaw Oo, and Tadashi Yokoyama. 2022. "Plant Growth and Yield Response to Salinity Stress of Rice Grown under the Application of Different Nitrogen Levels and Bacillus pumilus Strain TUAT-1" Crops 2, no. 4: 435-444. https://doi.org/10.3390/crops2040031