
Angiodrastic Chemokines Production by Colonic Cancer Cell Lines
by

Emmanouil George
1, 
Moursellas Andrew
1,
Tzardi Maria
2,
Voumvouraki Argyro
1 and
Kouroumalis Elias
1,* 1
Gastroenterology Research Laboratory, Medical School, University of Crete, 71500 Heraklion, Greece
2
Department of Pathology, Medical School, University of Crete, 71500 Heraklion, Greece
*
Author to whom correspondence should be addressed.
Onco 2022, 2(2), 69-84; https://doi.org/10.3390/onco2020006
Submission received: 21 March 2022
/
Revised: 17 April 2022
/
Accepted: 25 April 2022
/
Published: 29 April 2022
Abstract
:Simple Summary
Colorectal cancer (CRC) is the fourth most common type of cancer and the third in terms of global mortality. Angiogenesis is believed to play a vital role in cancer development. In our previous work, we have demonstrated the significantly increased expression of angiogenetic chemokines (VEGF and CXCL8) in tumor tissues compared to normal colorectal epithelial tissue. However, the origin of these chemokines in the malignant tissue remains to be identified. Therefore, we studied the production of angiogenesis-related chemokines in two well-established colonic epithelial cell lines (HT-29 and Caco-2) under various culture conditions. Results showed that there were significant quantitative and time-related differences in chemokine production between these two cell lines.
Abstract
Purpose: To study the production of angiodrastic chemokines by colonic cancer cell lines. Methods: A pro-angiogenic factor (VEGF), two angiogenic chemokines (CXCL8, CXCL6), and one angiostatic (CXCL4) chemokine were measured by ELISA in the supernatants of the colon cancer cell lines HT-29 and Caco-2. Cells were cultured for 24 h in the presence of serum from cancer patients or healthy individuals. Results were analyzed by one-way ANOVA and the General Linear Model for repeated measures. Results: Colonic epithelial cells are potent producers of angiodrastic chemokines. HT-29 and Caco-2 cells produce all four chemokines under basal conditions and 24 h after incubation with human serum. The secretion response, however, was completely different. HT-29 cells produce more CXCL8 and VEGF irrespective of culture conditions, while Caco-2 cells seem unresponsive with respect to CXCL6 and CXCL4. Moreover, HT-29 cells produce more CXCL8 and VEGF when incubated with cancer serum, contrary to Caco-2 cells which produce more CXCL4 under the same conditions. Conclusions: The two colon cancer cell lines were producers of all chemokines studied, but their responses were not uniform under similar culture conditions. CXCL8 and VEGF are differently regulated compared to CXCL4 and CXCL6 in these two cell lines
1. Introduction
Colorectal cancer (CRC) is one of the most common causes of cancer mortality worldwide. It has been suggested that the interplay between the cancer cells themselves and the surrounding mesenchymal cells of the bowel wall stroma plays a vital role in cancer induction and progress. Interactions between these cell types can either be through a direct cell contact or through a paracrine signaling pathway involving growth factors, cytokines, and chemokines [1,2].
We have recently demonstrated that the angiogenic factors CXCL6 and VEGF are increased in colorectal cancer tissue. More importantly, increased levels of cancer tissue CXCL8 and CXCL4 chemokines were associated with poor survival, but the cellular origin of these chemokines was not identified [3].
In an effort to see if malignant colon epithelial cells could be a source of angiodrastic factors, we used the colonic epithelial cell lines HT-29 and Caco-2, which are two of the best-characterized cell lines. The human colon adenocarcinoma cell line HT-29 was isolated from a primary tumor of a Caucasian female in 1964 [4]. The Caco-2 cell line was established in 1977 from a human colorectal adenocarcinoma [5]. These two cell lines have a different metastatic potential and a different cytokine response profile after stimulation. We used the collective term angiodrastic to include both chemokines and other factors which either promote or inhibit the tumor vascular environment.
CXCL8, CXCL6, and CXCL4 are three angiodrastic chemokines, all belonging to the CXC chemokine family, but only the first two bear the ELR amino acid profile. The presence of this profile characterizes CXC chemokines promoting angiogenesis, while its absence indicates an angiostatic potential [6,7]. CXCL8 was the first chemokine with a proven pro-angiogenic activity overexpressed in cancer-surrounding stromal cells. Moreover, an autocrine growth factor role has been described for CXCL8 [8,9,10]. CXCL6 also bears the ELR amino acid profile and shares a 31% amino acid sequence homology with CXCL8. Therefore, it is not surprising that these two chemokines have similar properties [11,12]. Both CXCL8 and CXCL6 bind to the same receptors, CXCR1 and CXCR2, that mediate their chemotactic and angiogenic activities [13,14].
CXCL4, on the other hand, lacks the ELR amino acid profile, and it is the first described angiostatic chemokine inhibiting the angiogenic effects of VEGF and bFGF [15,16]. CXCL4 also inhibits endothelial cell chemotaxis [17]. CXCL4 circulates in two isoforms, the CXCL4 and CXCL4L1 variants, differing only by three amino acids. Although they both have angiostatic properties, the CXCL4L1 is the more potent factor [18].
VEGF is one of the major pro-angiogenesis factors in tumor biology. Its binding to three structurally similar receptors leads to endothelial cell proliferation, migration, survival, and angiogenesis [19,20]. Moreover, VEGF is capable of attracting macrophages that also participate in tumor development and progression [21].
In the current study, we investigated the secretion of three angiodrastic cytokines (CXCL8, CXCL4, and CXCL6) and the angiogenic factor VEGF by the HT-29 and Caco-2 malignant colonic cell lines under various cultural conditions.
2. Materials and Methods
The human colonic epithelial carcinoma cell lines HT-29 and Caco-2 were purchased from the European Collection of Animal Cell Cultures. Human serum used in cultures was available from five colon cancer patients selected according to the following criteria, as reported previously [3]:
- (a)
- Only patients with stages II and III according to TNM classification (stages B and C according to Dukes classification, as modified by Astler-Coller) were included. Metastatic disease (type IV) patients were excluded.
- (b)
- Only patients with a curative (R0) surgical resection who did not receive adjuvant chemotherapy (either refused or were not considered as candidates by the attending doctors) were selected.
The study was conducted in accordance with the Declaration of Helsinki and was approved by the Ethics Committee of the University Hospital of Heraklion, Heraklion. Since sera were randomly and anonymously selected without any use of demographic, clinical, or laboratory data from a large sera collection and were used only for in vitro cultures without clinical details, the Ethics Committee felt that there was no need for further consent.
2.1. Cell Cultures
Cell incubation was performed in a Thermo Scientific Incubator at 37 °C and 5% CO2. HT-29 and Caco-2 were cultured in McCoy’s 5A and minimum essential medium alpha (MEM), respectively, supplemented with 10% fetal bovine serum (FBS), penicillin/streptomycin (10 μ/mL and 10 mg/mL), and fungizone (0.5 mg/mL). Both cell lines were cultured in 75 cm2 Nunc flasks. When cultures became 90–95% confluent, the cells were removed from flasks and seeded in six-well plates at a 3 × 104 cell concentration, and were maintained at 37 °C, until confluent. Then, the culture medium was removed, and cultures were washed and re-cultured in fresh medium without FBS for 24 h. Cells were then stimulated with either 10% serum from colon cancer patients or 10% serum from healthy individuals for 2, 6, 12, and 24 h. Unstimulated cells were cultured with medium alone and served as controls. Five separate experiments were performed for each cell line and stimulant. Independent wells for each time point and stimulant were used. Supernatants were collected at each time point and stored at −80 °C until use.
2.2. Enzyme-Linked Immunosorbent Assay (ELISA)
ELISAs with commercially available monoclonal antibodies and biotinylated antibodies obtained from the R&D Systems (Minneapolis, MN, USA) were set up.
The antibodies were reconstituted with sterile Tris-Buffered Saline (TBS) according to the manufacturer’s instructions. After reconstitution, antibodies were aliquoted and stored at −20 °C. Then, 96-well plates were initially incubated overnight with 100 μL of PBS containing the captured antibody according to the manufacturer’s instructions (CXCL8 0.5 μg/mL, CXCL4 2 μg/mL, CXCL6 0.5 μg/mL, and VEGF 1 μg/mL), followed by 1 h incubation with 300 μL of PBS and 1% BSA to block all non-specific binding sites. Then, 100 μL of each supernatant was added in each well and incubated for 2 h at room temperature. Next, 100 μL of PBS containing the detection antibody was added at concentrations suggested by the manufacturer (CXCL8 0.2 μg/mL, CXCL4 100 ng/mL, CXCL6 100 ng/mL, and VEGF 0.4 μg/mL) and incubated for 2 h at room temperature, followed by 100 μL of Streptavidin-HRP for 30 min. Finally, 100 μL of Substrate Buffer was added in every sample, incubated for 20 min, and 50 μL of Stop Solution (2 NH2SO4) was added to each well and optical density was determined at 450 nm. Between steps, all wells were washed 3 times with 400 μL of PBS and 0.05% Tween 20.
Commercially available recombinant human CXCL8, CXCL4, CXCL6, and VEGF were obtained (R&D Systems Inc., Mineapolis, MN, USA) and used for generating standard curves. Every sample was run in duplicate.
2.3. Statistics
Statistical analysis was performed with the IBM SPSS statistics software version 19.
The Kolmogorov–Smirnov test was used to check for normality. One-way ANOVA with the Scheffé post hoc analysis was used for comparison between values (Ca serum stimulation, control serum stimulation, and culture medium) at each time point. After verification of normality, time (2, 4, 6, 12, and 24 h), cell lines (HT-29 and Caco-2), and the interaction between time and cell lines were checked as probable differentiation factors within groups. The General Linear Model for Repeated Measures test was used for testing differences between HT-29 and Caco-2 measurements.
Statistical significance was set at the 5% level (p = 0.05).
3. Results
3.1. Measurements over Time
3.1.1. HT-29—CXCL8
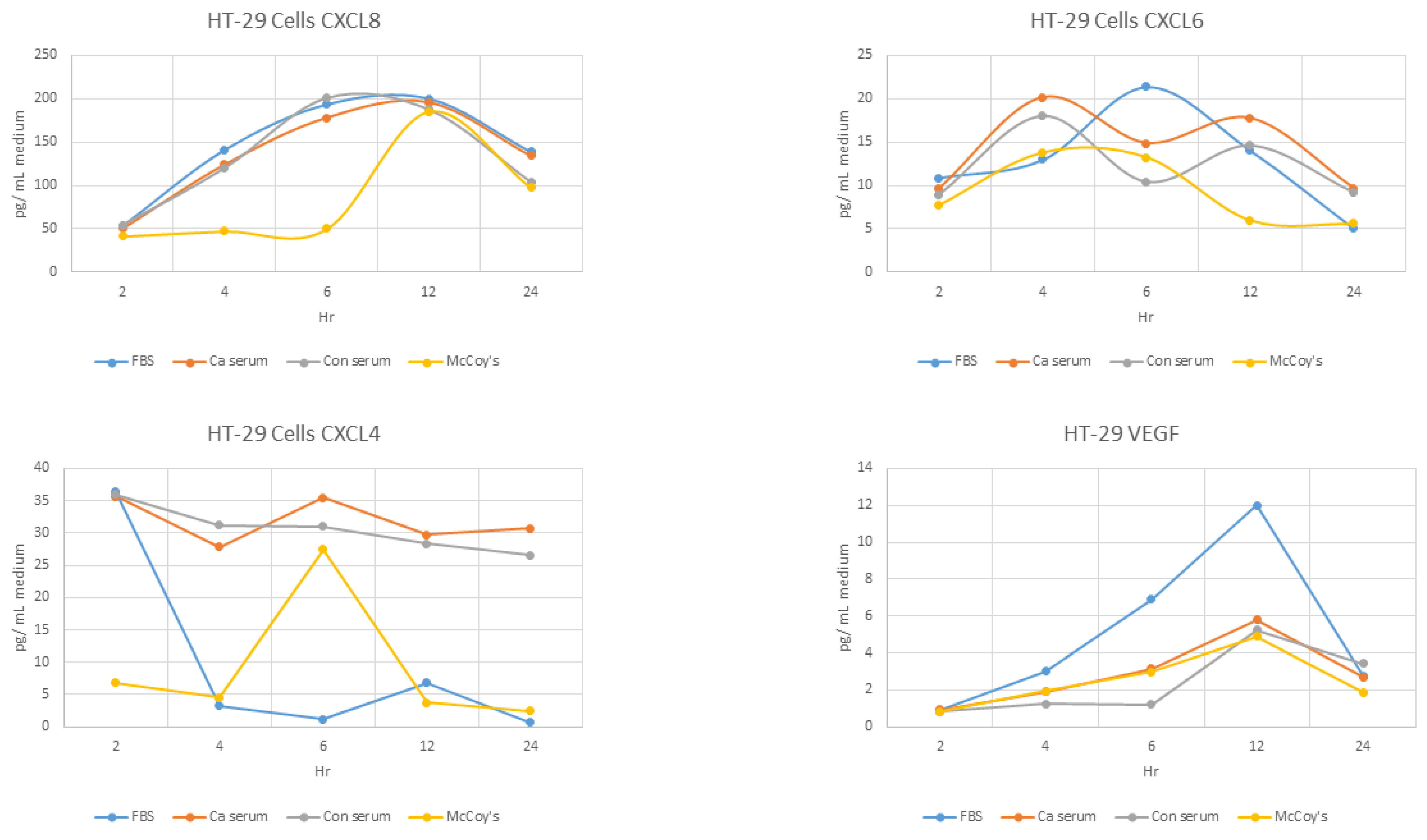
Overall, there was an early statistically significant difference at 2, 4, and 6 h of incubation between base values (unstimulated medium) and both cancer and normal serum, an effect that seems to be abolished at 12 and 24 h (p > 0.05) (Table 1, Figure 1).
Induction of CXCL8 by cancer serum was significantly less than induction by normal serum (p = 0.033) at 6 h.
3.1.2. HT-29—CXCL6
No difference was observed at any time point (p > 0.05) (Figure 1).
3.1.3. HT-29—CXCL4
3.1.4. HT-29—VEGF
Overall, a significant difference existed at all time points (p < 0.05), with the exception of the second hour (p > 0.05).
Individual comparisons revealed that there was no difference between basal and sera incubations at 4, 6, and 12 h incubations (p > 0.05), but at 24 h, the basal values were lower than both sera values. Moreover, cancer serum significantly reduced VEGF secretion compared to normal serum(Table 3, Figure 1).
3.1.5. Caco-2—CXCL8
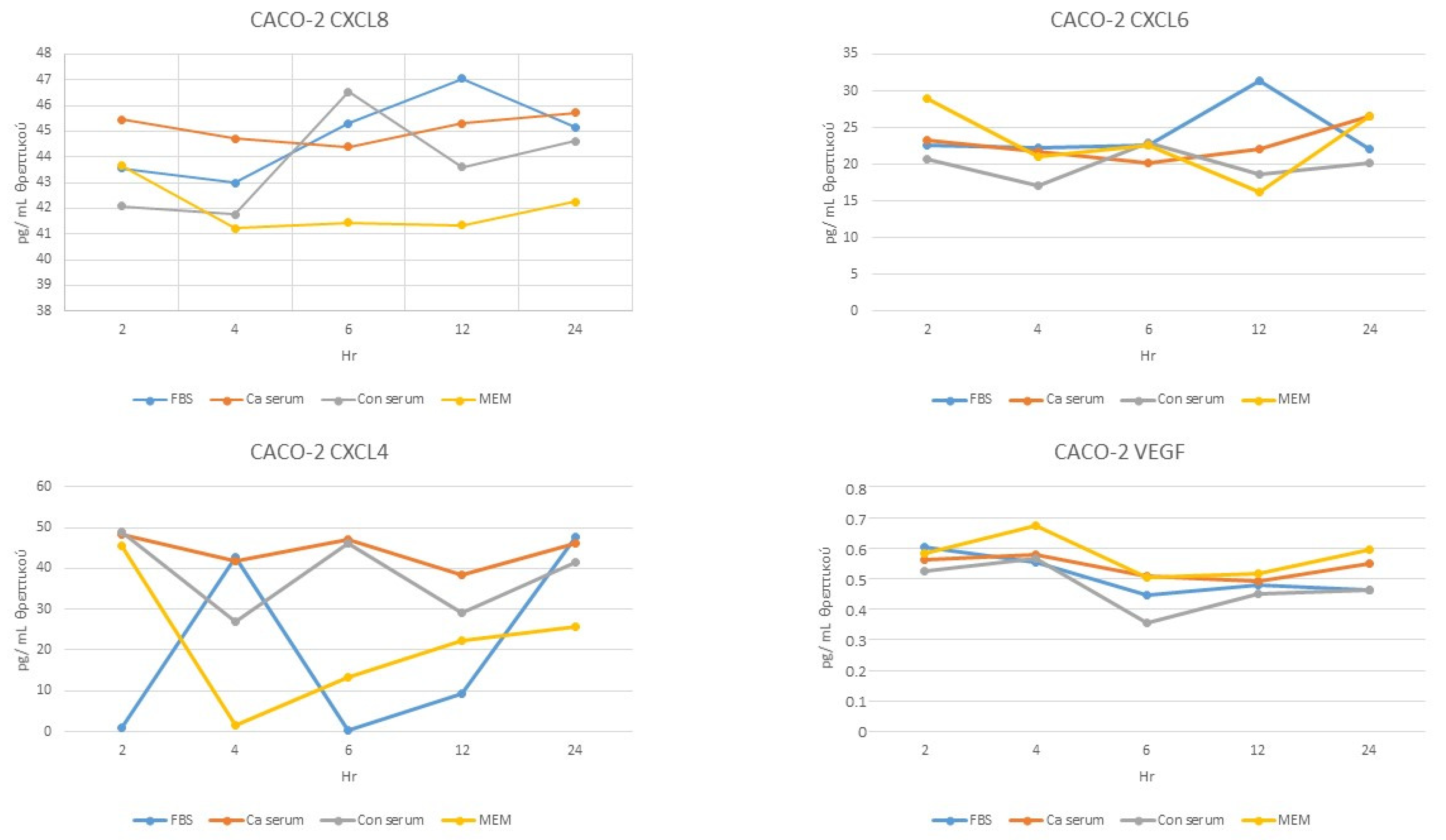
- Overall, there was a significant difference only at 4, 6, and 12 h (p < 0.05).
3.1.6. Caco-2—CXCL6
No difference was observed at any time point (p > 0.05) (Figure 2).
3.1.7. Caco-2—CXCL4
- Overall, significant differences existed at all time points (p < 0.05).
3.1.8. Caco-2—VEGF
No difference was observed at any time point (p > 0.05) (Figure 2).
3.2. Measurements over Time
3.2.1. CXCL8—Ca Serum
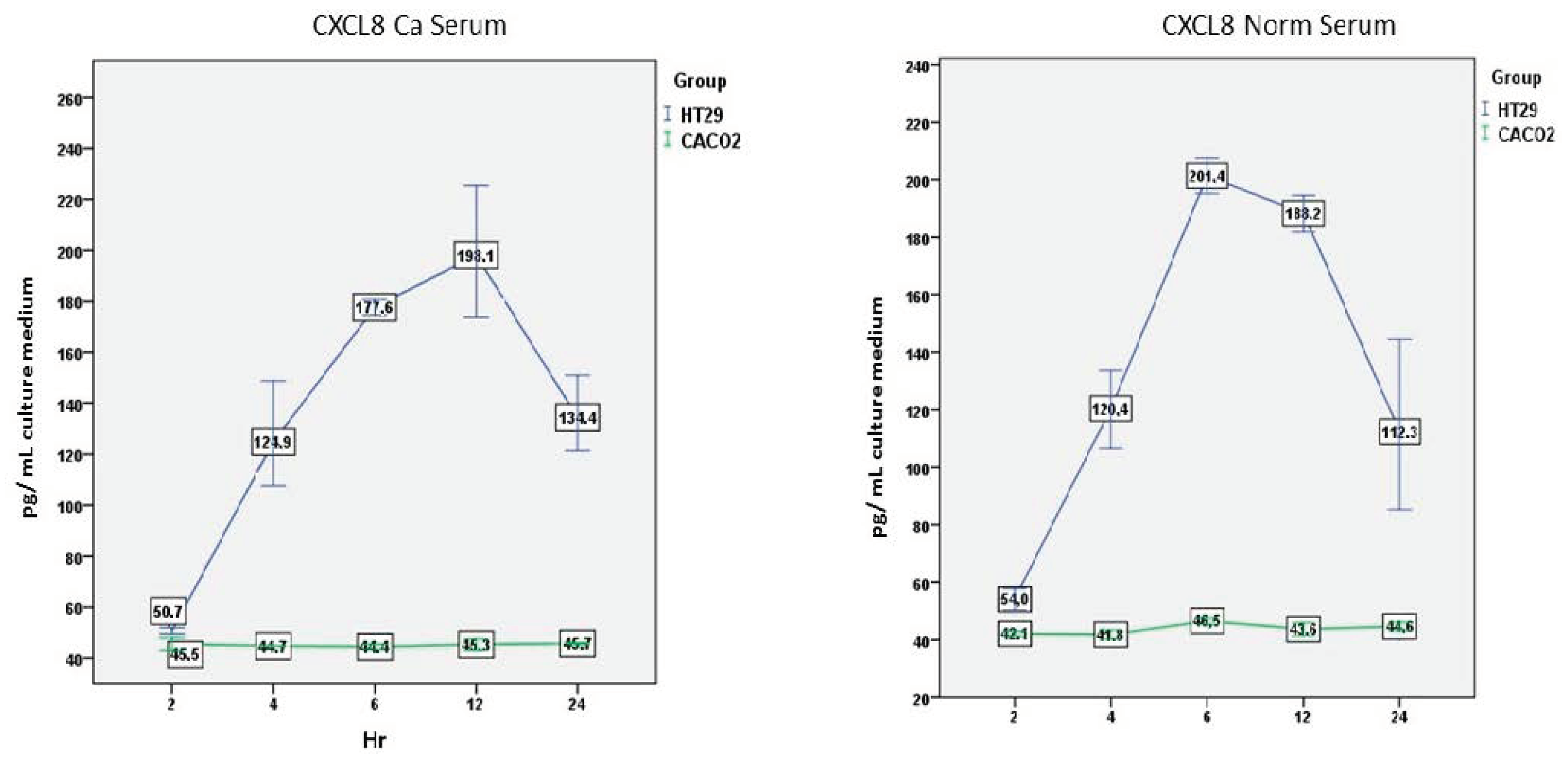
There was an overall statistically significant effect of the cell line (F(1,2) = 224.031, p = 0.004) and the time (F(4,8) = 10.519, p = 0.003), but also of the interaction between the cell line and the time (F(4,8) = 10.660, p = 0.003).
HT-29 cells produced significantly higher amounts of CXCL8 compared to Caco-2 cells at all time points (p < 0.05), with exception of the second hour (Figure 3).
3.2.2. CXCL8—Normal Serum
Results were similar after incubation with normal serum. There was a significant overall cell line effect (F(1,2) = 247.398, p = 0.004), time effect (F(4,8) = 8.775, p = 0.005), and interaction between time and cell line (F(4,8) = 8.028, p = 0.007) (Figure 3).
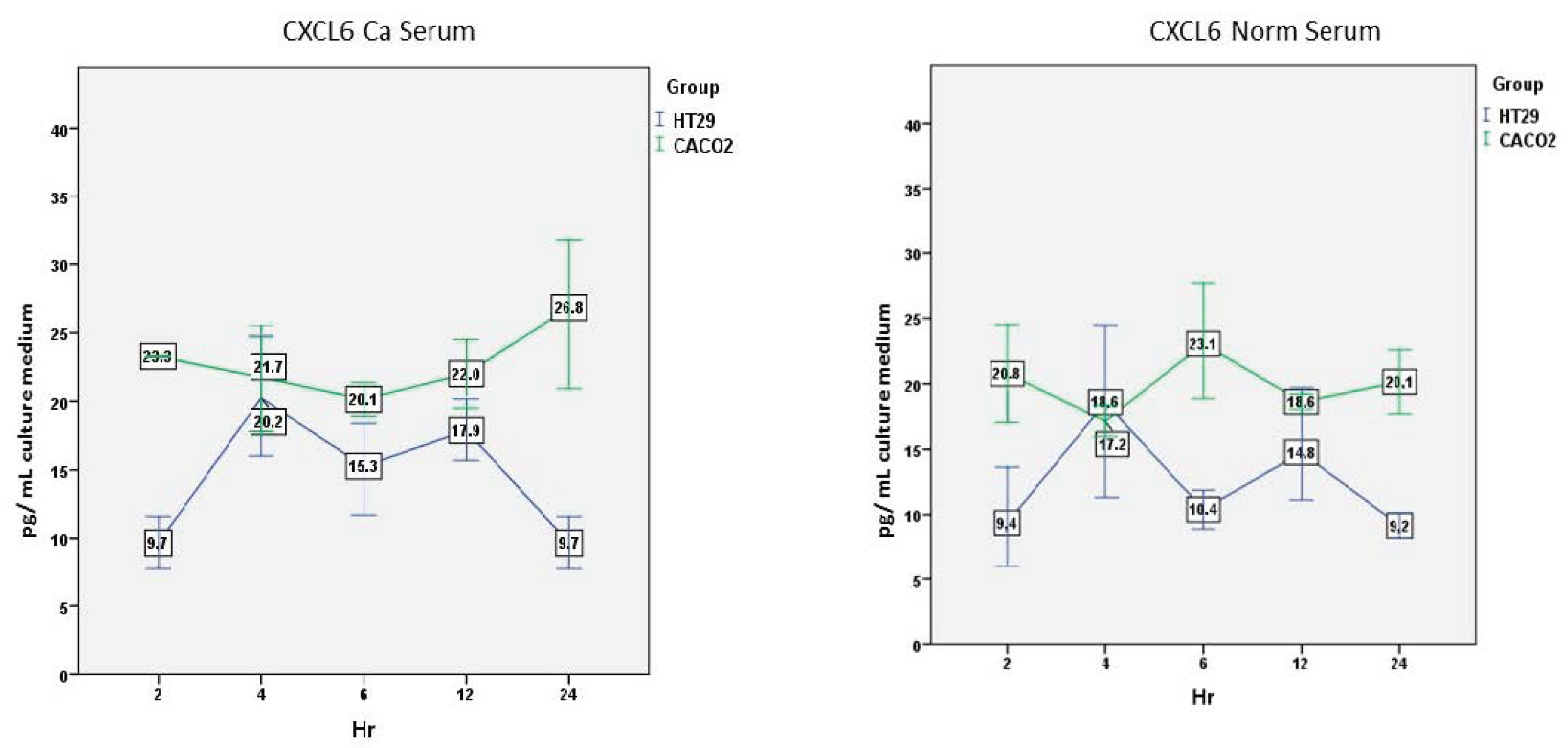
3.2.3. CXCL6—Ca Serum
There was a significant effect of the cell line (F(1,2) = 32.527, p = 0.029). This was not the case, however, for the time (F(4,8) = 0.458, p = 0.76) or the interaction between time and cell line (F(4,8) = 1.601, p = 0.26).
The time response was different here. Caco-2 cells produced higher levels of CXCL6 than HT-29 cells, which were relatively stable over time. On the other hand, HT-29 cells’ secretion was characterized by peaks which were significantly lower at 2 h (p = 0.005) (Figure 4).
3.2.4. CXCL6—Normal Serum
No significant effect on the cell line (F(1,2) = 4.399, p = 0.17), time (F(4,8) = 0.432, p = 0.78), or the interaction between time and cell line (F(4,8) = 2.287, p = 0.14) was found.
A similar picture to the incubation with cancer serum was also evident here, with higher values for the Caco-2 cells at all time points. Lower levels reached significance only at 24 h (p = 0.014) (Figure 4).
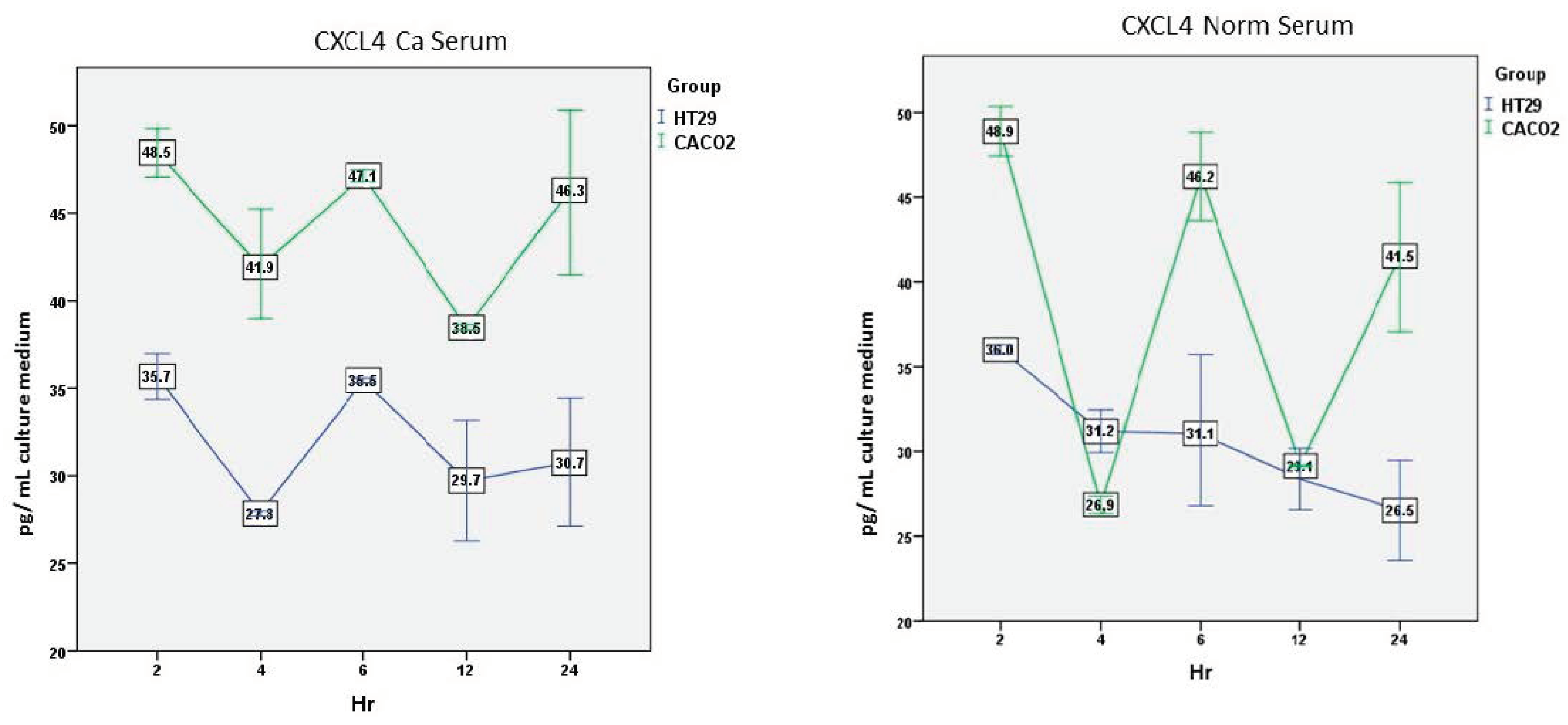
3.2.5. CXCL4—Ca Serum
There was a significant overall effect of the cell line (F(1,2) = 84.060, p = 0.012) and time (F(4,8) = 8.165, p = 0.006), but not of the interaction between time and cell line (F(4,8) = 1.037, p = 0.44).
Again, Caco-2 cells secreted significantly higher amounts of CXCL4 compared to HT-29 cells at all time points except 24 h (p < 0.05) (Figure 5).
3.2.6. CXCL4—Normal Serum
Overall, there was a significant effect of the cell line (F(1,2) = 33.158, p = 0.029), the time (F(4,8) = 24.209, p < 0.001), and the interaction between time and cell line (F(4,8) = 13.861, p = 0.001).
Caco-2 cells’ secretion was characterized by peaks which were significantly higher than HT-29 cell levels at 2, 6, and 24 h (p < 0.05). In contrast, HT-29 cells’ secretion was characterized by a gradual reduction over time (Figure 5).
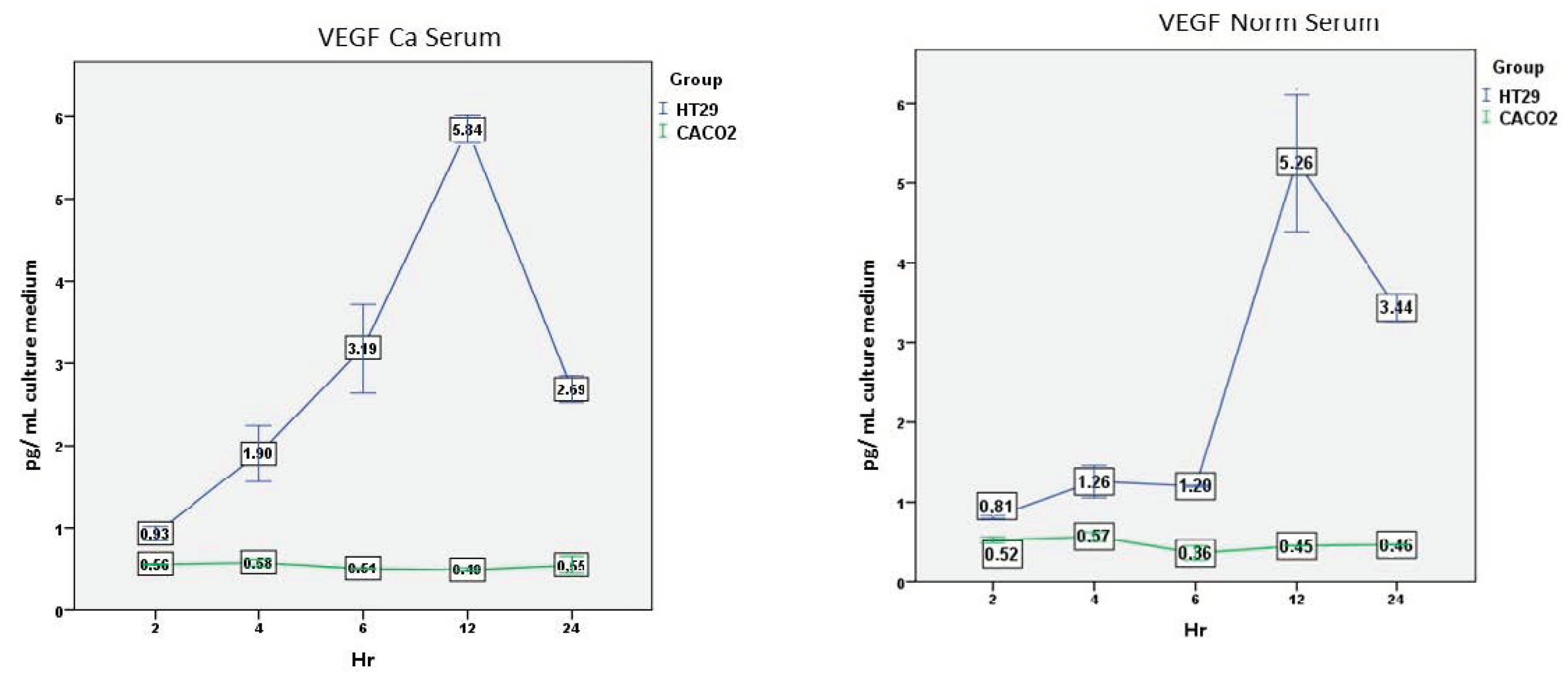
3.2.7. VEGF—Ca Serum
Overall, there was a significant effect of the cell line (F(1,2) = 1287.683, p = 0.001), the time (F(4,8) = 16.161, p = 0.001), and the interaction between time and cell line (F(4,8) = 17.331, p = 0.001) (Figure 6).
Secretion over time was similar to the CXCL8 secretion. HT-29 cells produced significantly higher levels compared to Caco-2 cells at all time points (p < 0.05), with a peak at 12 h. On the contrary, Caco-2 cells’ secretion was consistently low over time.
3.2.8. VEGF—Normal Serum
Overall, there was a significant effect of the cell line (F(1,2) = 1367.788, p = 0.007), the time (F(4,8) = 46.825, p = 0.001), and the interaction between time and cell line (F(4,8) = 48.448, p = 0.001).
HT-29 cells produced significantly higher levels compared to Caco-2 cells at all time points (p < 0.05), with a peak at 12 h. On the contrary, Caco-2 cells’ secretion was consistently low over time (Figure 6).
4. Discussion
The progress of colorectal carcinoma depends on various factors, both local and systemic. Among local factors, chemokine secretion is crucial. Chemokines are humoral agents with a dual action. They recruit immunocytes and behave as either angiogenic or angiostatic factors. Many cells, including macrophages and T lymphocytes, are able to produce chemokines. However, there is evidence that epithelial colonic enterocytes are also able to secrete C-X-C and C-C chemokines [22].
In this study, we attempted to answer four main questions: (1) Do HT-29 and Caco-2 cells produce angiodrastic cytokines in basal conditions? (2) Is there a difference between HT-29 and Caco-2 cells in response to whole serum from cancer patients compared to controls? (3) Is there a difference in the time course response between cancer patients and healthy individuals? (4) Is there a functional significance particularly when sera from cancer patients are compared to normal sera?
In a previous paper, we have demonstrated that colorectal cancer tissue contains increased amounts of the angiogenic factors CXCL6 and VEGF compared to normal adjacent tissue. More importantly, the five-year survival of patients was inversely correlated to increased levels of tissue CXCL8 and CXCL4 [3].
However, the biological behavior of colon cancer cells is not well-studied. Therefore, we studied HT-29 and Caco-2 cells in cultures because colonic epithelial cells have been found to be a major source of chemokine production [22]. We incubated these cell lines with sera from either colon cancer patients or healthy controls.
It has been reported that exposure of fibroblasts to whole serum results in changes in the expression of a large number of genes, many of which are known to be involved in wound healing. Genes induced in the fibroblast serum-response program are expressed in tumors (including colorectal cancer) by both the tumor cells themselves and the tumor-associated fibroblasts [23]. Moreover, the gene expression signature derived from serum-exposed fibroblasts in a subset of breast tumors has been found to correlate with poor clinical outcome, and may improve the risk stratification independent of currently employed clinico-pathological risk factors [24].
Since there is an interplay between different interleukins and chemokines, we decided to examine the net effect that the whole serum might have on well-characterized cell lines. HT-29 cells have been extensively used mostly because they express characteristics of mature intestinal cells. For this reason, HT-29 is considered a pluripotent intestinal cell line which can be used for the study of the structural and molecular events involved in cell differentiation [25]. Both cell lines secrete a variety of cytokines and growth factors, but secretion of chemokines has not been adequately studied [2,26].
4.1. Do HT-29 and Caco-2 Cells Produce Angiodrastic Cytokines in Basal Conditions?
Our results indicate that HT-29 cells under basic conditions (cultured in MEM) secreted considerable quantities of CXCL8, VEGF, CXCL4, and CXCL6. Secretion of CXCL8 in Caco-2 cells was similar to HT-29, but CXCL4 and CXCL6 secretion was significantly higher, while VEGF secretion was significantly lower.
There are conflicting and limited data on CXCL8 production, mostly by HT-29 cells. CXCL8 was detected in conditioned media from both unstimulated (1.1 ng/mL) and IL-1-beta-stimulated (16.1 ng/mL) HT-29 cultures [27]. Similarly, low levels of CXCL8 secretion by HT-29 cells were also reported in another study, while unstimulated Caco-2 cells were producing ten times lower levels of CXCL8 [28]. In contrast, no measurable levels of CXCL8 by HT-29 cells were reported in a subsequent study [29].
Very low levels of basal expression (mRNA) of CXCL8 and CXCL6 by HT-29 cells were recently reported, which considerably increased after incubation with IL-17 and TNF-a. However, no data on protein secretion were presented [30,31].
CXCL4 is secreted by activated T cells and monocytes, but data on CXCL4 secretion by HT-29 and Caco-2 cells do not exist [32].
Additionally, data on VEGF secretion in basal conditions are limited. Caco-2 cells express VEGF in basal conditions measured by real-time PCR. However, the well-known PGE2 target gene, VEGF, was not modulated in Caco-2 cells after exogenous PGE2 incubation [33].
An observation of our study that possibly explains the differences in the reported results is that basal secretion is dependent on the constituents of the culture medium. When fetal bovine serum was included in the culture medium, basal secretion was usually highly increased (data not shown). Therefore, direct comparisons can be made only when medium without the addition of bovine serum is used. This is the case of a report where HT-29 cells were shown to produce a considerable basal secretion of VEGF. The culture medium contained 10% fetal calf serum and levels were comparable to our results when FBS was used [26].
4.2. Are There Differences between HT-29 and Caco-2 Cells?
Differential chemokine production by colonic epithelial cells is thought to contribute to the characteristic increased infiltration of a selected population of cells in colorectal cancer. Colonic epithelial cell chemokine production can be differentially regulated by T cell-derived cytokines and suggests an interplay between epithelial cells and T lymphocytes that is potentially important in cancer. Thus, IL-13 enhances IL-1a-induced CXCL8 secretion by the colonic epithelial cell line HT-29, and the combination of TNF-a and IFN-γ causes secretion of C-C chemokines such as RANTES and MCP-1 via activation of a wortmannin-sensitive phosphatidylinositol 3-kinase [29].
Data on functional differences between the two cells lines are few. Expression of CXCL8 was different in cell lines with different metastatic potential. The KM12L4 cell line with a high metastatic potential expressed high levels of CXCL8, while cells without metastatic potential such as Caco-2 cells had the lowest expression [34].
We observed similar results in our study. HT-29 cells secreted significantly higher quantities of CXCL8 and VEGF at all time points compared with Caco-2 cells. The reason for this difference can only be speculative. All sera (whether normal or cancer) contain a mixture of cytokines that obviously activate HT-29 but not Caco-2 cells. This requires further investigation. Another explanation could be the degree of their methylation. Caco-2 cells have a higher percentage of methylation compared with the HT-29 cells, where a pattern of hypo-methylation is prevalent. This different pattern has been related to the expression of CXCL8. Cancer patients were found to express higher values of CXCL8 in cancer tissue in parallel with CXCL8 hypo-methylation [35].
Another possible explanation, at least for our VEGF results, could be the fact that HT-29 cells express COX-2 in contrast to Caco-2 cells. The VEGF gene is an established target for Prostaglandin E2 [33]. However, celecoxib, a selective COX-2 inhibitor, increased the VEGF levels in a COX-2-independent manner, activating ER stress [36].
The situation was exactly the opposite with CXCL6, and particularly with CXCL4. Caco-2 cells secreted higher levels of these two chemokines compared with HT-29 cells. This finding has not been reported before. Differential expression of chemokines by HT-29 and Caco-2 cells have been previously reported for C-C chemokines. In a study from our laboratory, HT-29 and Caco-2 cell lines showed a differential secretion of the C-C chemokines MIG, i-TAC, and IP-10 in response to activation by different cytokines. HT-29 cells secreted C-C chemokines upon incubation with IL-4, IL-10, and IL-13, but Caco-2 cells responded only when all three cytokines were acting simultaneously. Moreover, HT-29 cells constitutionally secreted IP-10 and i-TAC chemokines, while Caco-2 cells did not [37].
It is certain that differences between the two cell lines extend beyond the chemokine secretion. A very recent study identified a differential expression profiling comparing HT-29 and Caco-2 cells. These two cell lines differed in 1795 genes, 168 proteins, and 160 miRNAs, but their functional significance is unknown at present [38]. Additional functional differences between the two cell lines have been observed. TGF-b1 suppressed proliferation of HT-29 cells but had no effect on Caco-2 cells. In contrast to HT-29 cells, Caco-2 cells show a decreased expression of the two main TGF-b1 receptors, RI and RII [39]. A report from the same group on the effect of ciprofloxacin on proliferation and survival has shown that ciprofloxacin can suppress proliferation of HT-29 cells but has no effect on Caco-2 cells [40].
4.3. Is There a Difference in the Time Course Response between Cancer and Normal Serum Incubation?
In an earlier study, production of CXCL8 by Caco-2 and HT-29 cells was investigated. HT-29 cells secreted substantial amounts of CXCL8 if stimulated with IL-1-beta, TNF-alpha, or IFN-gamma, but Caco-2 cells responded only to IL-1-beta. Bacterial lipopolysaccharide (LPS) was also an efficient stimulus of CXCL8 release in HT-29 cells, whereas Caco-2 cells were completely unresponsive to LPS. CXCL8 secretion was greater at 4 h after stimulation [28].
In our time course experiments, CXCL8 production was constant over 24 h in Caco-2 cells, whether incubated with cancer serum or serum from healthy individuals. HT-29 cells, on the other hand, showed a highly significant increase protracted over 24 h of CXCL8 production when incubated with either cancer or normal serum. In accordance with the previous report, in this study, the increase for both sera was evident in 4 h, but the peak secretion was at 6 h for normal and at 12 h for cancer serum, while the decline was steeper after incubation with normal serum.
Similar results have been reported before, when HT-29 cells were incubated with a mixture of IFN-γ and TNF-a. A pattern similar to our incubation experiments with normal serum was found, with a peak at 4 h and a steep decline at 24 h [29].
Our VEGF results showed a similar picture in Caco-2 cells. They were completely unresponsive to incubation with either normal or cancer serum. Again, HT-29 cells responded to incubation with both sera, but there was an earlier response after incubation with cancer serum.
The situation was completely different for CXCL6, and particularly for CXCL4. Thus, CXLL6 production was always higher in Caco-2 cells and relatively constant over 24 h, irrespective of the serum used in incubation experiments. In contrast, HT-29 cells showed two secretory peaks at 4 and 12 h. CXCL4 secretion was also higher in Caco-2 cells over 24 h. Three peaks at 2, 6, and 24 h were identified when cells were incubated with cancer serum. Similar peaks were also found in HT-29 cells, but at a lower level of expression. Incubation with normal serum showed that HT-29 cells were rather unresponsive, while high peaks were again evident at 2, 6, and 24 h in Caco-2 cells.
4.4. Functional Significance of the Findings
We have demonstrated that colonic cancer cell lines secrete angiodrastic chemokines. HT-29 cells, but not Caco-2 cells, responded to incubation with sera by augmenting CXCL8 and VEGF production. CXCL8 and VEGF are angiogenic factors and their presence will therefore increase neo-angiogenesis and tumor progression [41].
Cell lines with a low metastatic potential, such as the Caco-2 cells, secrete very low levels of CXCL8 in contrast to cells with a higher metastatic potential, such as the HT-29 cells. Moreover, it has been shown that the addition of exogenous CXCL8 in cultures of colon cancer cell lines increases cell proliferation, thus indicating an autocrine trophic role for this chemokine contributing to the metastatic potential of cancer cells [34].
Furthermore, elevated levels of CXCR2 (a receptor for CXCL8) in the tumor microenvironment are associated with enhanced growth of human and murine colon cancer cells, increased peri-tumoral angiogenesis, and extravasation of cancer cells into the lungs and liver in vivo [42]. It should be noted, however, that colonic cells lose the expression of the CXCR2 receptor (one of the two receptors responsible for CXCL8 effects) as they are transformed into malignant cells. Thus, highly differentiated colon cancer cell lines such as the HT-29 and Caco-2 cells do not express CXCR2 receptors [8].
In accordance with the above findings, we have reported that high tumor levels of CXCL8 were significantly correlated with a reduced five-year survival of patients [3].
A recent paper elegantly describes an additional detrimental effect of CXCL8 that may also influence patient survival. CXCL8 derived from tumors contributes to the chemotactic recruitment of myeloid-derived suppressor cells (MDSC) and to their functional control. Monocytic but not granulocytic MDSC (MoMDSC) exert a suppressor activity on the proliferation of autologous T cells isolated from the circulation of cancer patients, and therefore create a pro-tumoral leukocyte microenvironment in cancer. MoMDSC were also attracted by CXCL8-containing culture supernatants of HT-29 colon carcinoma cells and by CXCL8-containing sera of patients [43].
Augmented secretion of VEGF from HT-29 cells, but not from Caco-2 cells, after incubation with cancer or normal serum (and FBS), may indicate that one of its functions is to act as a survival factor in colorectal cancer cells, demonstrating its distinct role in cancer cell survival, according to previous reports [44,45].
Thus, our findings may in part explain why Caco-2 cells have a limited metastatic potential as opposed to HT-29 cells. This is further supported by a recent report on the resistance of colonic cancer cell lines to the anti-tumoral agent bevacizumab. For the bevacizumab-sensitive DLD-1 tumor cells, treatment was accompanied by downregulation of the HIF–VEGF–VEGFR axis, whereas the same treatment resulted in activation of autocrine VEGF signaling in the bevacizumab-resistant HT-29 tumors. Furthermore, in the HT-29 tumors, bevacizumab treatment was associated with much stronger activation of autocrine VEGF signaling. These data indicate that intrinsic bevacizumab-resistance is clearly associated with the high levels of VEGF in the tumor environment that render the endothelial cells more resistant to bevacizumab [46]. A recent study adds a further explanation for the lack of metastatic potential of the Caco-2 cells as opposed to HT-29 cells. Inhibition of intracrine VEGF signaling strongly inhibited colorectal cancer cell migration and invasion by regulating proteins involved in cell motility [47].
The effects of these two pro-angiogenic factors are not independent. Hypoxia induces the production of VEGF through the HIF-1-alpha mediator. The hypoxic induction of VEGF in HIF-1-alpha knockdown DLD-1 colon cancer cells was only partially blocked. In addition to the preserved expression of VEGF, CXCL8 was induced by hypoxia in DLD-1 (HIF-kd) but not in DLD-1 (HIF-wt) cells. A neutralizing antibody to CXCL8 substantially inhibited angiogenesis and tumor growth in DLD-1 (HIF-kd) but not in DLD-1 (HIF-wt) xenografts, verifying the functional significance of this CXCL8 response. Thus, compensatory pathways can be activated to preserve the tumor angiogenic response [48].
Functional data that could be related to our findings on CXCL4 are scarce. We previously demonstrated that patients with increased CXCL4 levels in cancer tissue survived less often in five years compared to those with lower levels [3]. This might be contradictory to the known angiostatic effect of the platelet-produced CXCL4 chemokine. However, a new role for a platelet-unrelated CXCL4 has been described, supporting the development of colon cancer in vivo. The proliferation of T regulatory cells was enhanced, while the function of Treg was impaired, by CXCL4. Importantly, the newly identified pro-tumor effect of CXCL4 is connected to 5-FU chemotherapy, and hence, it might have potential clinical significance in oncology [49,50]. These data indicate that CXCL4 released from tumor cells (exemplified by Caco-2 and HT-29 cells of our study) reduced anti-tumor immunity, which may facilitate regrowth of the escaped colon cancer cells (after the surgical excision in our patients) and lead to tumor relapse and metastasis.
On the other hand, data on CXCL6 are even rarer. CXCL6 is a CXC chemokine that functionally uses both the CXCL8 receptors to chemo-attract neutrophils. We found that the pattern of its production is different from CXCL8 and VEGF and that Caco-2 cells produce more CXCL6 (and CXCL4) than HT-29 cells.
In accordance with our findings, different regulation of CXCL8 and CXCL6 secretion has been described in other cell systems. IL-1-beta was the predominant CXCL6 inducer in fibroblasts, chondrocytes, and endothelial cells, whereas CXCL8 was upregulated in these cells by TNF-alpha. In contrast, lipopolysaccharide (LPS) was a relatively better stimulus for CXCL6 compared to CXCL8 in fibroblasts. The kinetics of CXCL6 induction in fibroblasts differed from those of CXCL8. Freshly isolated peripheral blood mononuclear leukocytes, which are a good source of CXCL8, failed to produce CXCL6. Quantitatively, secretion of CXCL6 always remained inferior to that of CXCL8, as was also the case in our experiments.
Why this is happening and what the functional significance of this might be is currently not known [51].
5. Conclusions
In conclusion, we demonstrated that colonic epithelial cells are potent producers of angiodrastic chemokines. HT-29 and Caco-2 cells produced all four chemokines under basal conditions and over 24 h after incubation with either normal or cancer human serum. However, the production was not uniform for all chemokines. The secretion response of these two cell lines was completely different. HT-29 cells produced more CXCL8 and VEGF irrespective of serum incubation, while Caco-2 cells were relatively unresponsive. The opposite was true for CXCL6 and CXCL4. Moreover, HT-29 cells produced more CXCL8 and VEGF over 24 h when incubated with cancer serum. On the other hand, Caco-2 cells produced more CXCL4 when incubated with cancer serum.
Author Contributions
E.G., M.A., T.M., V.A. and K.E. have all directly participated in the planning, execution, or analysis of this study. E.G. conducted all experiments and measurements mentioned in this paper. The contents of this manuscript have not been previously published. The contents of this manuscript are not now under consideration for publication elsewhere. There are no directly related manuscripts or abstracts published by any authors of this paper. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki and according to Institutional Review Board of University Hospital of Heraklion there was no need for ethical approval, since this was an in vitro study, not a clinical one, and no demographic, clinical or laboratory details of any patients were used.
Informed Consent Statement
Patient consent was waived since sera were randomly and anonymously selected without any use of demographic, clinical, or laboratory data from a large sera collection and were used only for in vitro cultures without clinical details. The Ethics Committee of the University Hospital felt that there was no need for further consent.
Data Availability Statement
The data presented in this study are available upon request from the corresponding author. The data are not publicly available due to privacy (they are part of the Ph.D. thesis of E.G.).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Herszenyi, L.; Tulassay, Z. Epidemiology of gastrointestinal and liver tumors. Eur. Rev. Med. Pharmacol. Sci. 2010, 14, 249–258. [Google Scholar] [PubMed]
- Hemmings, C. Is carcinoma a mesenchymal disease? The role of the stromal microenvironment in carcinogenesis. Pathology 2013, 45, 371–381. [Google Scholar] [CrossRef] [PubMed]
- Emmanouil, G.; Ayiomamitis, G.; Zizi-Sermpetzoglou, A.; Tzardi, M.; Moursellas, A.; Voymvouraki, A.; Kouroumalis, E. Angiodrastic Chemokines in Colorectal Cancer: Clinicopathological Correlations. Anal. Cell. Pathol. 2018, 2018, 1616973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fogh, J.; Trempe, G. New human tumor cell lines. In Human Tumor Cell In Vitro, 1st ed.; Fogh, J., Ed.; Springer: New York, NY, USA, 1975; pp. 115–159. [Google Scholar] [CrossRef]
- Fogh, J.; Wright, J.C.; Loveless, J.D. Absence of HeLa cell contamination in 169 cell lines derived from human tumours. J. Natl. Cancer Inst. 1977, 58, 209–214. [Google Scholar] [CrossRef]
- Strieter, R.M.; Polverini, P.J.; Kunkel, S.L.; Arenberg, D.A.; Burdick, M.D.; Kasper, J.; Dzuiba, J.; van Damme, J.; Walz, A.; Marriott, D.; et al. The functional role of the ELR motif in CXC chemokine-mediated angiogenesis. J. Biol. Chem. 1995, 270, 27348–27357. [Google Scholar] [CrossRef] [Green Version]
- Bonecchi, R.; Galliera, E.; Borroni, E.M.; Corsi, M.M.; Locati, M.; Mantovani, A. Chemokines and chemokine receptors: An overview. Front. Biosci. (Landmark Ed.) 2009, 14, 540–551. [Google Scholar] [CrossRef] [Green Version]
- Sturm, A.; Baumgart, D.C.; d’Heureuse, J.H.; Hotz, A.; Wiedenmann, B.; Dignass, A.U. CXCL8 modulates human intestinal epithelial cells through a CXCR1 dependent pathway. Cytokine 2005, 29, 42–48. [Google Scholar] [CrossRef]
- Cui, G.; Yuan, A.; Goll, R.; Vonen, B.; Florholmen, J. Dynamic changes of interleukin-8 network along the colorectal adenoma-carcinoma sequence. Cancer Immunol. Immunother. 2009, 58, 1897–1905. [Google Scholar] [CrossRef]
- Ning, Y.; Manegold, P.C.; Hong, Y.K.; Zhang, W.; Pohl, A.; Lurje, G.; Winder, T.; Yang, D.; LaBonte, M.J.; Wilson, M.L.; et al. Interleukin-8 is associated with proliferation, migration, angiogenesis and chemosensitivity in vitro and in vivo in colon cancer cell line models. Int. J. Cancer 2010, 128, 2038–2049. [Google Scholar] [CrossRef] [Green Version]
- Strieter, R.M.; Polverini, P.J.; Arenberg, D.; Kunkel, S.L. The role of CXC chemokines as regulators of angiogenesis. Shock 1995, 4, 155–160. [Google Scholar] [CrossRef]
- Vandercappellen, J.; van Damme, J.; Struyf, S. The role of CXC chemokines and their receptors in cancer. Cancer Lett. 2008, 267, 226–244. [Google Scholar] [CrossRef] [PubMed]
- Wuyts, A.; van Osselaer, N.; Haelens, A.; Samson, I.; Herdewijn, P.; Ben-Baruch, A.; Oppenheim, J.J.; Proost, P.; van Damme, J. Characterization of synthetic human granulocyte chemotactic protein 2: Usage of chemokine receptors CXCR1 and CXCR2 and in vivo inflammatory properties. Biochemistry 1997, 36, 2716–2723. [Google Scholar] [CrossRef] [PubMed]
- Heidemann, J.; Ogawa, H.; Dwinell, M.B.; Rafiee, P.; Maaser, C.; Gockel, H.R.; Otterson, M.F.; Ota, D.M.; Lugering, N.; Domschke, W.; et al. Angiogenic effects of interleukin 8 (CXCL8) in human intestinal microvascular endothelial cells are mediated by CXCR2. J. Biol. Chem. 2003, 278, 8508–8515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maione, T.E.; Gray, G.S.; Petro, J.; Hunt, A.J.; Donner, A.L.; Bauer, S.I.; Carson, H.F.; Sharpe, R.J. Inhibition of angiogenesis by recombinant human platelet factor-4 and related peptides. Science 1990, 247, 77–79. [Google Scholar] [CrossRef]
- Gupta, S.K.; Singh, J.P. Inhibition of endothelial cell proliferation by platelet factor-4 involves a unique action on S phase progression. J. Cell Biol. 1994, 127, 1121–1127. [Google Scholar] [CrossRef]
- Shellenberger, T.D.; Wang, M.; Gujrati, M.; Jayakumar, A.; Strieter, R.M.; Burdick, M.D.; Ioannides, C.G.; Efferson, C.L.; El-Naggar, A.K.; Roberts, D.; et al. BRAK/CXCL14 is a potent inhibitor of angiogenesis and a chemotactic factor for immature dendritic cells. Cancer Res. 2004, 64, 8262–8270. [Google Scholar] [CrossRef] [Green Version]
- Struyf, S.; Burdick, M.D.; Proost, P.; van Damme, J.; Strieter, R.M. Platelets release CXCL14L1, a nonallelic variant of the chemokine platelet factor-4/CXCL4 and potent inhibitor of angiogenesis. Circ. Res. 2004, 95, 855–857. [Google Scholar] [CrossRef]
- Kowanetz, M.; Ferrara, N. Vascular endothelial growth factor signaling pathways: Therapeutic perspective. Clin. Cancer Res. 2006, 12, 5018–5022. [Google Scholar] [CrossRef] [Green Version]
- Aldebasi, Y.; Rahmani, A.; Khan, A.; Aly, S. The effect of vascular endothelial growth factor in the progression of bladder cancer and diabetic retinopathy. Int. J. Clin. Exp. Med. 2013, 6, 239–251. [Google Scholar]
- Tchaikovski, V.; Fellbrich, G.; Waltenberger, J. The molecular basis of VEGFR-1 signal transduction pathways in primary human monocytes. Arter. Thromb. Vasc. Biol. 2008, 28, 322–328. [Google Scholar] [CrossRef] [Green Version]
- Kucharzik, T.; Hudson, J.T., III; Lugering, A.; Abbas, J.A.; Bettini, M.; Lake, J.G.; Evans, M.E.; Ziegler, T.R.; Merlin, D.; Madara, J.L.; et al. Acute induction of human IL-8 production by intestinal epithelium triggers neutrophil infiltration without mucosal injury. Gut 2005, 54, 1565–1572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, H.Y.; Sneddon, J.B.; Alizadeh, A.A.; Sood, R.; West, R.B.; Montgomery, K.; Chi, J.-T.; van De Rijn, M.; Botstein, D.; O Brown, P. Gene expression signature of fibroblast serum response predicts human cancer progression: Similarities between tumors and wounds. PLoS Biol. 2004, 2, e7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, H.Y.; Nuyten, D.S.A.; Sneddon, J.B.; Hastie, T.; Tibshirani, R.; Sørlie, T.; Dai, H.; He, Y.D.; Veer, L.J.V.; Bartelink, H.; et al. Robustness, scalability, and integration of a wound-response gene expression signature in predicting breast cancer survival. Proc. Natl. Acad. Sci. USA 2005, 102, 3738–3743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huet, C.; Sahuquillo-Merino, C.; Coudrier, E.; Louvard, D. Absorptive and mucus-secreting subclones isolated from a multipotent intestinal cell line (HT-29) provide new models for cell polarity and terminal differentiation. J. Cell Biol. 1987, 105, 345–357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desai, S.; Kumar, A.; Laskar, S.; Pandey, B.N. Cytokine profile of conditioned medium from human tumor cell lines after acute and fractionated doses of gamma radiation and its effect on survival of bystander tumor cells. Cytokine 2013, 61, 54–62. [Google Scholar] [CrossRef] [PubMed]
- Kelly, C.P.; Keates, S.; Siegenberg, D.; Linevsky, J.K.; Pothoulakis, C.; Brady, H.R. IL-8 secretion and neutrophil activation by HT-29 colonic epithelial cells. Am. J. Physiol.-Gastrointest. Liver Physiol. 1994, 267, G991–G997. [Google Scholar] [CrossRef]
- Schuerer-Maly, C.C.; Eckmann, L.; Kagnoff, M.F.; Falco, M.T.; Maly, F.E. Colonic epithelial cell lines as a source of interleukin-8: Stimulation by inflammatory cytokines and bacterial lipopolysaccharide. Immunology 1994, 81, 85–91. [Google Scholar]
- Kolios, G.; Wright, K.L.; Jordan, N.J.; Leithead, J.B.; Robertson, D.A.; Westwick, J. C-X-C and C-C chemokine expression and secretion by the human colonic epithelial cell line, HT-29: Differential effect of T lymphocyte-derived cytokines. Eur. J. Immunol. 1999, 29, 530–536. [Google Scholar] [CrossRef]
- Wang, Y.L.; Fang, M.; Wang, X.M.; Liu, W.Y.; Zheng, Y.J.; Wu, X.B.; Tao, R. Proinflammatory effects and molecular mechanisms of interleukin-17 in intestinal epithelial cell line HT-29. World J. Gastroenterol. 2014, 20, 17924–17931. [Google Scholar] [CrossRef]
- Zhang, M.; Wang, G.; Tao, Y.; Zhang, H. The proinflammatory effect and molecular mechanism of IL-17 in the intestinal epithelial cell line HT-29. J. BUON 2015, 20, 120–127. [Google Scholar]
- Schaffner, P.; Rhyn, G.; Schoedon, D.J. Schaer, Regulated expression of platelet factor 4 in human monocytes-role of PARs as a quantitatively important monocyte activation pathway. J. Leukoc. Biol. 2005, 78, 202–209. [Google Scholar] [CrossRef] [PubMed]
- Mauritz, I.; Westermayer, S.; Marian, B.; Erlach, N.; Grusch, M.; Holzmann, K. Prostaglandin E(2) stimulates progression-related gene expression in early colorectal adenoma cells. Br. J. Cancer 2006, 94, 1718–1725. [Google Scholar] [CrossRef] [PubMed]
- Li, A.; Varney, M.L.; Singh, R.K. Expression of Interleukin 8 and Its Receptors in Human Colon Carcinoma Cells with Different Metastatic Potentials. Clin. Cancer Res. 2001, 7, 3298–3304. [Google Scholar] [PubMed]
- Dimberg, J.; Ström, K.; Löfgren, S.; Zar, N.; Lindh, M.; Matussek, A. DNA promoter methylation status and protein expression of interleukin-8 in human colorectal adenocarcinomas. Int. J. Color. Dis. 2011, 27, 709–714. [Google Scholar] [CrossRef] [Green Version]
- Xu, B.; Wang, Y.; Yang, J.; Zhang, Z.; Zhang, Y.; Du, H. Celecoxib induces apoptosis but up-regulates VEGF via endoplasmic reticulum stress in human colorectal cancer in vitro and in vivo. Cancer Chemother. Pharmacol. 2016, 77, 797–806. [Google Scholar] [CrossRef]
- Manousou, P.; Kolios, G.; Drygiannakis, I.; Pyrovolaki, K.; Bourikas, L.; Papadaki, H.A.; Kouroumalis, E. Expression of a splice variant of CXCR3 in Crohn’s disease patients; Indication for a lymphocyte—Epithelial cell interaction. J. Gastroenterol. Hepatol. 2008, 23, 1823–1833. [Google Scholar] [CrossRef]
- O’Sullivan, F.; Keenan, J.; Aherne, S.; O’Neill, F.; Clarke, C.; Henry, M.; Meleady, P.; Breen, L.; Barron, N.; Clynes, M.; et al. Parallel mRNA, proteomics and miRNA expression analysis in cell line models of the intestine. World J. Gastroenterol. 2017, 23, 7369–7386. [Google Scholar] [CrossRef]
- Biasi, F.; Tessitore, L.; Zanetti, D.; Cutrin, J.C.; Zingaro, B.; Chiarpotto, E.; Zarkovic, N.; Serviddio, G.; Poli, G. Associated changes of lipid peroxidation and transforming growth factor beta1 levels in human colon cancer during tumour progression. Gut 2002, 50, 361–367. [Google Scholar] [CrossRef] [Green Version]
- Zanetti, D.; Poli, G.; Vizio, B.; Zingaro, B.; Chiarpotto, E.; Biasi, F. 4-hydroxynonenal and transforming growth factor-beta1 expression in colon cancer. Mol. Asp. Med. 2003, 24, 273–280. [Google Scholar] [CrossRef]
- Brew, R.; Erikson, J.S.; West, D.C.; Kinsella, R.A.; Slavin, J.; Christmas, S.E. Interleukin-8 as an autocrine growth factor for human colon carcinoma cells in vitro. Cytokine 2000, 12, 78–85. [Google Scholar] [CrossRef]
- Lee, Y.S.; Choi, I.; Ning, Y.; Kim, N.Y.; Khatchadourian, V.; Yang, D.; Chung, H.K.; Choi, D.; LaBonte, M.J.; Ladner, R.D.; et al. Interleukin-8 and its receptor CXCR2 in the tumour microenvironment promote colon cancer growth, progression and metastasis. Br. J. Cancer 2012, 106, 1833–1841. [Google Scholar] [CrossRef] [PubMed]
- Alfaro, C.; Teijeira, A.; Oñate, C.; Pérez, G.; Sanmamed, M.F.; Andueza, M.-P.; Alignani, D.; Labiano, S.; Azpilikueta, A.; Rodriguez-Paulete, A.; et al. Tumor-Produced Interleukin-8 Attracts Human Myeloid-Derived Suppressor Cells and Elicits Extrusion of Neutrophil Extracellular Traps (NETs). Clin. Cancer Res. 2016, 22, 3924–3936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhattacharya, R.; Ye, X.C.; Wang, R.; Ling, X.; McManus, M.; Fan, F.; Boulbes, D.; Ellis, L.M. Intracrine VEGF Signaling Mediates the Activity of Prosurvival Pathways in Human Colorectal Cancer Cells. Cancer Res. 2016, 76, 3014–3024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samuel, S.; Fan, F.; Dang, L.H.; Xia, L.; Gaur, P.; Ellis, L.M. Intracrine vascular endothelial growth factor signaling in survival and chemoresistance of human colorectal cancer cells. Oncogene 2010, 30, 1205–1212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mésange, P.; Poindessous, V.; Sabbah, M.; Escargueil, A.E.; de Gramont, A.; Larsen, A.K. Intrinsic bevacizumab resistance is associated with prolonged activation of autocrine VEGF signaling and hypoxia tolerance in colorectal cancer cells and can be overcome bynintedanib, a small molecule angiokinase inhibitor. Oncotarget 2014, 5, 4709–4721. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharya, R.; Fan, F.; Wang, R.; Ye, X.; Xia, L.; Boulbes, D.; Ellis, L.M. Intracrine VEGF signalling mediates colorectal cancer cell migration and invasion. Br. J. Cancer 2017, 117, 848–855. [Google Scholar] [CrossRef] [Green Version]
- Mizukami, Y.; Jo, W.-S.; Duerr, E.-M.; Gala, M.; Li, J.; Zhang, X.; Zimmer, M.A.; Iliopoulos, O.; Zukerberg, L.R.; Kohgo, Y.; et al. Induction of interleukin-8 preserves the angiogenic response in HIF-1alpha-deficient colon cancer cells. Nat. Med. 2005, 11, 992–997. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Gao, J.; Wang, X.; Deng, S.; Ye, H.; Guan, W.; Wu, M.; Zhu, S.; Yu, Y.; Han, W. CXCL4 mediates tumor regrowth after chemotherapy by suppression of antitumor immunity. Cancer Biol. Ther. 2015, 16, 1775–1783. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.Y.; Battaglia, M.; Lee, S.H.; Sun, Q.H.; Aster, R.H.; Visentin, G.P. Platelet factor 4 differentially modulates CD4 + CD25 + (regulatory) versus CD4 + CD25-(nonregulatory) T cells. J. Immunol. 2005, 174, 2680–2686. [Google Scholar] [CrossRef] [Green Version]
- Wuyts, A.; Struyf, S.; Gijsbers, K.; Schutyser, E.; Put, W.; Conings, R.; Lenaerts, J.-P.; Geboes, K.; Opdenakker, G.; Menten, P.; et al. The CXC chemokine GCP-2/CXCL6 is predominantly induced in mesenchymal cells by interleukin-1β and is down-regulated by interferon-γ: Comparison with interleukin-8/CXCL8. Lab. Investig. 2003, 83, 23–34. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Secretion of angiodrastic chemokines by HT-29 cells over time. Standard error bars have been omitted for clarity.
Figure 1.
Secretion of angiodrastic chemokines by HT-29 cells over time. Standard error bars have been omitted for clarity.

Figure 2.
Secretion of angiodrastic chemokines by Caco-2 cells over time. Standard error bars have been omitted for clarity.
Figure 2.
Secretion of angiodrastic chemokines by Caco-2 cells over time. Standard error bars have been omitted for clarity.

Figure 3.
Comparison of CXCL8 secretion over time by the two cell lines after incubation with cancer or normal serum. Mean and standard error (vertical bars) of five separate experiments are presented.
Figure 3.
Comparison of CXCL8 secretion over time by the two cell lines after incubation with cancer or normal serum. Mean and standard error (vertical bars) of five separate experiments are presented.

Figure 4.
Comparison of CXCL6 secretion over time by the two cell lines after incubation with cancer or normal serum. Mean and standard error (vertical bars) of five separate experiments are presented. For statistics, see the text.
Figure 4.
Comparison of CXCL6 secretion over time by the two cell lines after incubation with cancer or normal serum. Mean and standard error (vertical bars) of five separate experiments are presented. For statistics, see the text.

Figure 5.
Comparison of CXCL4 secretion over time by the two cell lines after incubation with cancer or normal serum. Mean and standard error (vertical bars) of five separate experiments are presented. For statistics, see the text.
Figure 5.
Comparison of CXCL4 secretion over time by the two cell lines after incubation with cancer or normal serum. Mean and standard error (vertical bars) of five separate experiments are presented. For statistics, see the text.

Figure 6.
Comparison of VEGF secretion over time by the two cell lines after incubation with cancer or normal serum. Mean and standard error (vertical bars) of five separate experiments are presented. For statistics, see the text.
Figure 6.
Comparison of VEGF secretion over time by the two cell lines after incubation with cancer or normal serum. Mean and standard error (vertical bars) of five separate experiments are presented. For statistics, see the text.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
p-Value differences between base values (unstimulated medium) and both cancer and normal serum at 2, 4, and 6 h of incubation.
Table 1.
p-Value differences between base values (unstimulated medium) and both cancer and normal serum at 2, 4, and 6 h of incubation.
| Medium and Time | Type of Stimulation | p-Value |
|---|---|---|
| McCoy’s 2 h | Ca_Serum | 0.014 |
| Nor_Serum | 0.004 | |
| McCoy’s 4 h | Ca_Serum | 0.019 |
| Nor_Serum | 0.023 | |
| McCoy’s 6 h | Ca_Serum | 0.000 |
| Nor_Serum | 0.000 |
Bold marks statistical significance as p < 0.005.
Table 2.
Statistically significant differences between base values (medium) and both cancer and normal serum at all time points (p < 0.05).
Table 2.
Statistically significant differences between base values (medium) and both cancer and normal serum at all time points (p < 0.05).
| Medium and Time | Type of Stimulation | Mean Difference | p-Value |
|---|---|---|---|
| McCoy’s 2 h | Ca_Serum | −28.729202 * | 0.001 |
| Nor_Serum | −290.058917 * | 0.001 | |
| McCoy’s 4 h | Ca_Serum | −23.142217 * | 0.001 |
| Nor_Serum | −26.488188 * | 0.001 | |
| McCoy’s 6 h | Ca_Serum | −8.049175 | 0.213 |
| Nor_Serum | −3.654608 | 0.710 | |
| McCoy’s 12 h | Ca_Serum | −25.851839 * | 0.002 |
| Nor_Serum | −24.495701 * | 0.003 | |
| acCoy’s 24 h | Ca_Serum | −28.196170 * | 0.004 |
| Nor_Serum | −23.976026 * | 0.007 |
Bold marks statistical significance as p < 0.005; * marks statistical significant differences as demonstrated by the respective p value.
Table 3.
Difference between basal and sera incubations at 24 h and cancer and normal serum incubations at 24 h.
Table 3.
Difference between basal and sera incubations at 24 h and cancer and normal serum incubations at 24 h.
| Medium and Time | Type of Stimulation | Mean Difference | p-Value |
|---|---|---|---|
| McCoy’s 24 h | Ca_Serum | −1.350560 * | 0.001 |
| Nor_Serum | −2.100735 * | 0.000 | |
| Ca_Serum 24 h | Nor_Serum | −0.750176 * | 0.007 |
| MacCoy’s | 1.350560 * | 0.001 |
Bold marks statistical significance as p < 0.005; * marks statistical significant differences as demonstrated by the respective p value.
Table 4.
Cancer serum incubation significantly increased CXCL8 secretion at 4 h compared with either MEM or normal serum incubation, while CXCL8 secretion was significantly lower at 6 h under MEM incubation.
Table 4.
Cancer serum incubation significantly increased CXCL8 secretion at 4 h compared with either MEM or normal serum incubation, while CXCL8 secretion was significantly lower at 6 h under MEM incubation.
| Medium and Time | Type of Stimulation | Mean Difference | p-Value |
|---|---|---|---|
| Ca_Serum 4 h | Nor_Serum | 2.945237 * | 0.044 |
| MEM | 3.498874 * | 0.025 | |
| MEM 6 h | Ca_Serum | −2.963012 * | 0.034 |
| Nor_Serum | −5.099393 * | 0.005 |
Bold marks statistical significance as p < 0.005; * marks statistical significant differences as demonstrated by the respective p value.
Table 5.
Cancer serum produced significantly more CXCL4 compared to normal serum at 4 h (p < 0.008), and 6 h cancer and normal sera incubation produced significantly higher secretion compared to basal conditions.
Table 5.
Cancer serum produced significantly more CXCL4 compared to normal serum at 4 h (p < 0.008), and 6 h cancer and normal sera incubation produced significantly higher secretion compared to basal conditions.
| Medium and Time | Type of Stimulation | Mean Difference | p-Value |
|---|---|---|---|
| MEM 4 h | Ca_Serum | −40.393690 * | 0.000 |
| Nor_Serum | −25.335775 * | 0.001 | |
| MEM 6 h | Ca_Serum | −33.842377 * | 0.000 |
| Nor_Serum | −32.923005 * | 0.000 |
Bold marks statistical significance as p < 0.005; * marks statistical significant differences as demonstrated by the respective p value.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
George, E.; Andrew, M.; Maria, T.; Argyro, V.; Elias, K. Angiodrastic Chemokines Production by Colonic Cancer Cell Lines. Onco 2022, 2, 69-84. https://doi.org/10.3390/onco2020006
AMA Style
George E, Andrew M, Maria T, Argyro V, Elias K. Angiodrastic Chemokines Production by Colonic Cancer Cell Lines. Onco. 2022; 2(2):69-84. https://doi.org/10.3390/onco2020006
Chicago/Turabian StyleGeorge, Emmanouil, Moursellas Andrew, Tzardi Maria, Voumvouraki Argyro, and Kouroumalis Elias. 2022. "Angiodrastic Chemokines Production by Colonic Cancer Cell Lines" Onco 2, no. 2: 69-84. https://doi.org/10.3390/onco2020006