
Mechanistic Modelling of DNA Damage Repair by the Radiation Adaptive Response Mechanism and Its Significance

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Modelling of Radiation’s Influence on Cells
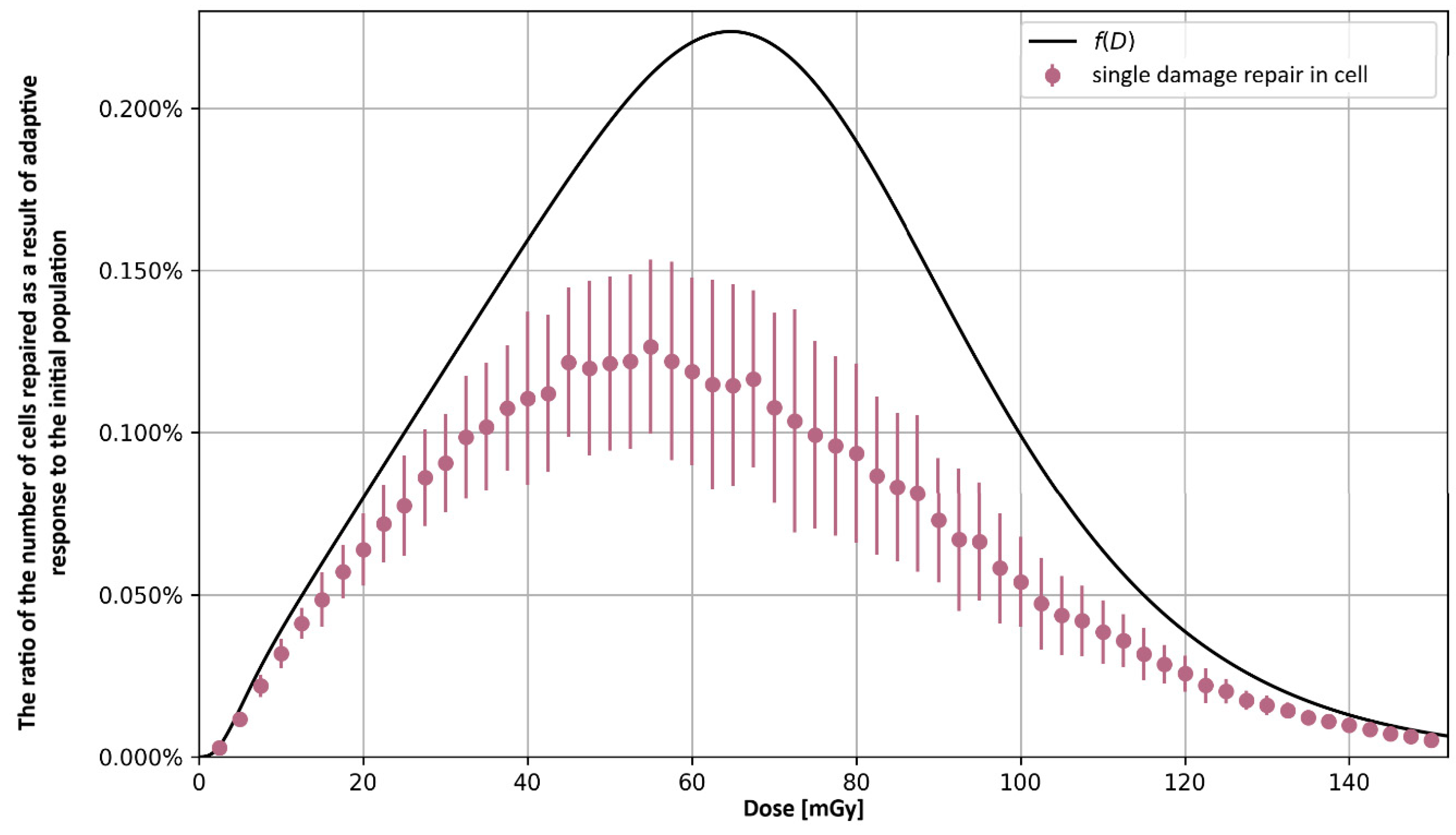
3. Radiation Adaptive Response—Enhancement of the Repair Process
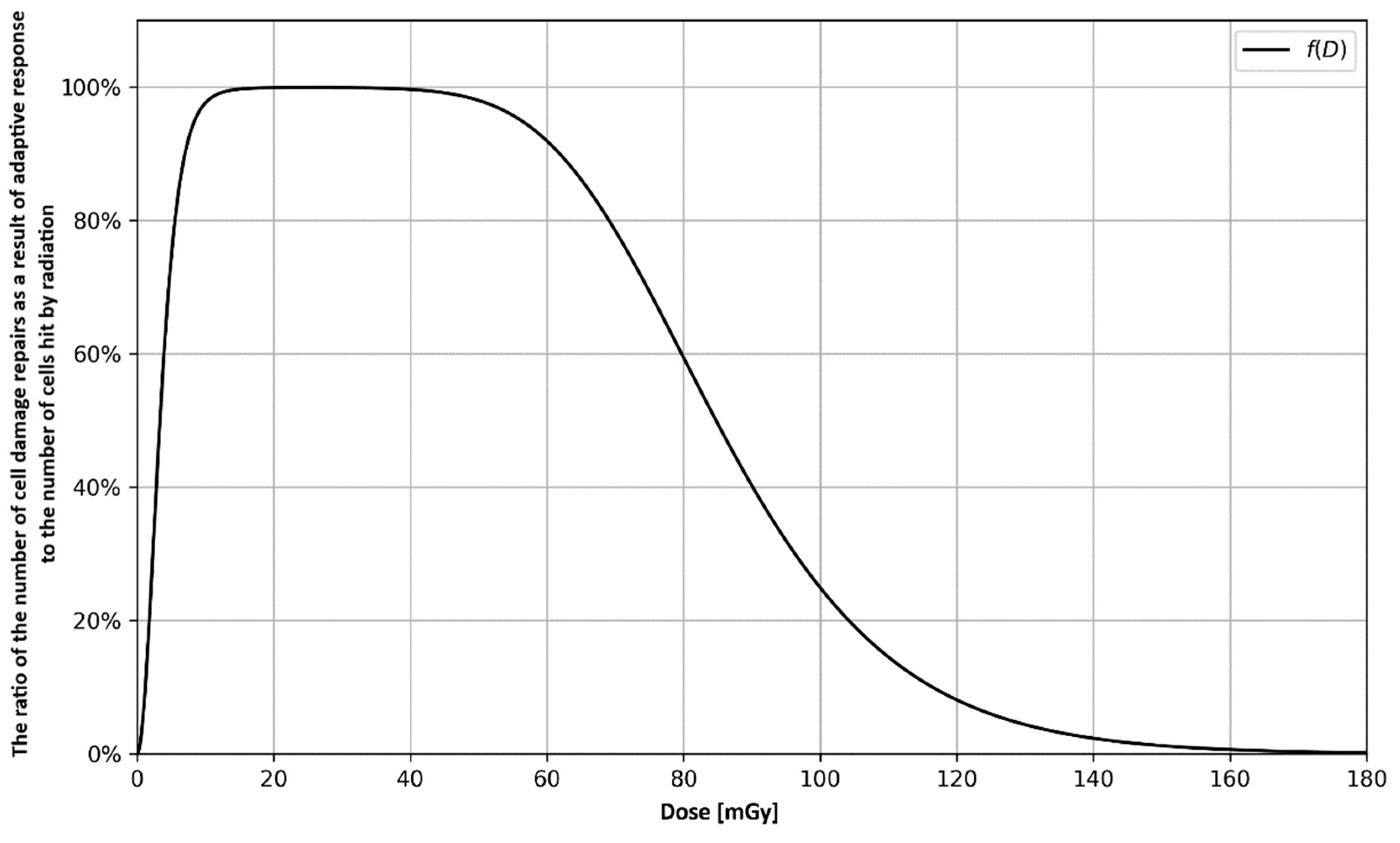
3.1. Analytical Approach
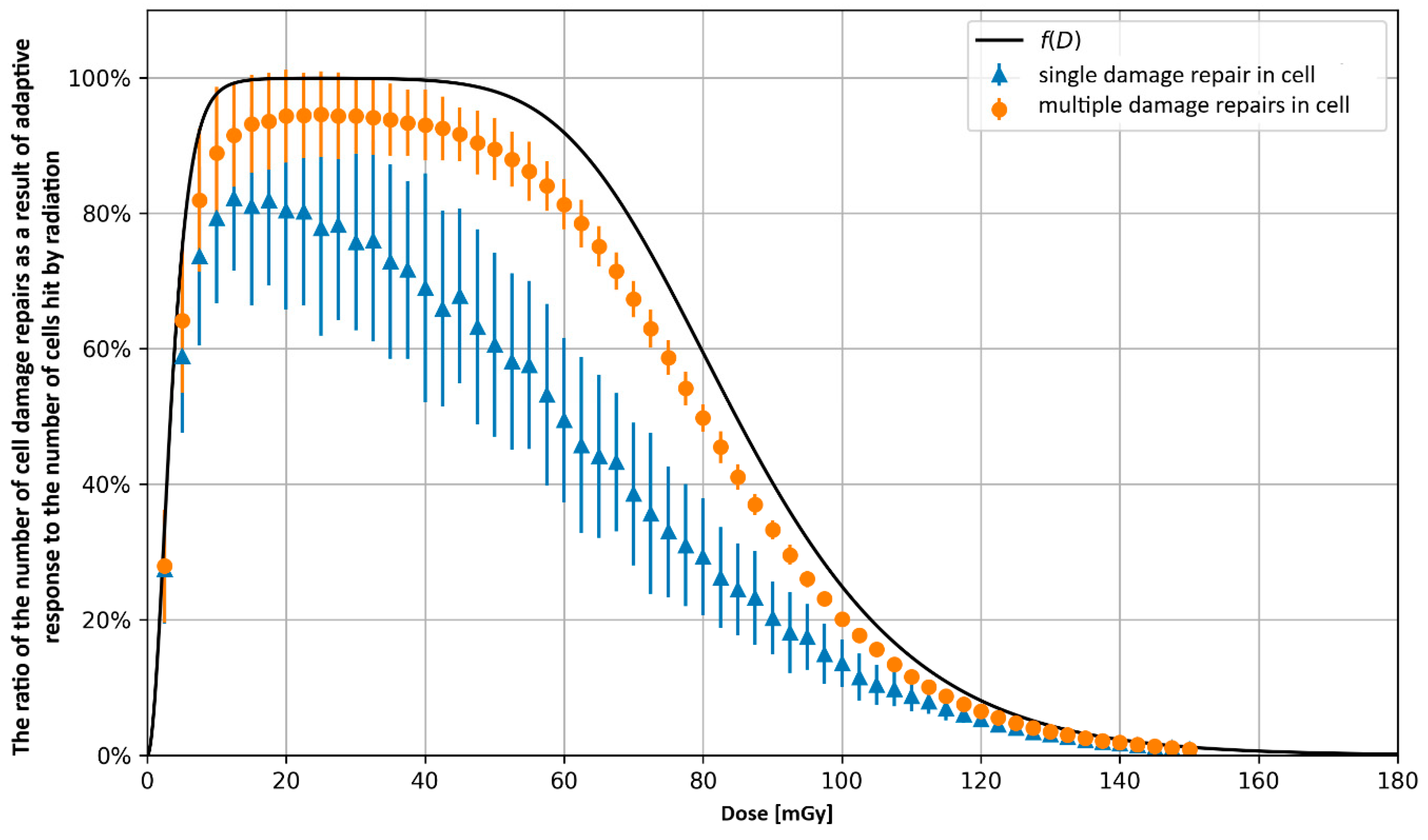
3.2. Monte Carlo Approach—Single Dose
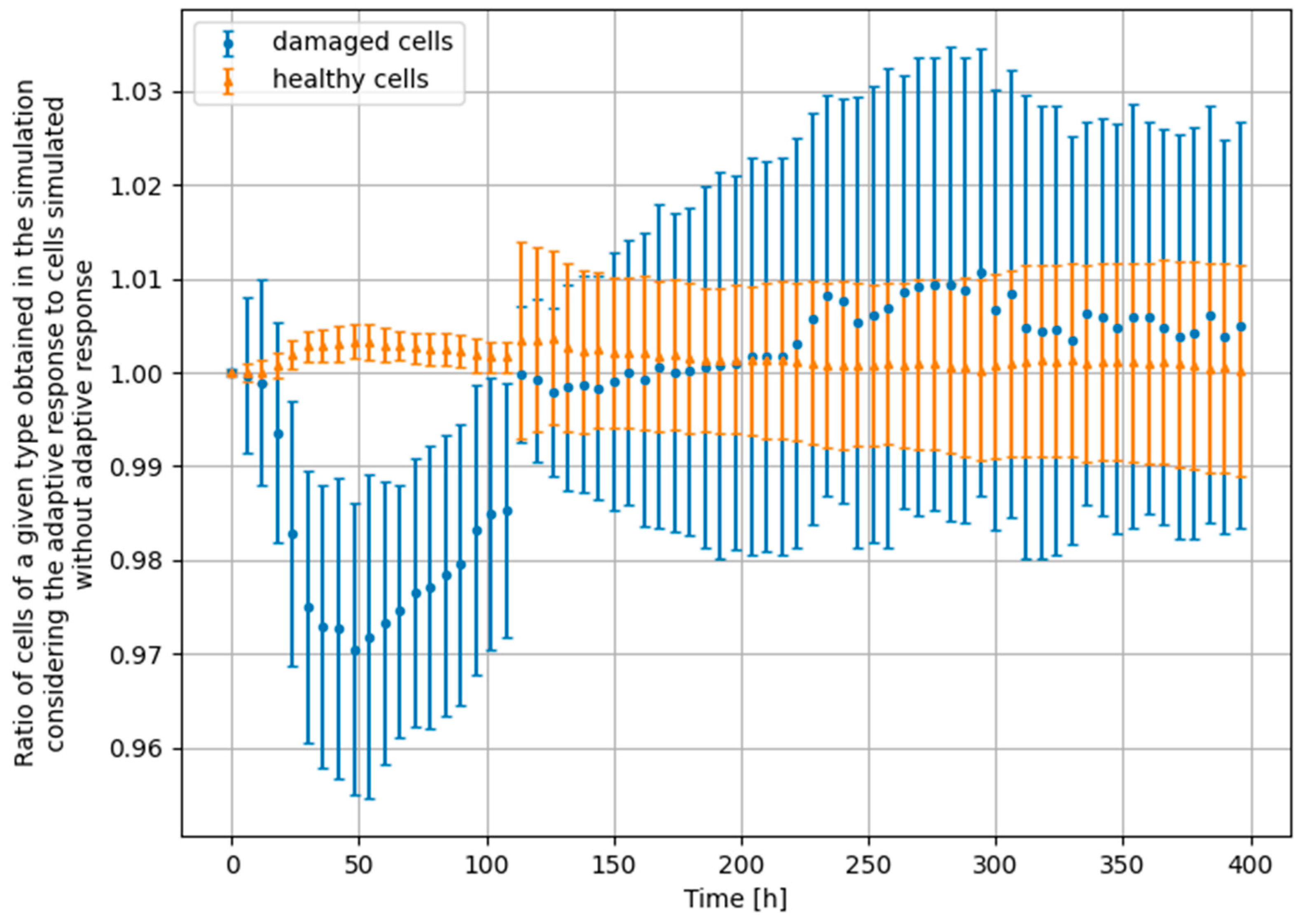
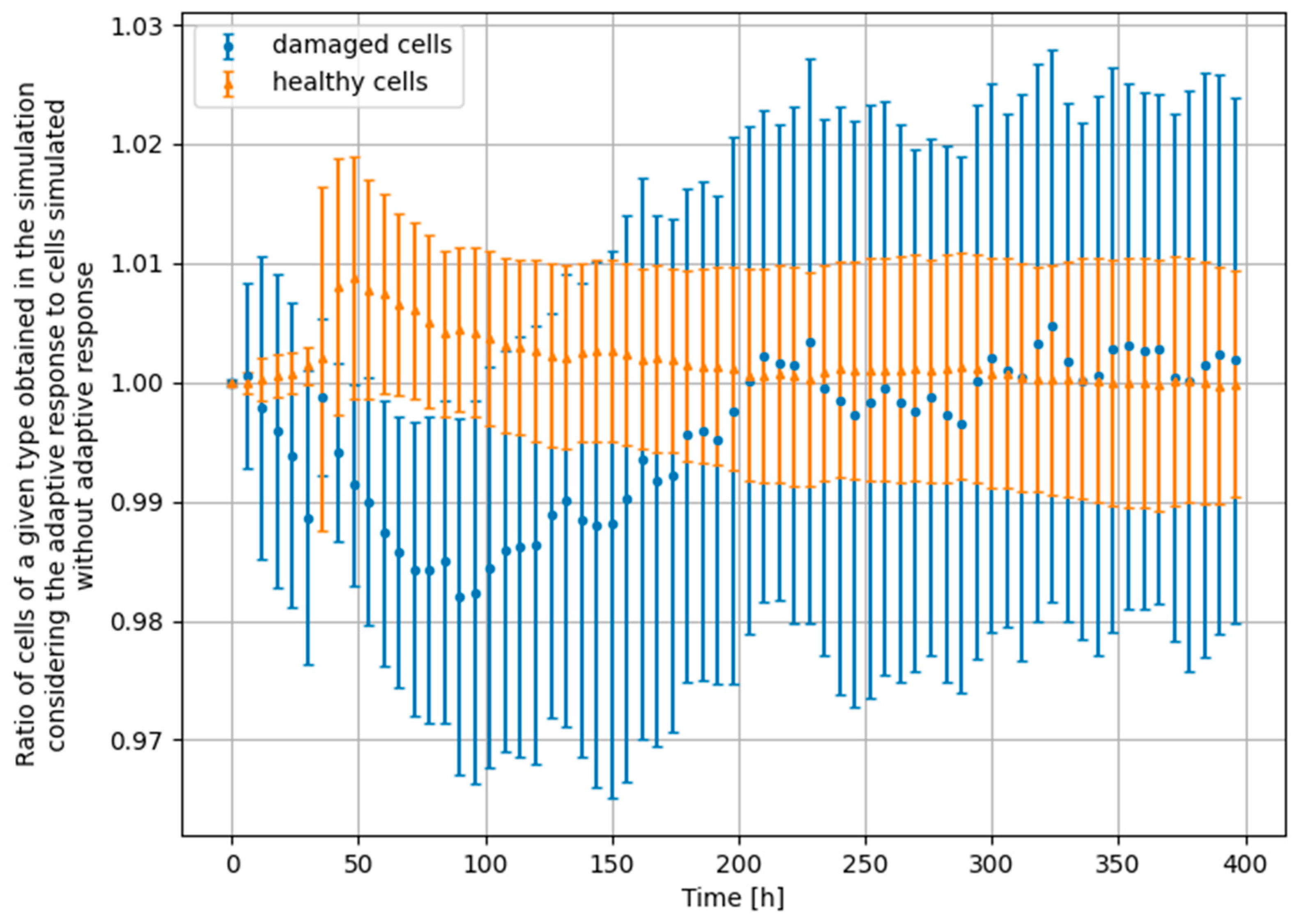
3.3. Monte Carlo Approach—Two Doses
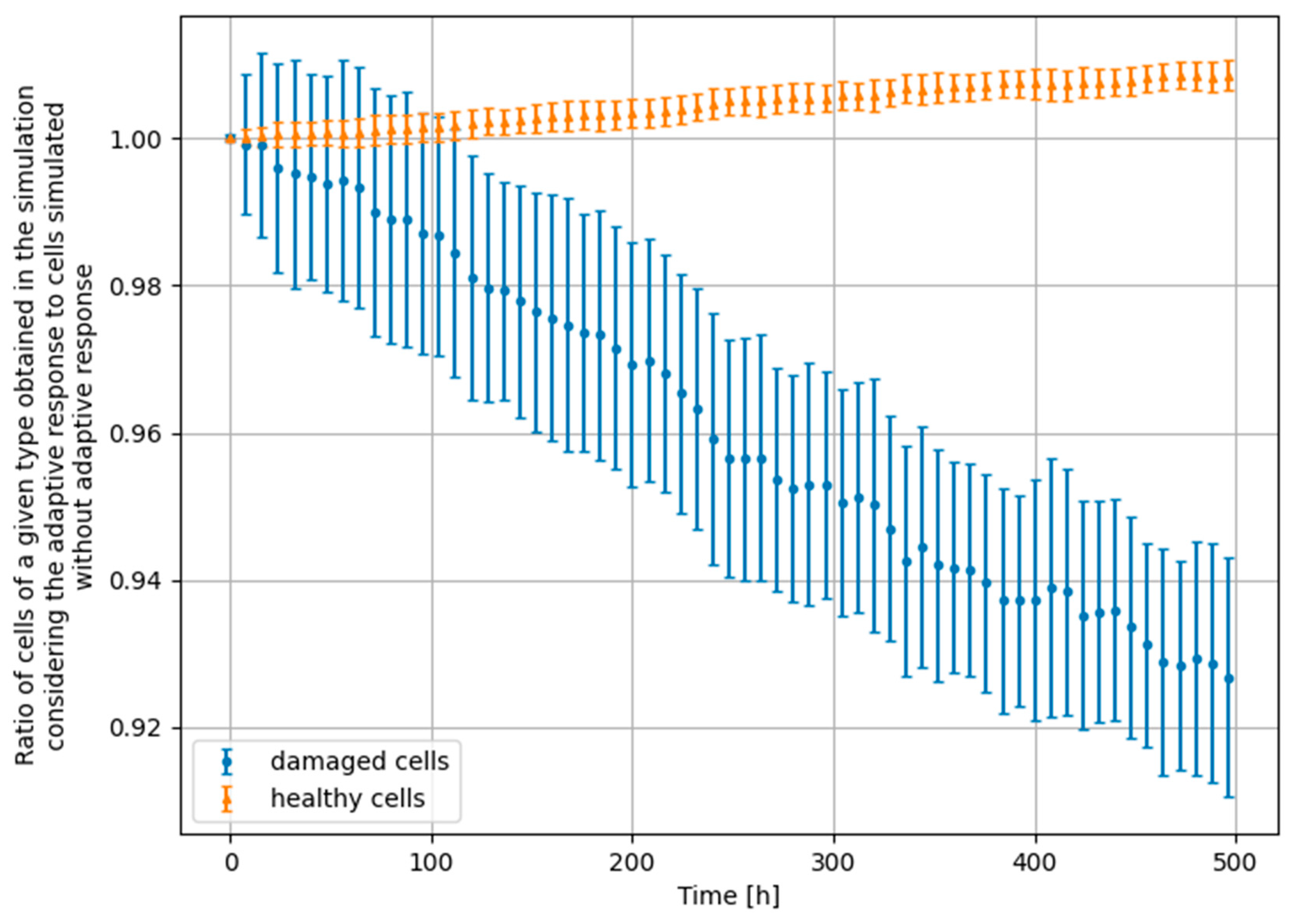
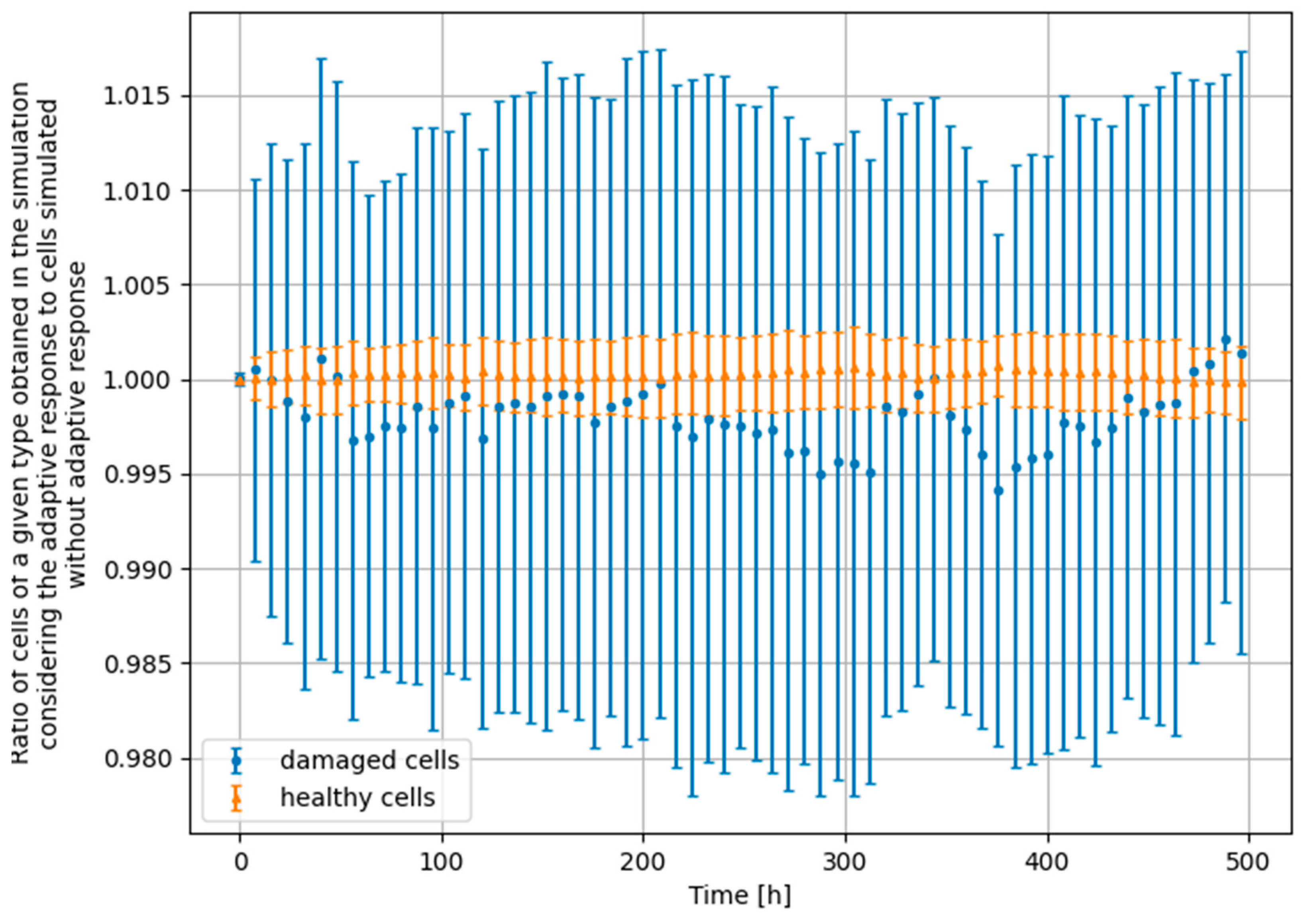
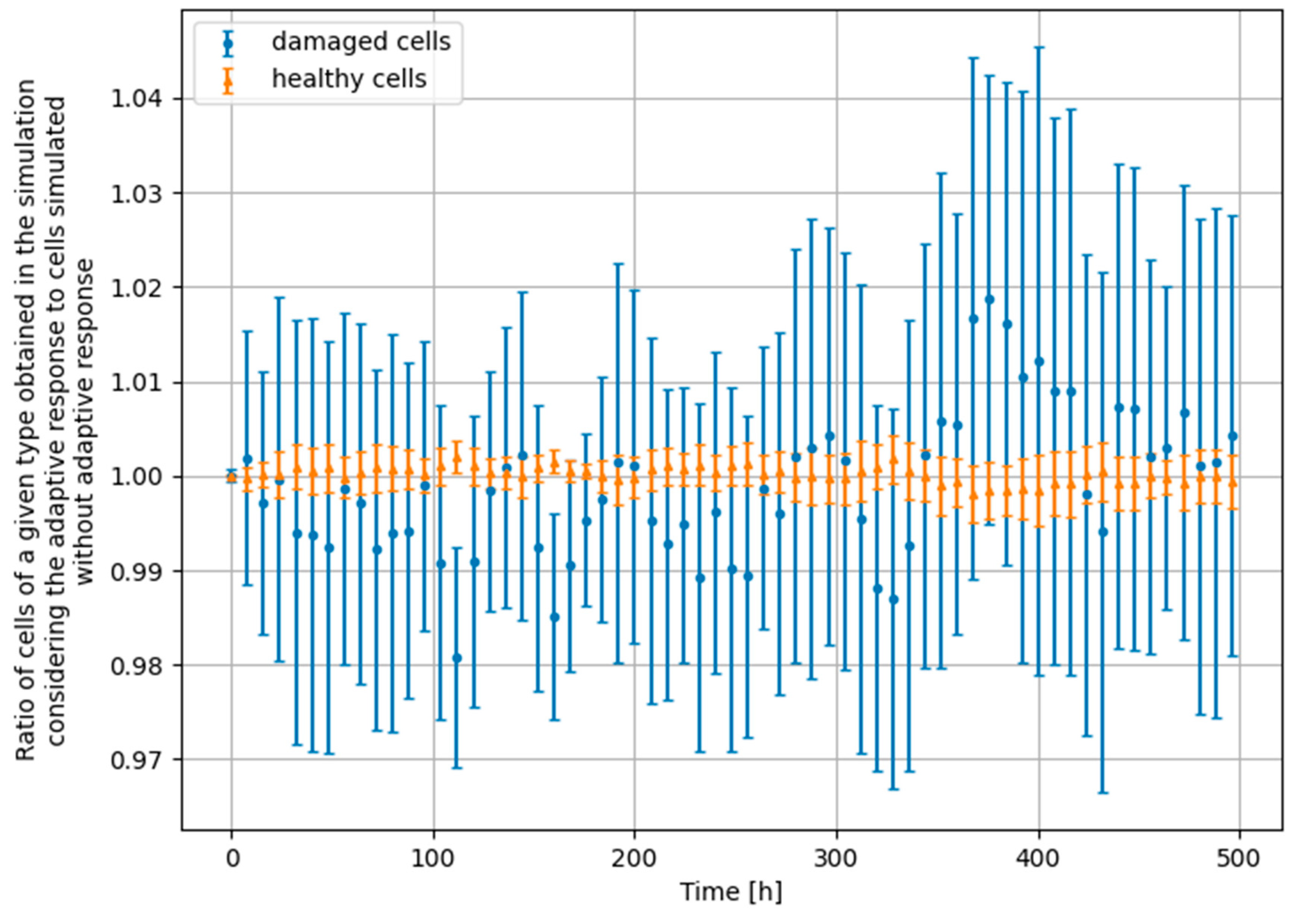
- Scenario no. 1: D1 = 25 mGy, Δt = 24 h, D2 = 1500 mGy
- Scenario no. 2: D1 = 25 mGy, Δt = 24 h, D2 = 4000 mGy
- Scenario no. 3: D1 = 25 mGy, Δt = 100 h, D2 = 1500 mGy
- Scenario no. 4: D1 = 100 mGy, Δt = 24 h, D2 = 1500 mGy
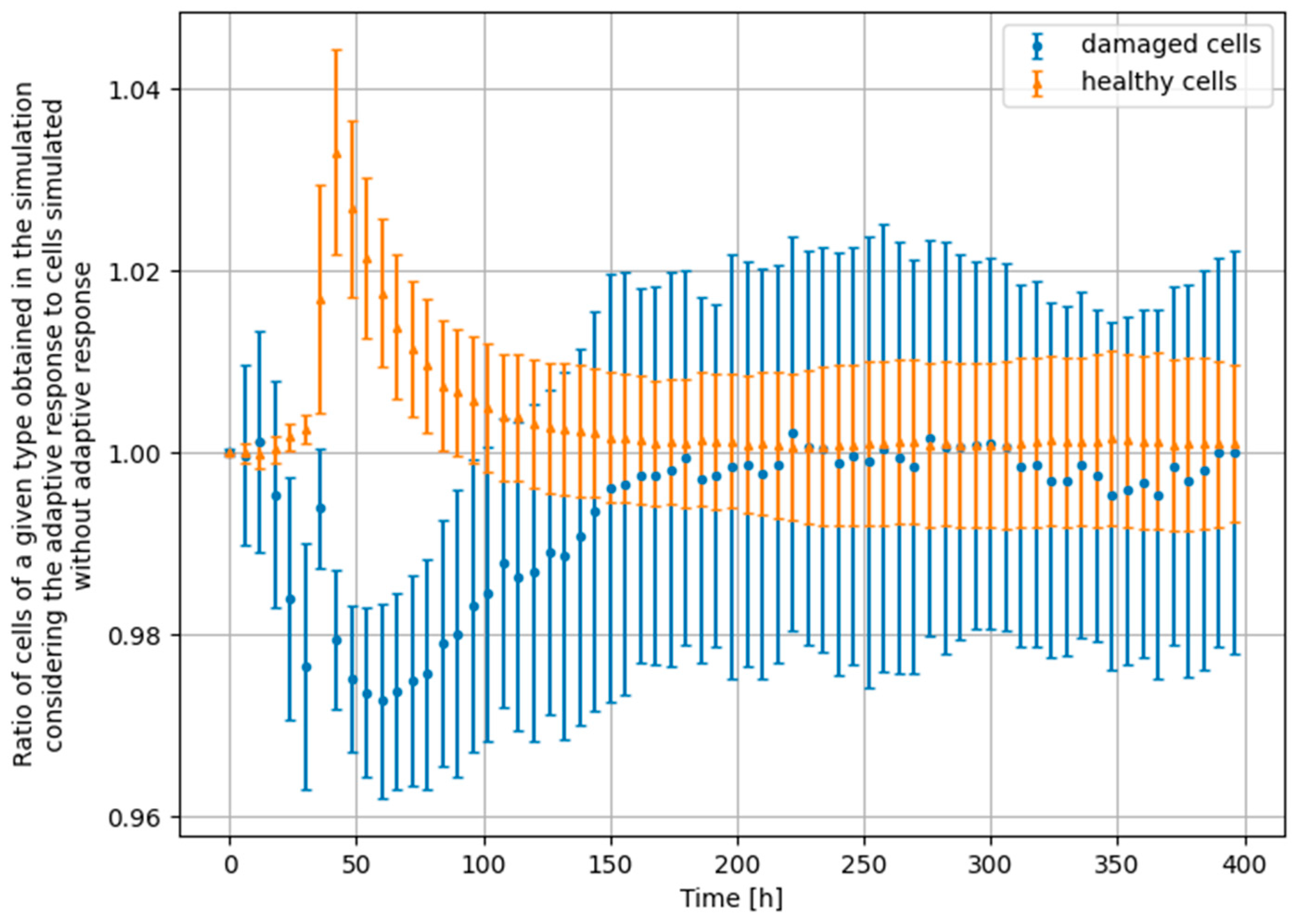
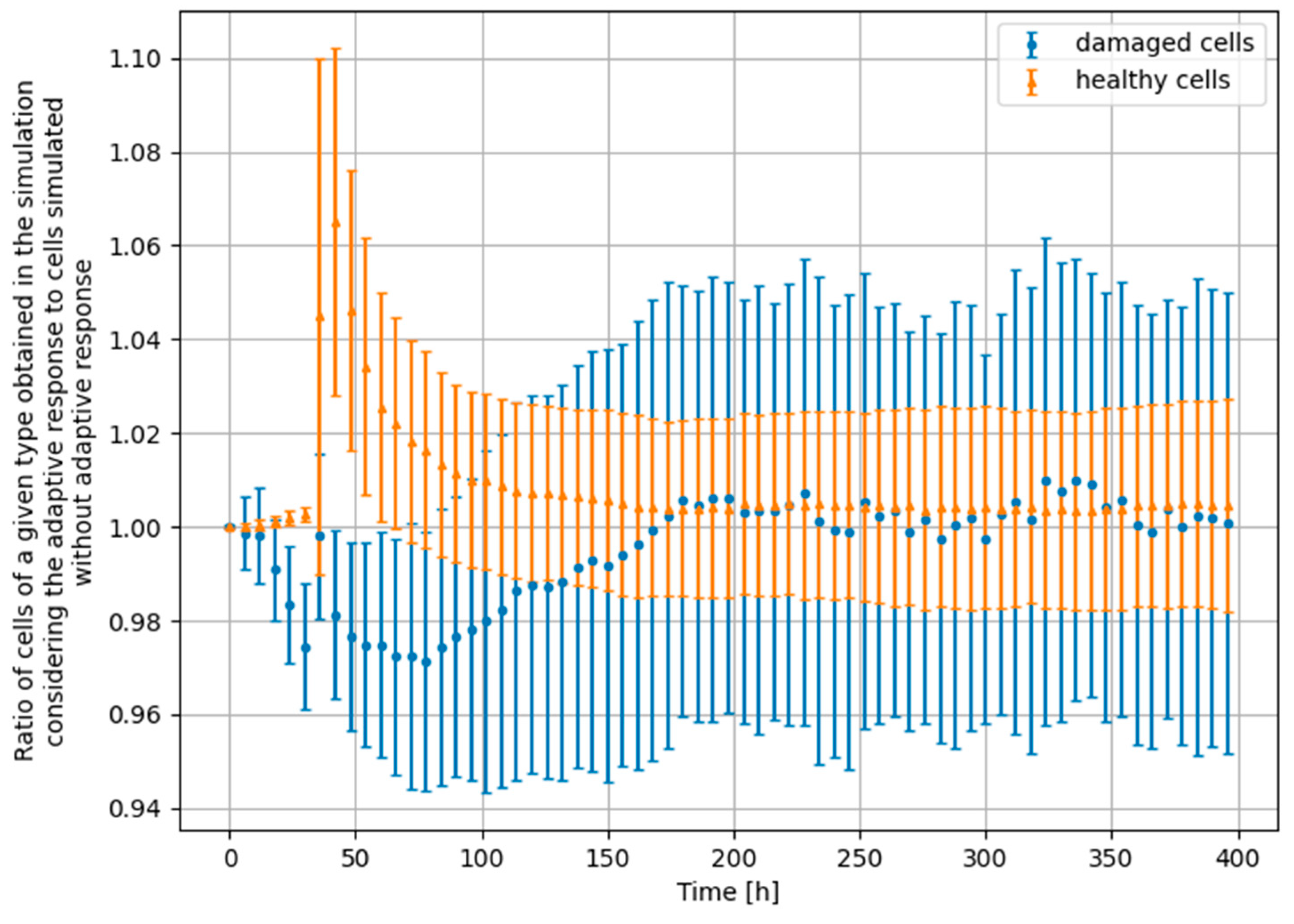
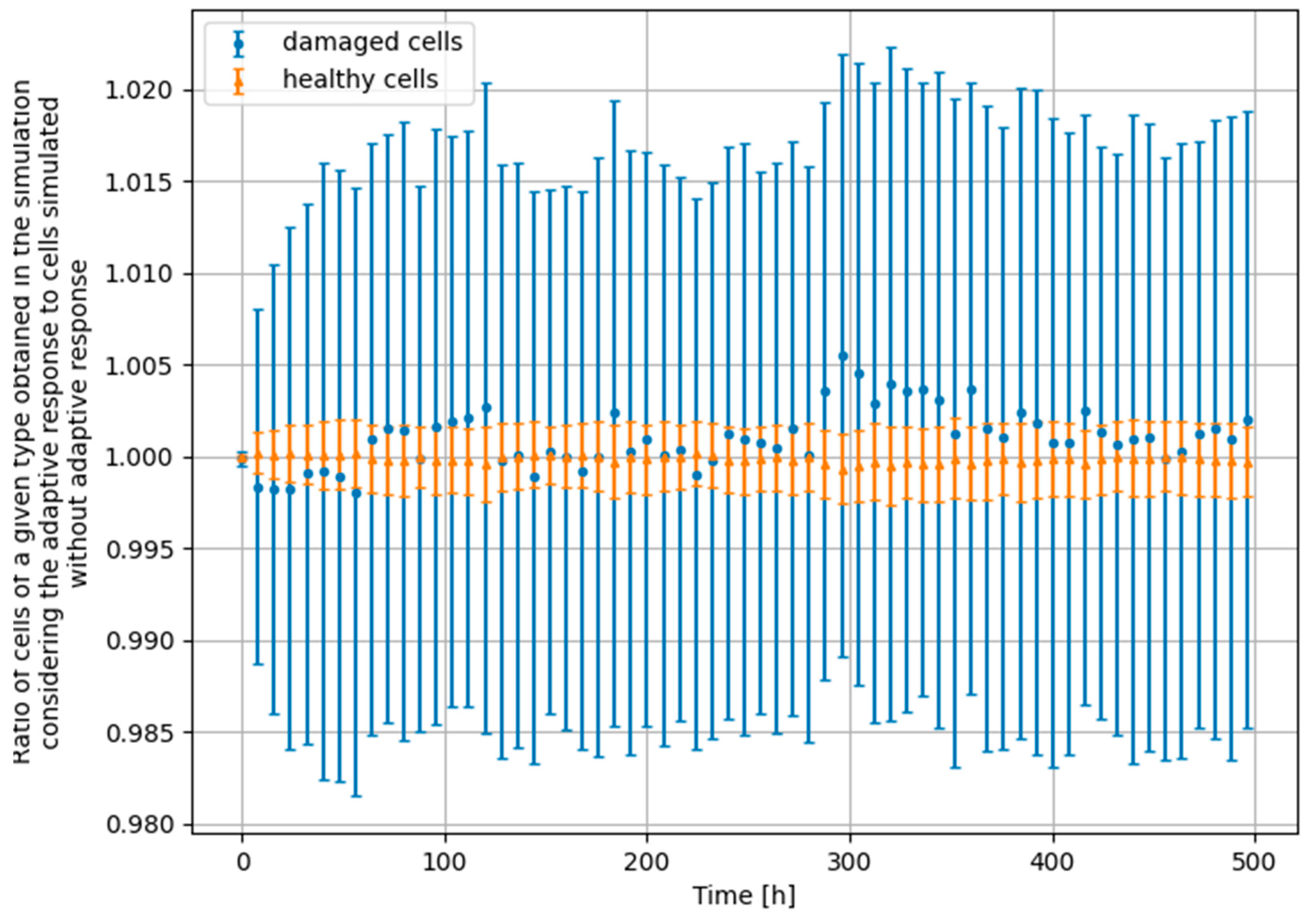
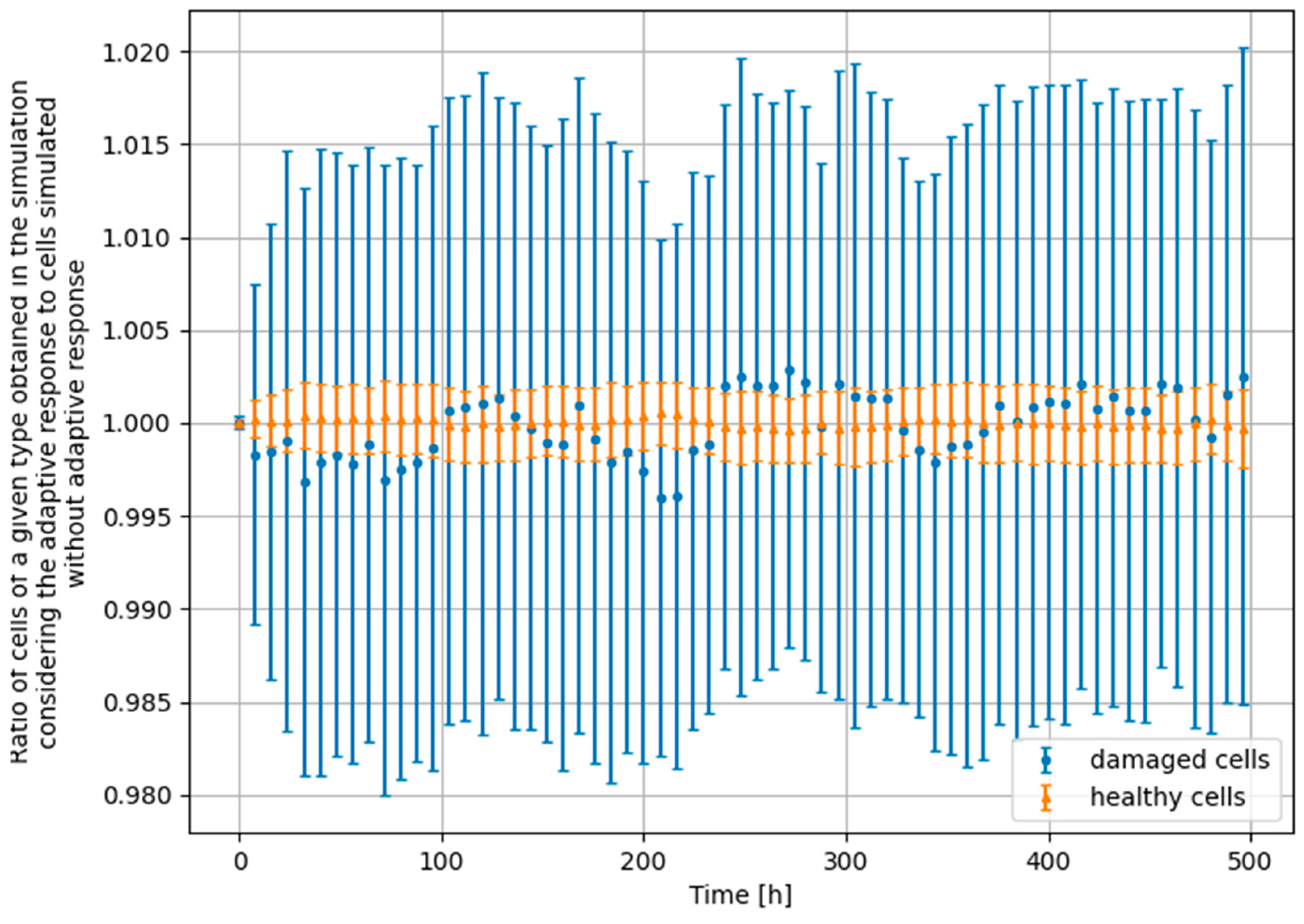
3.4. Monte Carlo Approach—Constant Dose-Rate
4. Significance of the Radioadaptive-Related Repair
5. Discussion and Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fornalski, K.W.; Adamowski, Ł.; Bugała, E.; Jarmakiewicz, R.; Kirejczyk, M.; Kopyciński, J.; Krasowska, J.; Kukulski, P.; Piotrowski, Ł.; Ponikowska, J.; et al. Biophysical modeling of the ionizing radiation influence on cells using the stochastic (Monte Carlo) and deterministic (analytical) approaches. Dose-Response, 2022; in-press (published on-line). [Google Scholar] [CrossRef]
- UNSCEAR (United Nations Scientific Committee on the Effects of Atomic Radiation) Report. Annex B: Adaptive Responses to Radiation in Cells and Organisms. 1994. Available online: https://www.unscear.org/docs/publications/1994/UNSCEAR_1994_Report.pdf (accessed on 13 February 2023).
- Raper, J.R. Effects of total surface beta irradiation. Radiology 1947, 49, 314–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olivieri, G.; Bodycote, J.; Wolff, S. Adaptive response of human lymphocytes to low concentrations of radioactive thymidine. Science 1984, 223, 594–597. [Google Scholar] [CrossRef] [PubMed]
- Sanders, C.L. Radiation Hormesis and the Linear-No-Threshold Assumption; Springer: New York, NY, USA, 2010. [Google Scholar]
- Wolff, S. The adaptive response in radiobiology: Evolving insights and implications. Environ. Health Perspect. 1998, 106 (Suppl. 1), 277–283. [Google Scholar] [PubMed]
- Aurengo, A.; Averbeck, D.; Bonnin, A.; Le Guen, B.; Masse, R.; Monier, R.; Tubiana, M.; Valleron, A.J.; de Vathaire, F. Dose-effect relationships and estimation of the carcinogenic effects of low doses of ionizing radiation. Rep. Fr. Acad. Sci. Natl. Acad. Med. 2005. Available online: https://www.radiochemistry.org/documents/html/033005_rad.html (accessed on 13 February 2023).
- Brooks, A.L.; Glines, W.M.; Waltar, A. Proceedings of the Joint American Nuclear Society and Health Physics Society Conference: Applicability of Radiation-Response Models to Low-Dose Protection Standards 1–3 October 2018, Pasco, Washington Introduction—Years in the Making. Health Phys. 2020, 118, 229–231. [Google Scholar] [CrossRef] [PubMed]
- Tapio, S.; Jacob, V. Radioadaptive response revisited. Radiat. Environ. Biophys. 2007, 46, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Guéguen, Y.; Bontemps, A.; Ebrahimian, T.G. Adaptive responses to low doses of radiation or chemicals: Their cellular and molecular mechanisms. Cell. Mol. Life Sci. 2019, 76, 1255–1273. [Google Scholar] [CrossRef]
- Feinendegen, L.E. The role of adaptive responses following exposure to ionizing radiation. Hum. Exp. Toxicol. 1999, 18, 426–432. [Google Scholar] [CrossRef]
- Feinendegen, L.E. Low Doses of Ionizing Radiation: Relationship between Biological Benefit and Damage Induction. A Synopsis. World J. Nucl. Med. 2005, 4, 21–34. [Google Scholar]
- Feinendegen, L.E. Quantification of adaptive protection following low-dose irradiation. Health Phys. 2016, 110, 276–280. [Google Scholar] [CrossRef]
- Smirnova, O.A.; Yonezawa, M. Radioprotection effect of low level preirradiation on mammals: Modeling and experimental investigations. Health Phys. 2003, 85, 150–158. [Google Scholar] [CrossRef] [PubMed]
- Smirnova, O.A.; Yonezawa, M. Radioresistance in mammals induced by low-level chronic irradiation: Modeling and experimental investigations. Health Phys. 2004, 87, 366–374. [Google Scholar] [CrossRef]
- Wodarz, D.; Sorace, R.; Komarova, N.L. Dynamics of cellular responses to radiation. PLoS Comput. Biol. 2014, 10, e1003513. [Google Scholar] [CrossRef] [PubMed]
- Bodgi, L.; Foray, N. The nucleo-shuttling of the ATM protein as a basis for a novel theory of radiation response: Resolution of the linear-quadratic model. Int. J. Radiat. Biol. 2016, 92, 117–131. [Google Scholar] [CrossRef] [PubMed]
- Devic, C.; Ferlazzo, M.L.; Foray, N. Influence of individual radiosensitivity on the adaptive response phenomenon: Toward a mechanistic explanation based on the nucleo-shuttling of ATM protein. Dose-Response 2018, 16, 1559325818789836. [Google Scholar] [CrossRef] [PubMed]
- Esposito, G.; Campa, A.; Pinto, M.; Simone, G.; Tabocchini, M.A.; Belli, M. Adaptive response: Modelling and experimental studies. Radiat. Prot. Dosim. 2011, 143, 320–324. [Google Scholar] [CrossRef]
- Socol, Y.; Shaki, Y.; Dobrzyński, L. Damped-oscillator model of adaptive response and its consequences. Int. J. Low Radiat. 2020, 11, 186–206. [Google Scholar] [CrossRef]
- Fornalski, K.W. Radioadaptation and radioresistance during deep space travels. J. Space Saf. Eng. 2022, 9, 385–389. [Google Scholar] [CrossRef]
- Fornalski, K.W.; Adamowski, Ł.; Dobrzyński, L.; Jarmakiewicz, R.; Powojska, A.; Reszczyńska, J. The radiation adaptive response and priming dose influence: The quantification of the Raper-Yonezawa effect and its three-parameter model for postradiation DNA lesions and mutations. Radiat. Environ. Biophys. 2022, 61, 221–239. [Google Scholar] [CrossRef]
- Dobrzyński, L.; Fornalski, K.W.; Reszczyńska, J.; Janiak, M.K. Modeling cell reactions to ionizing radiation: From a lesion to a cancer. Dose-Response 2019, 17, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Piotrowski, Ł. Modelowanie Odpowiedzi Grupy Komórek na Promieniowanie Jonizujące Metodą Monte Carlo. Bachelor’s Thesis, Faculty of Physics, Warsaw University of Technology, Warsaw, Poland, (supervisor: K.W. Fornalski). 2022. Available online: https://www.researchgate.net/publication/368275322_Modelowanie_odpowiedzi_grupy_komorek_na_promieniowanie_jonizujace_metoda_Monte_Carlo (accessed on 13 February 2023).
- Bugała, E. Modelowanie Radiacyjnej Odpowiedzi Adaptacyjnej dla Obszarów o Podwyższonym tle Promieniowania Jonizującego (in Polish). Master Thesis, Faculty of Physics, Warsaw University of Technology, Warsaw, Poland, (supervisor: K.W. Fornalski). 2022. Available online: https://www.researchgate.net/publication/368139245_Modelowanie_radiacyjnej_odpowiedzi_adaptacyjnej_dla_obszarow_o_podwyzszonym_tle_promieniowania_jonizujacego (accessed on 13 February 2023).
- Fornalski, K.W. Radiation adaptive response and cancer: From the statistical physics point of view. Phys. Rev. E 2019, 99, 22139. [Google Scholar] [CrossRef] [PubMed]












Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Piotrowski, Ł.; Krasowska, J.; Fornalski, K.W. Mechanistic Modelling of DNA Damage Repair by the Radiation Adaptive Response Mechanism and Its Significance. BioMedInformatics 2023, 3, 150-163. https://doi.org/10.3390/biomedinformatics3010011
Piotrowski Ł, Krasowska J, Fornalski KW. Mechanistic Modelling of DNA Damage Repair by the Radiation Adaptive Response Mechanism and Its Significance. BioMedInformatics. 2023; 3(1):150-163. https://doi.org/10.3390/biomedinformatics3010011
Chicago/Turabian StylePiotrowski, Łukasz, Julianna Krasowska, and Krzysztof W. Fornalski. 2023. "Mechanistic Modelling of DNA Damage Repair by the Radiation Adaptive Response Mechanism and Its Significance" BioMedInformatics 3, no. 1: 150-163. https://doi.org/10.3390/biomedinformatics3010011