
Effect of Pre-Germinative Treatments on Eight Priority Native Species for Reforestation in the Tropical Deciduous Forest
 , and
, and
Abstract
:1. Introduction
Selected Species
2. Materials and Methods
2.1. Pre-Germinative Treatments
- (1)
- For thermal scarification (TS), the seeds were immersed in water at a temperature between 70 °C and 94 °C.
- (2)
- The mechanical scarification (MS) consisted of the removal of a portion of 1 mm (or 0.5 mm in the case of D. viscosa) of the seed using diagonal cut pliers (Truper, model PM-CD4, Mexico City, Mexico). The size of the removed portion was dependent upon the total size of the seed and thickness of the seed coat.
- (3)
- For chemical scarification (CS), the seeds were immersed in 98% sulfuric acid for 20 to 45 min with constant manual agitation.
2.2. Statistical Analysis
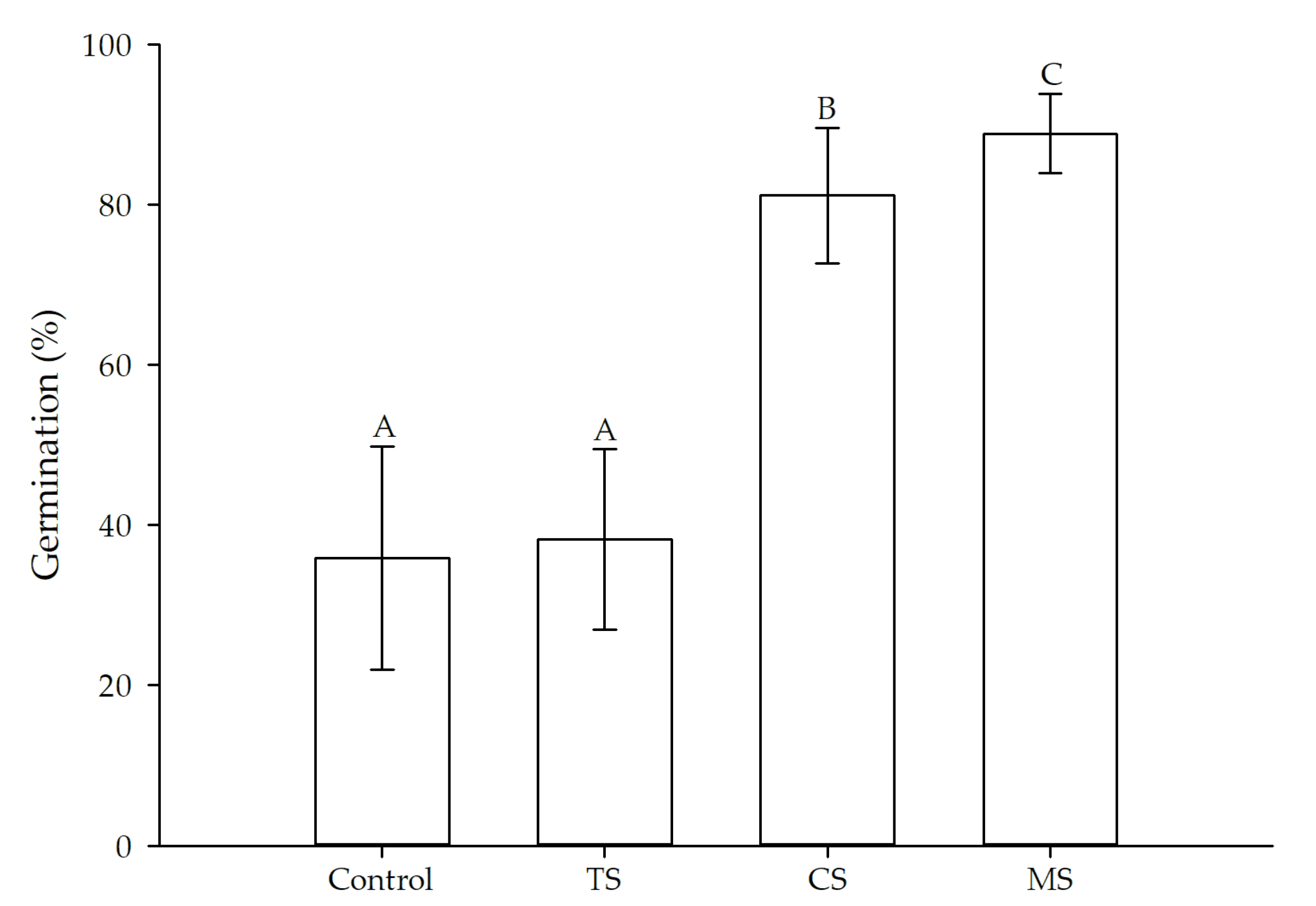
3. Results
3.1. Treatments
3.2. Species
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Schröder, J.M.; Rodríguez, L.P.; Günter, S. Research trends: Tropical dry forests: The neglected research agenda? For. Policy Econ. 2020, 122, 102333. [Google Scholar] [CrossRef]
- FAO. The State of the World’s Forests 2018-Forest Pathways to Sustainable Development; FAO: Rome, Italy, 2018; pp. i–xi. Available online: https://www.fao.org/publications/card/en/c/I9535EN/ (accessed on 30 March 2023).
- Portillo-Quintero, C.; Sanchez-Azofeifa, A.; Calvo-Alvarado, J.; Quesada, M.; do Espirito Santo, M.M. The role of tropical dry forests for biodiversity, carbon and water conservation in the neotropics: Lessons learned and opportunities for its sustainable management. Reg. Environ. Change 2015, 15, 1039–1049. [Google Scholar] [CrossRef]
- Gillespie, T.W.; Grijalva, A.; Farris, C.N. Diversity, composition, and structure of tropical dry forests in Central America. Plant Ecol. 2000, 147, 37–47. [Google Scholar] [CrossRef]
- Harker, M.L.; Hernndez-López, L.; Muñiz-Castro, M.A. Flora del bosque tropical caducifolio en una zona con suelos yesosos y calcáreos de Colima, México. Acta Bot. Mex. 2021, 128, 2448–7589. [Google Scholar] [CrossRef]
- Meave, J.; Romero-Romero, M.; Salas-Morales, S.; Pérez-García, E.; Gallardo-Cruz, J. Diversidad, amenazas y oportunidades para la conservación del bosque tropical caducifolio en el estado de Oaxaca, México. Ecosistemas 2012, 21, 85–100. Available online: https://www.revistaecosistemas.net/index.php/ecosistemas/article/view/29 (accessed on 2 February 2023).
- INEGI. Cobertura de Uso Del Suelo y Vegetación Serie VI; Mexico. 2017. Available online: https://www.inegi.org.mx/temas/usosuelo/#descargas (accessed on 20 March 2023).
- FAO. Resilient Livelihoods—Disaster Risk Reduction for Food and Nutrition Security; FAO: Rome, Italy, 2013; pp. 2–4. Available online: https://www.fao.org/fsnforum/resources/reports-and-briefs/resilient-livelihoods-disaster-risk-reduction-food-and-nutrition (accessed on 19 January 2023).
- Vieira, D.L.M.; Scariot, A. Principles of natural regeneration of tropical dry forests for restoration. Restor. Ecol. 2006, 14, 11–20. [Google Scholar] [CrossRef]
- FAO. Trees, Forests and Land Use in Drylands: The First Global Assessment—Full Report; FAO: Rome, Italy, 2019; pp. 49–51. Available online: https://www.fao.org/documents/card/en/c/ca7148en/ (accessed on 19 January 2023).
- Stan, K.; Sanchez-Azofeifa, A. Tropical Dry Forest Diversity, Climatic Response, and Resilience in a Changing Climate. Forests 2019, 10, 443. [Google Scholar] [CrossRef]
- Méndez-Toribio, M.; Martínez-Garza, C.; Ceccon, E.; Guariguata, M.R. La Restauración de Ecosistemas Terrestres en México. Estado actual, Necesidades y Oportunidades; Documentos Ocasionales; Centro para la Investigación Forestal Internacional: Bogor, Indonesia, 2018; pp. 1–7. Available online: https://www.cifor.org/knowledge/publication/6853/ (accessed on 12 December 2022).
- Álvarez-Aquino, C.; Williams-Linera, G. Seedling survival and growth of tree species: Site condition and seasonality in tropical dry forest restoration. Bot. Sci. 2012, 90, 341–351. [Google Scholar] [CrossRef]
- Pezzini, F.F.; Ranieri, B.D.; Brandão, D.O.; Fernandes, G.W.; Quesada, M.; Espírito-Santo, M.M.; Jacobi, C.M. Changes in tree phenology along natural regeneration in a seasonally dry tropical forest. Plant Biosyst. 2014, 148, 965–974. [Google Scholar] [CrossRef]
- Alberto, F.J.; Aitken, S.N.; Alía, R.; González-Martínez, S.C.; Hänninen, H.; Kremer, A.; Lefèvre, F.; Lenormand, T.; Yeaman, S.; Whetten, R.; et al. Potential for evolutionary responses to climate change—Evidence from tree populations. Glob. Change Biol. 2013, 19, 1645–1661. [Google Scholar] [CrossRef]
- Luna-Nieves, A.L.; Meave, J.A.; Morellato, L.P.C.; Ibarra-Manríquez, G. Reproductive phenology of useful Seasonally Dry Tropical Forest trees: Guiding patterns for seed collection and plant propagation in nurseries. For. Ecol. Manag. 2017, 393, 52–62. [Google Scholar] [CrossRef]
- Quesada, M. Succession and management of tropical dry forests in the Americas: Review and new perspectives. For. Ecol. Manag. 2009, 258, 1014–1024. [Google Scholar] [CrossRef]
- Derroire, G.; Tigabu, M.; Odén, P.C.; Healey, J.R. The Effects of Established Trees on Woody Regeneration during Secondary Succession in Tropical Dry Forests. Biotropica 2016, 48, 290–300. [Google Scholar] [CrossRef]
- Gordillo-Ruiz, M.C.; Pérez-Farrera, M.A.; Castillo-Santiago, M. Estructura y composición arbórea del bosque tropical caducifolio secundario en la Depresión Central, Chiapas, México. Madera Bosques 2020, 26, e2632055. [Google Scholar] [CrossRef]
- Understanding Poverty: Urban Development. Available online: https://www.worldbank.org/en/topic/urbandevelopment (accessed on 31 March 2023).
- Park, B.J.; Furuya, K.; Kasetani, T.; Takayama, N.; Kagawa, T.; Miyazaki, Y. Relationship between psychological responses and physical environments in forest settings. Landsc. Urban Plan. 2011, 102, 24–32. [Google Scholar] [CrossRef]
- Tyrväinen, L.; Ojala, A.; Korpela, K.; Lanki, T.; Tsunetsugu, Y.; Kagawa, T. The influence of urban green environments on stress relief measures: A field experiment. J. Environ. Psychol. 2014, 38, 1–9. [Google Scholar] [CrossRef]
- Castell, C. Naturaleza y salud: Una Alianza Necesaria. Gac. Sanit. 2020, 34, 194–196. [Google Scholar] [CrossRef]
- Coppel, G.; Wüstemann, H. The impact of urban green space on health in Berlin, Germany: Empirical findings and implications for urban planning. Landsc. Urban Plan. 2017, 167, 410–418. [Google Scholar] [CrossRef]
- Martínez-Soto, J.; Montero-López, M.; Córdova-Vázquez, A. Restauración Psicológica y Naturaleza Urbana: Algunas Implicaciones Para La Salud Mental. Salud Ment. 2014, 37, 217–224. Available online: https://www.scielo.org.mx/scielo.php?script=sci_arttext&pid=S0185-33252014000300005 (accessed on 1 April 2023). [CrossRef]
- Kowarik, I. Novel Urban Ecosystems, Biodiversity, and Conservation. Environ. Pollut. 2011, 159, 1974–1983. [Google Scholar] [CrossRef]
- Kleyer, M.; Minden, V. Why functional ecology should consider all plant organs: An allocation-based perspective. Basic Appl. Ecol. 2015, 16, 1–9. [Google Scholar] [CrossRef]
- Grubb, P.J. The maintenance of species-Richness in plant communities: The importance of the regeneration niche. Biol. Rev. 1977, 52, 107–145. [Google Scholar] [CrossRef]
- Jiménez-Alfaro, B.; Silveira, F.A.O.; Fidelis, A.; Poschold, P. Seed germination traints can contribute better to plant community ecology. J. Veg. Sci. 2016, 27, 637–645. [Google Scholar] [CrossRef]
- Schleicher, A.; Biedermann, R.; Kleyer, M. Dispersal traits determine plant response to habitat connectivity in an urban landscape. Landsc. Ecol. 2011, 26, 529–540. [Google Scholar] [CrossRef]
- Sánchez-Gómez, A.; Rosendo-Ponce, A.; Vargas-Romero, J.M.; Rosales-Martínez, F.; Platas-Rosado, D.E.; Becerril-Pérez, D.M. Energía germinativa en guaje (Leucaena leucocephala cv. cunningham) con diferentes métodos de escarificación de la semilla. Agrociencia 2018, 52, 863–874. Available online: https://www.scielo.org.mx/scielo.php?script=sci_arttext&pid=S1405-31952018000600863 (accessed on 12 December 2022).
- Tadros, M.J.; Samarah, N.H.; Alqudah, A.M. Effect of different pre-sowing seed treatments on the germination of Leucaena leucocephala (Lam.) and Acacia farnesiana (L.). New For. 2011, 42, 397–407. [Google Scholar] [CrossRef]
- Vargas, G.H.; Velásquez, L.R.S.; Aragón, F. Tratamientos pregerminativos en cuatro especies arbóreas de uso forrajero de la selva baja caducifolia de la Sierra de Manantlán. For. Ver. 2001, 3, 9–15. Available online: https://www.redalyc.org/articulo.oa?id=49730102 (accessed on 8 March 2023).
- Encino-Ruiz, L.; Linding-Cisneros, R.; Gomez-Romero, M.; Blanco-García, A. Desempeño de tres especies arbóreas del bosque tropical caducifolio en un ensayo de restauración ecológica. Bot. Sci. 2013, 91, 107–114. Available online: https://www.scielo.org.mx/scielo.php?script=sci_arttext&pid=S2007-42982013000100009 (accessed on 12 December 2022). [CrossRef]
- Benítez-Rodríguez, L.; Gamboa-de Buen, A.; Sánchez-Coronado, M.E.; Alvarado-López, S.; Soriano, D.; Méndez, I.; Vázquez-Santana, S.; Carabias-Lillo, J.; Mendoza, A.; Orozco-Segovia, A. Effects of seed burial on germination, protein mobilisation and seedling survival in Dodonaea viscosa. Plant. Biol. 2014, 16, 732–739. [Google Scholar] [CrossRef]
- Cervantes-Gutierrez, M.G. La reforestación en la Montaña de Guerrero: Una Estrategia Alternativa con Leguminosas Nativas. Master’s Thesis, UNAM, Mexico City, Mexico, 1996. Available online: https://repositorio.unam.mx/contenidos/la-reforestacion-en-la-montana-de-guerrero-una-estrategia-alternativa-con-leguminosas-353687?c=a88jq3&d=false&q=*:*&i=1&v=1&t=search_0&as=0 (accessed on 9 December 2022).
- Rivas-Medina, G.; Cervantes, G.G.; Castro, C.M.V.; Cohen, I.S.; Díaz, J.V. Morfología y escarificación de la semilla de mezquite, huizache y ahuehuete. Téc. Pecu. Méx. 2005, 43, 441–448. Available online: https://www.redalyc.org/articulo.oa?id=61343314 (accessed on 9 December 2022).
- Ffolliott, P.F.; Thames, J.L. Collection, Handling, Storage and Pre-Treatment of Prosopis Seeds in Latin America, 1st ed.; Food & Agriculture Organization of the United Nations: Tucson, AZ, USA, 1983; pp. 2–33. Available online: https://www.fao.org/3/Q2180E/Q2180E00.htm#TOC (accessed on 1 February 2023).
- Coronado-Perez, B. Efecto de dos Tratamientos Pregerminativos en el Crecimiento de Senna septemtrionalis Bajo Diferentes Condiciones de Estrés Hídrico. Bachelor’s Thesis, UNAM, Tlalnepantla de Baz, Mexico, 2018. [Google Scholar]
- De Melo, P.A.F.R.; Silva, K.B.; Alves, E.U.; de Medeiros, R.L.S.; Neto, A.P.A.; Pinto, K.M.S.; Leite, W.S.; Matos, V.P. Morphological analysis of fruits, seeds, and seedling germination Acacia farnesiana (L.) Willd. Afr. J. Agric. 2016, 11, 2912–2919. [Google Scholar] [CrossRef]
- Jayasuriya, K.G.; Wijetunga, A.S.; Baskin, J.M.; Baskin, C.C. Seed dormancy and storage behaviour in tropical Fabaceae: A study of 100 species from Sri Lanka. Seed Sci. Res. 2013, 23, 257–269. [Google Scholar] [CrossRef]
- Sobrevilla-Solís, J.A.; López-Herrera, M.; López-Escamilla, A.L.; Romero-Bautista, L. Evaluación de diferentes tratamientos pregerminativos y osmóticos en la germinación de semillas Prosopis laevigata (Humb. y Bonpl. Ex Willd) MC Johnston. In Estudios Científicos en el Estado de Hidalgo y Zonas Aledañas; Pulido-Flores, G., Monks, S., Eds.; University of Nebraska–Lincoln Libraries: Lincoln, NE, USA, 2013; Volume 2, pp. 83–95. Available online: https://digitalcommons.unl.edu/zeabook/16 (accessed on 11 December 2022).
- Cervantes, M.; Ceccon, E.; Bonfil, C. Germination of stored seeds of four tree species from the tropical dry forest of Morelos, Mexico. Bot. Sci. 2014, 92, 281–287. [Google Scholar] [CrossRef]
- Al-Namazi, A.A.; Al-Ammari, B.S.; Davy, A.J.; Al-Turki, T.A. Seed dormancy and germination in Dodonaea viscosa (Sapindaceae) from south-western Saudi Arabia. Saudi J. Biol. Sci. 2020, 27, 2420–2424. [Google Scholar] [CrossRef]
- Silva-Farías, M.A. Ensayos de Procedencias y Características de las Semillas y la Madera de Albizia plurijuga (Standl.) Britt. et Rose. Master’s Thesis, Universidad Michoacana de San Nicolás Hidalgo, Michoacan, Mexico, 2012. [Google Scholar]
- Sepúlveda-Ríos, O. Técnicas de pre germinación en leucaena (Leucaena leucocephala w.) y botón de oro (Tithonia diversifolia g.) como forrajeras con potencial para alimentación animal. Bachelor’s Thesis, Universidad Francisco de Paula Santander, Norte de Santander, Colombia, 2020. Available online: https://repositorio.ufps.edu.co/handle/ufps/4370 (accessed on 11 December 2022).
- Mendonça, A.J.T.; Silva, M.C.C.; Berto, F.H.R.; Gondim, A.R.D.O.; Medeiros, M.N.V.; Lins, W.L. Superação de dormência em sementes de Leucaena leucocephala (Lam.) de Wit. com métodos físicos e químicos. Rev. Verde Agro. Des. Sust 2020, 15, 325–329. [Google Scholar] [CrossRef]
- Rueda-Sánchez, A.; Benavides-Solorio, J.D.; Saenz-Reyez, J.T.; Muñoz Flores, H.J.; Prieto-Ruiz, J.Á.; Orozco-Gutiérrez, G. Calidad de planta producida en los viveros forestales de Nayarit. Rev. Mex. Cienc. For. 2014, 5, 58–73. Available online: http://www.scielo.org.mx/scielo.php?script=sci_arttext&pid=S2007-11322014000200005&lng=es&tlng=es (accessed on 10 December 2022). [CrossRef]
- Baskin, C.C.; Baskin, J.M. Seeds: Ecology, Biogeography, and, Evolution of Dormancy and Germination, 2nd ed.; Academic Press: San Diego, CA, USA, 1998; pp. 27–42. [Google Scholar]
- Baskin, J.M.; Baskin, C.C.; Li, X. Taxonomy, anatomy and evolution of physical dormancy in seeds. Plant Species Biol. 2000, 15, 139–152. [Google Scholar] [CrossRef]
- Baskin, J.M.; Davis, B.H.; Baskin, C.C.; Gleason, S.M.; Cordell, S. Physical dormancy in seeds of Dodonaea viscosa (Sapindales, Sapindaceae) from Hawaii. Seed Sci. Res. 2004, 14, 81–90. [Google Scholar] [CrossRef]
- Teketay, D. The effect of different pre-sowing seed treatments, temperature and light on the germination of five Senna species from Ethiopia. New For. 1996, 11, 155–171. Available online: https://link.springer.com/article/10.1007/BF00033411 (accessed on 4 December 2022). [CrossRef]
- Kimura, E.; Islam, M.A. Seed Scarification Methods and their Use in Forage Legumes. Res. J. Seed Sci. 2012, 5, 38–50. Available online: https://scialert.net/abstract/?doi=rjss.2012.38.50 (accessed on 4 December 2022).
- Vari, A.K.; Jethani, I.; Sharma, S.P.; Khanna, M.; Barnwal, S. Seed coat-imposed dormancy in Sesbania spp. and treatments to improve germination. Seed. Sci. Technol. 2007, 35, 318–325. [Google Scholar] [CrossRef]
- Benayas, J.M.R.; Newton, A.C.; Diaz, A.; Bullock, J.M. Enhancement of Biodiversity and Ecosystem Services by Ecological Restoration: A Meta-Analysis. Science 1979, 325, 1121–1124. [Google Scholar] [CrossRef]
- Benítez, G.; Equihua, M. Diagnóstico de la situación de los viveros oficiales de Veracruz y su papel para apoyar programas de reforestación y restauración. Rev. Chapingo 2002, 8, 5–12. Available online: https://www.redalyc.org/articulo.oa?id=62980101 (accessed on 7 February 2023).
- Rodríguez-Larramendi, L.A.; Sánchez-Cortés, M.S.; Gordillo-Ruiz, M.C. Árboles útiles del bosque tropical caducifolio secundario en la Reserva Forestal Villa Allende, Chiapas, México. Acta Bot. Mex. 2018, 125, 189–214. [Google Scholar] [CrossRef]
- Rico-Arce, M.L.; Gale, S.L.; Maxted, N. A taxonomic study of Albizia (Leguminosae: Mimosoideae: Ingeae) in Mexico and Central America. An. Jardín Bot. Madr. 2008, 65, 255–305. Available online: https://dialnet.unirioja.es/servlet/articulo?codigo=2786975 (accessed on 29 January 2023). [CrossRef]
- Allen, O.; Allen, E. The Leguminosae—A Source Book of Characteristics, Uses, and Nodulation; The University of Wisconsin Press: Madison, WI, USA, 1981; pp. 30–32. [Google Scholar]
- Pérez, J.I.J. Funciones E Importancia Económica Del Recurso Chapulixtle (Dodonaea viscosa) En Un Ejido Del Subtrópico Mexicano. Obs. Iberoam. Desarro. Local 2013, 7. Available online: https://ideas.repec.org/a/erv/oidles/y2013i141.html (accessed on 3 February 2023).
- FAO. Trees and Shrubs of the Maldives, 1st ed.; FAO: Bangkok, Thailand, 2007; pp. 83–84. [Google Scholar]
- Cansino, J.V. Botánica Económica de Cuatro Especies de San Juan Ixcaquxstla, Mixteca Poblana. Bachelor’s Thesis, Universidad Autónoma de Chapingo, Texcoco, Mexico, 2001. Available online: https://es.slideshare.net/CharlieSC4/botnica-econmica-de-cuatro-especies-de-san-juan-ixcaquixtla (accessed on 4 December 2022).
- Peralta-Juárez, I.; Gómez-Campos, A.; Romero-Castillo, P.A.; Reyes-Dorantes, A. Uso antropocéntrico Del Guaje Leucaena esculenta (Moc. y Sessé Ex. DC.) Benth, en dos comunidades de la mixteca baja oaxaqueña. Polibotánica 2017, 14, 349–364. [Google Scholar] [CrossRef]
- Andrade, M.; Greter, R.; Hernandez, H.; Medina-Lemos, R.; Rico, L.; Sousa, M. Flora del Valle de Tehuacán-Cuicatlan, 1st ed.; UNAM: Mexico City, Mexico, 2012; pp. 38–43. [Google Scholar]
- De Stefano, R.D.; Tun, C.T.; Contreras, J.E.L.; Fernández-Concha, G.C.; Leopardi-Verde, C.L.; Ramírez-Prado, J.H.; Can-Itza, L.L.; Cen, I.T. Filogenia de Lysiloma (Fabaceae), un género restringido a Megaméxico con especies atípicas en las Antillas y Florida. Acta Bot. Mex. 2021, 128, e1782. [Google Scholar] [CrossRef]
- Rodríguez-Sauceda, E.N. Análisis técnico del árbol de mezquite (Prosopis laevigata Humb. & Bonpl. ex Willd.) en Mexico. Ra Ximhai 2014, 10, 173–193. Available online: https://www.redalyc.org/articulo.oa?id=46131111013 (accessed on 4 December 2022).
- Palacios-Romero, A.; Jiménez-Muñoz, E.; Rodríguez-Laguna, R.; Razo-Zárate, R. Distribución potencial de Prosopis laevigata (Humb. et Bonpl. ex Willd.) M.C. Johnst. en el estado de Hidalgo, México. Rev. Mex. Cienc. For. 2020, 12, 71–87. [Google Scholar] [CrossRef]
- García-López, J.C.; Duran-Garcia, H.M.; De-Nova, J.A.; Álvarez-Fuentes, G.; Pinos-Rodriguez, J.; Lee-Rangel, H.A.; Lopez Aguirre, S.; Ruiz-Tavares, D.; Rendon-Huerta, J.; Vicente-Martínez, J.; et al. Producción y contenido nutrimental de vainas de tres variantes de mezquite (Prosopis laevigata) en el altiplano potosino, México. Agrociencia 2020, 53, 821–832. Available online: https://agrociencia-colpos.org/index.php/agrociencia/article/view/1846 (accessed on 3 February 2023).
- Lewis, G.P.; Irwin, H.S.; Barneby, R.C. The American Cassiinae: A Synoptical Revision of Leguminosae Tribe Cassieae Subtribe Cassiinae in the New World. Kew Bull. 1984, 39, 664–666. [Google Scholar] [CrossRef]
- Rzedowski, J.; Rzedowski, G.C. Flora del Bajío y de Regiones Adyacentes. Familia Leguminosae: Subfamilia Caesalpinioideae; Instituto de Ecología A.C.: Michoacan, Mexico, 1997; pp. 98–101. [Google Scholar]
- García-Martínez, M.A.; Rodríguez, A. Vegetation and phanerogamic flora of Piedras Bola protected area in Jalisco, Mexico. Polibotánica 2018, 48, 71–90. [Google Scholar] [CrossRef]
- Arana-Argáez, V.E.; Domínguez, F.; Moreno, D.A.; Isiordia-Espinoza, M.A.; Lara-Riegos, J.C.; Ceballos-Góngora, E.; Zapata-Morales, J.R.; Franco-de la Torre, L.; Sánchez-Enríquez, S.; Alonso-Castro, A.J. Anti-inflammatory and antinociceptive effects of an ethanol extract from Senna septemtrionalis. Inflammopharmacology 2020, 28, 541–549. [Google Scholar] [CrossRef] [PubMed]
- Mimosaceae = Leguminosae en parte Acacia farnesiana (L.) Willd. Huizache. Malezas de México. Available online: http://www.conabio.gob.mx/malezasdemexico/mimosaceae/acacia-farnesiana/fichas/ficha.htm (accessed on 3 February 2023).
- Erkovan, H.İ.; Clarke, P.J.; Whalley, R.D.B. A review on General Description of Vachellia farnesiana (L.) Wight &Arn. Atatürk Univ. J. Agric. Fac. 2016, 47, 71–76. Available online: https://dergipark.org.tr/en/pub/ataunizfd/issue/27764/293472 (accessed on 3 February 2023).
- López-Merlín, D.; Soto-Pinto, L.; Jiménez-Ferrer, G.; Hernández-Daumás, S. Relaciones alométricas para la predicción de biomasa forrajera y leña de Acacia pennatula y Guazuma ulmifolia en dos comunidades del norte de Chiapas, México. Interciencia 2003, 28, 334–339. Available online: http://ve.scielo.org/scielo.php?script=sci_arttext&pid=S0378-18442003000600005&lng=es&tlng=es (accessed on 9 March 2023).
- Niembro-Rocas, A.; Vázquez-Torres, M.; Sánchez-Sánchez, O. Árboles de Veracruz: 100 Especies para la Reforestación Estratégica, 1st ed.; Secretaría de Educación del Estado de Veracruz: Xalapa-Enríquez, Mexico, 2010; pp. 26–27. Available online: https://libros.uv.mx/index.php/UV/catalog/book/FC138 (accessed on 2 February 2023).
- Ribeiro-Oliveira, J.P.; Ranal, M.A.; Garcia de Santana, D.; Pereira, L.A. Sufficient sample size to study seed germination. Aust. J. Bot. 2016, 64, 295–301. [Google Scholar] [CrossRef]
- Ribeiro-Oliveira, J.P.; Ranal, M.A. Sample size in studies on the germination process. Botany 2016, 94, 103–115. [Google Scholar] [CrossRef]
- Bradow, J.M.; Bauer, P.J. Germination and Seedling Development. In Physiology of Cotton, 1st ed.; Stewart, J.M., Oosterhuis, D.M., Heitholt, J.J., Mauney, J.R., Eds.; Springer: Dordrecht, The Netherlands, 2010; pp. 48–56. [Google Scholar] [CrossRef]
- Moares, F.G.D.; de Matos, V.P.; Silva, M.; da Ferreira, E.G.B.S.; Santos, H.H.D.; Rodrigues, I.A.S.; Bittar, S.M.B. Tratamentos pré-germinativos em sementes de Vachellia farnesiana (L.) Wight & Arn.—Leguminosae-Mimosoidae. Sci. Plena 2013, 8, 04732. Available online: https://www.scientiaplena.org.br/sp/article/view/1430 (accessed on 9 March 2023).
- Gómez-Ruiz, P.A.; Sáenz-Romero, C.; Lindig-Cisneros, R. Early performance of two tropical dry forest species after assisted migration to pine–oak forests at different altitudes: Strategic response to climate change. J. For. Res. 2020, 31, 1215–1223. [Google Scholar] [CrossRef]
- Campbell, R.K.; Sorensen, F.C. Genetic implications of nursery practices. In Forestry Nursery Manual: Production of Bareroot Seedlings, 1st ed.; Duryea, M.L., Landis, T.D., Eds.; Forestry Sciences: Dordrecht, The Netherlands, 1984; Volume 11, pp. 183–191. [Google Scholar] [CrossRef]
- Figueroa-Torres, I. Efecto en el Crecimiento de Albizia occidentalis en Inoculación Dual con Hongos Micorrícicos y Nanoparticulas como Factor de Tolerancia al Estrés Hídrico. Master’s Thesis, Universidad Michoacana de San Nicolas de Hidalgo, Michoacan, Mexico, 2021. Available online: http://bibliotecavirtual.dgb.umich.mx:8083/xmlui/handle/DGB_UMICH/6396 (accessed on 3 December 2022).






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Chemical Scarification | Thermal Scarification | |
|---|---|---|---|
| Temperature °C | Time | ||
| Albizia occidentalis | 20 min | 80 | 4 min |
| Dodonaea viscosa | 5 min | 80 | 20 min |
| Lysiloma divaricatum | 10 min | 80 | 3 min |
| Leucaena esculenta | 3 min | 80 | 3 min |
| Prosopis laevigata | 5 min | 94 | 45 s |
| Senna septemtrionalis | 45 min | 94 | 15 s |
| Vachellia farnesiana | 20 min | 70 | 20 min |
| Vachellia pennatula | 20 min | 70 | 20 min |
| Species | Control (%) | TS (%) | CS (%) | MS (%) |
|---|---|---|---|---|
| Albizia occidentalis | 88 ± 3.2 | 0 ± 0 | 91 ± 6.8 | 98 ± 2.3 |
| Dodonaea viscosa | 7 ± 5 | 61 ± 6 | 69 ± 10.5 | 67 ± 10.5 |
| Leucaena esculenta | 34 ± 5.1 | 23 ± 15.0 | 89 ± 8.2 | 72 ± 11.7 |
| Lysiloma divaricatum | 98 ± 2.3 | 40 ± 35.6 | 94 ± 6.9 | 100 ± 0 |
| Prosopis laevigata | 50 ± 14.7 | 3 ± 2 | 96 ± 4.6 | 96 ± 5.6 |
| Senna septemtrionalis | 3 ± 3.8 | 76 ± 10.3 | 88 ± 8.6 | 78 ± 9.5 |
| Vachellia farnesiana | 5 ± 6 | 21 ± 13.2 | 26 ± 2.3 | 100 ± 0 |
| Vachellia pennatula | 2 ± 4 | 82 ± 9.5 | 96 ± 3.2 | 100 ± 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Jesús-Velázquez, J.; Cisneros-Villaseñor, A.; Tamayo-Bustamante, R.A.; Girón-Gutiérrez, D.; Luna-Soria, H.; Cambrón-Sandoval, V.H. Effect of Pre-Germinative Treatments on Eight Priority Native Species for Reforestation in the Tropical Deciduous Forest. Conservation 2023, 3, 277-290. https://doi.org/10.3390/conservation3020019
De Jesús-Velázquez J, Cisneros-Villaseñor A, Tamayo-Bustamante RA, Girón-Gutiérrez D, Luna-Soria H, Cambrón-Sandoval VH. Effect of Pre-Germinative Treatments on Eight Priority Native Species for Reforestation in the Tropical Deciduous Forest. Conservation. 2023; 3(2):277-290. https://doi.org/10.3390/conservation3020019
Chicago/Turabian StyleDe Jesús-Velázquez, Juan, Alejandro Cisneros-Villaseñor, Ricardo Armando Tamayo-Bustamante, Dioseline Girón-Gutiérrez, Hugo Luna-Soria, and Víctor Hugo Cambrón-Sandoval. 2023. "Effect of Pre-Germinative Treatments on Eight Priority Native Species for Reforestation in the Tropical Deciduous Forest" Conservation 3, no. 2: 277-290. https://doi.org/10.3390/conservation3020019