
Living in a Thermally Diverse Environment: Field Body Temperatures and Thermoregulation in Hermann’s Tortoise, Testudo hermanni, in Montenegro

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
- (1)
- How effective is T. hermanni as a thermoregulator in attaining preferred body temperatures during different weather conditions and seasons? For example, when the environment begins to cool, do they increase thermoregulatory efforts or remain thermally passive and accept lower body temperatures? This is important because of the influence of body temperature levels on physiological processes.
- (2)
- What would be the thermal consequences for tortoises to remain in shaded areas avoiding open sunlit clearings or patches? Would they be capable of achieving preferred temperatures and, if so, how frequently?
- (3)
- What would be the thermal consequences for a tortoise to forage in open sunlit areas during hot weather for long periods?
- (4)
- Are there thermal consequences due to size differences in different individuals, for example between males and larger females?
- (5)
- We assumed that a tortoise could equilibrate body temperature in dappled sunlit areas by either being half in sun and half in shade or spending 50% of its time within sun and shade. To what extent do tortoise body temperatures agree with model temperatures in this type of habitat?
2. Methods
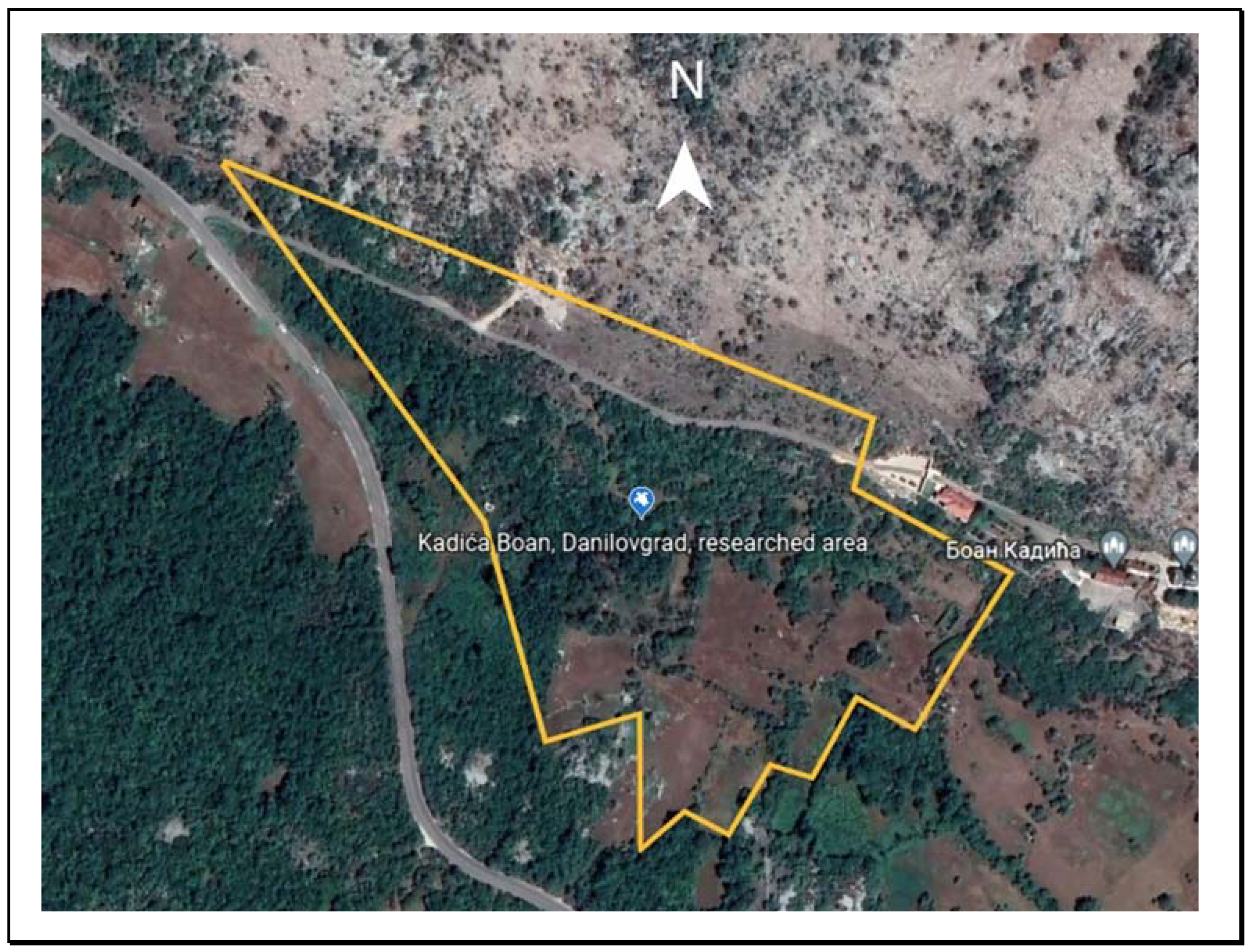
2.1. Species and Study Area
2.2. Statistic Analysis
3. Results
3.1. Tortoise Numbers, Sex Ratios and Their Sizes
3.2. Body Temperatures
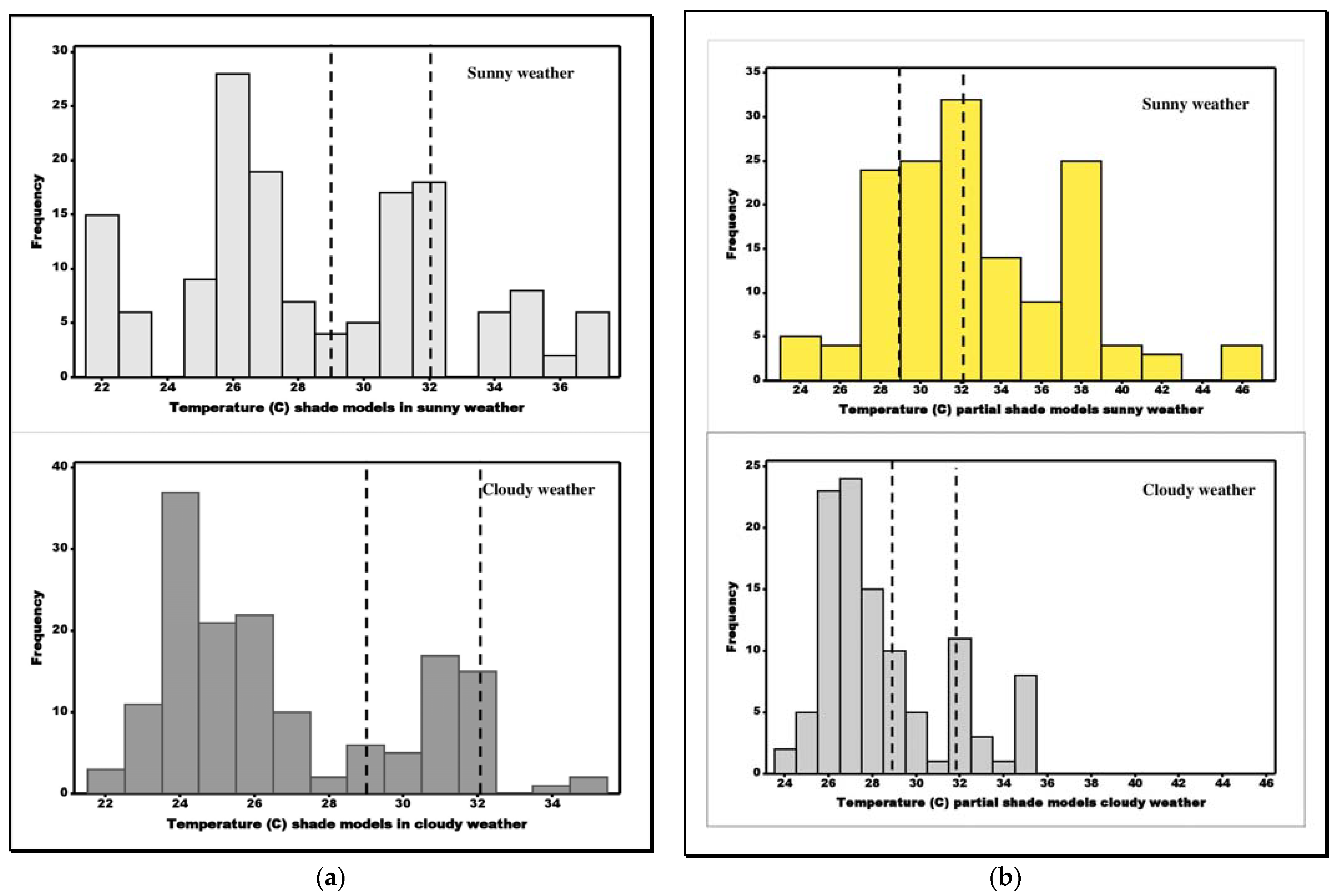
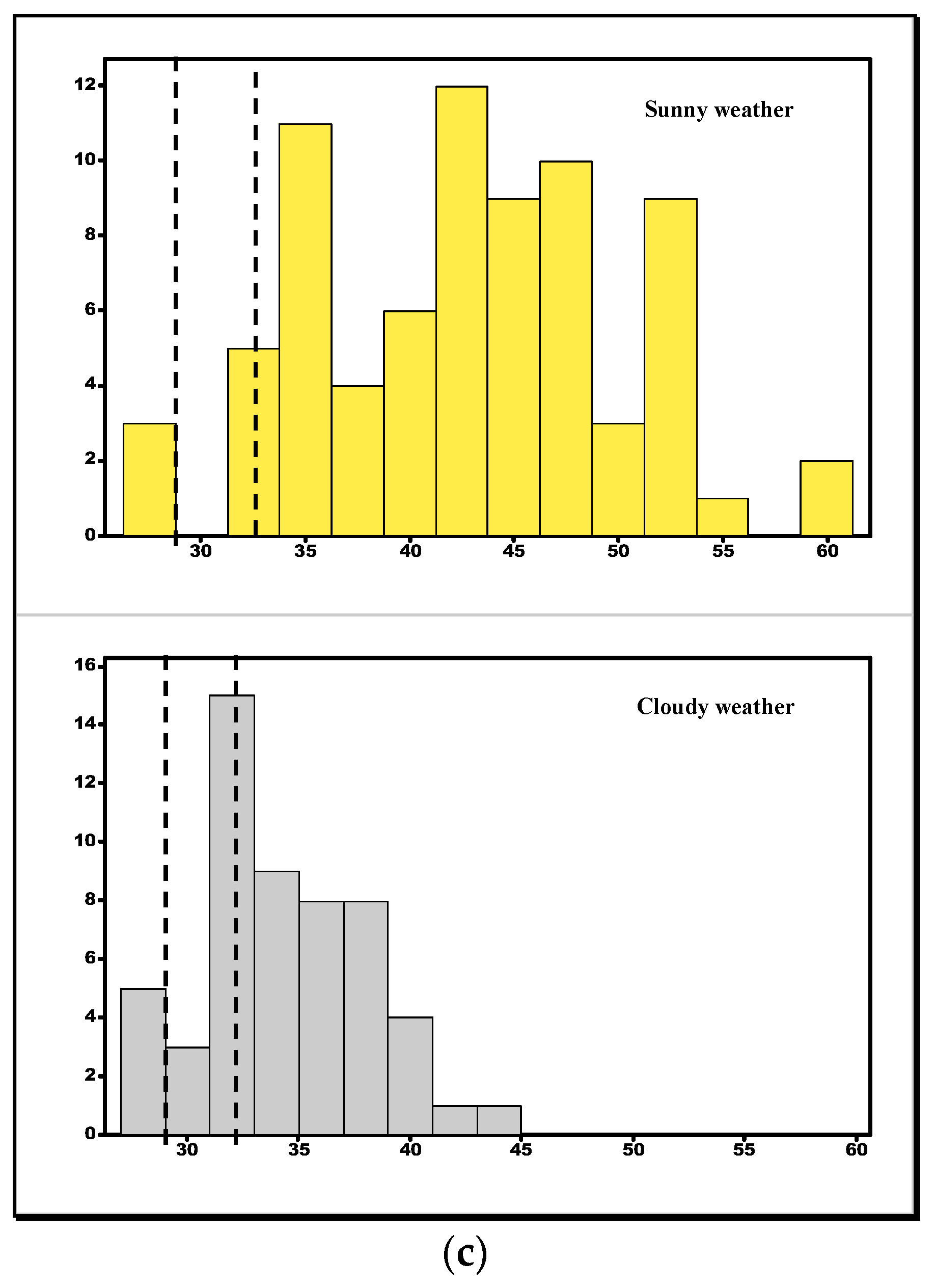
3.3. Comparison of Model Temperatures
3.4. Testing for Efficiency of Thermoregulation
4. Discussion
4.1. General Considerations
4.2. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Huey, R.B.; Slatkin, M. Costs and benefits of lizard thermoregulation. Q. Rev. Biol. 1976, 51, 363–384. [Google Scholar] [CrossRef] [PubMed]
- Hertz, P.E.; Huey, R.B.; Stevenson, R.D. Evaluating temperature regulation by field-active ectotherms: The fallacy of the inappropriate question. Am. Nat. 1993, 142, 796–818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bauwens, D.; Hertz, P.E.; Castilla, A.M. Thermoregulation in a lacertid lizard: The relative contributions of distinct behavioral mechanisms. Ecology 1996, 77, 1818–1830. [Google Scholar] [CrossRef]
- Castilla, A.M.; Van Damme, R.; Bauwens, D. Field body temperatures, mechanisms of thermoregulation and evolution of thermal characteristics in lacertid lizards. Nat. Croat. 1999, 8, 253–274. [Google Scholar]
- Grbac, I.; Bauwens, D. Constraints on temperature regulation in two sympatric Podacis lizards during autumn. Copeia 2001, 1, 178–186. [Google Scholar] [CrossRef]
- Blouin-Demers, G.; Nadeau, P. The cost-benefit model of thermoregulation does not predict lizard thermoregulatory behavior. Ecology 2005, 86, 560–566. [Google Scholar] [CrossRef]
- Dubois, Y.; Blouin-Demers, G.; Shipley, B.; Thomas, D. Thermoregulation and habitat selection in wood turtles Glyptemys insculpta: Chasing the sun slowly. J. Anim. Ecol. 2009, 78, 1023–1032. [Google Scholar] [CrossRef]
- Jayes, A.S.; Alexander, R.M. The gaits of chelonians: Walking techniques for very low speeds. J. Zool. 1980, 191, 353–378. [Google Scholar] [CrossRef]
- Branch, W.R. Preliminary observations on the ecology of the angulate tortoise (Chersina angulata) in Eastern Cape Province, South Africa. Amphibia-Reptilia 1984, 5, 43–55. [Google Scholar] [CrossRef]
- Swingland, I.R.; Frazier, J.G. The conflict between feeding and overheating in the Aldabran giant tortoise. In A Handbook of Biotelemetry and Radio Tracking; Amlaner, C.J., MacDonald, D.W., Eds.; Pergamon Press: Oxford, UK, 1979; pp. 611–615. [Google Scholar]
- Moulherat, S.; Delmas, V.; Slimani, T.; El Mouden, E.; Louzizi, T.; Lagarde, F.; Bonnet, X. How far can a tortoise walk in open habitat before overheating? Implications for conservation. J.Nat. Conserv. 2014, 22, 186–192. [Google Scholar] [CrossRef]
- Meek, R. Nutritional selection in Hermann`s tortoise Testudo hermanni, in Montenegro and Croatia. BCG Testudo 2010, 7, 88–95. [Google Scholar]
- Pulford, E.; Hailey, A.; Stubbs, D. Thermal relations of Testudo hermanni robertmertensi in S. France. Amphibia-Reptilia 1984, 5, 37–41. [Google Scholar]
- Meek, R. Thermoregulatory behaviour in a population of Hermann’s tortoise (Testudo hermanni) in southern Yugoslavia. Br. J. Herpetol. 1984, 6, 387–391. [Google Scholar]
- Meek, R. The thermal ecology of Hermann’s tortoise (Testudo hermanni) in summer and autumn in Yugoslavia. J. Zool. 1988, 215, 99–111. [Google Scholar] [CrossRef]
- Filippi, E.; Rugerio, L.; Capula, M.; Burke, R.L.; Luiselli, L. Population and Thermal Ecology of Testudo hermanni hermanni in the Tolfa Mountains of Central Italy. Chelonian Conserv. Biol. 2010, 9, 54–60. [Google Scholar] [CrossRef]
- Zimmerman, L.C.; O’Connor, M.P.; Kemp, S.A.; Spotila, S.R. Thermoregulation by desert tortoises (Gopherus agassizii) at the Desert Tortoise Conservation Center, Las Vegas, Nevada: Preliminary results. In Proceedings of the Symposium-Desert Tortoise Council, Acton, CA, USA, 1992; pp. 103–117. [Google Scholar]
- Vitt, L.J.; Sartorius, S.S. HOBO’s, Tidbits and lizard models: The utility of electronic devices in field studies of ectotherm thermoregulation. Funct. Ecol. 1999, 13, 670–674. [Google Scholar] [CrossRef]
- Meek, R. Thermoregulation and activity patterns in captive water dragons Physignathus cocincinus, in a naturalistic environment. Herpetol. J. 1999, 9, 137–146. [Google Scholar]
- Meek, R.; Jolley, E.; de Silva, A.; Goonewardene, S.; Drake, J.; Chalalochani, H.M.N.; Liyanage, P.; Abeysekera, T.S.; Mayadunna, M.D.I.P.K.; Somathilaka, S.A.U.S.; et al. Altitudinal differences in thermoregulatory behaviour in Calotes versicolor in the Knuckles region, Sri Lanka. The diversity of the Dumbara Mountains: The Knuckles Massif, Sri Lanka: With special reference to its herpetofauna. Lyriocephalus 2005, 6, 83–93. [Google Scholar]
- Shine, R.; Kearney, M. Field studies of reptile thermoregulation: How well do physical models predict operative temperatures? Funct. Ecol. 2001, 15, 282–288. [Google Scholar]
- Bakken, G.S. Measurement and Application of Operative and Standard Operative Temperature in Ecology. Am. Zool. 1992, 32, 194–216. Available online: https://www.jstor.org/stable/3883758 (accessed on 5 September 2022). [CrossRef]
- Meek, R.; Avery, R.A. Basking in the Australian Water Dragon Physignathus lesueurii; why do alpha males not respond to operative temperatures in the same way as adults and sub-adults? Amphibia-Reptilia 2008, 29, 257–262. [Google Scholar]
- Burić, M.; Milenković, M.; Ducic, M. The Specificities of the Climate of Danilovgrad (Montenegro). Glas. Srp. Geogr. Drus. 2019, 99, 19–28. Available online: http://www.doiserbia.nb.rs/Article.aspx?ID=0350-35931901019B#.Y8BKv33MKpo (accessed on 5 September 2022). [CrossRef]
- Cagle, F.R. A system of marking turtles for future identification. Copeia 1939, 1939, 170–173. [Google Scholar] [CrossRef]
- Stubbs, D.; Hailey, A.; Pulford, E.; Tyler, W. Population ecology of European tortoises: Review of field techniques. Amphibia-Reptilia 1984, 5, 57–68. [Google Scholar] [CrossRef]
- Willemsen, R.E.; Hailey, A. Sexual dimorphism of body size and shell shape in European tortoises. J. Zool. 2003, 260, 353–365. [Google Scholar] [CrossRef]
- Van Damme, R.; Bauwens, D.; Castilla, A.M.; Verheyen, R.F. Altitudinal variation of the thermal biology and running performance in the lizard Podarcis tiliguerta. Oecologia 1989, 80, 516–524. [Google Scholar] [CrossRef]
- Cherchi, M. Termoregulazione in Testudo hermanni. Bolletino Musei Inst. Biol. Univ. Genova 1956, 28, 9–87. [Google Scholar]
- Sears, M.W.; Angilletta, M.J.; Schuler, M.S.; Borchert, J.D.; Dilliplane, K.F.; Stegman, M.; Rusch, T.; Mitchell, W.A. Configuration of the thermal landscape determines thermoregulatory performance of ectotherms. Proc. Natl. Acad. Sci. USA 2016, 113, 10595–10600. [Google Scholar] [CrossRef] [Green Version]
- Troyer, K. Small differences in daytime body temperature affect digestion of natural food in a herbivorous lizard (Iguana iguana). Comp. Biochem. Physiol. 1987, 87A, 623–626. [Google Scholar] [CrossRef]
- Bjorndal, K.A. Fermentation in Reptiles and Amphibians. In Gastrointestinal Microbiology; Springer: Boston, MA, USA, 1997; pp. 199–230. [Google Scholar]
- Niu, C.; Zhang, T.; Sun, R. Food consumption and growth of juvenile Chinese soft-shelled turtles (Pelodiscus sinensis) in relation to body weight and water temperature. Asiat. Herpetol. Res. 1999, 8, 81–84. [Google Scholar]
- Mackie, R.I.; Rycyk, M.; Ruemmler, R.L.; Aminov, R.; Wikelski, M. Biochemical and microbiological evidence for fermentative digestion in free-living land iguanas (Conolophus pallidus) and marine iguanas (Amblyrhynchus cristatus) on the Galapagos archipelago. Physiol. Biochem. Zool. 2004, 77, 127–138. [Google Scholar] [CrossRef] [PubMed]
- Meek, R.; Inskeep, R. Aspects of the field biology of a population of Hermanns tortoise (Testudo hermanni) in southern Yugoslavia. Br. J. Herpetol. 1981, 6, 159–164. [Google Scholar]
- DeWitt, C.B. Precision of thermoregulation and its relation to environmental factors in the desert iguana, Dipsosaurus dorsalis. Physiol. Zool. 1967, 40, 49–66. [Google Scholar] [CrossRef]
- Grant, B.W.; Dunham, A.E. Thermally imposed time constraints on the activity of the desert lizard Sceloporus merriami. Ecology 1988, 69, 167. [Google Scholar] [CrossRef]
- Grant, B.W.; Dunham, A.E. Elevational covariation in environmental constraints and life histories of the desert lizard Sceloporus Merriami. Ecology 1990, 71, 1765–1776. [Google Scholar] [CrossRef]
- Haxhiu, I. Results of studies on the chelonians of Albania. Chelonian Conserv. Biol. 1995, 1, 324–326. [Google Scholar]
- Meek, R. Thermal loads experienced by a nesting female Testudo hermanni. Amphibia-Reptilia 1988, 9, 311–312. [Google Scholar] [CrossRef]
- Basson, C.H.; Levy, O.; Angilletta, M.J.; Clusella-Trullas, S. Lizards paid a greater opportunity cost to thermoregulate in a less heterogeneous environment. Funct. Ecol. 2017, 31, 856–865. [Google Scholar] [CrossRef] [Green Version]
- Hailey, A. Thermoregulation and activity metabolism in the armoured anguid Ophisaurus apodus. Br. J. Herpetol. 1984, 6, 391–398. [Google Scholar]
- Hailey, A.; Theophilidis, G. Cardiac response to stress and activity in the armoured legless lizard (Ophisaurus apodus): Comparison with snake and tortoise. Comp. Biochem. Physiol. 1987, 88, 201–206. [Google Scholar] [CrossRef]
- Meek, R. Field body temperature of the glass lizard Ophisaurus apodus in Yugoslavia. Amphib.-Reptil. 1986, 7, 43–49. [Google Scholar] [CrossRef]
- Christian, K.A.; Bedford, G.S.; Schultz, T.J. Energetic consequences of metabolic depression in tropical and temperate-zone lizards. Aust. J. Zool. 1999, 47, 133–141. [Google Scholar] [CrossRef]
- Rugiero, L.; Luiselli, L. Ecological modelling of habitat use and the annual activity patterns in an urban population of the tortoise, Testudo hermanni. Ital. J. Zool. 2006, 73, 219–225. [Google Scholar] [CrossRef]
- Cherchi, M. Uteriori richerchi sulla termoregulazione in Testudo hermanni. Bolletino Musei Inst. Biol. Univ. Genova 1960, 30, 35–60. (In Italian) [Google Scholar]
- Del Vecchio, S.; Burke, R.L.; Rugiero, L.; Capula, M.; Luiselli, L. The turtle is in the details: Microhabitat choice by Testudo hermanni is based on microscale plant distribution. Anim. Biol. 2011, 61, 249–261. [Google Scholar] [CrossRef] [Green Version]
- Stojadinović, D.; Milošević, Đ.; Sretić, K.; Cvetković, M.; Jovanović, T.; Jovanović, B.; Crnobrnja-Isailović, J.M. Activity patterns and habitat preference of eastern Hermann’s tortoise (Testudo hermanni boettgeri) in Serbia. Turk. J. Zool. 2017, 41, 1036–1044. [Google Scholar] [CrossRef]
- Vujović, A.; Iković, V.; Golubović, A.; Đorđević, S.; Pešić, V.; Tomović, L.J. Effects of fires and roadkills on the isolated population of Testudo hermanni Gmelin, 1789 (Reptilia: Testudinidae) in central Montenegro. Acta Zool. Bulg. 2015, 67, 75–84. [Google Scholar]
- Huey, R.B.; Kearney, M.R.; Krockenberger, A.; Holtum, A.M.; Jess, M.; Williams, S.E. Predicting organismal vulnerability to climate warming: Roles of behaviour, physiology and adaptation. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2012, 367, 1665–1679. [Google Scholar] [CrossRef] [Green Version]
- Park Williams, A.; Allen, C.D.; Macalady, A.K.; Griffin, D.; Woodhouse, C.A.; Meko, D.M.; Swetnam, T.W.; Rauscher, S.A.; Seager, R.; Grissino-Mayer, H.D. Temperature as a potent driver of regional forest drought stress and tree mortality. Nat. Clim. Chang 2013, 3, 292–297. [Google Scholar] [CrossRef]
- Luiselli, L.; Vignoli, L.; Rugiero, L.; Meek, R. Declining occupancy rates in the hibernacula of aspic vipers (Vipera aspis) in Italy and France; evidence for climatic effects? Herpetol. J. 2018, 28, 137–142. [Google Scholar]
- Kearney, M.; Shine, R.; Porter, W.P. The potential for behavioral thermoregulation to buffer “cold-blooded” animals against climate warming. Proc. Natl. Acad. Sci. USA 2009, 106, 3835–3840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westerling, A.L. Increasing western US forest wildfire activity: Sensitivity to changes in the timing of spring. Phil. Trans. R. Soc. B 2016, 371, 20150178. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vujović, A.; Pešić, V.; Meek, R. Living in a Thermally Diverse Environment: Field Body Temperatures and Thermoregulation in Hermann’s Tortoise, Testudo hermanni, in Montenegro. Conservation 2023, 3, 59-70. https://doi.org/10.3390/conservation3010005
Vujović A, Pešić V, Meek R. Living in a Thermally Diverse Environment: Field Body Temperatures and Thermoregulation in Hermann’s Tortoise, Testudo hermanni, in Montenegro. Conservation. 2023; 3(1):59-70. https://doi.org/10.3390/conservation3010005
Chicago/Turabian StyleVujović, Ana, Vladimir Pešić, and Roger Meek. 2023. "Living in a Thermally Diverse Environment: Field Body Temperatures and Thermoregulation in Hermann’s Tortoise, Testudo hermanni, in Montenegro" Conservation 3, no. 1: 59-70. https://doi.org/10.3390/conservation3010005