
Pre-Test Manipulation by Intraperitoneal Saline Injection with or without Isoflurane Pre-Treatment Does Not Influence the Outcome of Social Test in Male Mice

Abstract
:1. Introduction
2. Results
3. Discussion
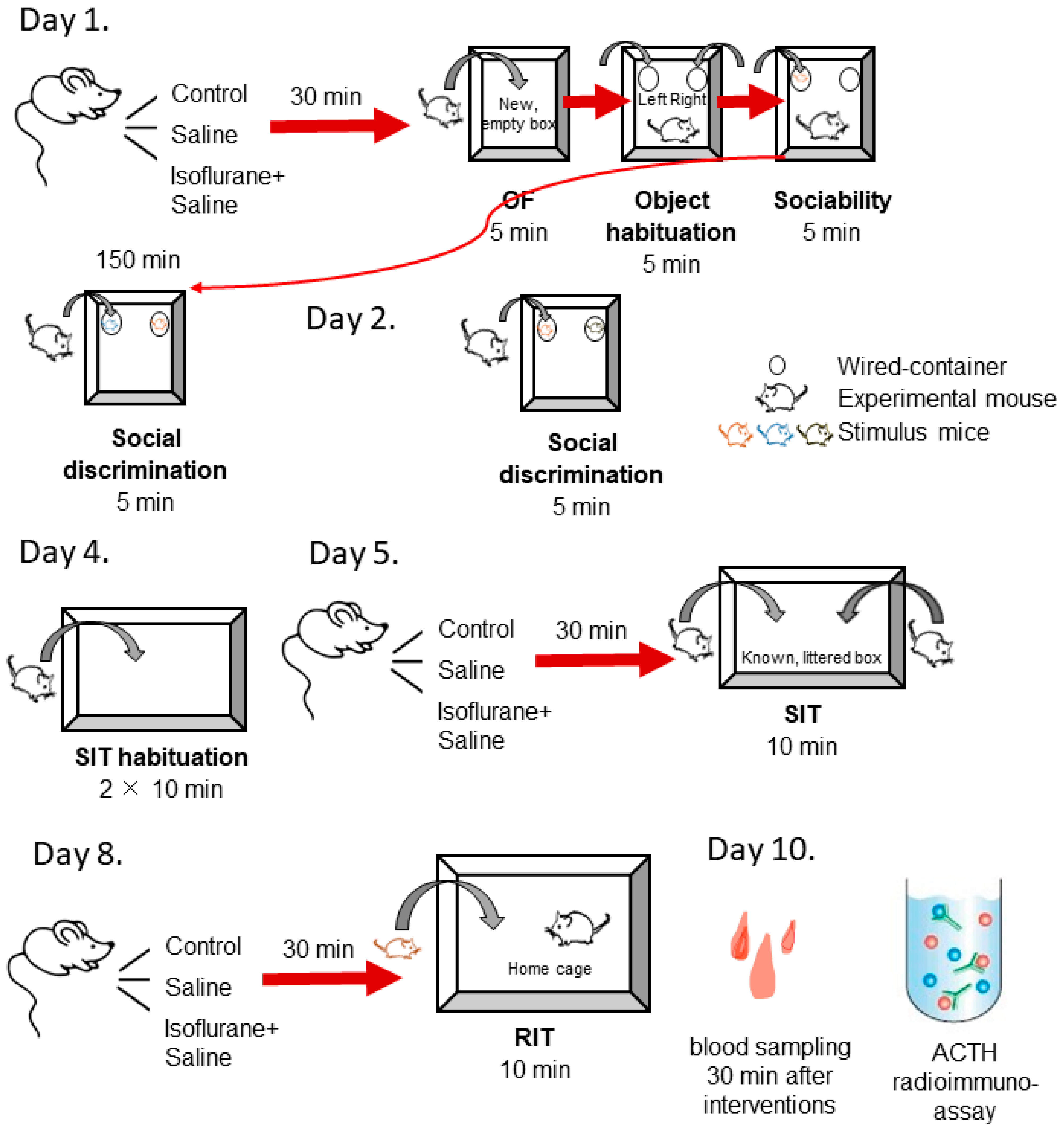
4. Materials and Methods
4.1. Animals
4.2. Behavioural Testing
4.2.1. Sociability Test
4.2.2. Social Interaction Test (SIT)
4.2.3. Resident–Intruder Test (RIT)
4.3. Stress Hormone Measurements
4.4. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lewis, D.I. Animal experimentation: Implementation and application of the 3Rs. Emerg. Top Life Sci. 2019, 3, 675–679. [Google Scholar] [PubMed]
- Laferriere, C.A.; Pang, D.S. Review of Intraperitoneal Injection of Sodium Pentobarbital as a Method of Euthanasia in Laboratory Rodents. J. Am. Assoc. Lab. Anim. Sci. 2020, 59, 254–263. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Cruz, L.; Bussey, T.J.; Saksida, L.M.; Heath, C.J. Using touchscreen-delivered cognitive assessments to address the principles of the 3Rs in behavioral sciences. Lab. Anim. (NY) 2021, 50, 174–184. [Google Scholar] [CrossRef] [PubMed]
- Turner, P.V.; Pang, D.S.; Lofgren, J.L. A Review of Pain Assessment Methods in Laboratory Rodents. Comp. Med. 2019, 69, 451–467. [Google Scholar] [CrossRef]
- Van der Mierden, S.; Leenaars, C.H.C.; Boyle, E.C.; Ripoli, F.L.; Gass, P.; Durst, M.; Goerlich-Jansson, V.C.; Jirkof, P.; Keubler, L.M.; Lewejohann, L. Measuring endogenous corticosterone in laboratory mice—A mapping review, meta-analysis, and open source database. ALTEX 2021, 38, 111–122. [Google Scholar] [CrossRef]
- Yarushkina, N.I. The role of hypothalamo-hypophyseal-adrenocortical system hormones in controlling pain sensitivity. Neurosci. Behav. Physiol. 2008, 38, 759–766. [Google Scholar] [CrossRef]
- Jacobsen, K.R.; Kalliokoski, O.; Teilmann, A.C.; Hau, J.; Abelson, K.S. The effect of isoflurane anaesthesia and vasectomy on circulating corticosterone and ACTH in BALB/c mice. Gen. Comp. Endocrinol. 2012, 179, 406–413. [Google Scholar] [CrossRef]
- Romero, L.M.; Fairhurst, G.D. Measuring corticosterone in feathers: Strengths, limitations, and suggestions for the future. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2016, 202, 112–122. [Google Scholar] [CrossRef] [Green Version]
- Scorrano, F.; Carrasco, J.; Pastor-Ciurana, J.; Belda, X.; Rami-Bastante, A.; Bacci, M.L.; Armario, A. Validation of the long-term assessment of hypothalamic-pituitary-adrenal activity in rats using hair corticosterone as a biomarker. FASEB J. 2015, 29, 859–867. [Google Scholar] [CrossRef] [Green Version]
- Rowland, N.E.; Toth, L.A. Analytic and Interpretational Pitfalls to Measuring Fecal Corticosterone Metabolites in Laboratory Rats and Mice. Comp. Med. 2019, 69, 337–349. [Google Scholar] [CrossRef]
- Palme, R. Non-invasive measurement of glucocorticoids: Advances and problems. Physiol. Behav. 2019, 199, 229–243. [Google Scholar] [CrossRef] [PubMed]
- Armario, A.; Labad, J.; Nadal, R. Focusing attention on biological markers of acute stressor intensity: Empirical evidence and limitations. Neurosci. Biobehav. Rev. 2020, 111, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Armario, A. The hypothalamic-pituitary-adrenal axis: What can it tell us about stressors? CNS Neurol. Disord. Drug Targets 2006, 5, 485–501. [Google Scholar] [CrossRef] [PubMed]
- Hurtubise, J.L.; Howland, J.G. Effects of stress on behavioral flexibility in rodents. Neuroscience 2017, 345, 176–192. [Google Scholar] [CrossRef]
- Haller, J.; Varga, B.; Ledent, C.; Barna, I.; Freund, T.F. Context-dependent effects of CB1 cannabinoid gene disruption on anxiety-like and social behaviour in mice. Eur. J. Neurosci. 2004, 19, 1906–1912. [Google Scholar] [CrossRef]
- Carnevali, L.; Montano, N.; Tobaldini, E.; Thayer, J.F.; Sgoifo, A. The contagion of social defeat stress: Insights from rodent studies. Neurosci. Biobehav. Rev. 2020, 111, 12–18. [Google Scholar] [CrossRef]
- Grimm, C.; Balsters, J.H.; Zerbi, V. Shedding Light on Social Reward Circuitry: (Un)common Blueprints in Humans and Rodents. Neuroscientist 2021, 27, 159–183. [Google Scholar] [CrossRef]
- Eslinger, P.J.; Anders, S.; Ballarini, T.; Boutros, S.; Krach, S.; Mayer, A.V.; Moll, J.; Newton, T.L.; Schroeter, M.L.; de Oliveira-Souza, R.; et al. The neuroscience of social feelings: Mechanisms of adaptive social functioning. Neurosci. Biobehav. Rev. 2021, 128, 592–620. [Google Scholar] [CrossRef]
- Ibanez, A.; Garcia, A.M.; Esteves, S.; Yoris, A.; Munoz, E.; Reynaldo, L.; Pietto, M.L.; Adolfi, F.; Manes, F. Social neuroscience: Undoing the schism between neurology and psychiatry. Soc. Neurosci. 2018, 13, 1–39. [Google Scholar] [CrossRef]
- Desmarais, P.; Lanctot, K.L.; Masellis, M.; Black, S.E.; Herrmann, N. Social inappropriateness in neurodegenerative disorders. Int. Psychogeriatr. 2018, 30, 197–207. [Google Scholar] [CrossRef]
- Szabo, S.; Yoshida, M.; Filakovszky, J.; Juhasz, G. “Stress” is 80 Years Old: From Hans Selye Original Paper in 1936 to Recent Advances in GI Ulceration. Curr. Pharm. Des. 2017, 23, 4029–4041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fazekas, C.L.; Bellardie, M.; Torok, B.; Sipos, E.; Toth, B.; Baranyi, M.; Sperlágh, B.; Dobos-Kovács, M.; Chaillou, E.; Zelena, D. Pharmacogenetic excitation of the median raphe region affects social and depressive-like behavior and core body temperature in male mice. Life Sci. 2021, 286, 120037. [Google Scholar] [CrossRef] [PubMed]
- Chaves, T.; Torok, B.; Fazekas, C.L.; Correia, P.; Sipos, E.; Várkonyi, D.; Hellinger, Á.; Erk, D.; Zelena, D. Median raphe region GABAergic neurons contribute to social interest in mouse. Life Sci. 2021, 289, 120223. [Google Scholar] [CrossRef] [PubMed]
- Camats-Perna, J.; Kalaba, P.; Ebner, K.; Sartori, S.B.; Vuyyuru, H.; Aher, N.Y.; Dragačević, V.; Singewald, N.; Engelmann, M.; Lubec, G. Differential Effects of Novel Dopamine Reuptake Inhibitors on Interference With Long-Term Social Memory in Mice. Front. Behav. Neurosci. 2019, 13, 63. [Google Scholar] [CrossRef]
- Sterrenburg, L.; Borch, A.; Peeters, B.W.; Pinter, O.; Zelena, D.; Roubos, E.W.; Kozicz, T. Acute ether stress differentially affects corticotropin-releasing factor and urocortin 1 in the Brattleboro rat. Brain Res. 2011, 1398, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Makara, G.B.; Domokos, A.; Mergl, Z.; Csabai, K.; Barna, I.; Zelena, D. Gender-specific regulation of the hypothalamo-pituitary-adrenal axis and the role of vasopressin during the neonatal period. Ann. N. Y. Acad. Sci. 2008, 1148, 439–445. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. World Health Organization Model List of Essential Medicines: 21st List 2019; No. WHO/MVP/EMP/IAU/2019.06; World Health Organization: Geneva, Switzerland, 2019; hdl:10665/325771.
- Richardson, C.A.; Flecknell, P.A. Anaesthesia and post-operative analgesia following experimental surgery in laboratory rodents: Are we making progress? Altern. Lab. Anim. 2005, 33, 119–127. [Google Scholar] [CrossRef]
- Mellon, R.D.; Simone, A.F.; Rappaport, B.A. Use of anesthetic agents in neonates and young children. Anesth. Analg. 2007, 104, 509–520. [Google Scholar] [CrossRef]
- Chirico, M.T.T.; Guedes, M.R.; Vieira, L.G.; Reis, T.O.; Dos Santos, A.M.; Souza, A.B.F.; Ribeiro, I.M.L.; Noronha, S.I.; Nogueira, K.O.; Oliveira, L.A.; et al. Lasting effects of ketamine and isoflurane administration on anxiety- and panic-like behavioral responses in Wistar rats. Life Sci. 2021, 276, 119423. [Google Scholar] [CrossRef]
- Hamden, J.E.; Salehzadeh, M.; Gray, K.M.; Forys, B.J.; Soma, K.K. Isoflurane stress induces glucocorticoid production in mouse lymphoid organs. J. Endocrinol. 2021, 251, 137–148. [Google Scholar] [CrossRef]
- Filaretova, L.; Morozova, O.; Laszlo, F.; Morschl, E.; Zelena, D. Does chronic stress enhance the risk of diseases? Endocr. Regul. 2013, 47, 177–188. [Google Scholar] [CrossRef] [PubMed]
- Zelena, D.; Filaretova, L.; Mergl, Z.; Barna, I.; Toth, Z.E.; Makara, G.B. Hypothalamic paraventricular nucleus, but not vasopressin, participates in chronic hyperactivity of the HPA axis in diabetic rats. Am. J. Physiol. Endocrinol. Metab. 2006, 290, E243–E250. [Google Scholar] [CrossRef] [PubMed]
- Zelena, D.; Barna, I.; Mlynarik, M.; Gupta, O.P.; Jezova, D.; Makara, G.B. Stress symptoms induced by repeated morphine withdrawal in comparison to other chronic stress models in mice. Neuroendocrinology 2005, 81, 205–215. [Google Scholar] [CrossRef] [PubMed]
- Bazhan, N.; Zelena, D. Food-intake regulation during stress by the hypothalamo-pituitary-adrenal axis. Brain Res. Bull. 2013, 95, 46–53. [Google Scholar] [CrossRef] [Green Version]
- Bajwa, N.M.; Lee, J.B.; Halavi, S.; Hartman, R.E.; Obenaus, A. Repeated isoflurane in adult male mice leads to acute and persistent motor decrements with long-term modifications in corpus callosum microstructural integrity. J. Neurosci. Res. 2019, 97, 332–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yonezaki, K.; Uchimoto, K.; Miyazaki, T.; Asakura, A.; Kobayashi, A.; Takase, K.; Goto, T. Postanesthetic effects of isoflurane on behavioral phenotypes of adult male C57BL/6J mice. PLoS ONE 2015, 10, e0122118. [Google Scholar] [CrossRef] [Green Version]
- Neudecker, V.; Perez-Zoghbi, J.F.; Coleman, K.; Neuringer, M.; Robertson, N.; Bemis, A.; Glickman, B.; Schenning, K.J.; Fair, D.A.; Martin, L.D.; et al. Infant isoflurane exposure affects social behaviours, but does not impair specific cognitive domains in juvenile non-human primates. Br. J. Anaesth. 2021, 126, 486–499. [Google Scholar] [CrossRef]
- Coleman, K.; Robertson, N.D.; Dissen, G.A.; Neuringer, M.D.; Martin, L.D.; Cuzon Carlson, V.C.; Kroenke, C.; Fair, D.; Brambrink, A.M. Isoflurane Anesthesia Has Long-term Consequences on Motor and Behavioral Development in Infant Rhesus Macaques. Anesthesiology 2017, 126, 74–84. [Google Scholar] [CrossRef] [Green Version]
- Diana, P.; Joksimovic, S.M.; Faisant, A.; Jevtovic-Todorovic, V. Early exposure to general anesthesia impairs social and emotional development in rats. Mol. Neurobiol. 2020, 57, 41–50. [Google Scholar] [CrossRef]
- Maloney, S.E.; Yuede, C.M.; Creeley, C.E.; Williams, S.L.; Huffman, J.N.; Taylor, G.T.; Noguchi, K.N.; Wozniak, D.F. Repeated neonatal isoflurane exposures in the mouse induce apoptotic degenerative changes in the brain and relatively mild long-term behavioral deficits. Sci. Rep. 2019, 9, 2779. [Google Scholar] [CrossRef] [Green Version]
- Lee, B.H.; Chan, J.T.; Kraeva, E.; Peterson, K.; Sall, J.W. Isoflurane exposure in newborn rats induces long-term cognitive dysfunction in males but not females. Neuropharmacology 2014, 83, 9–17. [Google Scholar] [CrossRef] [Green Version]
- Sasaki Russell, J.M.; Chinn, G.A.; Maharjan, D.; Eichbaum, Y.; Sall, J.W. Female rats are more vulnerable to lasting cognitive impairment after isoflurane exposure on postnatal day 4 than 7. Br. J. Anaesth. 2019, 122, 490–499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finsterwald, C.; Alberini, C.M. Stress and glucocorticoid receptor-dependent mechanisms in long-term memory: From adaptive responses to psychopathologies. Neurobiol. Learn Mem. 2014, 112, 17–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kruk, M.R.; Halasz, J.; Meelis, W.; Haller, J. Fast positive feedback between the adrenocortical stress response and a brain mechanism involved in aggressive behavior. Behav. Neurosci. 2004, 118, 1062–1070. [Google Scholar] [CrossRef]
- Zelena, D.; Kiem, D.T.; Barna, I.; Makara, G.B. Alpha 2-adrenoreceptor subtypes regulate ACTH and beta-endorphin secretions during stress in the rat. Psychoneuroendocrinology 1999, 24, 333–343. [Google Scholar] [CrossRef] [PubMed]
- Nishiyama, T. Effects of repeat exposure to inhalation anesthetics on liver and renal function. J. Anaesthesiol. Clin. Pharmacol. 2013, 29, 83–87. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | Saline | Isoflurane | F | p | |||
|---|---|---|---|---|---|---|---|
| General data | Body weight (g) | Initial | 25.28 ± 0.33 | 25.53 ± 0.59 | 25.17 ± 0.47 | 0.22 | 0.80 |
| Weight gain | 0.05 ± 0.35 | 0.58 ± 0.72 | 0.27 ± 0.87 | 0.17 | 0.84 | ||
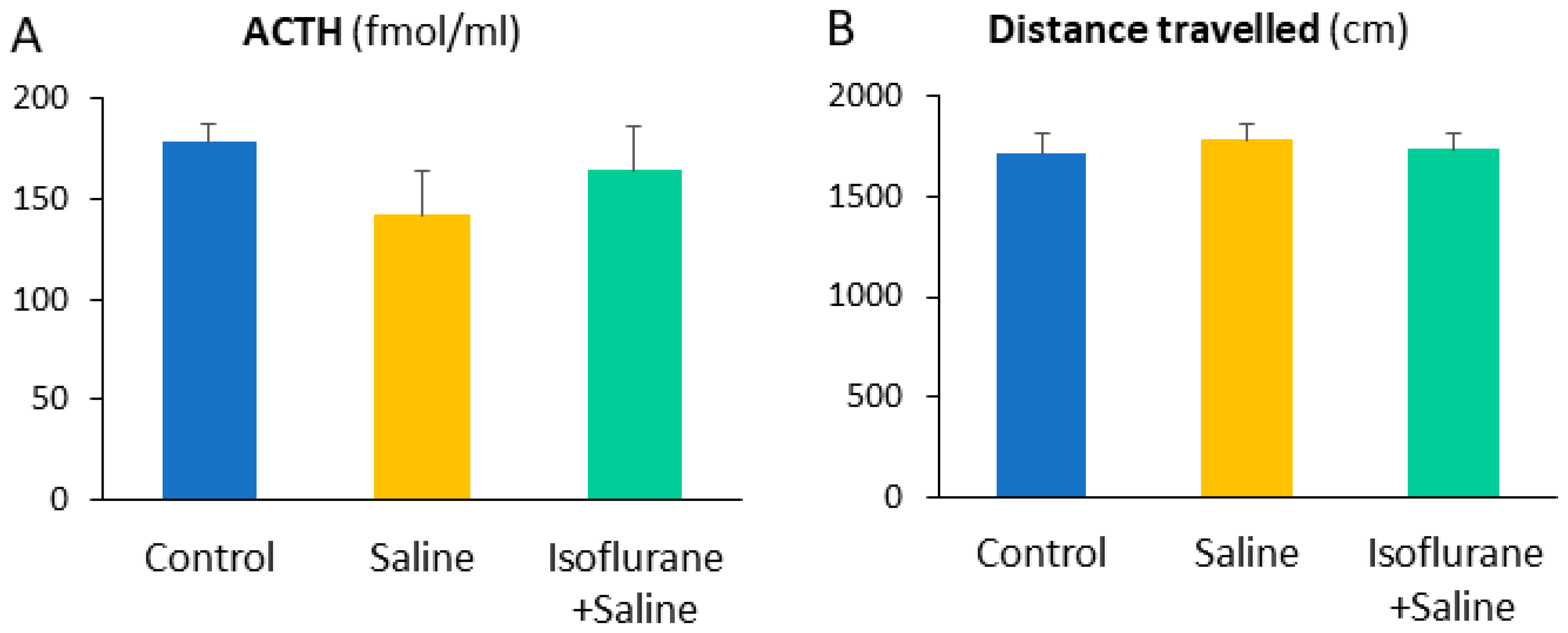
| ACTH (fmol/mL) | 178.38 ± 8.81 | 142.36 ± 22.30 | 163.63 ± 22.46 | 1.04 | 0.37 | ||
| Sociability | Open field | Distance | 1714.34 ± 96.88 | 1780.49 ± 82.21 | 1738.79 ± 76.48 | 0.15 | 0.86 |
| Object habituation | Left% | 8.1 ± 3.00 | 5.78 ± 0.93 | 7.07 ± 2.54 | 0.08 | 0.92 | |
| Right% | 6.46 ± 1.72 | 7.44 ± 1.40 | 7.96 ± 2.18 | ||||
| Sociability | Object% | 7.25 ± 1.88 | 4.84 ± 1.19 | 8.09 ± 1.99 | 0.65 | 0.53 | |
| Mice% | 21.71 ± 5.28 | 15.01 ± 3.74 | 17.25 ± 5.51 | ||||
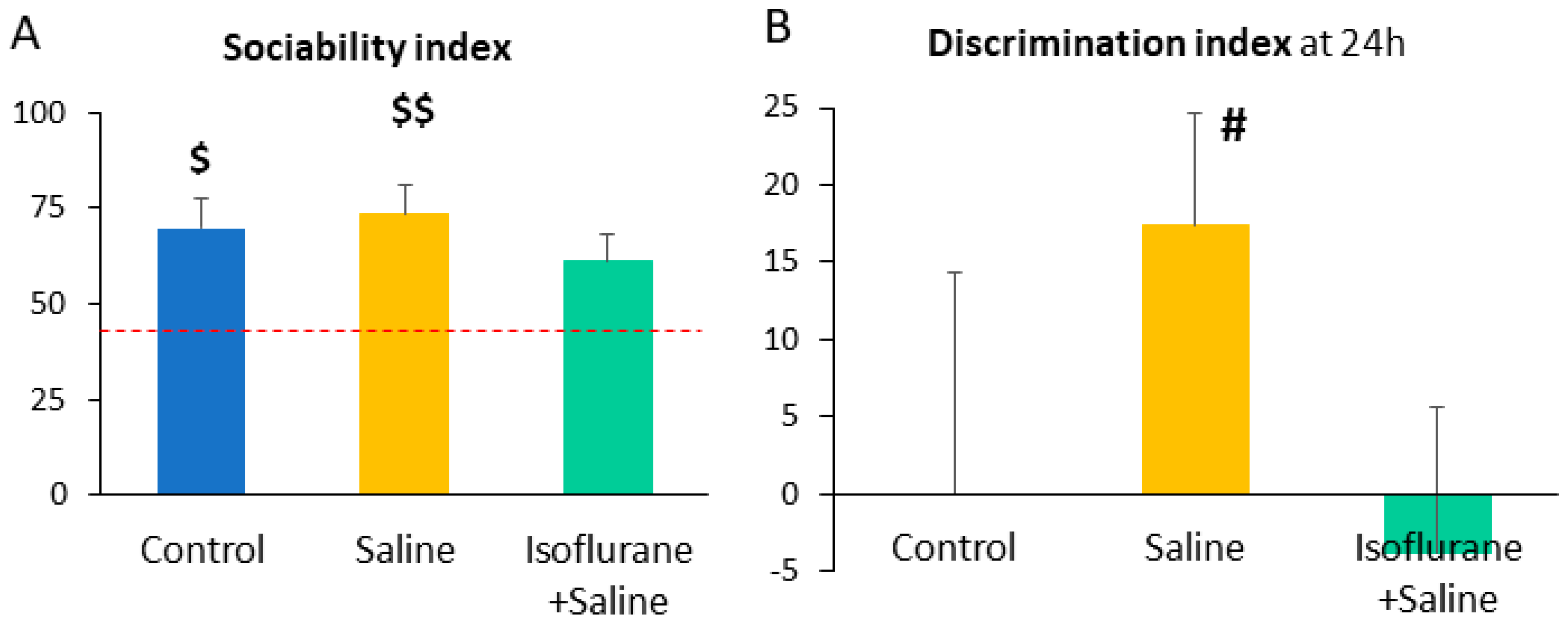
| SI | 69.90 ± 7.74 | 73.51 ± 7.55 | 61.08 ± 7.36 | 0.66 | 0.52 | ||
| Social discrimination | 2.5 h Old% | 14.14 ± 2.95 | 12.31 ± 3.25 | 14.03 ± 2.25 | 0.05 | 0.95 | |
| 2.5 h New% | 11.60 ± 2.71 | 15.76 ± 3.66 | 12.16 ± 2.70 | ||||
| 2.5 h DI | −6.81 ± 10.76 | 9.32 ± 11.02 | −12.84 ± 8.74 | 1.26 | 0.30 | ||
| 24 h Old% | 12.68 ± 3.27 | 12.42 ± 2.66 | 10.78 ± 1.56 | 0.48 | 0.63 | ||
| 24 h New% | 13.75 ± 2.88 | 16.01 ± 2.54 | 11.41 ± 2.20 | ||||
| 24 h DI | −0.03 ± 14.42 | 17.44 ± 7.25 | −3.74 ± 9.39 | 1.00 | 0.38 | ||
| Social interaction test (time% of 10 min) | Friendly social interaction | 19.05 ± 2.50 | 19.61 ± 1.94 | 24.00 ± 2.88 | 1.20 | 0.32 | |
| Aggression | 3.68 ± 1.71 | 7.74 ± 2.91 | 2.06 ± 0.83 | 2.11 | 0.14 | ||
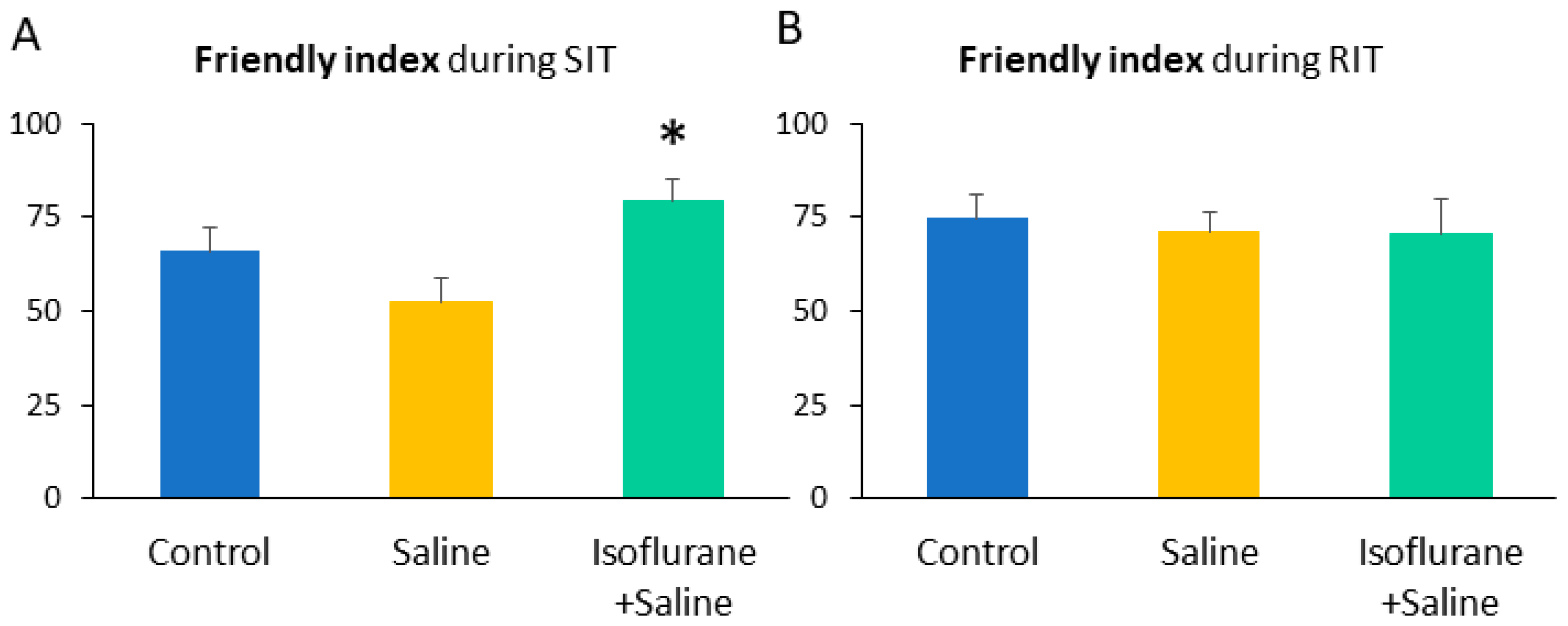
| Friendly index | 65.93 ± 6.14 | 52.61 ± 6.07 | 79.11 ± 5.91 * | 4.81 | 0.02 | ||
| Resident–intruder test (time% of 10 min) | Aggression | 5.77 ± 4.76 | 7.49 ± 7.97 | 3.31 ± 3.78 | 1.21 | 0.32 | |
| Friendly social interaction | 23.23 ± 10.34 | 23.42 ± 10.85 | 18.92 ± 11.59 | 0.51 | 0.61 | ||
| Friendly index | 74.75 ± 6.19 | 71.32 ± 5.28 | 70.63 ± 9.01 | 0.11 | 0.90 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pomogyi, F.; Török, B.; Zelena, D. Pre-Test Manipulation by Intraperitoneal Saline Injection with or without Isoflurane Pre-Treatment Does Not Influence the Outcome of Social Test in Male Mice. Stresses 2023, 3, 340-350. https://doi.org/10.3390/stresses3010025
Pomogyi F, Török B, Zelena D. Pre-Test Manipulation by Intraperitoneal Saline Injection with or without Isoflurane Pre-Treatment Does Not Influence the Outcome of Social Test in Male Mice. Stresses. 2023; 3(1):340-350. https://doi.org/10.3390/stresses3010025
Chicago/Turabian StylePomogyi, Flóra, Bibiána Török, and Dóra Zelena. 2023. "Pre-Test Manipulation by Intraperitoneal Saline Injection with or without Isoflurane Pre-Treatment Does Not Influence the Outcome of Social Test in Male Mice" Stresses 3, no. 1: 340-350. https://doi.org/10.3390/stresses3010025