


Expression Profile of Selected Genes Involved in Na+ Homeostasis and In Silico miRNA Identification in Medicago sativa and Medicago arborea under Salinity Stress

 ,
,
Abstract
:1. Introduction
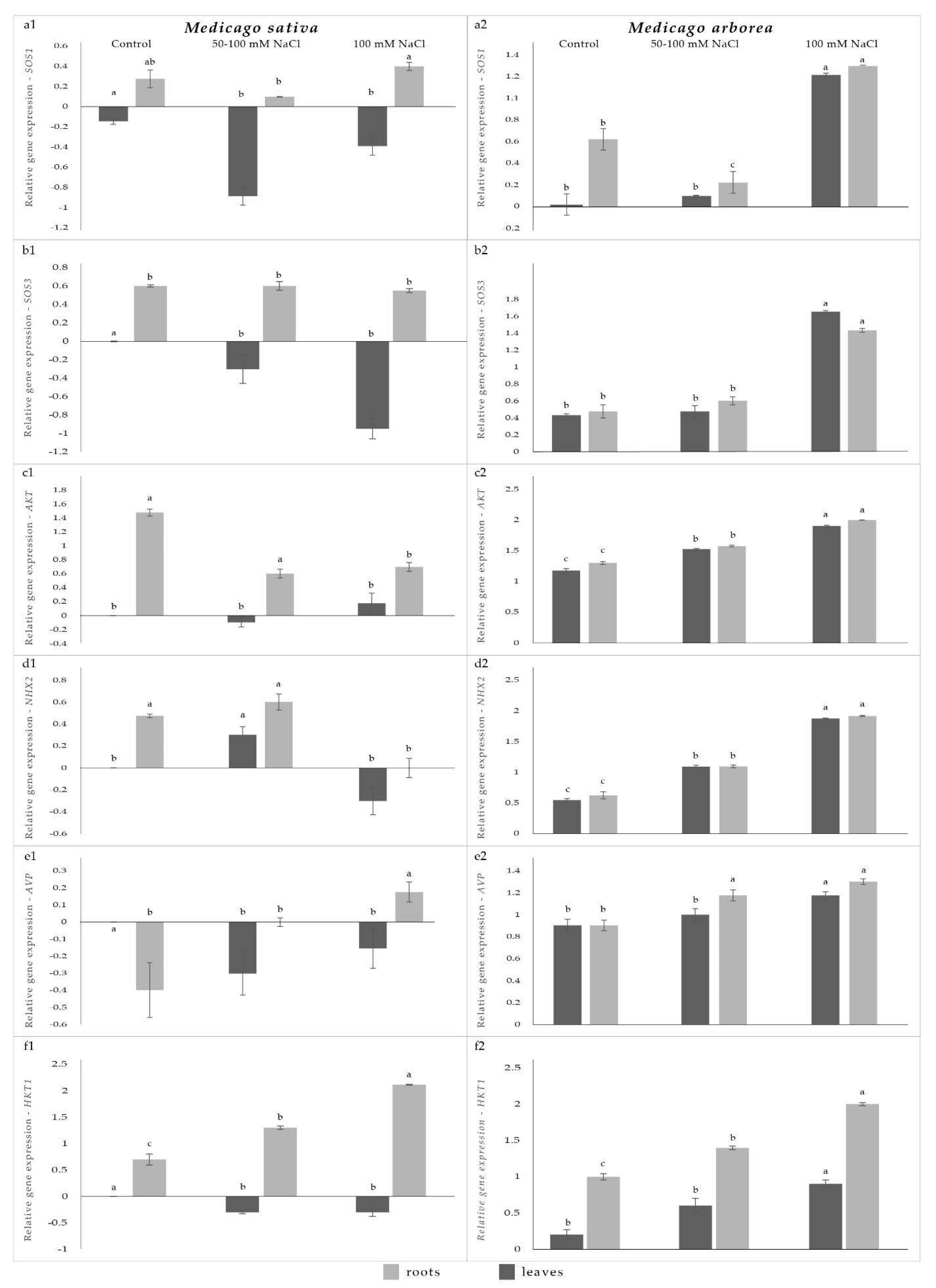
2. Results and Discussion
2.1. Gene Expression Levels

2.2. miRNA In Silico Analysis
3. Materials and Methods
3.1. Plant Material and Seed Pretreatment
3.2. Growth Conditions and Salt Stress Treatments
3.3. RNA Isolation and cDNA Synthesis
3.4. Real-Time PCR Experiments
3.5. Statistical Analysis
3.6. In Silico miRNA Analysis
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, C.M.; Zhang, J.L.; Liu, X.S.; Li, Z.; Wu, G.Q.; Cai, J.Y.; Flowers, T.J.; Wang, S.M. Puccinellia tenuiflora maintains a low Na+ level under salinity by limiting unidirectional Na+ influx resulting in a high selectivity for K+ over Na+. Plant Cell Environ. 2009, 32, 486–496. [Google Scholar] [CrossRef] [PubMed]
- Aisset Amel, M.Z. Effect of salinity and water stress on the germination of Medicago arborea L. seeds. J. Appl. Biol. Sci. 2016, 6, 113–121. [Google Scholar]
- Tani, E.; Sarri, E.; Goufa, M.; Asimakopoulou, G.; Psychogiou, M.; Bingham, E.; Skaracis, G.N.; Abraham, E.M. Seedling growth and transcriptional responses to salt shock and stress in Medicago sativa L.; Medicago arborea L.; and their hybrid (Alborea). Agronomy 2018, 8, 231. [Google Scholar] [CrossRef] [Green Version]
- Volkov, V.; Amtmann, A. Thellungiella halophila, a salt-tolerant relative of Arabidopsis thaliana, has specific root ion-channel features supporting K+/Na+ homeostasis under salinity stress. Plant J. 2006, 48, 342–353. [Google Scholar] [CrossRef]
- Pandita, D. Role of miRNA technology and miRNAs in abiotic and biotic stress resilience. In Plant Perspectives to Global Climate Changes; Aftab, T., Roychoudhury, A., Eds.; Academic Press: Cambridge, MA, USA, 2022; pp. 303–330. [Google Scholar]
- Zelm, E.V.; Zhang, Y.; Testerink, C. Salt tolerance mechanisms of plants. Annu. Rev. Plant Biol. 2020, 71, 403–433. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.K. Salt and drought stress signal transduction in plants. Annu. Rev. Plant Biol. 2002, 53, 247–273. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.Q.; Niu, Y.D.; Huridu, H.; Hao, J.F.; Qi, Z.; Hasi, A. Salicornia europaea L. Na+/H+ antiporter gene improves salt tolerance in transgenic alfalfa (Medicago sativa L.). Genet. Mol. Res. 2014, 13, 5350–5360. [Google Scholar] [CrossRef]
- Wu, G.-Q.; Wang, J.-L.; Li, S.-J. Genome-wide identification of Na+/H+ antiporter (NHX) genes in sugar beet (Beta vulgaris L.) and their regulated expression under salt stress. Genes 2019, 10, 401. [Google Scholar] [CrossRef] [Green Version]
- Sandhu, D.; Cornacchione, M.V.; Ferreira, J.F.S.; Suarez, D.L. Variable salinity responses of 12 alfalfa genotypes and comparative expression analyses of salt-response genes. Sci. Rep. 2017, 7, 42958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toranj, S.; Aliabad, K.K.; Abbaspour, H.; Saeedpour, A. Effect of salt stress on the genes expression of the vacuolar H+ -pyrophosphatase and Na+/H+ antiporter in Rubia tinctorum. Mol. Biol. Rep. 2020, 47, 235–245. [Google Scholar] [CrossRef] [PubMed]
- Sunkar, R.; Chinnusamy, V.; Zhu, J.; Zhu, J.K. Small RNAs as big players in plant abiotic stress responses and nutrient deprivation. Trends Plant Sci. 2007, 12, 301–309. [Google Scholar] [CrossRef]
- Xu, T.; Zhang, L.; Yang, Z.; Wei, Y.; Dong, T. Identification and functional characterization of plant miRNA under salt stress shed light on salinity resistance improvement through miRNA manipulation in crops. Front. Plant Sci. 2021, 12, 665439. [Google Scholar] [CrossRef]
- Zhang, Y.M.; Zhang, H.M.; Liu, Z.H.; Li, H.C.; Guo, X.L.; Li, G.L. The wheat NHX antiporter gene TaNHX2 confers salt tolerance in transgenic alfalfa by increasing the retention capacity of intracellular potassium. Plant Mol. Biol. 2015, 87, 317–327. [Google Scholar] [CrossRef]
- Barciszewska-Pacak, M.; Milanowska, K.; Knop, K.; Bielewicz, D.; Nuc, P.; Plewka, P.; Pacak, A.M.; Vazquez, F.; Karlowski, W.; Jarmolowski, A.; et al. Arabidopsis microRNA expression regulation in a wide range of abiotic stress responses. Front. Plant Sci. 2015, 6, 410. [Google Scholar] [CrossRef]
- Cao, X.; Wu, Z.; Jiang, F.; Zhou, R.; Yang, Z. Identification of chilling stress-responsive tomato microRNAs and their target genes by high-throughput sequencing and degradome analysis. BMC Genom. 2014, 15, 1130. [Google Scholar] [CrossRef] [Green Version]
- Shi, R.; Jiao, W.; Yun, L.; Zhang, Z.; Zhang, X.; Wang, Q.; Li, Y.; Mi, F. Utilization of transcriptome, small RNA, and degradome sequencing to provide insights into drought stress and rewatering treatment in Medicago ruthenica. Front. Plant Sci. 2021, 12, 675903. [Google Scholar] [CrossRef]
- Wani, S.H.; Kumar, V.; Khare, T.; Tripathi, P.; Shah, T.; Ramakrishna, C.; Aglawe, S.; Mangrauthia, S.K. miRNA applications for engineering abiotic stress tolerance in plants. Biologia 2020, 75, 1063–1081. [Google Scholar] [CrossRef]
- Sarri, E.; Termentzi, A.; Abraham, E.M.; Papadopoulos, G.K.; Baira, E.; Machera, K.; Loukas, V.; Komaitis, F.; Tani, E. Salinity stress alters the secondary metabolic profile of M. sativa, M. arborea and their hybrid (Alborea). Int. J. Mol. Sci. 2021, 22, 4882. [Google Scholar] [CrossRef] [PubMed]
- Mäser, P.; Eckelman, B.; Vaidyanathan, R.; Horie, T.; Fairbairn, D.J.; Kubo, M.; Yamagami, M.; Yamaguchi, K.; Nishimura, M.; Uozumi, N.; et al. Altered shoot/root Na+ distribution and bifurcating salt sensitivity in Arabidopsis by genetic disruption of the Na+ transporter AtHKT1. FEBS Lett. 2002, 531, 157–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.M.; Liu, Z.H.; Wen, Z.Y.; Zhang, H.M.; Yang, F.; Guo, X.L. The vacuolar Na+-H+ antiport gene TaNHX2 confers salt tolerance on transgenic alfalfa (Medicago sativa). Funct. Plant Biol. 2012, 39, 708–716. [Google Scholar] [CrossRef]
- Gaxiola, R.A.; Li, J.; Undurraga, S.; Dang, L.M.; Allen, G.J.; Alper, S.L.; Fink, G.R. Drought- and salt-tolerant plants result from overexpression of the AVP1 H+-pump. Proc. Natl. Acad. Sci. USA 2001, 98, 11444–11449. [Google Scholar] [CrossRef] [Green Version]
- Sandhu, D.; Kaundal, A.; Acharya, B.R.; Forest, T.; Pudussery, M.V.; Liu, X.; Ferreira, J.F.S.; Suarez, D.L. Linking diverse salinity responses of 14 almond rootstocks with physiological, biochemical, and genetic determinants. Sci. Rep. 2020, 10, 21087. [Google Scholar] [CrossRef] [PubMed]
- Arif, A.; Zafar, Y.; Arif, M.; Blumwald, E. Improved growth, drought tolerance, and ultrastructural evidence of increased turgidity in tobacco plants overexpressing Arabidopsis vacuolar pyrophosphatase (AVP1). Mol. Biotechnol. 2013, 54, 379–392. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Knapp, J.; Koirala, P.; Rajagopal, D.; Peer, W.A.; Silbart, L.K.; Murphy, A.; Gaxiola, R.A. Enhanced phosphorus nutrition in monocots and dicots over-expressing a phosphorus-responsive type I H+-pyrophosphatase. Plant Biotechnol. J. 2007, 5, 735–745. [Google Scholar] [CrossRef] [PubMed]
- Bhaskaran, S.; Savithramma, D.L. Co-expression of Pennisetum glaucum vacuolar Na+/H+ antiporter and Arabidopsis H+-pyrophosphatase enhances salt tolerance in transgenic tomato. J. Exp. Bot. 2011, 62, 5561–5570. [Google Scholar] [CrossRef] [PubMed]
- Jones-Rhoades, M.W.; Bartel, D.P. Computational identification of plant microRNAs and their targets, including a stress-induced miRNA. Mol. Cell 2004, 14, 787–799. [Google Scholar] [CrossRef]
- Mallory, A.C.; Bouché, N. MicroRNA-directed regulation: To cleave or not to cleave. Trends Plant Sci. 2008, 13, 359–367. [Google Scholar] [CrossRef] [PubMed]
- Cao, C.; Long, R.; Zhang, T.; Kang, J.; Wang, Z.; Wang, P.; Sun, H.; Yu, J.; Yang, Q. Genome-wide identification of microRNAs in response to salt/alkali stress in Medicago truncatula through high-throughput sequencing. Int. J. Mol. Sci. 2018, 19, 4076. [Google Scholar] [CrossRef] [Green Version]
- Jodder, J.; Das, R.; Sarkar, D.; Bhattacharjee, P.; Kundu, P. Distinct transcriptional and processing regulations control miR167a level in tomato during stress. RNA Biol. 2018, 15, 130–143. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.; Wang, Y.; Li, J. Global identification and analysis of microRNAs involved in salt stress responses in two alfalfa (Medicago sativa ‘Millennium’) lines. Can. J. Plant Sci. 2019, 100, 445–455. [Google Scholar] [CrossRef]
- Fu, R.; Zhang, M.; Zhao, Y.; He, X.; Ding, C.; Wang, S.; Feng, Y.; Song, X.; Li, P.; Wang, B. Identification of salt tolerance-related microRNAs and their targets in maize (Zea mays L.) using high-throughput sequencing and degradome analysis. Front. Plant Sci. 2017, 8, 864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Islam, W.; Waheed, A.; Naveed, H.; Zeng, F. MicroRNAs mediated plant responses to salt stress. Cells 2022, 11, 2806. [Google Scholar] [CrossRef]
- Kang, T.; Yu, C.Y.; Liu, Y.; Song, W.M.; Bao, Y.; Guo, X.T.; Li, B.; Zhang, H.X. Subtly manipulated expression of ZmmiR156 in tobacco improves drought and salt tolerance without changing the architecture of transgenic plants. Front. Plant Sci. 2020, 10, 1664. [Google Scholar] [CrossRef] [PubMed]
- Khraiwesh, B.; Zhu, J.K.; Zhu, J. Role of miRNAs and siRNAs in biotic and abiotic stress responses of plants. Biochim. Biophys. Acta Gene Regul. Mech. 2012, 1819, 137–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, A.; Ruiz-Manriquez, L.M.; Serrano-Cano, F.I.; Reyes-Pérez, P.R.; Tovar Alfaro, C.K.; Barrón Andrade, Y.E.; Paul, S. Identification of microRNAs and their expression in leaf tissues of guava (Psidium guajava L.) under salinity stress. Agronomy 2020, 10, 1920. [Google Scholar] [CrossRef]
- Song, C.; Zhang, D.; Zheng, L.; Zhang, J.; Zhang, B.; Luo, W.; Li, Y.; Li, G.; Ma, J.; Han, M. miRNA and degradome sequencing reveal miRNA and their target genes that may mediate shoot growth in spur type mutant “Yanfu 6”. Front. Plant Sci. 2017, 8, 441. [Google Scholar] [CrossRef] [Green Version]
- Suh, C. Evaluation of Bioactivity of Phytotoxins from Pathogenic Fungi of Orobanche sp. Ph.D. Thesis, Agricultural University of Athens, Athina, Greece, 2011. [Google Scholar] [CrossRef]

{kind=link}
| miRNA | Target Name and Accession Number | Alignment | Target Description | Inhibition Type | ||
|---|---|---|---|---|---|---|
| mtr-miR167b-3p | AKT1 XM_003609192 | miRNA Target | 20 1354 |  | M. truncatula potassium channel protein LOC11443237 AKT1 mRNA | Cleavage |
| mtr-miR156a | AVP XM_003609415.3 | miRNA Target | 21 2478 |  | M. truncatula pyrophosphate-energized vacuolar membrane proton pump (LOC11438181) AVP mRNA | Cleavage |
| mtr-miR2605 | SOS3 XM_003602196.3 | miRNA Target | 20 631 |  | M. truncatula calcineurin B-like protein 7 (LOC11440434), transcript variant X4, SOS3 mRNA | Cleavage |
| mtr-miR2670f | SOS3 XM_003602196.3 | miRNA Target | 21 280 |  | M. truncatula calcineurin B-like protein 7 (LOC11440434), transcript variant X4, SOS3 mRNA | Cleavage |
| mtr-miR2670g | SOS3 XM_003602196.3 | miRNA Target | 21 280 |  | M. truncatula calcineurin B-like protein 7 (LOC11440434), transcript variant X4, SOS3 mRNA | Cleavage |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sarri, E.; Kapazoglou, A.; Gerakari, M.; Abraham, E.M.; Bebeli, P.J.; Tani, E. Expression Profile of Selected Genes Involved in Na+ Homeostasis and In Silico miRNA Identification in Medicago sativa and Medicago arborea under Salinity Stress. Stresses 2023, 3, 331-339. https://doi.org/10.3390/stresses3010024
Sarri E, Kapazoglou A, Gerakari M, Abraham EM, Bebeli PJ, Tani E. Expression Profile of Selected Genes Involved in Na+ Homeostasis and In Silico miRNA Identification in Medicago sativa and Medicago arborea under Salinity Stress. Stresses. 2023; 3(1):331-339. https://doi.org/10.3390/stresses3010024
Chicago/Turabian StyleSarri, Efi, Aliki Kapazoglou, Maria Gerakari, Eleni M. Abraham, Penelope J. Bebeli, and Eleni Tani. 2023. "Expression Profile of Selected Genes Involved in Na+ Homeostasis and In Silico miRNA Identification in Medicago sativa and Medicago arborea under Salinity Stress" Stresses 3, no. 1: 331-339. https://doi.org/10.3390/stresses3010024