
Use of Plant Regulators for Activation of Antioxidant Enzymes in Basil Plants under Water Deficit Conditions
 , , and
, , and
Abstract
:1. Introduction
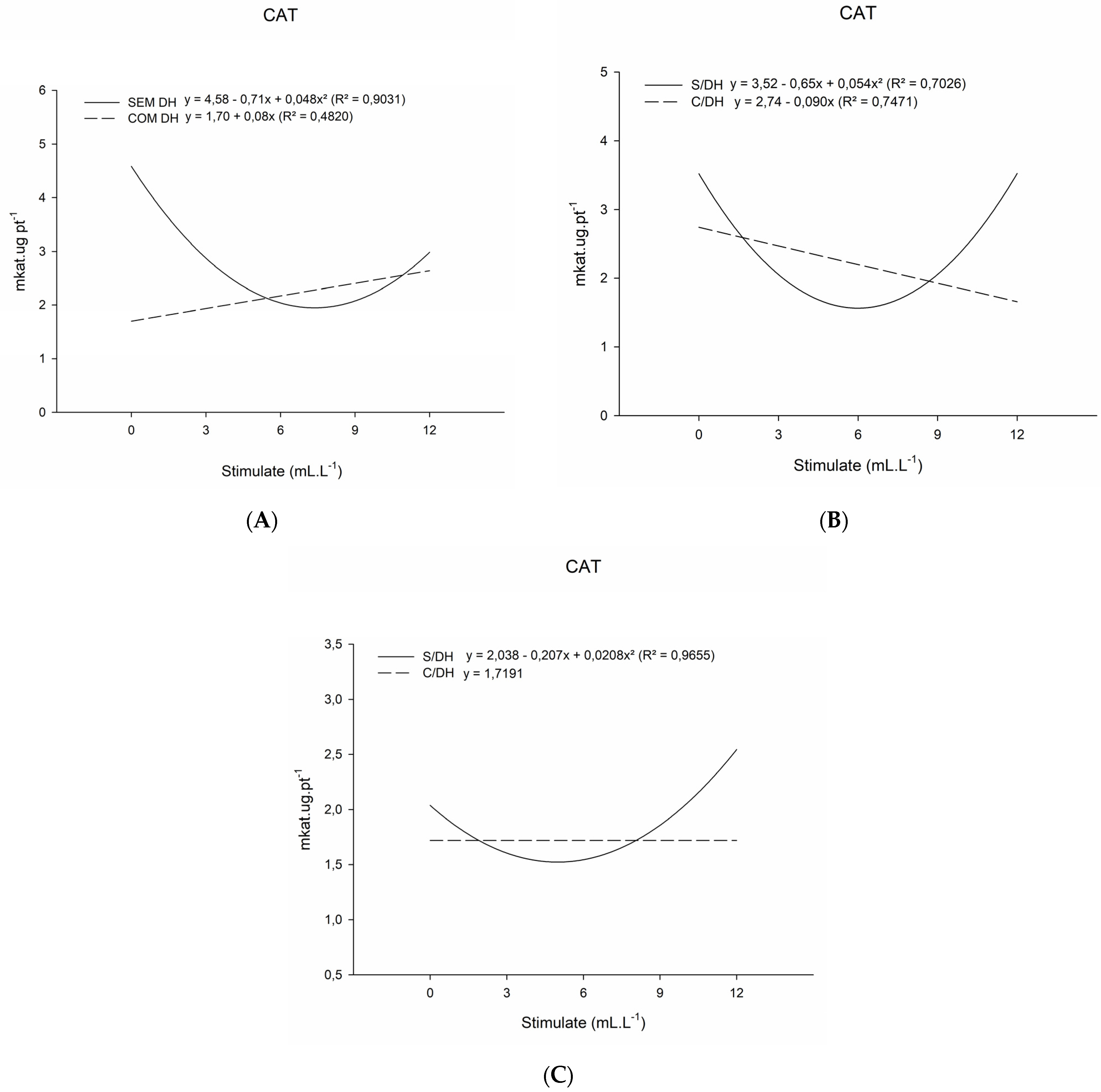
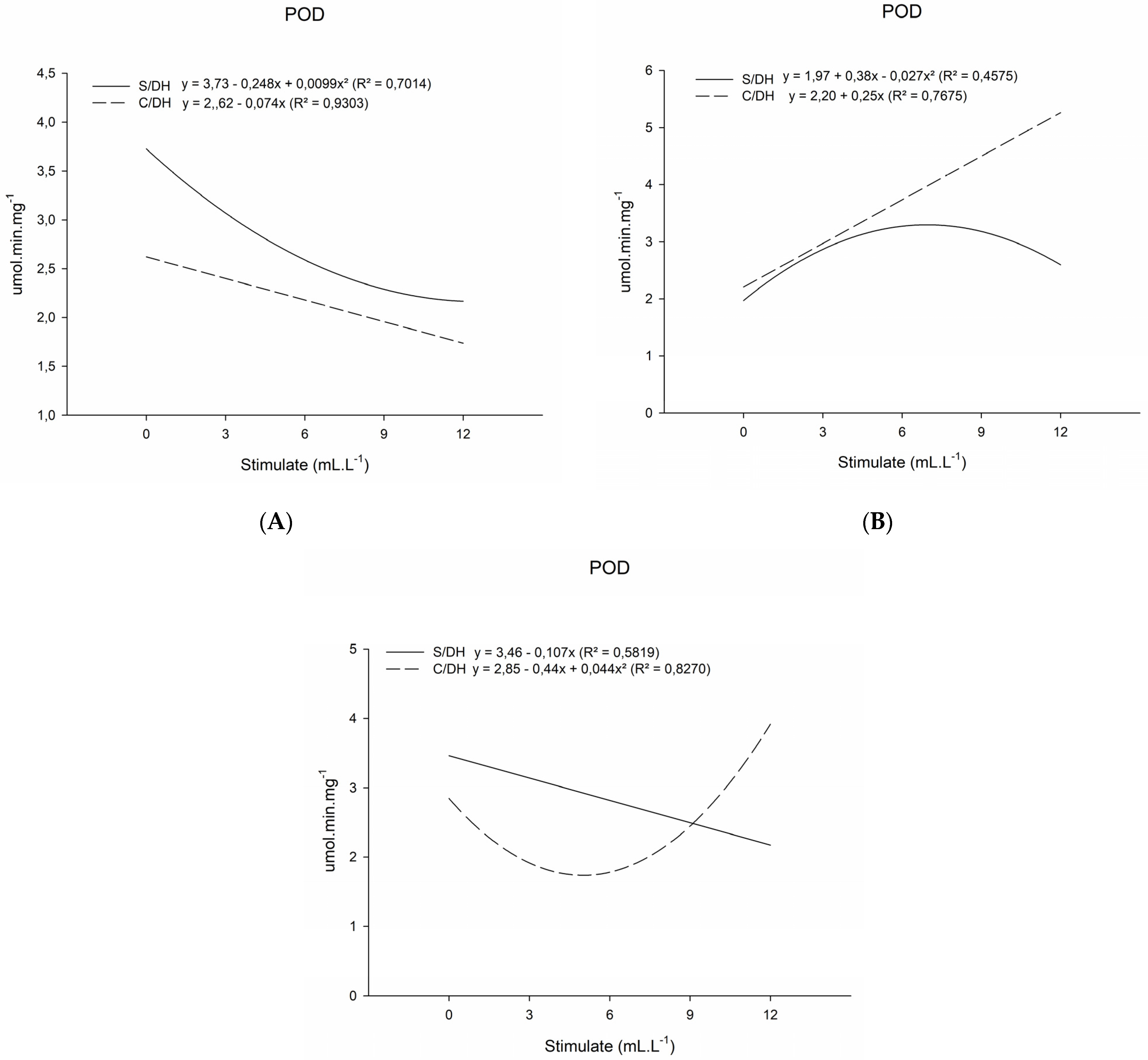
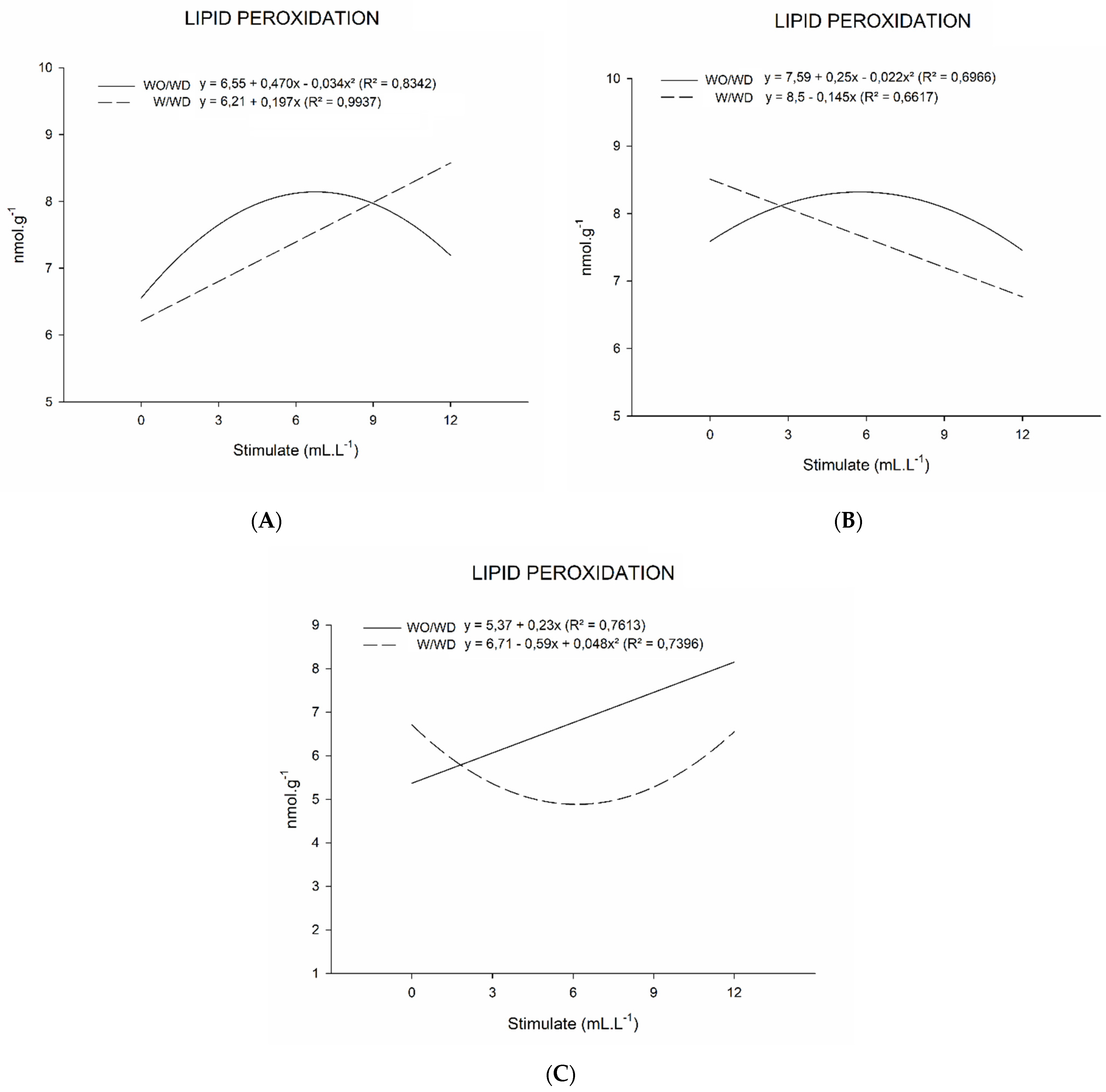
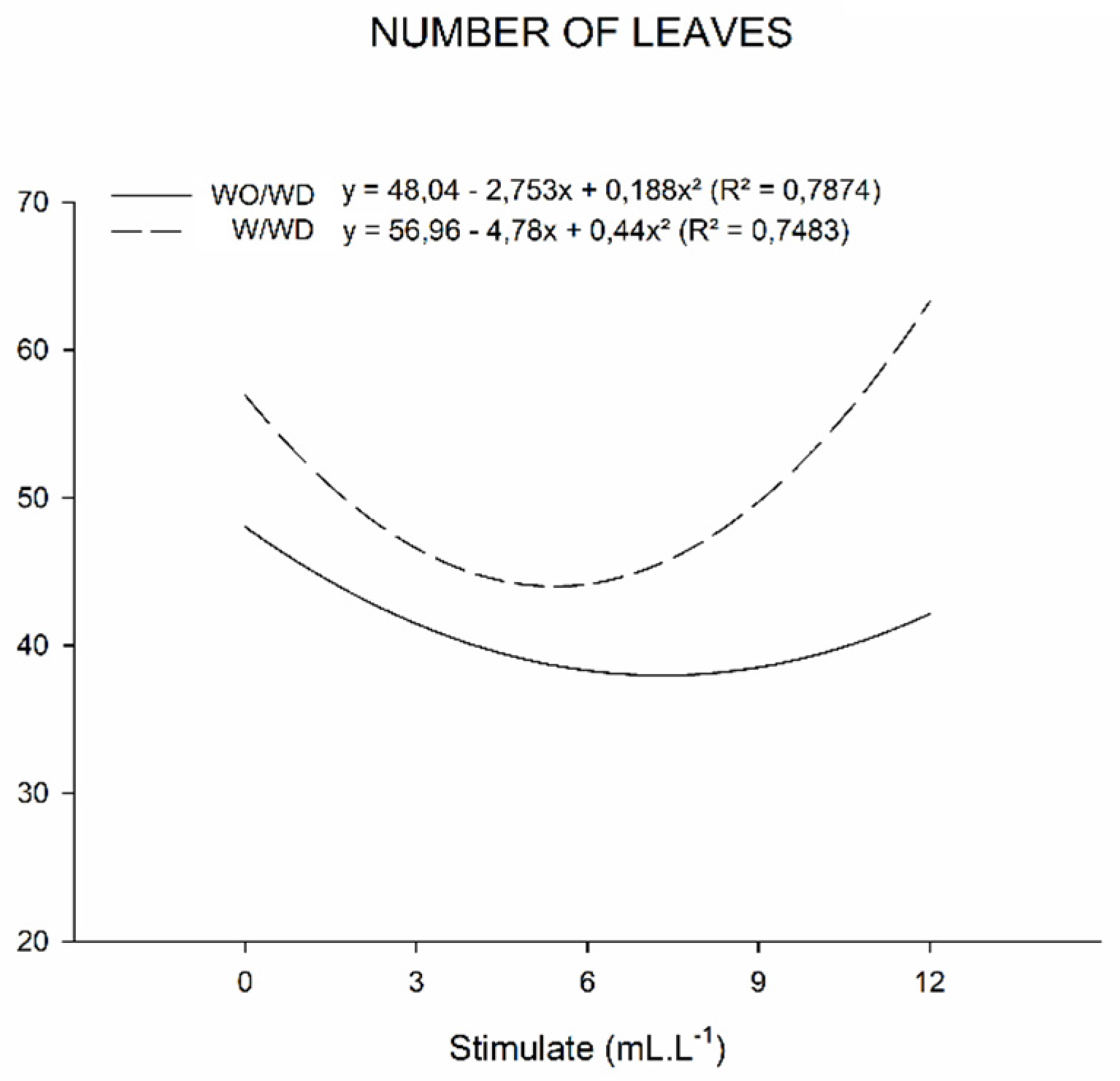
2. Results
2.1. Analysis of Antioxidative Enzyme Activity
2.2. Lipid Peroxidation
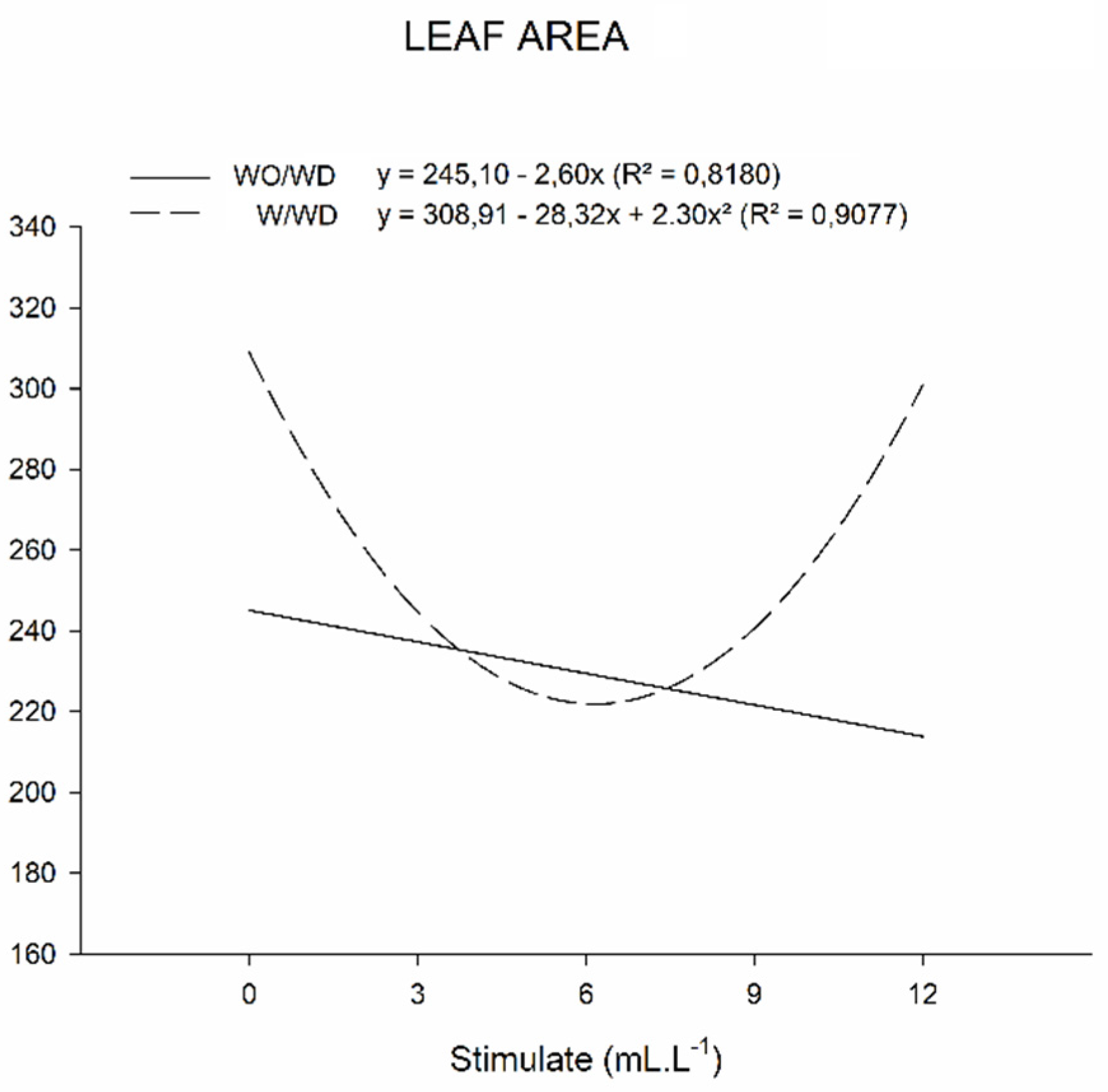
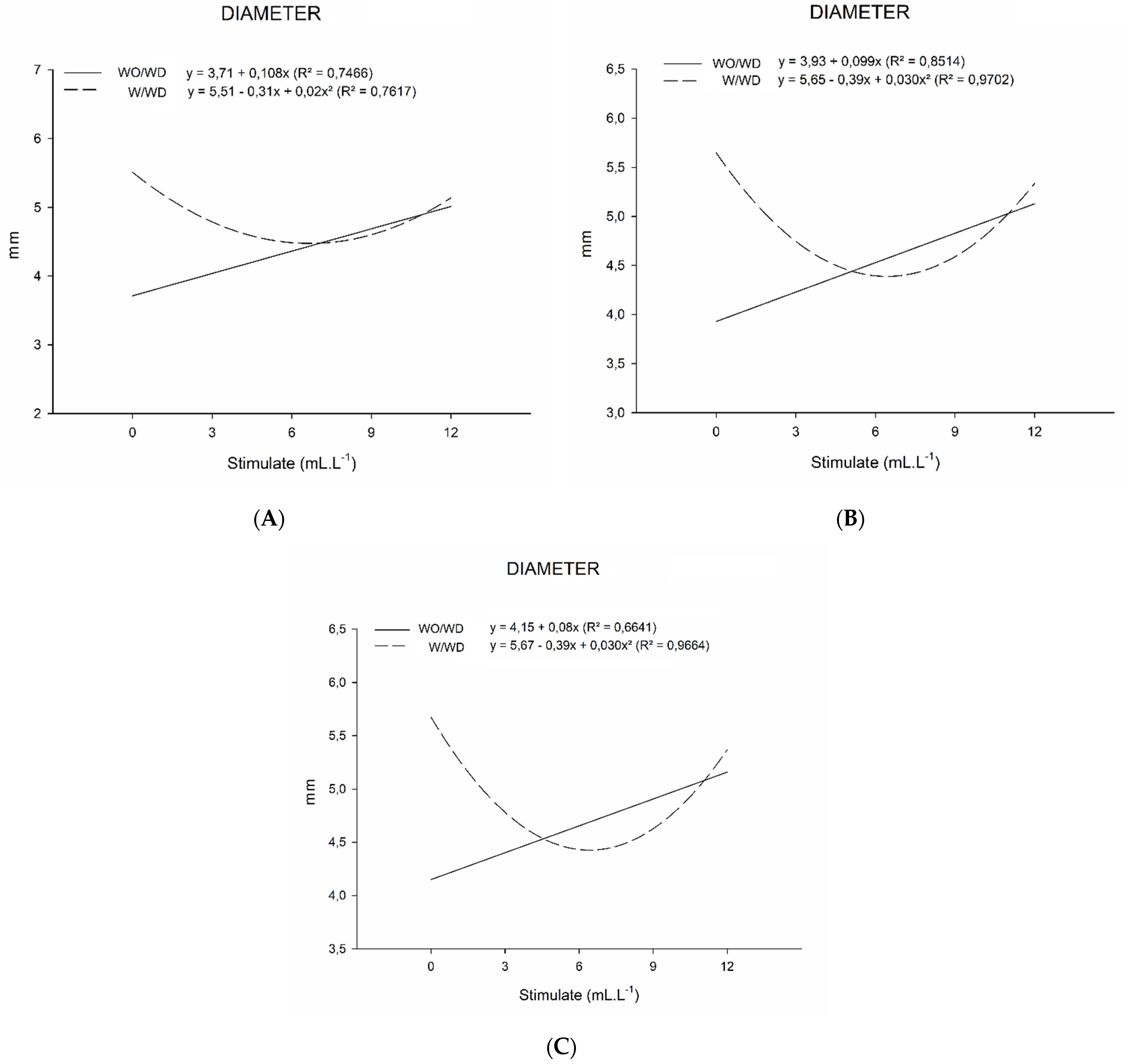
2.3. Growth Measurements
3. Discussion
3.1. Analysis of Antioxidative Enzyme Activity
3.2. Lipid Peroxidation
3.3. Growth Measurements
4. Materials and Methods
4.1. Experiment Setup and Conduction
4.2. Analysis of Antioxidative Enzyme Activity
4.3. Total Soluble Proteins
4.4. Lipid Peroxidation
4.5. Growth Measurements
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lorenzi, H.; Matos, F.J.A. Plantas Medicinais no Brasil: Nativas e Exóticas, 2nd ed.; Instituto Plantarum: Nova Odessa, Brazil, 2008. (In Portuguese) [Google Scholar]
- Barbosa, D.D. Desenvolvimento Vegetativo e Parâmetros Fisiológicos em Genótipos de Amendoim sob Déficit Hídrico e Inoculados com Rizóbios 76 f. Master’s Thesis, Universidade Estadual da Paraíba/Embrapa Algodão, Campina Grande, Brazil, 2016. [Google Scholar]
- Blank, A.F.; Filho, J.L.D.C.; Neto, A.L.D.S.; Alves, P.B.; Arrigoni-Blank, M.D.F.; Silva-Mann, R.; Mendonça, M.D.C. Morphological and agronomic characterization of basil and basil accessions. Hortic. Bras. 2004, 22, 113–116. [Google Scholar] [CrossRef]
- Simões, C.M.O.; Schenkel, E.P.; Mello, J.C.P.; Mentz, L.A.; Petrovick, P.R. Óleos Voláteis. In Farmacognosia: Da planta ao Medicamento 5; Editora da UFSC: Florianópolis, Brazil, 2004; p. 475. (In Portuguese) [Google Scholar]
- De Morais, L.A.S. Influência dos fatores abióticos na composição química dos óleos essenciais. Embrapa Meio Ambiente-Artig. Em An. De Congr. (ALICE) 2009, 27, S4050–S4063. [Google Scholar]
- Joly, R.J.; Zaerr, J.B.; Dalton, D.A.; Hanus, F.J.; Russell, S.A.; Evans, H.J. Alteration of Cell-Wall Water Content and Elasticity in Douglas-Fir during Periods of Water Deficit. Plant Physiol. 1987, 83, 418–422. [Google Scholar] [CrossRef]
- Kozlowski, T.T.; Pallardy, S. Acclimation and adaptive responses of woody plants to environmetal stresses. Bot. Rev. 2002, 68, 279–334. [Google Scholar] [CrossRef]
- Taiz, L.; Zeiger Ian Max Møller, E.; Murphy, A. Fisiologia e Desenvolvimento Vegetal—6a Edição; Artmed Editora: São Paulo, Brasil, 2017. [Google Scholar]
- Cavalcante, W.S.D.S.; Da Silva, N.F.; Teixeira, M.B.; Filho, F.R.C.; Nascimento, P.E.R.; Corrêa, F.R. Eficiência dos Bioestimulantes no Manejo do Déficit Hídrico na Cultura da Soja. Irriga 2020, 25, 754–763. [Google Scholar] [CrossRef]
- Torsian, W.S.; Kikuti, A.L.P.; Kikuti, H.; Pereira, C.E. Bioestimulantes no desenvolvimento da cana-de-açúcar. Magistra 2011, 31, 625–634. [Google Scholar]
- De Oliveira, F.A.; De Medeiros, J.F.; Da Cunha, R.C.; De Souza, M.W.L.; Lima, L.A. Use of biostimulants in relieving salt stress in popcorn. Rev. Ciência Agronômica 2016, 47, 307–315. [Google Scholar] [CrossRef]
- Reis, M.A.M. Bioestimulante Como Estratégia ao Déficit Hídrico em Feijoeiro Comum. 69 f. Doctoral Dissertation, Universidade Estadual Paulista Júlio de Mesquita Filho, Jaboticabal, São Paulo, Brazil, 2021. [Google Scholar]
- Freschi, J.; Magalhães Filho, J.R. O poder dos Hormônios Vegetais em Modular o Desenvolvimento da Planta. Available online: https://www.stoller.com.br/o-poder-dos-hormonios-vegetais-em-modular-o-desenvolvimento-da-planta/ (accessed on 9 December 2022).
- Lameira, O.A.; Pinto, J.E.B.P.; Arrigoni, M.F.B. Enraizamento de miniestacas de erva-baleeira. Hortic. Brasileira. 1997, 5, 114–116. [Google Scholar]
- El-Keltawi, N.E.; Croteau, R. Influence of foliar applied cytokinins on growth and essential oil content of several members of the lamiaceae. Phytochemistry 1987, 26, 891–895. [Google Scholar] [CrossRef]
- Paes, L.; Mendonça, M.S.; Casas, L. Aspectos Estruturais e Fitoquímicos de partes vegetativas de Costus spicatus (Jacq.) Sw. (Costaceae). Rev. Bras. de Plantas Med. 2012, 15, 380–390. [Google Scholar] [CrossRef]
- Spinelli, F.A.W.; Rademacher, B.E.; Sabatini, A.C.G.; Costa, A. Reduction of scabincidence (Venturia inaequalis) in apple with prohexadione-Ca and trinexapac-ethyl, two growth regulating acylcyclohexanediones. Crop Prot. 2010, 29, 691–698. [Google Scholar] [CrossRef]
- Souza, E.R. Fenologia e Mistura de Reguladores Vegetais e de Fertilizante Foliar no Metabolismo da Videira cv. Sweet Sunshine em Clima Semiárido 143 f. Doctor’s Dissertation, Universidade Estadual Paulista, São Paulo, Brazil, 2014. [Google Scholar]
- Kang, H.-M.; Saltveit, M.E. Reduced chilling tolerance in elongating cucumber seedling radicles is related to their reduced antioxidant enzyme and DPPH-radical scavenging activity. Physiol. Plant. 2002, 115, 244–250. [Google Scholar] [CrossRef] [PubMed]
- Souza, E.L.; Woyann, L.G.; Ahlert, R.J.; Almeida, A.M.; Costa, O. A Peroxidação lipídica em cultivares de arroz irrigado Submetidas ao estresse por ferro em hidroponia. In Proceedings of the Anais. CIC XX Congresso de iniciação científica. UFPEL, Pelotas, Brazil, 8–11 November 2011. [Google Scholar]
- Hu, Z.-H.; Shen, Y.-B.; Shen, F.-Y.; Su, X.-H. Effects of feeding Clostera anachoreta on hydrogen peroxide accumulation and activities of peroxidase, catalase, and ascorbate peroxidase in Populus simonii × P. pyramidalis ‘Opera 8277’ leaves. Acta Physiol. Plant. 2009, 31, 995–1002. [Google Scholar] [CrossRef]
- Nascimento, J.; Barrigossi, J.A. O Papel das Enzimas Antioxidantes na Defesa das Plantas Contra Insetos Herbívoros e Fitopatógenos. Agrar. Acad. 2014, 1, 234–250. [Google Scholar] [CrossRef]
- Rossi, V.S.; Costa, M.F. Mecanismo antioxidante em plantas. In Proceedings of the 10º mostra acadêmica UNIMEP, Piracicaba, Brazil, 10–12 November 2012; pp. 1–4. Available online: http://www.unimep.br/phpg/mostraacademica/anais/10mostra/4/31.pdf. (accessed on 5 July 2021).
- Barbosa, K.B.F.; Costa, N.M.B.; Alfenas, R.C.G.; Paula, S.O.; Minim, V.P.R.; Bressan, J. Estresse oxidativo: Conceito, implicações e fatores modulatórios. Rev. Nut. 2010, 23, 629–643. [Google Scholar] [CrossRef]
- Sofo, A.; Scopa, A.; Nuzzaci, M.; Vitti, A. Ascorbate Peroxidase and Catalase Activities and Their Genetic Regulation in Plants Subjected to Drought and Salinity Stresses. Int. J. Mol. Sci. 2015, 16, 13561–13578. [Google Scholar] [CrossRef]
- Prado, D.Z. Enraizamento de Eucalyptus Grandis x Eucalyptus Urophylla Pela Ação de Peróxido de Hidrogênio, Quercetina e Ácido Indolbutírico 106 f. Master’s Thesis, Curso de Ciências Biológicas (Botânica), Fisiologia e e Bioquímica Vegetal, Universidade Estadual Paulista, Júlio de Mesquita Filho, Botucatu, Brazil, 2014. [Google Scholar]
- Lopes, M.J.C.; Souza, I.R.P.; Magalhães, P.C.; Gama, E.E.G. Oxidação proteíca e peroxidação lípidica em plantas de diferentes ciclos de seleção do milho ‘SARACURA’, sob encharcamento contínuo. Rev. Bras. De Milho E Sorgo Lavras 2005, 4, 362–373. [Google Scholar] [CrossRef]
- Torres, T.P. Putrescina no Desenvolvimento do Tomateiro cv. Justyne em Condições de Estresse Hídrico 56 f. Master’s Thesis, Curso de Agronomia, Horticultura, Universidade Estadual Paulista Júlio de Mesquita Filho, Botucatu, Brazil, 2020. [Google Scholar]
- De Vasconcelos, A.C.F.; Zhang, X.; Ervin, E.H.; Kiehl, J.D.C. Enzymatic antioxidant responses to biostimulants in maize and soybean subjected to drought. Sci. Agricola 2009, 66, 395–402. [Google Scholar] [CrossRef]
- Deus, K.E. Atividade Enzimática e Expressão Diferencial da Superóxido Dismutase (SOD) em Plantas de Arroz de Terras Altas sob Deficiência Hídrica 131 f. Master’s Thesis, Curso de Biologia, Universidade Federal de Goiás, Goiânia, Brazil, 2014. [Google Scholar]
- Nunes, R.C.A.; Viana, R.S.; Machado, N.B., Neto. Atividade enzimática da superóxido dismutase em resposta aos fitorreguladores em Gerbera jamensonii. Comunicata Scientiae 2015, 6, 83–89. [Google Scholar]
- Moreira, B.R.A.; Viana, R.S.; Manarelli, F.; Viana, C.R.A.; Nakamune, A.C.M.S. Parâmetros Tecnológicos e Avaliação de Enzimas Antioxidantes da Cana-de-açúcar (Saccharum spp.) quando Aplicados Maturadores Químicos. Revista Virtual de Química 2018, 10, 1–23. [Google Scholar]
- Sousa, M.C. Reguladores Vegetais e Nutrientes Minerais no Metabolismo de Plantas de Tomateiro 76 f. Master’s Thesis, Curso de Agronomia, Horticultura, Universidade Estadual Paulista Júlio de Mesquita Filho, Botucatu, Brazil, 2016. [Google Scholar]
- Rossatto, T. Alterações Fisiológicas, Bioquímicas e Moleculares de Arroz, cv BRS AG, em Resposta ao Estresse Salino 117 f. Master’s Thesis, Curso de Biologia, Fisiologia Vegetal, Universidade Federal de Pelotas, Pelotas, Brazil, 2016. [Google Scholar]
- Bispo, G.L. Fenologia e desempenho ecofisiológico de Vasconcellea quercifolia A.St.-Hill 88 f. Doctoral Dissertation, Curso de Agronomia, Universidade Estadual Paulista (Unesp), Botucatu, Brazil, 2020. [Google Scholar]
- Fernandes, C.F.; Vieira, J.R., Jr.; Silva, D.S.G.; Alves, R.C. Estresse Oxidativo e o Mecanismo de Defesa de Plantas Contra Patógenos; Embrapa Rondônia: Porto Velho, Rondônia, Brazil, 2013. [Google Scholar]
- Sharma, S.; Ghoshal, C.; Arora, A.; Samar, W.; Nain, L.; Paul, D. Strain Improvement of Native Saccharomyces cerevisiae LN ITCC 8246 Strain Through Protoplast Fusion to Enhance Its Xylose Uptake. Appl. Biochem. Biotechnol. 2021, 193, 2455–2469. [Google Scholar] [CrossRef] [PubMed]
- Garcia, R.P.O. Efeitos de Giberelina Para Respostas em Plantas de Tomate à Deficiência Hídrica 71 f. Master’s Thesis, Curso de Agronomia, Fisiologia Vegetal, Universidade Federal de Viçosa, Viçosa, Brazil, 2014. [Google Scholar]
- Soares, A.M.S. Análise da Resposta Antioxidativa em Plantas de Ricinus Communis Submetidas ao Estresse por Metiljasmonato 93 f. Master’s Thesis, Curso de Biologia, Universidade Estadual do Norte Fluminense Darcy Ribeiro, Campos do Goytacazes, Brazil, 2006. [Google Scholar]
- Piza, I.M.T.; Lima, G.P.P.; Brasil, O.G. Peroxidase Activity and Proteins Levels on Pineapple Plants Micropropagated on Salinity Medium. Revista Brasileiro Agrociência 2003, 9, 361–366. [Google Scholar]
- Uprety, B.; Kaja, A.; Ferdoush, J.; Sen, R.; Bhaumik, S.R. Regulation of Antisense Transcription by NuA4 Histone Acetyltransferase and Other Chromatin Regulatory Factors. Mol. Cell. Biol. 2016, 36, 992–1006. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, J.M.; de Melo, N.F.; de Araujo, F.P.; Fernandes, K.V.S.; Pinto, M.S.T. Efeito do estresse hídrico sobre a atividade de enzimas antioxidantes em xilopódio de umbuzeiro (Spondias tuberosa. Arruda). In Proceedings of the XXII Congresso Brasileiro de Fruticultura, Bento Gonçalves, Brazil, 22–26 October 2012; pp. 573–576. Available online: https://www.alice.cnptia.embrapa.br/alice/bit-stream/doc/938810/1/00000003904SWB.pdf. (accessed on 5 June 2021).
- Camargo, R.B. Citocinina, piraclostrobina e putrescina: Influência no desenvolvimento de mudas em três cultivares de videiras em diferentes ambientes 80 f. Doctoral Dissertation, Curso de Agronomia, Horticultura, Universidade Estadual Paulista Júlio de Mesquita Filho, Botucatu, Brazil, 2016. [Google Scholar]
- Yamamoto, Y.; Kobayashi, Y.; Matsumoto, H. Lipid Peroxidation Is an Early Symptom Triggered by Aluminum, But Not the Primary Cause of Elongation Inhibition in Pea Roots. Plant Physiol. 2001, 125, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Junqua, M.; Biolley, J.-P.; Pie, S.; Kanoun-Boulé, M.; Duran, R.; Goulas, P. In vivo occurrence of carbonyl residues in Phaseolus vulgaris proteins as a direct consequence of a chronic ozone stress. Plant Physiol. Biochem. 2000, 38, 853–861. [Google Scholar] [CrossRef]
- Souza, J.B. Caracterização Fisiológica e Bioquímica de Linhagens de Milho Visando Tolerância ao Alumínio 70 f. Master’s Thesis, Universidade Federal de Minas Gerais, Belo Horizonte, Brazil, 2003. [Google Scholar]
- Giannopolotis, C.N.; Ries, S.K. Superoxide dismutases: I. Occurrence in higher plants. Plant Physiol. 1977, 59, 309–314. [Google Scholar] [CrossRef]
- Soares, C.S.M.; Ramos, A.P.; Cruz, N.F. Junior; Steffens, C.A.; Amarante, C.V.T. Ação de brassinosteroide e auxina sobre o estresse oxidativo em maçãs ’gala’. Available online: https://www.udesc.br/arquivos/udesc/id_cpmenu/3168/31_A__o_de_brassinosteroide_e_auxina_sobre_o_estresse_oxidativo_em_ma__s_Gala_1503494829714_3168.pdf (accessed on 11 January 2023).
- Santos, J.F. dos. Cultivo Hidropônico de Manjericão sob Estresse Salino: Crescimento, Produção e Aspectos Bioquímicos 44 f. Doctoral Dissertation, Curso de Engenharia Agrícola, Universidade Federal do Recôncavo da Bahia, Cruz das Almas, Brazil, 2017. [Google Scholar]
- Rossi, R. Nitrogênio em Cobertura e Bioestimulante Aplicado via Foliar em Feijoeiro de Inverno no Sistema Plantio Direto 62 f. Master’s Thesis, Curso de Agronomia, Universidade Estadual Paulista Júlio de Mesquita Filho, Ilha Solteira, Brazil, 2011. [Google Scholar]
- Gonçalves, B.H.L.; Souza, J.M.A.; Ferraz, R.A.; Tecchio, M.A.; Leonel, S. Efeito do bioestimulante Stimulate® no desenvolvimento de mudas de maracujazeiro cv. BRS Rubi do Cerrado. Revista de Ciências Agrárias 2017, 41, 147–155. [Google Scholar] [CrossRef]
- Souza, J.M.A.; Gonçalves, B.H.L.; Santos, A.M.F.; Ferraz, R.A.; Leonel, S. Efeito de bioestimulante no desenvolvimento inicial de plântulas do porta-enxerto cítrico tangerineira ‘Cleópatra’. Sci. Plena. 2013, 9, 1–8. [Google Scholar]
- Neiva, L.S. Filho; Souza, A.S. Stimulate® na Micropropagação da Mandioca (Manihot esculenta Crantz). In 6º Jornada Científica; Embrapa Mandioca e Fruticultura: Bahia, Brazil, 2012. [Google Scholar]
- Torres, R.C.; Borges, K.C.A.S. Effects of the action of gibberellin in the initial growth of pepper plants (Capsicum frutescens). Cadernos UniFOA (Centro Universitário De Volta Redonda) 2013, 8, 11–16. [Google Scholar]
- Tecchio, M.A.; Leonel, S.; Dos Reis, L.L.; Simonetti, L.M.; Da Silva, M.J.R. Stimulate no Desenvolvimento de Mudas de Kunquat ‘Nagami’. Irriga 2015, 1, 97–106. [Google Scholar] [CrossRef]
- De Oliveira, A.; Ferreira, G.; Rodrigues, J.D.; Ferrari, T.B.; Kunz, V.L.; Primo, M.A.; Poletti, L.D. Efeito de reguladores vegetais no desenvolvimento de mudas de Passiflora alata Curtis. Rev. Bras. de Frutic. 2005, 27, 9–13. [Google Scholar] [CrossRef]
- Dos Santos, L.P.; Barbacena, D.R.; Gonçalves, R.C.; Nascimento, C.A.C.; Carvalho, F.L.D.C.; França, L.C.; Adorian, G.C. Aplicação de Bioestimulante e Complexo de Nutrientes no Tratamento de Sementes de Soja. Agri-Environ. Sci. 2020, 6, 1–8. [Google Scholar] [CrossRef]
- Scalon, S.D.P.Q.; De Lima, A.A.; Filho, H.S.; Vieira, M.D.C. Germinação de sementes e crescimento inicial de mudas de Campomanesia adamantium Camb.: Efeito da lavagem, temperatura e de bioestimulantes. Rev. Bras. Sement. 2009, 31, 96–103. [Google Scholar] [CrossRef]
- Cerezer, B.; Cin, G.D. Ação de Giberelina e Auxina no Desenvolvimento da Planta e na Qualidade de Tomates ‘Gaúcho’ 42 f. Graduation Thesis, Curso de Agronomia, Instituto Federal de Santa Catarina, São Miguel do Oeste, Brazil, 2020. [Google Scholar]
- Martínez, L.D.O.; Mendoza, O.J.; Valenzuela, M.C.; Serrano, P.A.; Olarte, S.J. Efecto de las giberelinas sobre el crecimiento y calidad de plántulas de tomate. Biotecnia 2013, 15, 56–60. [Google Scholar] [CrossRef]
- Davies, P.J. The plant hormones: Biosynthesis, signal transduction, action, 3rd ed.; Springer: New York, NY, USA, 2010; p. 805. [Google Scholar]
- Leite, V.M.; Rosolem, C.A.; Rodrigues, J.D. Gibberellin and cytokinin effects on soybean growth. Sci. Agric. 2003, 60, 537–541. [Google Scholar] [CrossRef]
- Almeida, G.M.; Rodrigues, J.G.L. Desenvolvimento de plantas através da interferência de auxinas, citocininas, etileno e giberelinas. Braz. J. Appl. Technol. Agric. Sci. 2016, 1, 111–117. [Google Scholar] [CrossRef]
- Peixoto, P.H.P.; Cambraia, J.; Sant’Anna, R.; Mosquim, P.R.; Moreira, M.A. Aluminium effects on lipid peroxidation and the activities of enzymes of oxidative metabolism in sorghum. Revista Brasileira de Fisiologia Vegetal 1999, 11, 137–143. [Google Scholar]
- Teisseire, H.; Guy, V. Copper-induced changes in antioxidant enzymes activities in fronds of duckweed (Lemna minor). Plant Sci. 2000, 153, 65–72. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Abade, M.; Ávila, M.T.; Lima, L.G.S.; Silva, E.G.; Oliveira, C.S.; Silva, R.T.L. Determinação dos Teores de Proteínas Solúveis Totais em Plantas de Girassol (Helianthus annuus L.). In Proceedings of the VII Workshop de Agroenergia Matérias Primas Ribeirão Preto, Anais [...], Infobibos, Ribeirão Preto, Brazil, 29–30 March 2014; pp. 1–4. Available online: http://www.infobibos.com/Agroenergia/CD_2014/Resumos/ResumoAgroenergia_201 4_010.pdf (accessed on 5 September 2019).
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts: I. kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| F | SOD 20 DAT | SOD 35 DAT | SOD 50 DAT |
|---|---|---|---|
| Water Deficit (DH) | 46.96 ** | 50.85 ** | 119.56 ** |
| Plant Regulators (PR) | 9.95 ** | 46.10 ** | 9.74 ** |
| WD × PR | 40.12 ** | 54.46 ** | 32.18 ** |
| CV (%) | 4.09 | 5.25 | 5.5 |
| F | CAT 20 DAT | CAT 35 DAT | CAT 50 DAT |
|---|---|---|---|
| Water Deficit (WD) | 16.45 ** | 15.79 ** | 2.19 ns |
| Plant Regulators (PR) | 2.00 * | 3.91 * | 4.80 ** |
| WD × PR | 3.57 ** | 1.52 ns | 3.96 * |
| CV (%) | 25.88 | 27.81 | 26.17 |
| F | POD 20 DAT | POD 35 DAT | POD 50 DAT |
|---|---|---|---|
| Water Deficit (DH) | 47.55 ** | 44.86 ** | 3.75 ns |
| Plant Regulators (PR) | 33.02 ** | 25.07 ** | 4.14 ** |
| WD × PR | 6.04 ** | 23.98 ** | 12.15 ** |
| CV (%) | 10.93 | 13.88 | 20.97 |
| F | MDA 20 DAT | MDA 35 DAT | MDA 50 DAT |
|---|---|---|---|
| Water Deficit (DH) | 0.23 ns | 2.64 ns | 47.21 ** |
| Plant Regulators (PR) | 4.37 ** | 11.49 ** | 36.11 ** |
| WD × PR | 3.06 * | 2.77 * | 46.48 ** |
| CV (%) | 12.52 | 7.07 | 7.36 |
| F | Number of Leaves | Leaf Area |
|---|---|---|
| Water Deficit (DH) | 88.18 ** | 21.52 ** |
| Plant Regulators (PR) | 22.71 ** | 6.86 ** |
| WD × PR | 34.77 ** | 10.87 ** |
| CV (%) | 7.50 | 9.37 |
| F | Diameter 20 DAT | Diameter 35 DAT | DIAMETER 50 DAT |
|---|---|---|---|
| Water Deficit (DH) | 8.43 ** | 7.24 * | 7.23 * |
| Plant Regulators (PR) | 0.91 ns | 3.21 * | 6.35 ** |
| WD × PR | 5.76 ** | 6.12 ** | 8.27 ** |
| CV (%) | 12.76 | 10.27 | 7.87 |
| pH | MO | P | S | Al3+ | H + Al3+ | K | Ca | Mg | SB | CTC | V% |
|---|---|---|---|---|---|---|---|---|---|---|---|
| CaC2 | g/dm³ | mg/dm³ | mmolc/dm³ | ||||||||
| 4.0 | 11 | 1 | 18 | 10 | 63 | 1.25 | 5 | 1 | 7 | 70 | 10 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carvalho, B.L.; Aires, E.S.; Rodrigues, J.D.; Ono, E.O. Use of Plant Regulators for Activation of Antioxidant Enzymes in Basil Plants under Water Deficit Conditions. Stresses 2023, 3, 282-301. https://doi.org/10.3390/stresses3010021
Carvalho BL, Aires ES, Rodrigues JD, Ono EO. Use of Plant Regulators for Activation of Antioxidant Enzymes in Basil Plants under Water Deficit Conditions. Stresses. 2023; 3(1):282-301. https://doi.org/10.3390/stresses3010021
Chicago/Turabian StyleCarvalho, Beatriz Lívero, Eduardo Santana Aires, João Domingos Rodrigues, and Elizabeth Orika Ono. 2023. "Use of Plant Regulators for Activation of Antioxidant Enzymes in Basil Plants under Water Deficit Conditions" Stresses 3, no. 1: 282-301. https://doi.org/10.3390/stresses3010021