


Field Investigation into Tree Fates from Recent Apple Tree Decline: Abrupt Hydraulic Failure versus Gradual Hydraulic Loss


Abstract
:1. Introduction
2. Results
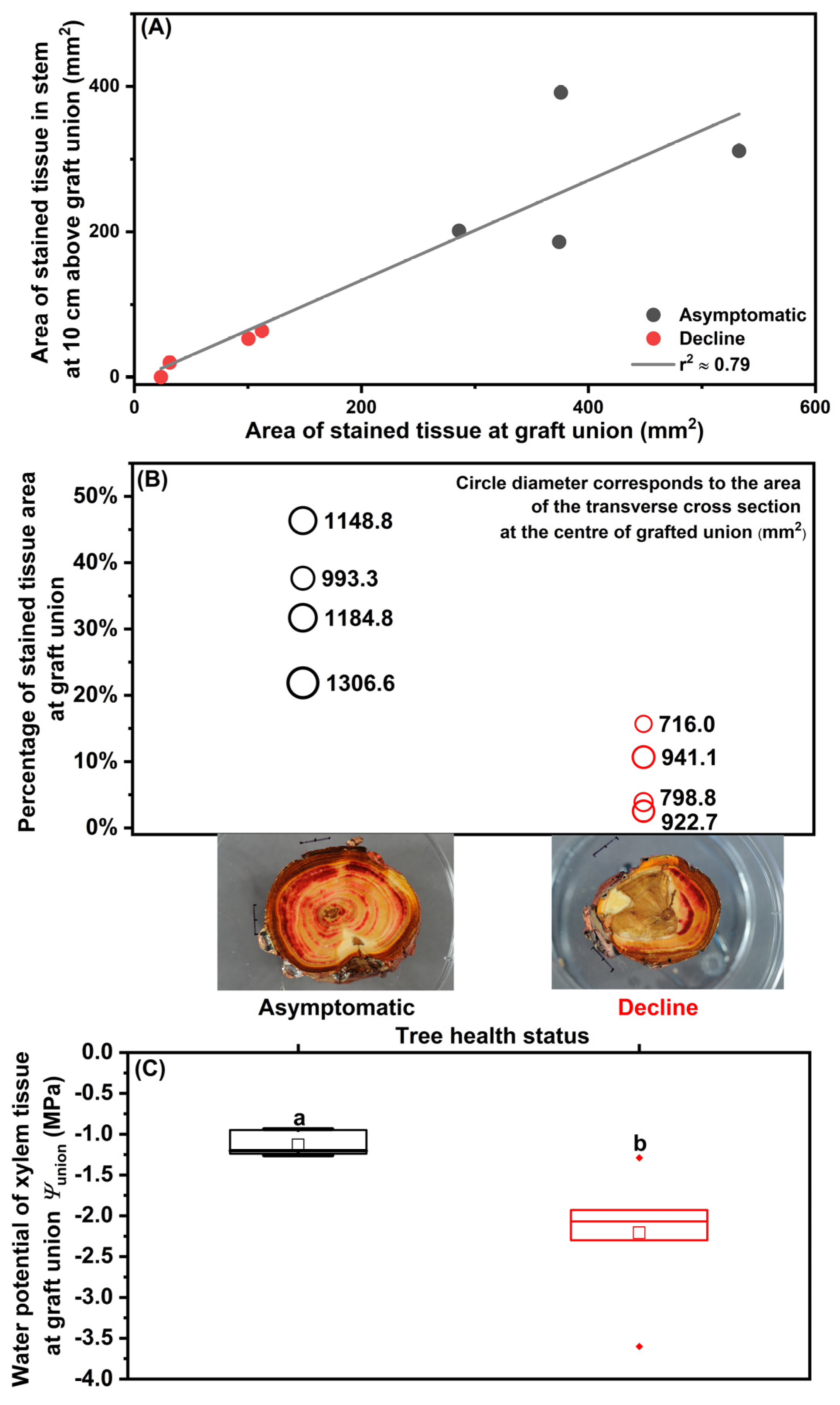
2.1. Hydraulic Characteristics Prior to Tree Mortality
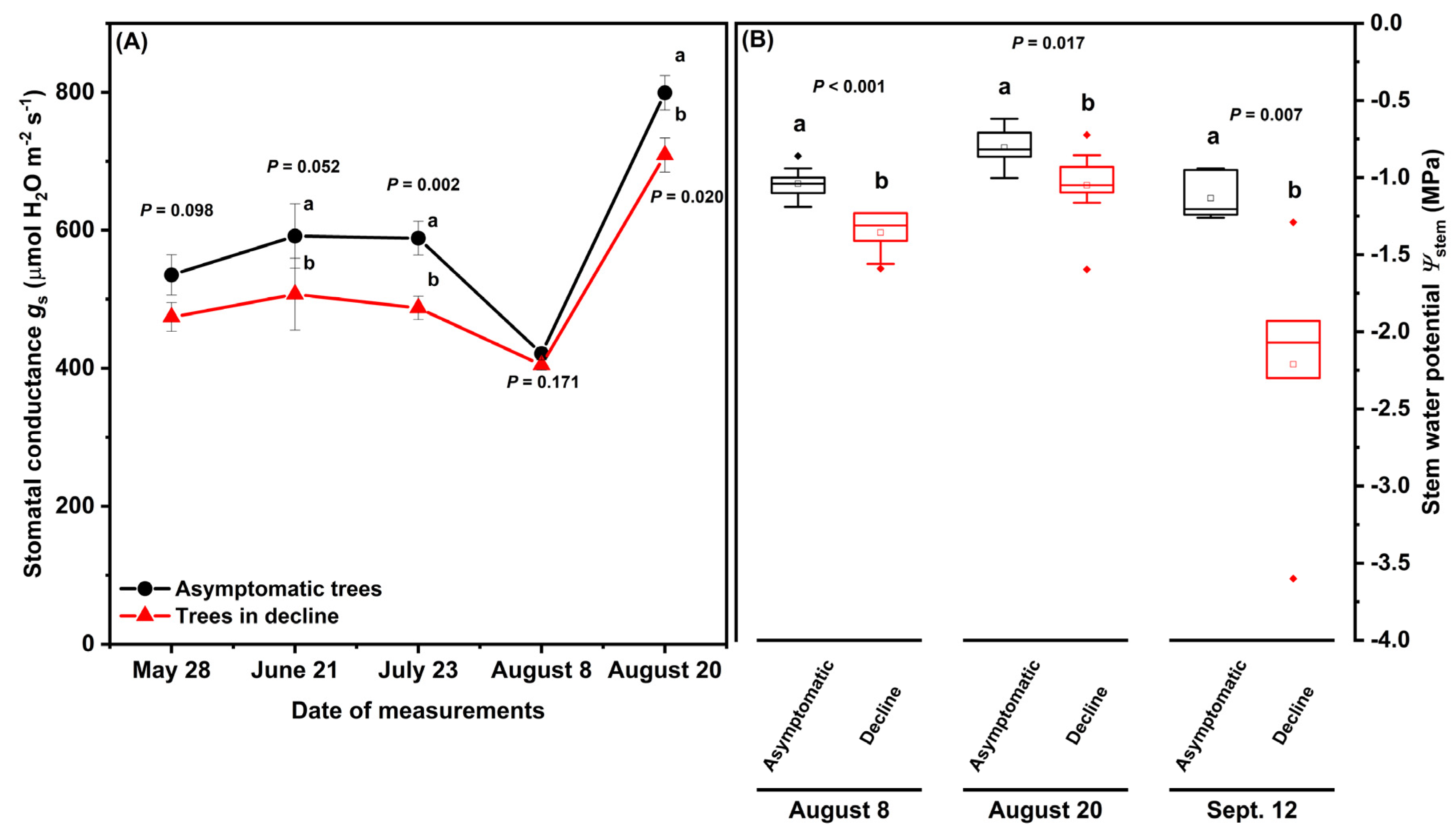
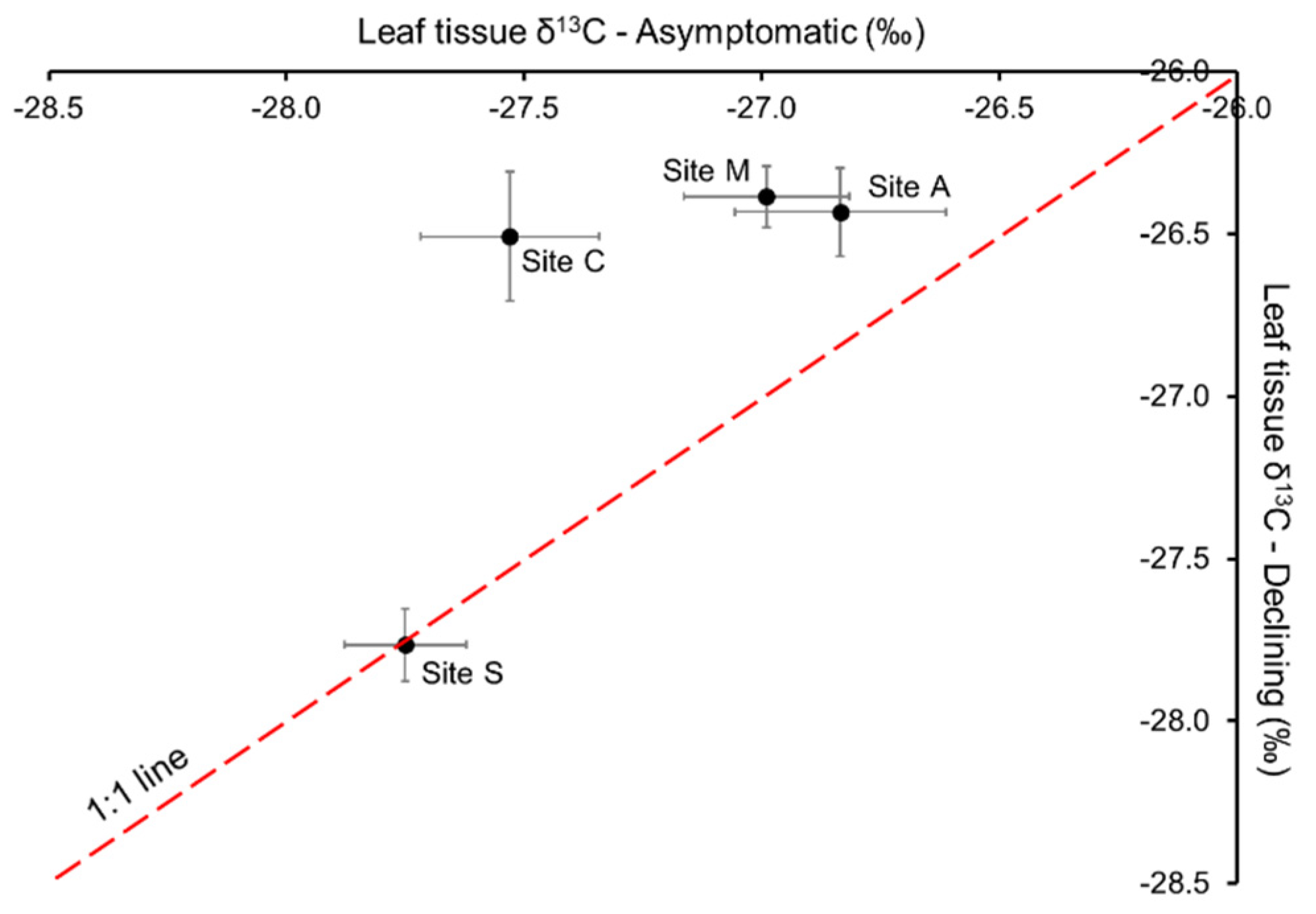
2.2. Water Relations and Fruit Carbohydrate Accumulation during Tree Decline
3. Discussion
3.1. Hydraulic Dysfunctions under Environmental Stresses
3.2. Horticultural Mitigation
3.3. Limitations and Future Perspectives
4. Materials and Methods
4.1. Site Selection and Symptom Description
4.2. Staining to Characterize Transporting Xylem Tissue
4.3. Tree Water Relations Measurement
4.4. Soil Organic Matter and Soil pH
4.5. Apple Fruit Quality Assessment
4.6. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rosenberger, D. Sudden Apple Decline: Trunk-Related Problems in Apples. Cornell University. 2018. Available online: http://www.hort.cornell.edu/expo/proceedings/2017/TreeFruitPestMGMT.AppleTrunkDisorders.Rosenberger.2017.pdf (accessed on 17 November 2022).
- Villani, S.M.; Calvin, J.; Kreis, R.; Schoof, S.; Walgenbach, J.F. Defining factors associated with rapid apple decline in the Southeastern United States. In Proceedings of the International Congress of Plant Pathology (ICPP) 2018: Plant Health in A Global Economy, St. Paul, MN, USA, 29 July–3 August 2018. [Google Scholar]
- Singh, J.; Silva, K.J.P.; Fuchs, M.; Khan, A. Potential role of weather, soil and plant microbial communities in rapid decline of apple trees. PLoS ONE 2019, 14, e0213293. [Google Scholar] [CrossRef] [PubMed]
- Stokstad, E. Rapid apple decline has researchers stumped. Science 2019, 363, 1259. [Google Scholar] [CrossRef]
- MacDonald, J.L.; Hannam, K.; Xu, H. Signs and symptoms of sudden apple decline in British Columbia, impacts on tree physiology, and the potential role of environmental stressors. In Proceedings of the Canadian Phytopathological Society Annual Meeting, Virtual, 4–8 July 2022. [Google Scholar]
- Xiao, H.; Hao, W.; Storoschuk, G.; MacDonald, J.L.; Sanfaçon, H. Characterizing the Virome of Apple Orchards Affected by Rapid Decline in the Okanagan and Similkameen Valleys of British Columbia (Canada). Pathogens 2022, 11, 1231. [Google Scholar] [CrossRef] [PubMed]
- Landsberg, J.; Waring, R.; Ryan, M. Water relations in tree physiology: Where to from here? Tree Physiol. 2017, 37, 18–32. [Google Scholar] [CrossRef] [PubMed]
- Cohen, S.; Naor, A.; Bennink, J.; Grava, A.; Tyree, M. Hydraulic resistance components of mature apple trees on rootstocks of different vigours. J. Exp. Bot. 2007, 58, 4213–4224. [Google Scholar] [CrossRef] [PubMed]
- Bauerle, T.L.; Centinari, M.; Bauerle, W.L. Shifts in xylem vessel diameter and embolisms in grafted apple trees of differing rootstock growth potential in response to drought. Planta 2011, 234, 1045–1054. [Google Scholar] [CrossRef]
- Olien, W.; Lakso, A.N. Effect of rootstock on apple (Malus domestica) tree water relations. Physiol. Plant. 1986, 67, 421–430. [Google Scholar] [CrossRef]
- Atkinson, C.J.; Webster, A.D.; Vaughan, S.; Lucas, A.S. Effects of root restriction on the physiology of apple tree growth. Acta Hortic. 1997, 451, 587–598. [Google Scholar] [CrossRef]
- Jones, H.G. How do rootstocks control shoot water relations? New. Phytol. Found. 2012, 194, 301–303. [Google Scholar] [CrossRef]
- Tworkoski, T.; Fazio, G. Effects of size-controlling apple rootstocks on growth, abscisic acid, and hydraulic conductivity of scion of different vigor. Int. J. Fruit Sci. 2015, 15, 369–381. [Google Scholar] [CrossRef]
- Marini, R.P.; Fazio, G. Apple rootstocks: History, physiology, management and breeding—Stresses influencing rootstock performance. In Horticultural Reviews; Warrington, I., Ed.; John Wiley & Sons, Inc.: Oxford, UK, 2018; Volume 45, pp. 225–258. [Google Scholar]
- Xu, H.; Ediger, D. Rootstocks with different vigor influenced scion–water relations and stress responses in AmbrosiaTM apple trees (Malus domestica var. Ambrosia). Plants 2021, 10, 614. [Google Scholar] [CrossRef]
- Xu, H.; Watanabe, Y.; Ediger, D.; Yang, X.; Iritani, D. Characteristics of sunburn browning fruit and rootstock-dependent damage-free yield of Ambrosia™ apple after sustained summer heat events. Plants 2022, 11, 1201. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Blatt, S.; Ediger, D. Tools for climate resilience in tree fruit I: Large-dwarfing rootstocks can alleviate sunburn damage in “Buckeye Gala” apple. Can. J. Plant Sci. 2022. e-First. [Google Scholar] [CrossRef]
- Rasool, A.; Mansoor, S.; Bhat, K.; Hassan, G.; Baba, T.; Alyemeni, M.; Alsahli, A.; El-Serehy, H.; Paray, B.; Ahmad, P. Mechanisms underlying graft union formation and rootstock scion interaction in horticultural plants. Front. Plant Sci. 2020, 11, 590847. [Google Scholar] [CrossRef]
- Klein, T.; Hoch, G.; Yakir, D.; Körner, C. Drought stress, growth and nonstructural carbohydrate dynamics of pine trees in a semi-arid forest. Tree Physiol. 2014, 34, 981–992. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, C.M.; Priestley, C.A. Carbohydrate reserves in deciduous fruit trees. Hort. Rev. 1988, 10, 403–430. [Google Scholar]
- Cailleret, M.; Jansen, S.; Robert, E.M.; Desoto, L.; Aakala, T.; Antos, J.A.; Beikircher, B.; Bigler, C.; Bugmann, H.; Caccianiga, M.; et al. A synthesis of radial growth patterns preceding tree mortality. Glob. Change Biol. 2017, 23, 1675–1690. [Google Scholar] [CrossRef]
- Robinson, T. Apple Rootstock Performance—An Eastern Perspective. SCRI Apple Root to Fruit Webinar on Rootstock and Nutrition 2022. Virtual. March 1 2022. Available online: https://youtube.com/playlist?list=PLYLbxsK4pTXXoyNLVkWZjKprOVOkkoQCL (accessed on 20 March 2022).
- Pacific Northwest Pest Management Handbooks. Insect/Tree Fruit Crops/Apple Pest—Apple-Shothole Borer. A Pacific northwest Extension Publication. Available online: https://pnwhandbooks.org/insect/tree-fruit/apple/apple-shothole-borer (accessed on 30 December 2022).
- Úrbez-Torres, J.R.; Boule, J.; O’Gorman, D.T. First report of Diplodia seriata and D. mutila causing apple dieback in British Columbia. Plant Dis. 2016, 100, 1243. [Google Scholar] [CrossRef]
- Trifilò, P.; Kiorapostolou, N.; Petruzzellis, F.; Vitti, S.; Petit, G.; Gullo, M.A.L.; Nardini, A.; Casolo, V. Hydraulic recovery from xylem embolism in excised branches of twelve woody species: Relationships with parenchyma cells and non-structural carbohydrates. Plant Physiol. Biochem. 2019, 139, 513–520. [Google Scholar] [CrossRef]
- Choi, B.H.; Bhusal, N.; Jeong, W.T.; Park, I.H.; Han, S.G.; Yoon, T.M. Waterlogging tolerance in apple trees grafted on rootstocks from G, CG, and M series. Hort. Environ. Biotech. 2020, 61, 685–692. [Google Scholar] [CrossRef]
- Zhang, P.; Cui, Z.; Xu, H.; Ali, A.; Zhang, X.; Liu, X.; Zhang, Y.; Zhou, X.; Lu, Z. Thirst or Malnutrition: The impacts of invasive insect Agrilus mali on the physiological status of wild apple trees. Forests 2020, 11, 440. [Google Scholar] [CrossRef] [Green Version]
- Farquar, G.D.; Ehleringer, J.R.; Hubick, K.T. Carbon isotope discrimination and photosynthesis. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1989, 40, 503–537. [Google Scholar] [CrossRef]
- Casagrande Biasuz, E.; Kalcsits, L.A. Apple rootstocks affect functional leaf traits with consequential effects on carbon isotope composition and vegetative vigour. AoB Plants 2022, 14, plac020. [Google Scholar] [CrossRef]
- Kalcsits, L.; Valverdi, N.; Reid, M. Timing of water limitations affect source to sink differences in δ13C composition in apple. Acta Hortic. 2022, 1335, 437–444. [Google Scholar] [CrossRef]
- Valverdi, N.A.; Kalcsits, L. Apple rootstock genotype affects scion responses to water limitations under field conditions. Acta Physiol. Plantatum 2022, 43, 97. [Google Scholar] [CrossRef]
- Zebarth, B.J.; Neilsen, G.H.; Hogue, E.; Neilsen, D. Influence of organic waste on selected soil physical and chemical properties. Can. J. Soil Sci. 1999, 79, 501–504. [Google Scholar] [CrossRef]
- Neilsen, G.H.; Hogue, E.J.; Forge, T.; Neilsen, D. Surface application of mulches and biosolids affect orchard soil properties after 7 years. Can. J. Soil Sci. 2003, 83, 131–137. [Google Scholar] [CrossRef]
- Environment Canada. Station Results. Historic Data; Station: Summerland, BC. Available online: https://climate.weather.gc.ca/historical_data/search_historic_data_e.html (accessed on 5 April 2021).
- O’Leary, M.H. Carbon isotopes in photosynthesis. Bioscience 1988, 38, 328–336. [Google Scholar] [CrossRef]
- Suni, M.; Nyman, M.; Eriksson, N.A.; Björk, L.; Björck, I. Carbohydrate composition and content of organic acids in fresh and stored apples. J. Sci. Food Agric. 2000, 80, 1538–1544. [Google Scholar] [CrossRef]
- Andrews, P.K.; Marquez, C.S. Graft incompatibility. Hortic. Rev. 2010, 15, 183–232. [Google Scholar]
- Kalcsits, L.; Asteggiano, L.; Schmidt, T.; Musacchi, S.; Serra, S.; Layne, D.R.; Mupambi, G. Shade netting reduces sunburn damage and soil moisture depletion in ‘Granny Smith’ apples. Acta Hortic. 2018, 1228, 85–90. [Google Scholar] [CrossRef]
- Mupambi, G.; Anthony, B.M.; Layne, D.R.; Musacchi, S.; Serra, S.; Schmidt, T.; Kalcsits, L.A. The influence of protective netting on tree physiology and fruit quality of apple: A review. Sci. Hortic. 2018, 236, 60–72. [Google Scholar]
- Boini, A.; Manfrini, L.; Morandi, B.; Corelli Grappadelli, L.; Predieri, S.; Daniele, G.M.; López, G. High levels of shading as a sustainable application for mitigating drought, in modern apple production. Agronomy 2021, 11, 422. [Google Scholar] [CrossRef]
- Lulane, E.B.; Dzikiti, S.; Volschenk, T.; Lötze, E.; Midgley, S.J.E. Quantifying water saving benefits of fixed white protective netting in irrigated apple orchards under Mediterranean-type climate conditions in South Africa. Sci. Hortic. 2022, 305, 111439. [Google Scholar] [CrossRef]
- Naschitz, S.; Naor, A.; Genish, S.; Wolf, S.; Goldschmidt, E.E. Internal management of non-structural carbohydrate resources in apple leaves and branch wood under a broad range of sink and source manipulations. Tree Physiol. 2010, 30, 715–727. [Google Scholar] [CrossRef]
- Tomasella, M.; Petrussa, E.; Petruzzellis, F.; Nardini, A.; Casolo, V. The possible role of non-structural carbohydrates in the regulation of tree hydraulics. Int. J. Mol. Sci. 2019, 21, 144. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.K.; Park, J.; Hwang, I. Investigating water transport through the xylem network in vascular plants. J. Exp. Bot. 2014, 65, 1895–1904. [Google Scholar] [CrossRef]
- Olmstead, M.A.; Lang, N.S.; Lang, G.A.; Ewers, F.W.; Owens, S.A. Examining the vascular pathway of sweet cherries grafted onto dwarfing rootstocks. HortScience 2006, 41, 674–679. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Fields, J.S.; Fonteno, W.C.; Jackson, B.E.; Heitman, J.L.; Owen Jr, J.S. The Use of Dewpoint Hygrometry to Measure Low Water Potentials in Soilless Substrate Components and Composites. Agronomy 2020, 10, 1393. [Google Scholar] [CrossRef]
- Scholander, P.F.; Bradstreet, E.D.; Hemmingsen, E.A.; Hammel, H.T. Sap Pressure in Vascular Plants: Negative hydrostatic pressure can be measured in plants. Science 1965, 148, 339–346. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Site | Number of Trees | Scion | Rootstock | Planting Date | Soil Texture | SOM (%) | Soil pH |
|---|---|---|---|---|---|---|---|
| A | 10 | Salish | M9 | 2013 | S, LS, SL | 4.3 ± 0.3 | 6.0 ± 0.1 |
| S | 12 | Ambrosia | M9 | 2003 | S, LS | 1.6 ± 0.1 | 6.8 ± 0.1 |
| C | 10 | Ambrosia | M9 | 2009 | LS, SL | 2.0 ± 0.1 | 7.8 ± 0.1 |
| M | 10 | Ambrosia | M9 | 2012 | LS, SL | 2.6 ± 0.3 | 6.2 ± 0.1 |
| P | 6 | Ambrosia | M9/B9 | 2003 | LS, SL | 2.2 ± 0.2 | 6.2 ± 0.2 |
| Year | Site | Status | Fruit Mass (g) | DMC (%) | Fruit DM (g) | SSC (%) | Firmness (lbs) | Ψfruit (MPa) |
|---|---|---|---|---|---|---|---|---|
| 2019 | S | A | 164.93 ± 6.13 | 11.99 ± 0.20 | 21.55 ± 0.91 | 13.13 ± 0.18 | 16.81 ± 0.16 | −1.69 ± 0.05 a |
| D | 128.76 ± 4.96 * | 11.73 ± 0.15 * | 18.34 ± 0.58 * | 12.78 ± 0.17 | 17.70 ± 0.21 * | −1.66 ± 0.05 a | ||
| F(1,118) = 89.9 p < 0.001 | F(1,118) = 3.9 p = 0.0497 | F(1,118) = 80.9 p < 0.001 | F(1,22) = 2.0 p = 0.17 | F(1,58) = 11.2 p < 0.01 | F(1,16) = 0.2 p = 0.65 | |||
| C | A | 238.96 ± 6.51 | 14.56 ± 0.09 | 34.75 ± 0.93 | 14.11 ± 0.24 | 12.28 ± 0.28 | −1.96 ± 0.04 b | |
| D | 198.77 ± 5.28 * | 14.68 ± 0.08 | 29.16 ± 0.77 * | 17.21 ± 0.09 * | 12.05 ± 0.28 | −2.39 ± 0.04 c | ||
| F(1,98) = 23.0 p < 0.001 | F(1,98) = 1.0 p = 0.33 | F(1,98) = 21.2 p < 0.001 | F(1,38) = 171.5 p < 0.001 | F(1,55) = 0.4 p = 0.55 | F(1,28) = 65.3 p < 0.001 | |||
| M | A | 275.80 ± 4.37 | 13.94 ± 0.06 | 38.45 ± 0.64 | 14.13 ± 0.09 | 14.92 ± 0.20 | −1.89 ± 0.03 b | |
| D | 237.11 ± 5.24 * | 13.82 ± 0.07 | 32.74 ± 0.72 * | 14.59 ± 0.12 * | 15.33 ± 0.23 | −1.87 ± 0.03 b | ||
| F(1,98) = 32.1 p < 0.001 | F(1,98) = 1.6 p = 0.21 | F(1,98) = 35.0 p < 0.001 | F(1,43) = 10.0 p = 0.003 | F(1,58) = 1.8 p = 0.19 | F(1,38) = 0.2 p = 0.69 | |||
| 2020 | S | A | 242.75 ± 7.05 | 15.60 ± 0.45 | 37.94 ± 1.04 | 13.78 ± 0.16 | 17.15 ± 0.46 | −1.95 ± 0.02 C |
| D | 244.80 ± 6.62 | 15.56 ± 0.68 | 38.24 ± 1.03 | 13.82 ± 0.28 | 17.39 ± 0.29 | −1.96 ± 0.02 C | ||
| F(1,93) = 0.04 p = 0.83 | F(1,93) = 0.05 p = 0.83 | F(1,112) = 0.04 p = 0.84 | F(1,17) = 0.04 p = 0.85 | F(1,17) = 0.24 p = 0.63 | F(1,28) = 0.02 p = 0.88 | |||
| C | A | 207.38 ± 7.91 | 14.65 ± 0.16 | 30.43 ± 1.38 | 12.65 ± 0.26 | 17.50 ± 0.31 | −1.87 ± 0.02 A | |
| D | 127.76 ± 6.66 * | 15.26 ± 0.56 | 19.59 ± 1.37 * | 13.39 ± 0.51 * | 21.93 ± 1.11 * | −2.09 ± 0.04 BC | ||
| F(1,99) = 104.8 p < 0.001 | F(1,99) = 3.6 p = 0.06 | F(1,99) = 77.8 p < 0.001 | F(1,14) = 4.9 p = 0.04 | F(1,18) = 14.7 p = 0.001 | F(1,23) = 8.7 p = 0.007 | |||
| M | A | 283.89 ± 4.86 | 15.41 ± 0.08 | 43.78 ± 0.71 | 13.77 ± 0.12 | 18.69 ± 0.40 | −1.87 ± 0.01 BC | |
| D | 177.28 ± 7.67 * | 16.07 ± 0.33 * | 28.49 ± 1.33 * | 15.00 ± 1.14 * | 20.87 ± 0.51 * | −2.22 ± 0.04 D | ||
| F(1,98) = 322.4 p < 0.001 | F(1,98) = 10.8 p = 0.001 | F(1,98) = 223.0 p < 0.001 | F(1,18) = 12.0 p = 0.003 | F(1,18) = 11.3 p = 0.003 | F(1,28) = 35.8 p < 0.001 | |||
| P | A | 273.69 ± 9.31 | 14.72 ± 0.23 | 40.47 ± 1.88 | 12.68 ± 0.18 | 17.36 ± 0.25 | −1.73 ± 0.01 A | |
| D | 225.57 ± 9.25 * | 14.74 ± 0.20 | 33.08 ± 1.14 * | 12.91 ± 0.35 | 18.35 ± 0.40 * | −1.76 ± 0.01 A | ||
| F(1,98) = 28.2 p < 0.001 | F(1,98) = 0.1 p = 0.75 | F(1,98) = 27.4 p < 0.001 | F(1,18) = 0.3 p = 0.58 | F(1,18) = 4.5 p < 0.05 | F(1,28) = 0.4 p = 0.53 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, H.; Hannam, K.D.; MacDonald, J.L.; Ediger, D. Field Investigation into Tree Fates from Recent Apple Tree Decline: Abrupt Hydraulic Failure versus Gradual Hydraulic Loss. Stresses 2023, 3, 256-269. https://doi.org/10.3390/stresses3010019
Xu H, Hannam KD, MacDonald JL, Ediger D. Field Investigation into Tree Fates from Recent Apple Tree Decline: Abrupt Hydraulic Failure versus Gradual Hydraulic Loss. Stresses. 2023; 3(1):256-269. https://doi.org/10.3390/stresses3010019
Chicago/Turabian StyleXu, Hao, Kirsten D. Hannam, Jesse L. MacDonald, and Danielle Ediger. 2023. "Field Investigation into Tree Fates from Recent Apple Tree Decline: Abrupt Hydraulic Failure versus Gradual Hydraulic Loss" Stresses 3, no. 1: 256-269. https://doi.org/10.3390/stresses3010019