
Protective Effect of Biflavones from Platonia insignis Mart. against Ethidium Bromide-Induced Lethality in Staphylococcus aureus
 , , and
, , and 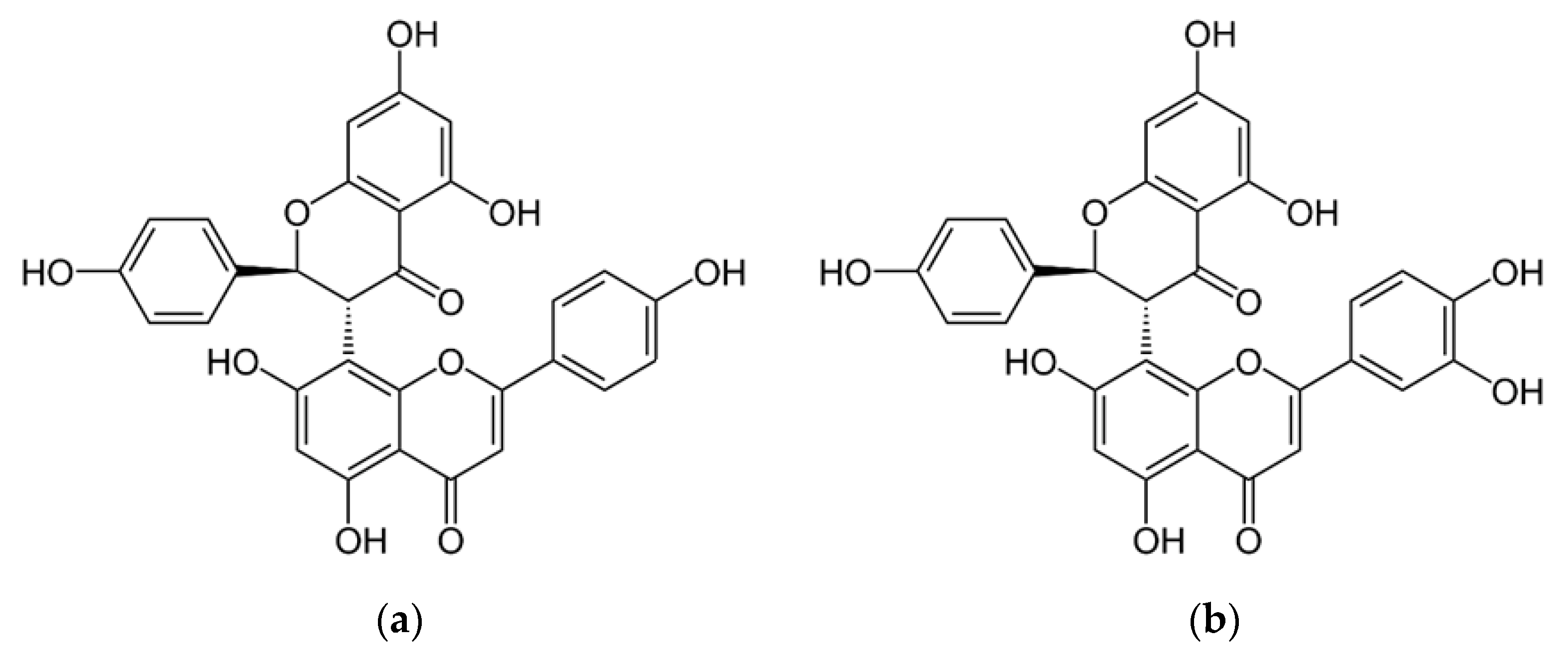{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
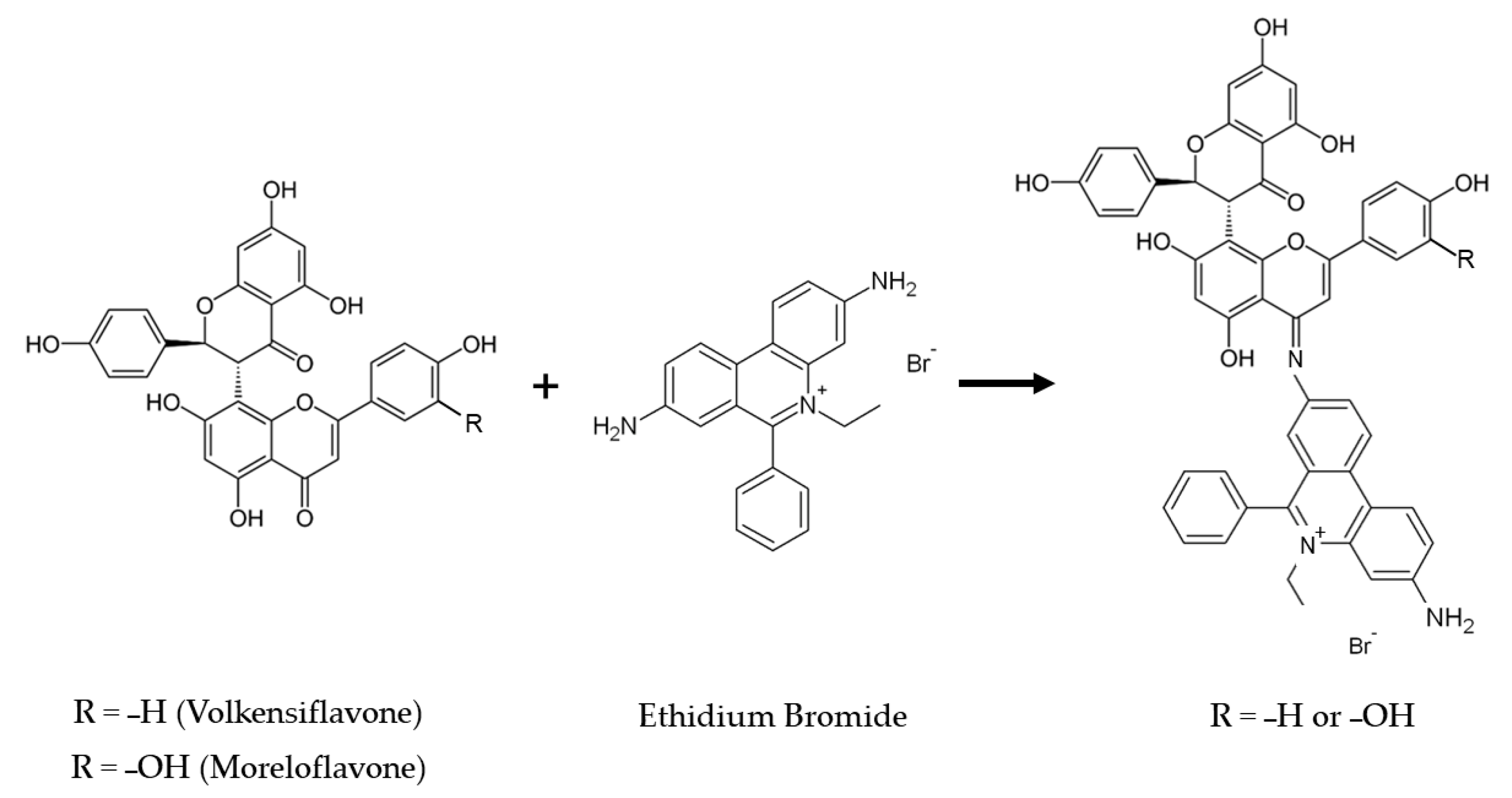
2.1. Bacterial Strains and Chemical Compounds
2.2. Extraction of the Biflavones Mixture
2.3. Determination of the Minimum Inhibitory Concentration
2.4. Evaluation of the Protective Effect of BFs against Lethality Induced by EtBr in Staphylococcus aureus
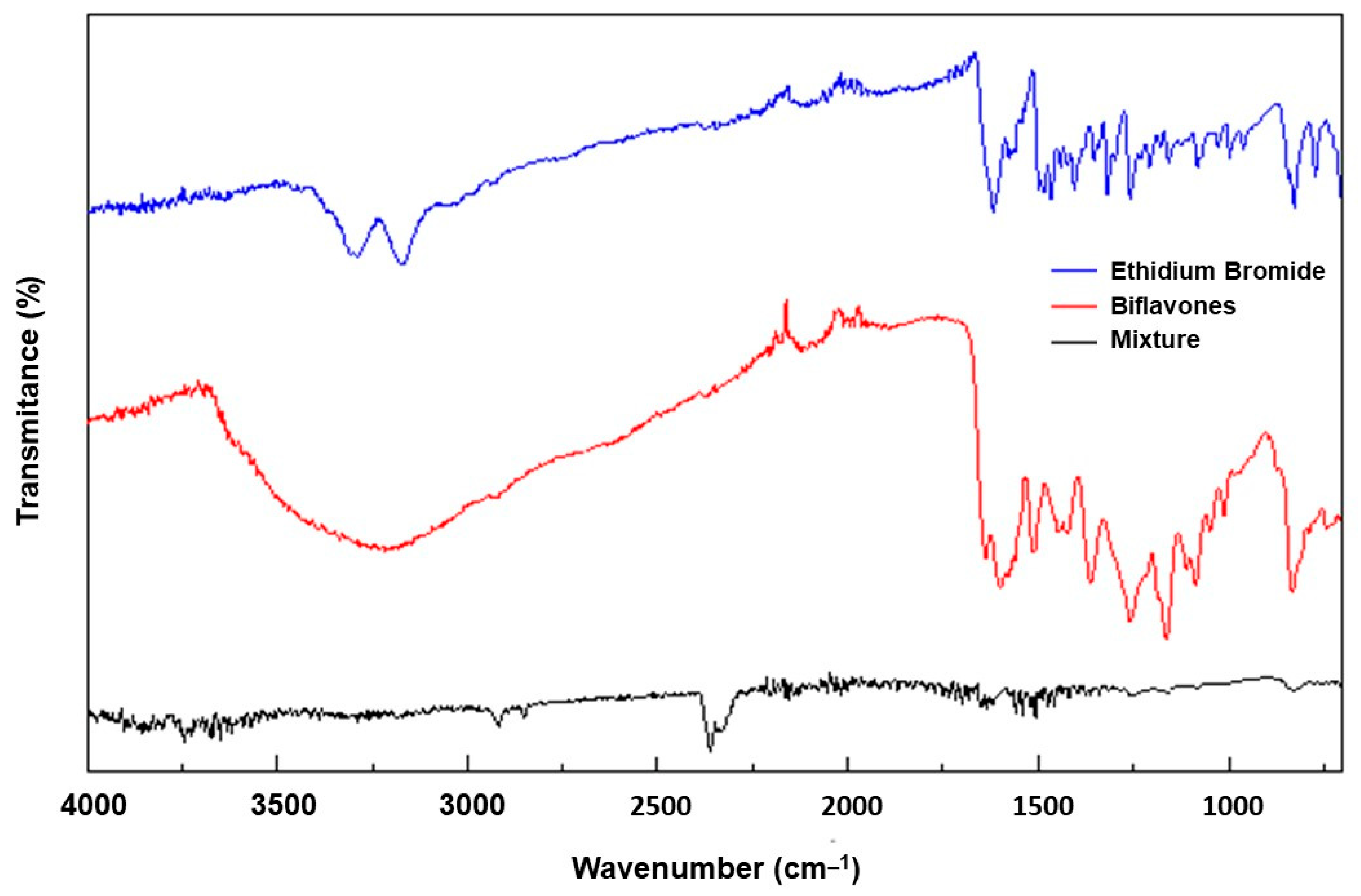
2.5. Spectroscopic Analysis in the Infrared Region
2.6. Statistical Analysis
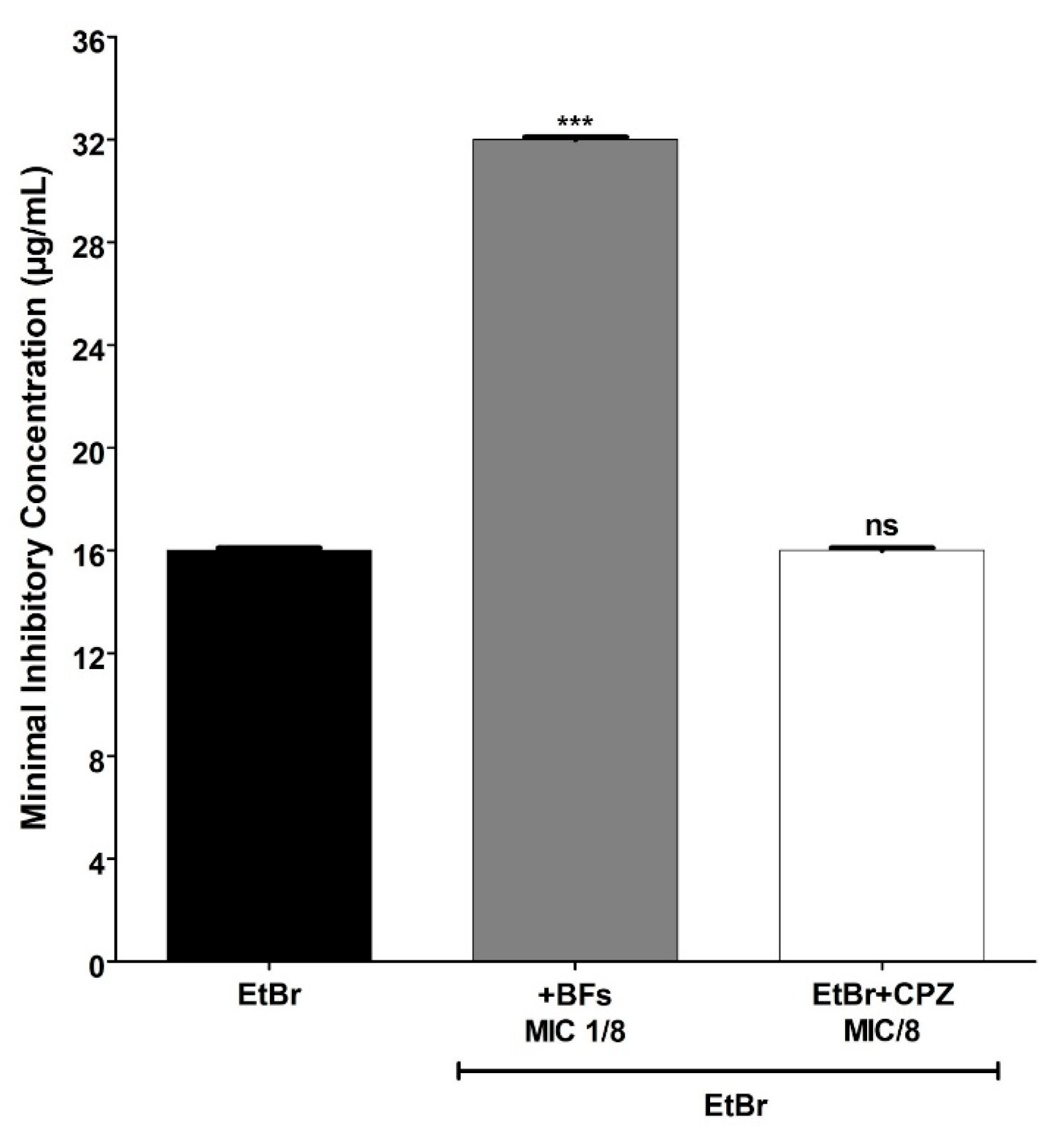
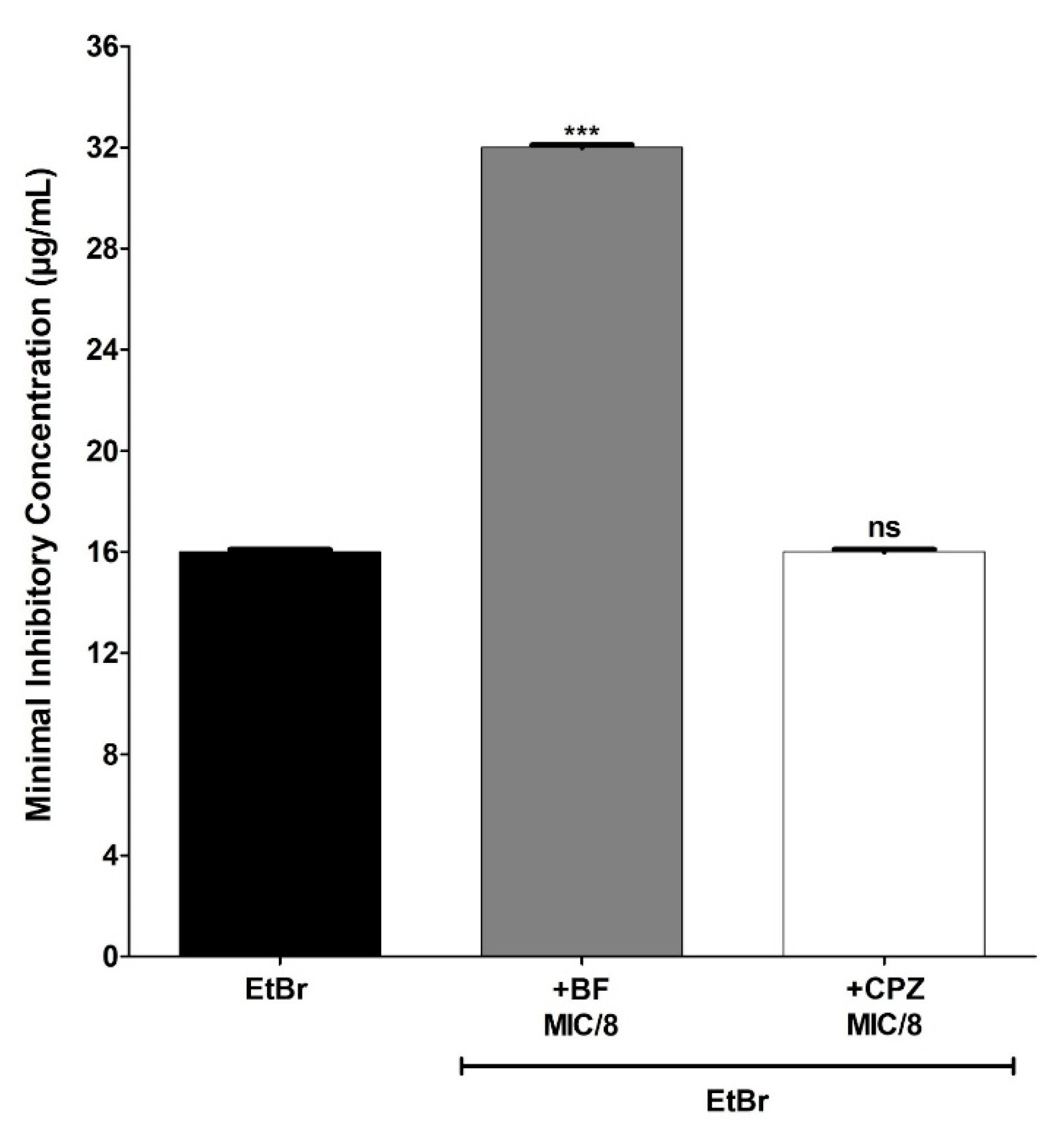
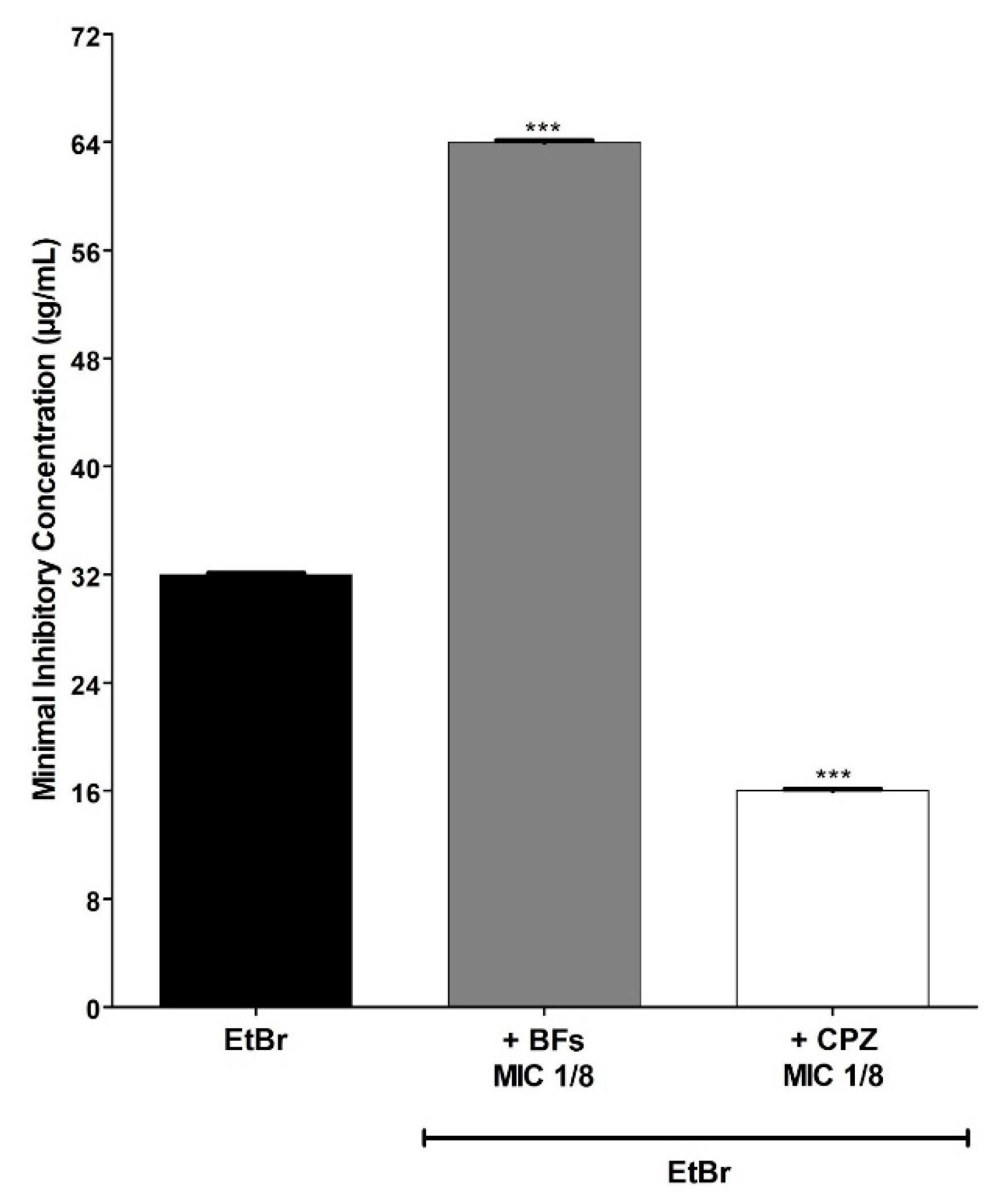
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Galindo-Murillo, R.; Cheatham, T.E. Ethidium bromide interactions with DNA: An exploration of a classic DNA-ligand complex with unbiased molecular dynamics simulations. Nucleic Acids Res. 2021, 49, 3735–3747. [Google Scholar] [CrossRef] [PubMed]
- LePecq, J.B.; Paoletti, C.A. Fluorescent complex between ethidium bromide and nucleic acids: Physical-chemical characterization. J. Mol. Biol. 1967, 27, 87–106. [Google Scholar] [CrossRef] [PubMed]
- Burns, V.W.F. Fluorescence decay time characteristics of the complex between ethidium bromide and nucleic acids. Arch. Biochem. Biophys. 1969, 133, 420–424. [Google Scholar] [CrossRef] [PubMed]
- Possato, B.; Dalmolin, L.F.; Pereira, L.M.; Alves, J.Q.; Silva, R.T.C.; Gelamo, R.V.; Yatsuda, A.P.; Lopez, R.F.V.; Albuquerque, S.D.; Leite, N.B.; et al. Gold (III) complexes with thiosemicarbazonate ligands as potential anticancer agents: Cytotoxicity and interactions with biomolecular targets. Eur. J. Pharm. Sci. 2021, 162, 105834. [Google Scholar] [CrossRef] [PubMed]
- Burns, V.W.F. Fluorescence polarization characteristics of the complexes between ethidium bromide and rRNA, tRNA, and DNA. Arch. Biochem. Biophys. 1971, 145, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Alnajrani, M.N.; Alsager, O.A. Decomposition of DNA staining agent ethidium bromide by gamma irradiation: Conditions, kinetics, by-products, biological activity, and removal from wastewater. J. Hazard. Mater. 2020, 389, 122142. [Google Scholar] [CrossRef]
- Burns, V.W.F. Location and molecular characteristics of fluorescent complexes of ethidium bromide in the cell. Exp. Cell Res. 1972, 75, 200–206. [Google Scholar] [CrossRef]
- Braga, R. Plantas do Nordeste: Especialmente do Ceará, 2nd ed.; Imprensa Official: Fortaleza, Brazil, 1960; 540p. [Google Scholar]
- Cavalcante, P.B. Frutas Comestíveis da Amazônia, 1st ed.; Museu Paraense Emílio Goeldi: Pará, Brazil, 1996; 279p. [Google Scholar]
- Nascimento, W.M.O.D.; Carvalho, J.E.U.D.; Muller, C.H. Ocorrência e distribuição geográfica do bacurizeiro. Rev. Bras. Frutic. 2007, 29, 657–660. [Google Scholar] [CrossRef]
- Sinimbú-Neto, F.D.A.; Martins, A.B.G.; Barbosa, J.C. Viabilidade in vitro de grãos de pólen de bacurizeiro—Clusiaceae. Rev. Bras. Frutic. 2011, 33, 593–600. [Google Scholar] [CrossRef]
- Souza, A.C.; Alves, M.M.D.M.; Brito, L.M.; Oliveira, L.G.D.C.; Sobrinho-Júnior, E.P.C.; Costa, I.C.G.; Freitas, S.D.L.; Rodrigues, K.A.D.F.; Chaves, M.H.; Arcanjo, D.D.R.; et al. Platonia insignis Mart., a Brazilian Amazonian plant: The stem barks extract and its main constituent lupeol exert antileishmanial effects involving macrophages activation. eCAM 2017, 2017, 3126458. [Google Scholar] [CrossRef] [Green Version]
- Mendes, M.B.; Silva-Filho, J.C.; Sabino, C.K.B.; Arcanjo, D.D.R.; Sousa, C.M.M.; Costa, I.C.G.; Chaves, M.H.; Oliveira, R.D.C.M.; Oliveira, A.P. Pharmacological evidence of 2-adrenergic receptors in the hypotensive effect of Platonia insignis Mart. J. Med. Food 2014, 17, 1079–1085. [Google Scholar] [CrossRef] [PubMed]
- Costa-Júnior, J.S.; de Almeida, A.A.C.; Costa, J.P.; Citó, A.M.G.L.; Saffi, J.; de Freitas, R.M. Superoxide dismutase and catalase activities in rat hippocampus pretreated with garcinielliptone FC from Platonia insignis. Pharm. Biol. 2012, 50, 453–457. [Google Scholar] [CrossRef] [PubMed]
- Costa-Júnior, J.S.; Freitas, R.M.; Citó, A.M.G.L.; Henriques, J.A.P.; Saffi, J. Evaluation of effects of ethanolic extract (EE) from Platonia insignis Mart. on pilocarpine-induced seizures. J. Biol. Sci. 2010, 10, 747–753. [Google Scholar] [CrossRef]
- Costa-Júnior, J.S.; Ferraz, A.B.; Feitosa, C.M.; Citó, A.M.G.L.; Freitas, R.M.; Saffi, J. Evaluation of antioxidant effects in vitro of garcinielliptone FC (GFC) isolated from Platonia insignis Mart. J. Med. Plants Res. 2011, 5, 293–329. [Google Scholar] [CrossRef]
- Costa-Junior, J.S.; de Almeida, A.A.C.; Ferraz, A.D.B.F.; Rossatto, R.R.; Silva, T.G.; Silva, P.B.N.; Militão, G.C.G.; Citó, A.M.G.L.; Santana, L.C.L.R.; Carvalho, F.A.D.A.; et al. Cytotoxic and leishmanicidal properties of garcinielliptone FC, a prenylated benzophenone from Platonia insignis. Nat. Prod. Res. 2013, 27, 470–474. [Google Scholar] [CrossRef]
- Silva, A.K.F.; Reis, A.C.; Pinheiro, E.E.A.; Sousa, J.N.; Oliveira, F.A.A.; Moura, A.K.S.; Lima-neto, J.S.; Citó, A.M.G.L.; Siqueira-Júnior, J.P.; Kaatz, G.W.; et al. Modulation of the drug resistance by Platonia insignis Mart. extract, ethyl acetate fraction and Morelloflavone/Volkensiflavone (Biflavonoids) in Staphylococcus aureus strains overexpressing efflux pump genes. Curr. Drug Metab. 2021, 21, 114–122. [Google Scholar] [CrossRef]
- Bezerra, E.A.; Alves, M.M.M.; Lima, S.K.R.; Pinheiro, E.E.A.; Amorim, L.V.; Lima Neto, S.K.R.; Carvalho, F.A.A.; Citó, A.M.G.L.; Arcanjo, D.D.R. Biflavones from Platonia insignis Mart. Flowers promote In Vitro antileishmanial and immunomodulatory effects Against internalized amastigote forms of Leishmania amazonensis. Pathogens 2021, 10, 1166. [Google Scholar] [CrossRef]
- Verdi, L.G.; Pizzolatti, M.G.; Montanher, A.B.P.; Brighente, I.M.C.; Smânia Júnior, A.; Smânia, E.D.F.A.; Simionatto, E.L.; Monache, F.D. Antibacterial and brine shrimp lethality tests of biflavonoids and derivatives of Rheedia gardneriana. Fitoterapia 2004, 75, 360–363. [Google Scholar] [CrossRef]
- Lin, Y.M.; Anderson, H.; Flavin, M.T.; Pai, Y.H.S.; Mata-Greenwood, E.; Pengsuparp, T.; Pezzuto, J.M.; Schinazi, R.F.; Hughes, S.H.; Chen, F.C. In Vitro anti-HIV activity of biflavonoids isolated from Rhus succedanea and Garcinia multiflora. J. Nat. Prod. 1997, 60, 884–888. [Google Scholar] [CrossRef]
- Gontijo, V.S.; De Souza, T.C.; Rosa, I.A.; Soares, M.G.; Da Silva, M.A.; Vilegas, W.; Viegas, C.; Dos Santos, M.H. Isolation and evaluation of the antioxidant activity of phenolic constituents of the Garcinia brasiliensis epicarp. Food Chem. 2012, 132, 1230–1235. [Google Scholar] [CrossRef] [Green Version]
- Otuki, M.F.; Bernardi, C.A.; Prudente, A.S.; Laskoski, K.; Gomig, F.; Horinouchi, C.D.S.; Guimarães, C.L.; Ferreira, J.; Delle-Monache, F.; Cechinel-Filho, V.; et al. Garcinia gardneriana (Planchon & Triana) Zappi. (Clusiaceae) as a topical anti-inflammatory alternative for cutaneous inflammation. Basic Clin. Pharmacol. Toxicol. 2011, 109, 56–62. [Google Scholar]
- Tuansulong, K.A.; Hutadilok-Towatana, N.; Mahabusarakam, W.; Pinkaew, D.; Fujise, K. Morelloflavone from Garcinia dulcis as a novel biflavonoid inhibitor of HMG-CoA reductase. Phyther. Res. 2011, 25, 424–428. [Google Scholar] [CrossRef] [PubMed]
- Santos, J.F.S.; Tintino, S.R.; Silva, A.R.P.; Barbosa, C.R.S.; Scherf, J.R.; Silveira, Z.S.; Freitas, T.S.; Lacerda-Neto, L.J.; Barros, L.M.; Menezes, I.R.A.; et al. Enhancement of the antibiotic activity by quercetin against Staphylococcus aureus efflux pumps. J. Bioenerg. Biomembr. 2021, 53, 157–167. [Google Scholar] [CrossRef]
- Hedge, A.H.; Prashanth, S.N.; Seetharamappa, J. Interaction of antioxidant flavonoids with calf thymus DNA analyzed by spectroscopic and electrochemical methods. J. Pharm. Biomed. Anal. 2012, 63, 40–46. [Google Scholar] [CrossRef]
- Neyfakh, A.A.; Borsch, C.M.; Kaatz, G.W. Fluoroquinolone resistance protein NorA of Staphylococcus aureus is a multidrug efflux transporter. Antimicrob. Agents Chemother. 1993, 37, 128–129. [Google Scholar] [CrossRef] [PubMed]
- Schindler, B.D.; Kaatz, G.W. Multidrug efflux pumps of Gram-positive bacteria. Drug Resist. Updat. 2016, 27, 1–13. [Google Scholar] [CrossRef]
- Du, D.; Wang-Kan, X.; Neuberger, A.; van Veen, H.W.; Pos, K.M.; Piddock, L.J.; Luisi, B.F. Multidrug efflux pumps: Structure, function and regulation. Nat. Rev. Microbiol. 2018, 16, 523–539. [Google Scholar] [CrossRef]
- Shriram, V.; Khare, T.; Bhagwat, R.; Shukla, R.; Kumar, V. Inhibiting bacterial efflux pumps via phyto-therapeutics to combat threatening antimicrobial resistance. Front. Microbiol. 2018, 9, 2990. [Google Scholar] [CrossRef]
- Kim, M.; Jee, S.C.; Kim, K.S.; Kim, H.S.; Yu, K.N.; Sung, J.S. Quercetin and Isorhamnetin Attenuate Benzo [a] pyrene-Induced Toxicity by Modulating Detoxification Enzymes through the AhR and NRF2 Signaling Pathways. Antioxidants 2021, 10, 787. [Google Scholar] [CrossRef]
- Vipin, C.; Mujeeburahiman, M.; Ashwini, P.; Arun, A.B.; Rekha, P.D. Antibiofilm and cytoprotective activities of quercetin against Pseudomonas aeruginosa isolates. Lett. Appl. Microbiol. 2019, 68, 464–471. [Google Scholar] [CrossRef]
- Wang, H.L.; Xing, G.D.; Qian, Y.; Sun, X.F.; Zhong, J.F.; Chen, K.L. Dihydromyricetin attenuates heat stress-induced apoptosis in dairy cow mammary epithelial cells through suppressing mitochondrial dysfunction. Ecotoxicol. Environ. Saf. 2021, 214, 112078. [Google Scholar] [CrossRef] [PubMed]
- Geng, Y.; Wu, Z.; Buist-Homan, M.; Blokzijl, H.; Moshage, H. Hesperetin protects against palmitate-induced cellular toxicity via induction of GRP78 in hepatocytes. Toxicol. Appl. Pharmacol. 2020, 404, 115183. [Google Scholar] [CrossRef] [PubMed]










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moura, A.S.G.; Barreto, J.d.A.F.; Silva, A.K.F.e.; Nascimento, M.L.L.B.d.; Pinheiro, E.E.A.; Moura, A.K.S.; Sousa, J.M.d.C.e.; Lima-Neto, J.d.S.; Citó, A.M.d.G.L.; Coutinho, H.D.M.; et al. Protective Effect of Biflavones from Platonia insignis Mart. against Ethidium Bromide-Induced Lethality in Staphylococcus aureus. Compounds 2023, 3, 194-204. https://doi.org/10.3390/compounds3010016
Moura ASG, Barreto JdAF, Silva AKFe, Nascimento MLLBd, Pinheiro EEA, Moura AKS, Sousa JMdCe, Lima-Neto JdS, Citó AMdGL, Coutinho HDM, et al. Protective Effect of Biflavones from Platonia insignis Mart. against Ethidium Bromide-Induced Lethality in Staphylococcus aureus. Compounds. 2023; 3(1):194-204. https://doi.org/10.3390/compounds3010016
Chicago/Turabian StyleMoura, Andressa Sabrina Guimarães, Júlia de Andrade Ferreira Barreto, Andressa Kelly Ferreira e Silva, Maria Luísa Lima Barreto do Nascimento, Emanuelly Elanny Andrade Pinheiro, Arkellau Kenned Silva Moura, João Marcelo de Castro e Sousa, José de Souza Lima-Neto, Antonia Maria das Graças Lopes Citó, Henrique Douglas Melo Coutinho, and et al. 2023. "Protective Effect of Biflavones from Platonia insignis Mart. against Ethidium Bromide-Induced Lethality in Staphylococcus aureus" Compounds 3, no. 1: 194-204. https://doi.org/10.3390/compounds3010016