
Molecular Detection of Cryptosporidium Species in Wildlife and Humans at the Wildlife-Human Interface around Queen Elizabeth National Park, Uganda
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
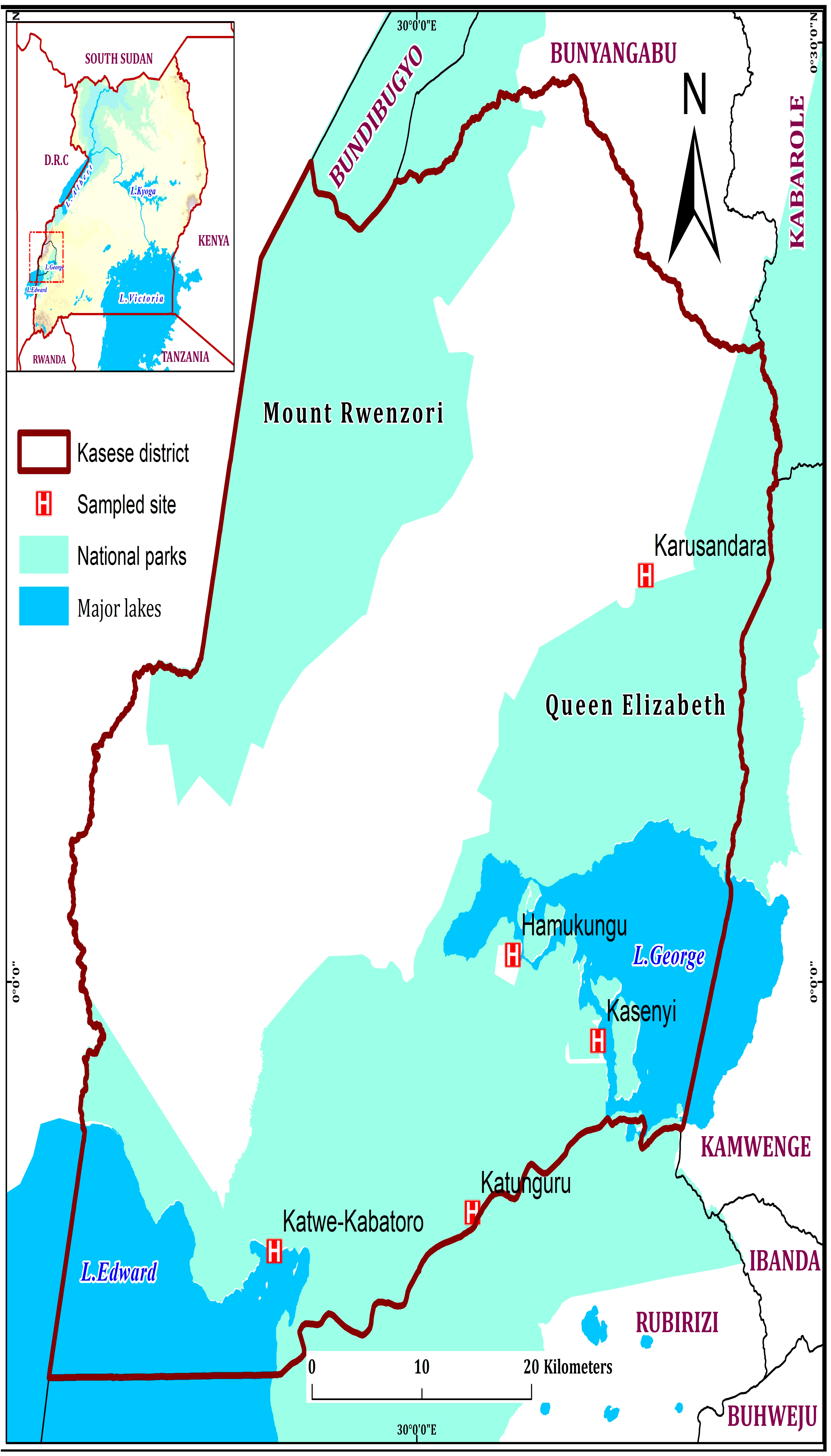
2.1. Study Area
2.2. Study Population
2.3. Study Design and Sample Calculation
Exclusion Criteria
2.4. Sample Collection, Processing, Transportation, and Storage
2.5. Microscopic Detection of Cryptosporidium Oocysts in Stool
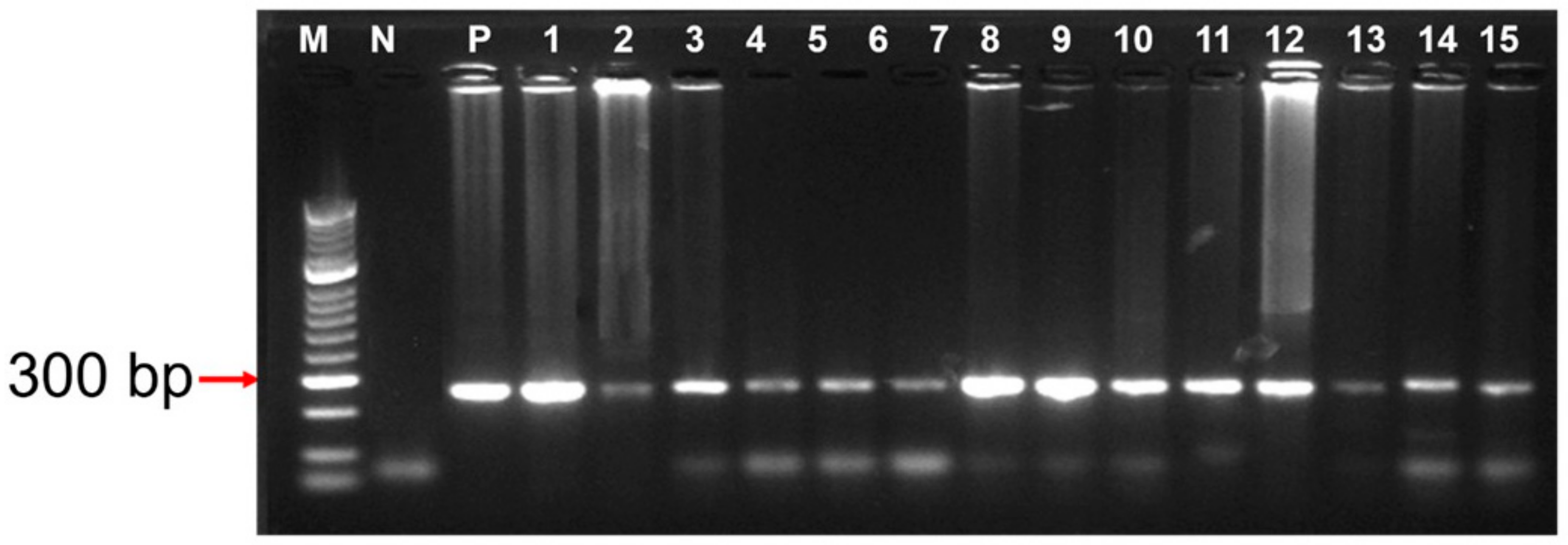
2.6. DNA Extraction and PCR Amplification
2.7. PCR Amplicon Sequencing
2.8. Data Analysis
2.9. Ethical Considerations
3. Results
3.1. Prevalence of Cryptosporidium Species
3.2. Microscopy Thick Smear Sensitivity and Specificity on Stool Samples
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Carmena, D. Waterborne transmission of Cryptosporidium and Giardia: Detection, surveillance and implications for public health. In Current Research, Technology and Education Topics in Applied Microbiology and Microbial Biotechnology; Formatex Research Center: Badajoz, Spain, 2010; Volume 20, pp. 3–4. [Google Scholar]
- Efstratiou, A.; Ongerth, J.E.; Karanis, P. Waterborne transmission of protozoan parasites: Review of worldwide outbreaks—An update 2011–2016. Water Res. 2017, 114, 14–22. [Google Scholar] [CrossRef] [PubMed]
- Ryan, U.; Hijjawi, N.; Xiao, L. Foodborne cryptosporidiosis. Int. J. Parasitol. 2018, 48, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Thompson, R.C.; Palmer, C.S.; O’Handley, R. The public health and clinical significance of Giardia and Cryptosporidium in domestic animals. Vet. J. 2008, 177, 18–25. [Google Scholar] [CrossRef]
- Savioli, L.; Smith, H.; Thompson, A. Giardia and Cryptosporidium join the ’Neglected Diseases Initiative. Trends Parasitol. 2006, 22, 203–208. [Google Scholar] [CrossRef] [PubMed]
- Kotloff, K.L.; Nataro, J.P.; Blackwelder, W.C.; Nasrin, D.; Farag, T.H.; Panchalingam, S.; Wu, Y.; Sow, S.O.; Sur, D.; Breiman, R.F.; et al. Burden and aetiology of diarrhoeal disease in infants and young children in developing countries (the Global Enteric Multicenter Study, GEMS): A prospective, case-control study. Lancet 2013, 382, 209–222. [Google Scholar] [CrossRef]
- Striepen, B. Parasitic infections: Time to tackle cryptosporidiosis. Nature 2013, 503, 189–191. [Google Scholar] [CrossRef]
- Baldursson, S.; Karanis, P. Waterborne transmission of protozoan parasites: 2004–2010. Water Res. 2011, 45, 6603–6614. [Google Scholar] [CrossRef]
- Aldeyarbi, H.M.; Abu El-Ezz, N.M.T.; Karanis, P. Cryptosporidium and cryptosporidiosis: The African perspective. Environ. Sci. Pollut. Res. Int. 2016, 23, 13811–13821. [Google Scholar] [CrossRef]
- Squire, S.A.; Ryan, U. Cryptosporidium and Giardia in Africa: Current and future challenges. Parasites Vectors 2017, 10, 195–227. [Google Scholar] [CrossRef]
- Fayer, R.; Ungar, B.L. Cryptosporidium spp. and cryptosporidiosis. Microbiol. Rev. 1986, 50, 458–483. [Google Scholar] [CrossRef]
- Cabada, M.M.; White, A.C. Treatment of cryptosporidiosis: Do we know what we think we know? Curr. Opin. Infect. Dis. 2010, 23, 494–499. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.J.; Li, J.Q.; Chen, Y.C.; Zhang, L.X.; Xiao, L.H. Widespread occurrence of Cryptosporidium infections in patients with HIV/AIDS: Epidemiology, clinical feature, diagnosis, and therapy. Acta Trop. 2018, 187, 257–263. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, R.M.; Shaffie, R.; Kang, G.; Ward, H.D. Cryptosporidiosis in patients with HIV/AIDS. Aids 2011, 25, 549–560. [Google Scholar] [CrossRef] [PubMed]
- Hotez, P.J. Could Nitazoxanide be added to other essential medicines for integrated neglected tropical disease control and elimination? PLoS Negl. Trop. Dis. 2014, 8, e2758. [Google Scholar] [CrossRef]
- Khalil, I.A.; Troeger, C.; Rao, P.C.; Blacker, B.F.; Brown, A.; Brewer, T.G.; Colombara, D.V.; De Hostos, E.L.; Engmann, C.; Guerrant, R.L.; et al. Morbidity, mortality, and long-term consequences associated with diarrhoea from Cryptosporidium infection in children younger than 5 years: A meta-analyses study. Lancet Glob. Health 2018, 6, e758–e768. [Google Scholar] [CrossRef]
- Nizeyi, J.B.; Mwebe, R.; Nanteza, A.; Cranfield, M.R.; Kalema, G.R.N.N.; Graczyk, T.K. Cryptosporidium sp. and Giardia sp. infections in mountain gorillas (Gorilla gorilla beringei) of the Bwindi Impenetrable National Park, Uganda. J. Parasitol. 1999, 85, 1084–1088. [Google Scholar] [CrossRef]
- Graczyk, T.K.; DaSilva, A.J.; Cranfield, M.R.; Nizeyi, J.B.; Kalema, G.R.; Pieniazek, N.J. Cryptosporidium parvum genotype 2 infections in free-ranging mountain gorillas (Gorilla gorilla beringei) of the Bwindi Impenetrable National Park, Uganda. Parasitol. Res. 2001, 87, 368–370. [Google Scholar] [CrossRef]
- Salyer, S.J.; Gillespie, T.R.; Rwego, I.B.; Chapman, C.A.; Goldberg, T.L. Epidemiology and molecular relationships of Cryptosporidium spp. in people, primates, and livestock from Western Uganda. PLoS Negl. Trop. Dis. 2012, 6, e1597. [Google Scholar] [CrossRef]
- Tumwine, J.K.; Kekitiinwa, A.; Nabukeera, N.; Akiyoshi, D.E.; Rich, S.M.; Widmer, G.; Feng, X.; Tzipori, S. Cryptosporidium parvum in children with diarrhoea in Mulago Hospital, Kampala, Uganda. Am. J. Trop. Med. Hyg. 2003, 68, 710–715. [Google Scholar] [CrossRef]
- Kankya, C.; Okello, J.; Baguma, J.N.; Wambi, R.; Ninsiima, L.R.; Tubihemukama, M.; Kulabako, C.T.; Munyeme, M.; Hartmack, S.; Okello, W. Prevalence and factors associated with cryptosporidiosis among livestock and dogs in Kasese District, Uganda: A cross-sectional study. medRxiv 2023. [Google Scholar] [CrossRef]
- Available online: www.kasese.go.ugKaseseDistrict (accessed on 30 January 2023).
- NECOC. Kasese Hazard, Risk and Vulnerability Profiles—Western Uganda Region. Available online: opm.go.ug (accessed on 30 January 2023).
- Kuzehkanan, B.A.; Rezaeian, M.; Zeraati, H.; Mohebali, M.; Meamar, A.R.; Babaei, Z.; Kashi, L.; Heydarnezhadi, M.; Rezaie, S. A sensitive and specific PCR based method for identification of Cryptosporidium Sp. using new primrs from 18S Ribosomal RNA. Iran J. Parasitol. 2011, 6, 1–7. [Google Scholar]
- Casemore, D.P.; Armstrong, M.; Sands, R.L. Laboratory diagnosis of cryptosporidiosis. J. Clin. Path. 1985, 38, 1337–1341. [Google Scholar] [CrossRef] [PubMed]
- Hall, T. BioEdit: An important software for molecular biology. GERF Bull. Biosci. 2011, 2, 60–61. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol, Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Nizeyi, J.B.; Sebunya, D.; Dasilva, A.J.; Cranfield, M.R.; Pieniazek, N.J.; Graczyk, T.K. Cryptosporidiosis in people sharing habitats with free-ranging mountain gorillas (Gorilla gorilla beringei), Uganda. Am. J. Trop. Med. Hyg. 2002, 66, 442–444. [Google Scholar] [CrossRef]
- Casemore, D.P.; Wright, S.E.; Coop, R.L. Cryptosporidiosis–Human and animal epidemiology. In Cryptosporidium and Cryptosporidiosis; Fayer, R., Ed.; CRC Press: Boca Raton, FL, USA, 1997; pp. 65–92. [Google Scholar]
- Karanis, P.; Kourenti, C.; Smith, H. Waterborne transmission of protozoan parasites: A worldwide review of outbreaks and lessons learnt. J. Water Health 2007, 5, 1–38. [Google Scholar] [CrossRef]
- Alexander, K.A.; Carzolio, M.; Goodin, D.; Vance, E. Climate change is likely to worsen the public health threat of diarrheal disease in Botswana. Int. J. Environ. Res. Public Health 2013, 10, 1202–1230. [Google Scholar] [CrossRef]
- Parsons, M.B.; Travis, D.; Lonsdorf, E.V.; Lipende, I.; Roellig, D.M.A.; Kamenya, S.; Zhang, H.; Xiao, L.; Gillespie, T.R. Epidemiology and molecular characterization of Cryptosporidium spp. in humans, wild primates, and domesticated animals in the greater Gombe ecosystem, Tanzania. PLoS Negl. Trop. Dis. 2015, 10, e0003529. [Google Scholar] [CrossRef]
- Xiao, L.H.; Feng, Y.Y. Zoonotic cryptosporidiosis. FEMS Immunol. Med. Microbiol. 2008, 52, 309–323. [Google Scholar] [CrossRef]
- Shoukry, N.M.; Dawoud, H.A.; Haridy, F.M. Studies on zoonotic cryptosporidiosis parvum in Ismailia Governorate, Egypt. J Egypt Soc. Parasitol. 2009, 39, 479–488. [Google Scholar]
- Samn, K.; Samn, A.; Abou El-Nour, M. A survey of Giardia and Cryptosporidium spp. in Rural and Urban community in North Delta, Egypt. N. Y. Sci. J. 2012, 5, 49–54. [Google Scholar]
- Cama, V.A.; Bern, C.; Roberts, J.; Cabrera, L.; Sterling, C.R.; Ortega, Y.; Gilman, R.H.; Xiao, L. Cryptosporidium species and subtypes and clinical manifestations in children, Peru. Emerg. Infect. Dis. 2008, 14, 1567–1574. [Google Scholar] [CrossRef] [PubMed]
- Bern, C.; Hernandez, B.; Lopez, M.B.; Arrowood, M.J.; De Merida, A.M.; Klein, R.E. The contrasting epidemiology of Cyclospora and Cryptosporidium among outpatients in Guatemala. Am. J. Trop. Med. Hyg. 2000, 63, 231–235. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Maboud, A.I.; Rossignol, J.F.; Mostafa, M.S. Cryptosporidiosis in Benha, study of some modalities in diagnosis and treatment. J. Egypt. Soc. Parasitol. 2000, 30, 717–725. [Google Scholar]
- Sinyangwe, N.N.; Siwila, J.; Muma, J.B.; Chola, M.; Michelo, C. Factors associated with Cryptosporidium infection among adult HIV positive population in contact with livestock in Namwala District, Zambia. Front. Public Health 2020, 8, 74. [Google Scholar] [CrossRef]
- Yongsi, H.B.N. Suffering for water, suffering from water: Access to drinking water and associated health risks in Cameroon. J. Health Popul. Nutr. 2010, 28, 424–435. [Google Scholar] [CrossRef]
- Ayalew, D.; Boelee, E.; Endeshaw, T.; Petros, B. Cryptosporidium and Giardia infection and drinking water sources among children in Lege Dini, Ethiopia. Trop. Med. Int. Health 2008, 13, 472–475. [Google Scholar] [CrossRef]
- Raccurt, C.P.; Brasseur, P.; Verdier, R.I.; Li, X.; Eyma, E.; Stockman, C.P.; Agnamey, P.; Guyot, K.; Totet, A.; Liautaud, B.; et al. Human cryptosporidiosis and Cryptosporidium spp. in Haiti. Trop. Med. Int. Health 2006, 11, 929–934. [Google Scholar] [CrossRef]
- Akiyoshi, D.E.; Tumwine, J.K.; Bakeera-Kitaka, S.; Tzipori, S. Subtype analysis of Cryptosporidium isolates from children in Uganda. J. Parasitol. 2006, 92, 1097–1100. [Google Scholar] [CrossRef]
- Graczyk, T.K.; Fayer, R.; Cranfield, M.R. Zoonotic potential of cross-transmission of Cryptosporidium parvum: Implications for waterborne cryptosporidiosis. Parasitol. Today 1997, 13, 348–351. [Google Scholar] [CrossRef]
- Fayer, R.; Morgan, U.; Upton, S.J. Epidemiology of Cryptosporidium: Transmission, detection and identification. Int. J. Parasitol. 2000, 30, 1305–1322. [Google Scholar] [CrossRef] [PubMed]
- Helmy, Y.A.; Krücken, J.; Nöckler, K.; von Samson-Himmelstjerna, G.; Zessin, K.-H. Molecular epidemiology of Cryptosporidium in livestock animals and humans in the Ismailia province of Egypt. Vet. Parasitol. 2013, 193, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Ukwah, B.N.; Ezeonu, I.M.; Ezeonu, C.T.; Roellig, D.; Xiao, L. Cryptosporidium species and subtypes in diarrheal children and HIV-infected persons in Ebonyi and Nsukka, Nigeria. J. Infect. Dev. Ctries 2017, 11, 173–179. [Google Scholar] [CrossRef] [PubMed]
- Adamu, H.; Petros, B.; Hailu, A.; Petry, F. Molecular characterization of Cryptosporidium isolates from humans in Ethiopia. Acta Trop. 2010, 115, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Samra, N.A.; Jori, F.; Samie, A.; Thompson, P. The prevalence of Cryptosporidium spp. oocyst in wild mammals in the Kruger National Park, South Africa. Vet. Parasitol. 2011, 175, 155–159. [Google Scholar] [CrossRef]
- Ramakers, C.; Ruijter, J.M.; Deprez, R.H.L.; Moorman, A.F.M. Assumption-free analysis of quantitative real-time polymerase chain reaction (PCR) data. Neurosci. Lett. 2003, 339, 62–66. [Google Scholar] [CrossRef]
- Khurana, S.; Sharma, P.; Sharma, A.; Malla, N. Evaluation of Ziehl-Neelsen staining, auramine phenol staining, antigen detection enzyme linked immunosorbent assay and polymerase chain reaction, for the diagnosis of intestinal cryptosporidiosis. Trop. Parasitol. 2012, 2, 20–23. [Google Scholar] [CrossRef]
- Lendner, M.; Etzold, M.; Daugschies, A. Kryptosporidiose--ein Update [Cryptosporidiosis--an update]. Berl. Munch Tierarztl. Wochenschr. 2011, 124, 473–484. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Microscopy | PCR | ||||
|---|---|---|---|---|---|
| Characteristic | Number Analyzed | Number Positive | % Positivity (95% CI) | Number Positive | % Positivity (95% CI) |
| Location | |||||
| Kasenyi HCII | 79 | 22 | 27.8 (18.3–39.1) | 19 | 24.1 (15.1–35) |
| Karusandara HCIII | 71 | 18 | 25.4 (15.8–37.1) | 13 | 18.3 (10.1–29.3) |
| Katwe-Kabatooro HCII | 49 | 8 | 16.3 (7.3–29.7) | 12 | 24.5 (13.3–38.9) |
| Katunguru HCII | 45 | 7 | 15.6 (6.5–29.5) | 11 | 24.4 (12.9–39.5) |
| Hamukungu HCII | 38 | 5 | 13.2 (4.4–28.1) | 3 | 7.9 (1.7–21.4) |
| Age group (Years) | |||||
| <5 | 87 | 27 | 31 (21.5–41.9) | 30 | 34.5 (24.6–45.4) |
| 5–18 | 89 | 28 | 31.5 (22–42.2) | 28 | 31.5 (22–42.2) |
| 19–35 | 70 | 17 | 24.3 (14.8–36) | 19 | 27.1 (17.2–39.1) |
| 36–65 | 28 | 3 | 10.7 (2.3–28.2) | 1 | 3.6 (0.1–18.3) |
| >65 | 6 | 0 | 0 (0–45.9) | 0 | 0 (0–45.9) |
| Not indicated | 2 | 0 | 0 (0–84.2) | 0 | 0 (0–84.2) |
| Gender | |||||
| Female | 151 | 81 | 53.6 (45.4–61.8) | 86 | 57 (48.7–65) |
| Male | 129 | 59 | 45.7 (36.9–54.7) | 55 | 42.6 (34–51.6) |
| Not indicated | 2 | 0 | 0 (0–84.2) | 0 | 0 (0–84.2) |
| Stool consistency | |||||
| Watery | 104 | 38 | 36.5 (27.3–46.6) | 38 | 36.5 (27.3–46.6) |
| Watery with rice pellets | 35 | 4 | 11.4 (3.2–26.7) | 4 | 11.4 (3.2–26.7) |
| Solid hard | 77 | 22 | 28.6 (18.8–40) | 22 | 28.6 (18.8–40) |
| Mucoid | 39 | 5 | 12.8 (4.3–27.4) | 5 | 12.8 (4.3–27.4) |
| Solid soft | 24 | 2 | 8.3 (1–27) | 3 | 12.5 (2.7–32.4) |
| Not indicated | 3 | 0 | 0 (0–70.8) | 0 | 0 (0–70.8) |
| Sampling Site | Wildlife Species Sampled (No. Positive/ No. Sampled) | Overall | |||||
|---|---|---|---|---|---|---|---|
| Buffalo | Elephant | Hippopotamus | Uganda Kob | Warthogs | Water Buck | % Positivity | |
| Camp site 2 | 0 | 100% (5/5) | 0 | 0 | 0 | 0 | 100% (5/5) |
| Channel track | 0 | 100% (5/5) | 100% (4/4) | 0 | 0 | 0 | 100% 9/9) |
| Janet track | 78% (7/9) | 0 | 0 | 0 | 0 | 0 | 78% (7/9) |
| Kaguta track | 0 | 0 | 0 | 0 | 100% (1/1) | 45% (5/11) | 50% (6/12) |
| Kasenyi Koblek | 0 | 0 | 0 | 100% (1/1) | 0 | 0 | 100% (1/1) |
| Katunguru gate | 0 | 0 | 0% (0/1) | 0 | 0% (0/1) | 100% (2/2) | 50% (2/4) |
| Katwe fishing village | 0 | 50% (1/2) | 0% (0/1) | 0 | 0 | 50% (1/2) | 40% (2/5) |
| Lake Katwe near institution | 0 | 67% (2/3) | 0 | 0 | 0 | 0 | 67% (2/3) |
| Mweya Jet | 67% (2/3) | 0 | 0 | 0 | 75% (3/4) | 0 | 71% (5/7) |
| Mweya peninsula | 0 | 0 | 0 | 100% (1/1) | 91% (20/22) | 0 | 91% (21/23) |
| Nyamunuka stretch | 0 | 67% (2/3) | 0 | 0 | 0 | 0 | 67% (2/3) |
| Research Koblek | 93% (14/15) | 0 | 0 | 0 | 0 | 0 | 93% (14/15) |
| Researchers track | 0 | 0 | 0 | 50% (2/4) | 0 | 0 | 50% (2/4) |
| Overall | 85% (23/27) | 83% (15/18) | 67% (4/6) | 67% (4/6) | 86% (24/28) | 53% (8/15) | 78% (78/100) |
| Number of Samples with the Following Results Microscopy | ||||
|---|---|---|---|---|
| Positive | Negative | Total | ||
| PCR | Positive | 61 | 52 | 113 |
| Negative | 87 | 82 | 169 | |
| Total | 148 | 134 | 282 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mugasa, C.M.; Mirembe, B.B.; Ochwo, S.; Nkamwesiga, J.; Ndekezi, C.; Tusabe, T.; Musoba, A.; Kankya, C. Molecular Detection of Cryptosporidium Species in Wildlife and Humans at the Wildlife-Human Interface around Queen Elizabeth National Park, Uganda. Parasitologia 2023, 3, 181-193. https://doi.org/10.3390/parasitologia3020019
Mugasa CM, Mirembe BB, Ochwo S, Nkamwesiga J, Ndekezi C, Tusabe T, Musoba A, Kankya C. Molecular Detection of Cryptosporidium Species in Wildlife and Humans at the Wildlife-Human Interface around Queen Elizabeth National Park, Uganda. Parasitologia. 2023; 3(2):181-193. https://doi.org/10.3390/parasitologia3020019
Chicago/Turabian StyleMugasa, Claire Mack, Bernadette Basuta Mirembe, Sylvester Ochwo, Joseph Nkamwesiga, Christian Ndekezi, Tobias Tusabe, Abubakar Musoba, and Clovice Kankya. 2023. "Molecular Detection of Cryptosporidium Species in Wildlife and Humans at the Wildlife-Human Interface around Queen Elizabeth National Park, Uganda" Parasitologia 3, no. 2: 181-193. https://doi.org/10.3390/parasitologia3020019