
A New Giant Petrel (Macronectes, Aves: Procellariidae) from the Pliocene of Taranaki, New Zealand †

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
3. Systematics
- Order Procellariiformes Fürbringer, 1888
- Family Procellariidae Leach, 1820
- Genus Macronectes Richmond, 1905
- Macronectes tinae sp. nov.
- ZooBank reg. nr.: urn:lsid:zoobank.org:act:EB59C374-6AE6-4FB8-8D22-0F2747CD6F3A
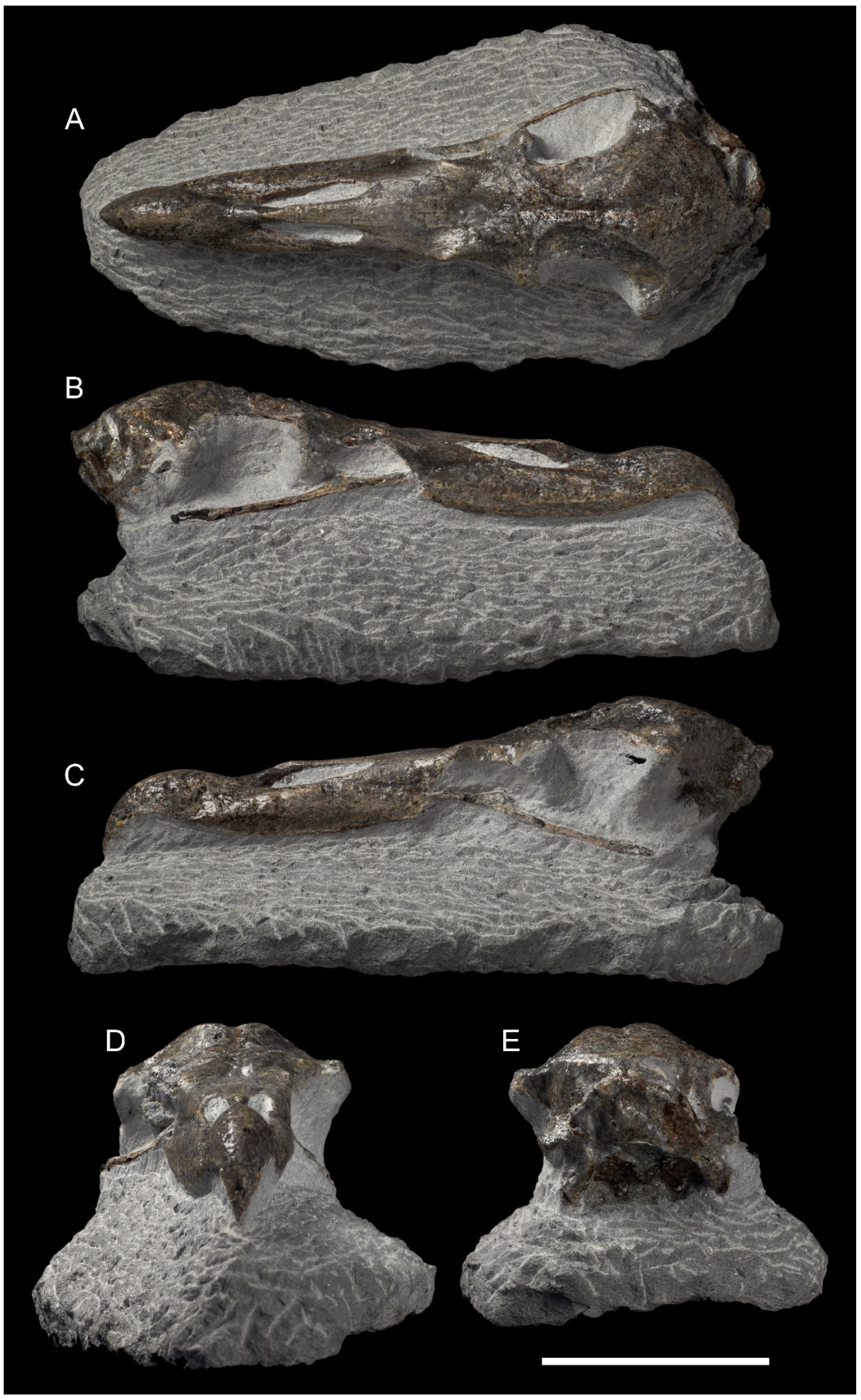
- Holotype: NMNZ S.048502 (col. Alastair Johnson, 2017): largely complete skull (Figure 1).
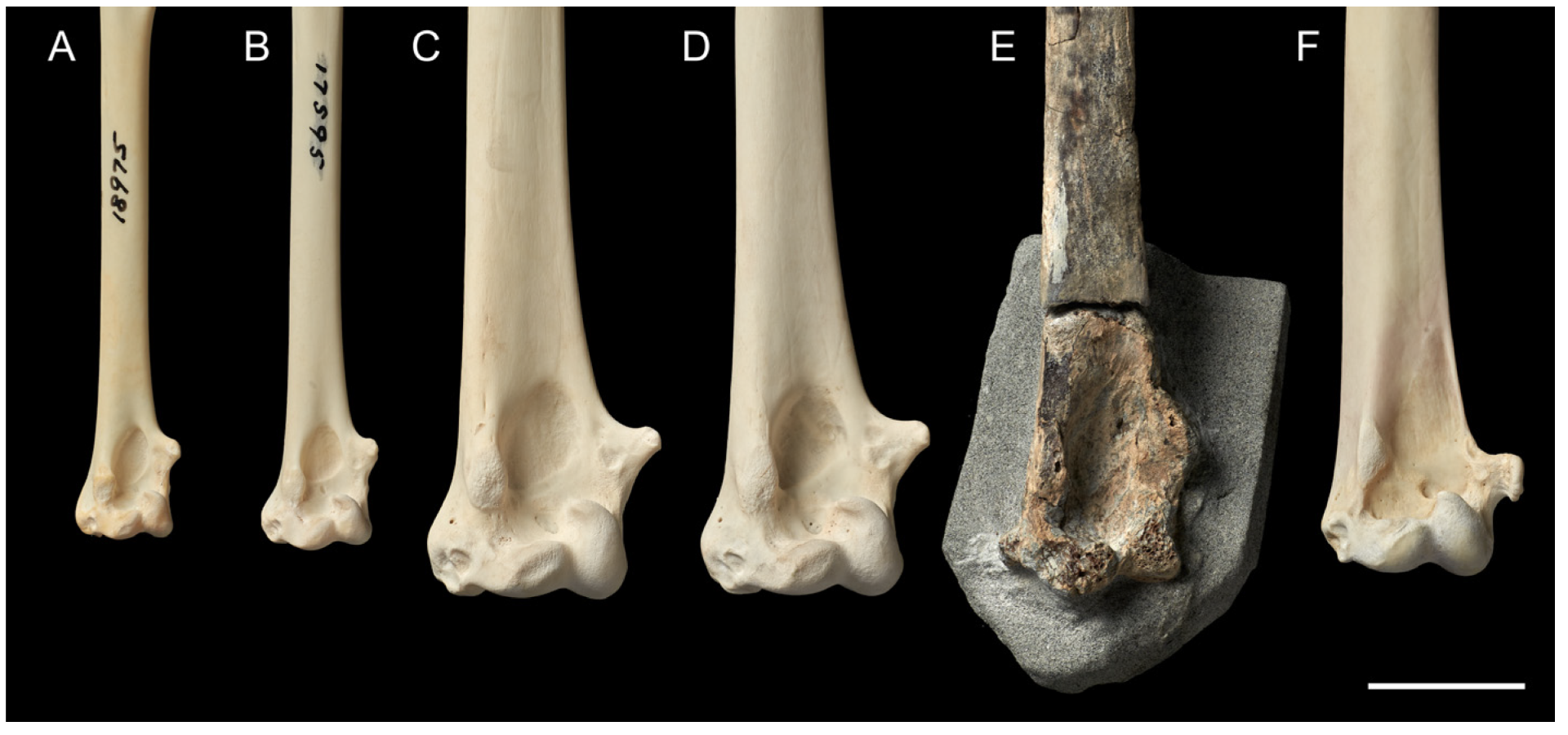
- Paratype: NMNZ S.048870 (col. Alastair Johnson, 2019): left humerus, fragmentary, only the shaft and distal end remaining (Figure 3A). The shaft is to be broken near where the crista deltopectoralis would terminate.
- Type locality and stratum: New Zealand, North Island, southern Taranaki, Hāwera. Tangahoe Formation. The holotype and paratype were surface collected as beach boulders and do not have an exact Fossil Record Electronic Database number, but see Q21/f0002 for nearby location.
- Age: Late Pliocene, Piacenzian (“Waipipian Stage” in the New Zealand scale): constrained to 3.36–3.06 Ma, based on the oxygen isotope stage and magnetic polarity data [10].
- Etymology: The specific epithet honours Tina King, the late partner of fossil collector Alastair Johnson. This giant petrel skull was her favourite fossil, hence the homage.
- Measurements: Skull: FTD = 7.2 mm; MFF = 6.9 mm; MIF = 20.5 mm; MUW = 12.6 mm; NL = 23.1 mm; NW = 4.5 mm; PFNW = 22.2 mm; PoW = 48.8 mm (estimate); PrW = 34.9 mm (estimate); TL = 148 mm. Humerus: total (preserved) length = 166 mm; DW = 26.8 mm; SbW = 14.7 mm; SbD = 6.8 mm.
- Diagnosis: Skull: overall smaller size; proportionately longer apertura nasi ossea; apparently shallower os supraocciptale. Humerus: shaft proportionately craniocaudally less deep, with more delicate appearance than congeners; medial portion of epicondylus ventralis more prominent; fossa medialis brachialis proportionately larger, elongated, and nearly fusiform.
- Differential diagnosis: There is little intraspecific and interspecific variation in the skull and humerus morphology between M. giganteus and M. halli ([16,18]; this study: Table S1), barring sexual dimorphism (males are larger; [16,20,21]) and the slightly smaller average size of M. halli (the size range of the two species completely overlap; [1,7]). That is to be expected from taxa with little genetic distinction [6]. As such, morphological comparisons can be made between M. tinae sp. nov. and both living Macronectes spp. simultaneously.
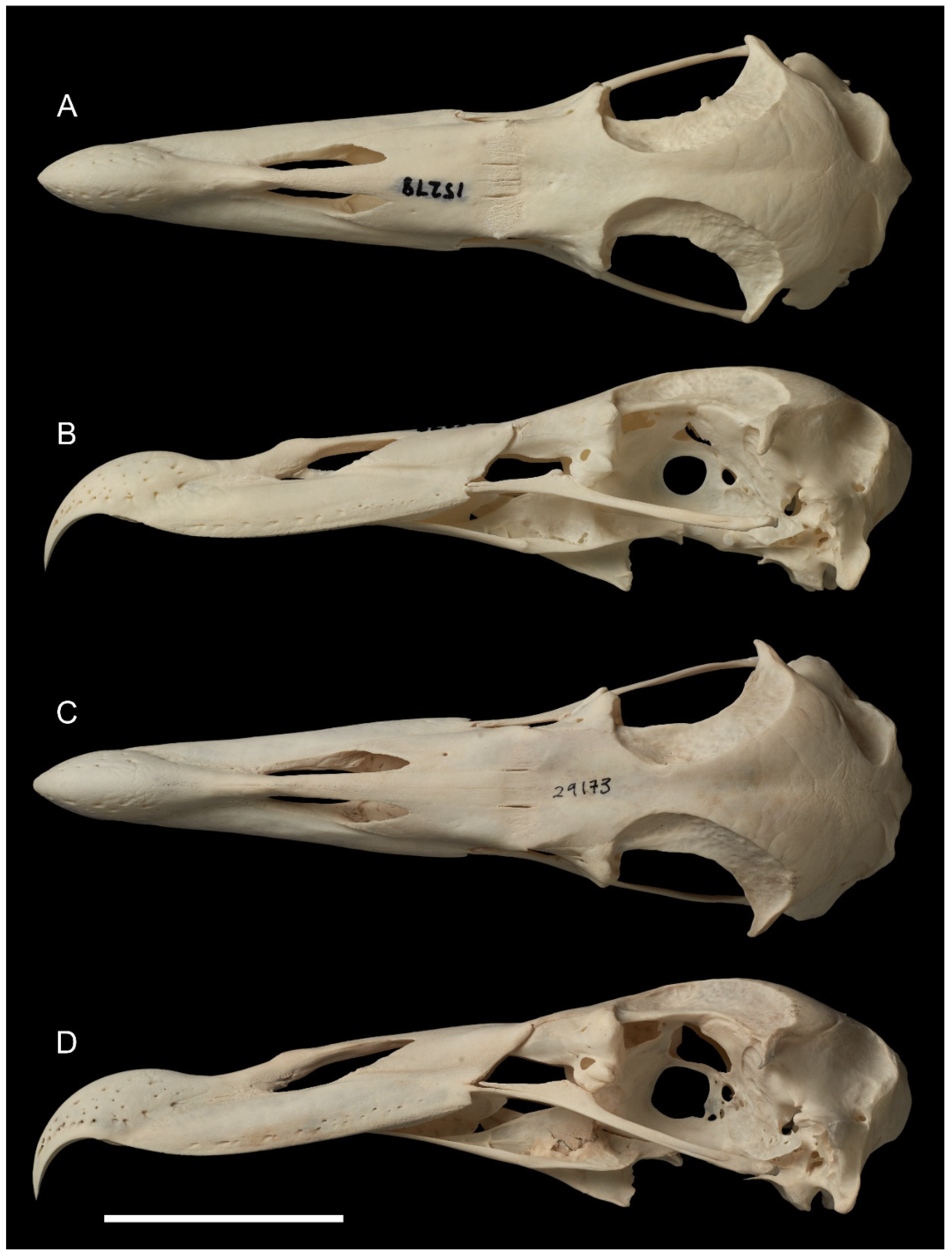
- Skull: The skull of M. tinae sp. nov. (Figure 1) is smaller than all Macronectes spp. in the NMNZ collection (Table S1) and can be instantly diagnosed by its size. Barring the size difference, almost all other structures are the same as in living Macronectes spp. (Figure 2), with two exceptions: the fossa temporalium and the os supraocciptale.
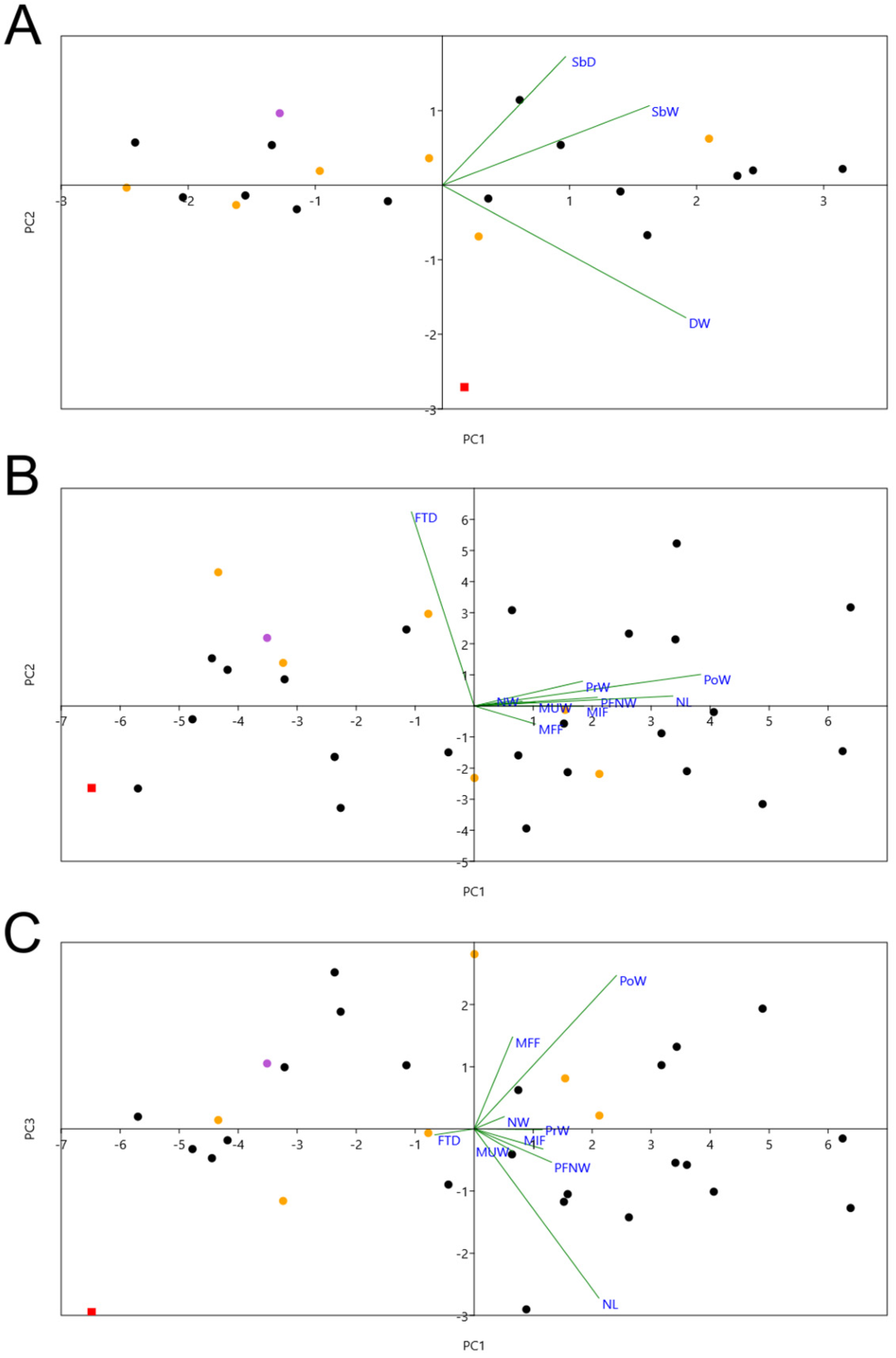
- Humerus: The humerus of M. tinae sp. nov. is about as big as the smallest Macronectes spp. in the NMNZ collection (e.g., NMNZ OR.015606; Table S1). The distal end of the fossil is more delicate than that of living Macronectes spp., with its shaft being proportionately less deep (Figure 3; Table S1: larger [SbW/SbD] measure in M. tinae sp. nov.). According to the PCA, PC1 explains 81% of variance and PC2 17% (Table S3). Larger PC1 values mean greater W, SbW, and SbD, while larger PC2 values mean greater SbW and SbD (thicker shaft), but lower W (smaller distal end). By plotting PC1 × PC2 (Figure 5A), it is clear that M. tinae sp. nov. is separated from the two recent species (which largely overlap) due to its different proportions, as explained above.
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Carboneras, C. Family Procellariidae (petrels and shearwaters). In Handbook of the Birds of the World; del Hoyo, J., Elliot, A., Sargatal, J., Eds.; Lynx: Barcelona, Spain, 1992; pp. 216–239. [Google Scholar]
- Checklist Committee OSNZ. Checklist of the Birds of New Zealand, 5th ed.; Ornithological Society of New Zealand Occasional Publication No. 1; Ornithological Society of New Zealand: Wellington, New Zealand, 2022; 335p. [Google Scholar]
- Bourne, W.P.; Warham, J. Geographical variation in the giant petrels of the genus Macronectes. Ardea 1966, 54, 45–67. [Google Scholar]
- Voisin, J.-F. Les pétrels géants Macronectes giganteus et M. halli de l’île de la Possession (archipel Crozet). Oiseau 1968, 38, 95–122. [Google Scholar]
- Rheindt, F.E.; Austin, J. Major analytical and conceptual shortcomings in a recent taxonomic revision of the Procellariiformes—A reply to Penhallurick and Wink (2004). Emu-Austral Ornithol. 2005, 105, 181–186. [Google Scholar] [CrossRef]
- Penhallurick, J.; Wink, M. Analysis of the taxonomy and nomenclature of the Procellariiformes based on complete nucleotide sequences of the mitochondrial cytochrome b gene. Emu 2004, 104, 125–147. [Google Scholar] [CrossRef]
- Marchant, S.; Higgins, P.J. Handbook of Australian, New Zealand and Antarctic Birds. Vol. 1: Ratites to Ducks, Part A—Ratites to Petrels. Oxford University Press: Melbourne, Australia, 1990; 736p. [Google Scholar]
- Avery, G.A.; Klein, R.G. Review of fossil phocid and otariid seals from the southern and western coasts of South Africa. Trans. R. Soc. S. Afr. 2011, 66, 14–24. [Google Scholar] [CrossRef]
- Tennyson, A.J.D. Holocene bird bones found at the subantarctic Auckland Islands. Notornis 2020, 67, 269–294. [Google Scholar]
- Naish, T.R.; Wehland, F.; Wilson, G.S.; Browne, G.H.; Cook, R.A.; Morgans, H.E.G.; Rosenberg, M.; King, P.R.; Smale, D.; Nelson, C.S.; et al. An integrated sequence stratigraphic, palaeoenvironmental, and chronostratigraphic analysis of the Tangahoe Formation, southern Taranaki coast, with implications for mid-Pliocene (c. 3.4–3.0 Ma) glacio-eustatic sea-level changes. J. R. Soc. N. Z. 2005, 35, 151–196. [Google Scholar] [CrossRef] [Green Version]
- McKee, J. Geology and vertebrate palaentology of the Tangahoe Formation, South Taranaki Coast, New Zealand. Geol. Soc. N. Z. Misc. Publ. B 1994, 80, 63–91. [Google Scholar]
- Thomas, D.B.; Tennyson, A.J.D.; Scofield, R.P.; Heath, T.A.; Pett, W.; Ksepka, D.T. Ancient crested penguin constrains timing of recruitment into seabird hotspot. Proc. R. Soc. B Boil. Sci. 2020, 287, 20201497. [Google Scholar] [CrossRef]
- Gilbert, B.M.; Martin, L.D.; Savage, H.G. Avian Osteology; Modern Printing: Laramie, WY, USA, 1981; 252p. [Google Scholar]
- Baumel, J.J.; Witmer, L.M. Osteologia. In Handbook of Avian Anatomy: Nomina Anatomica Avium; Baumel, J.J., King, A.S., Breazile, J.E., Evans, H.E., Vanden Berge, J.C., Eds.; Nuttall Ornithological Club: Cambridge, UK, 1993; pp. 45–132. [Google Scholar]
- Livezey, B.C.; Zusi, R.L. Higher-order phylogeny of modern birds (Theropoda, Aves: Neornithes) based on comparative anatomy. II. Analysis and discussion. Zool. J. Linn. Soc. 2007, 149, 1–95. [Google Scholar] [CrossRef] [Green Version]
- Piro, A.; Hospitaleche, C.A. Skull morphology and ontogenetic variation of the Southern Giant Petrel Macronectes giganteus (Aves: Procellariiformes). Polar Biol. 2019, 42, 27–45. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. Past: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 4. [Google Scholar]
- Mazzochi, M.S.; Carlos, C.J. Skull morphology of four Antarctic fulmarine petrels (Aves: Procellariiformes): Insights into their feeding biology. Polar Biol. 2022, 45, 191–201. [Google Scholar] [CrossRef]
- Mayr, G.; Smith, T. Phylogenetic affinities and taxonomy of the Oligocene Diomedeoididae, and the basal divergences amongst extant procellariiform bird. Zool. J. Linn. Soc. 2012, 166, 854–875. [Google Scholar] [CrossRef]
- Conroy, J.W.H. Ecological aspects of the biology of the Giant Petrel, Macronectes giganteus (Gmelin), in the maritime Antarctic. Br. Antarct. Surv. Sci. Rep. 1972, 75, 1–74. [Google Scholar]
- Carlos, C.J.; Voisin, J.F. Identifying giant petrels, Macronectes giganteus and M. halli, in the field and in the hand. Seabird 2008, 21, 1–15. [Google Scholar]
- Hendy, A.J.; Kamp, P.J.; Vonk, A.J. Late Miocene turnover of molluscan faunas, New Zealand: Taxonomic and ecological reassessment of diversity changes at multiple spatial and temporal scales. Palaeogeogr. Palaeoclim. Palaeoecol. 2009, 280, 275–290. [Google Scholar] [CrossRef]
- Salomon, M.; Voisin, J.-F. Ecogeographical variation in the Southern Giant Petrel (Macronectes giganteus). Can. J. Zool. 2010, 88, 195–203. [Google Scholar] [CrossRef]
- Schodde, R.; Tennyson, A.J.; Groth, J.G.; Lai, J.; Scofield, R.; Steinheimer, F.D. Settling the name Diomedea exulans Linnaeus, 1758 for the Wandering Albatross by neotypification. Zootaxa 2017, 4236, 135–148. [Google Scholar] [CrossRef]
- Prince, P.A.; Morgan, R.A. Diet and feeding ecology of Procellariiformes. In Seabird: Feeding Ecology and Role in Marine Ecosystems; Croxall, J.P., Ed.; Cambridge University Press: Cambridge, UK, 1987; pp. 135–171. [Google Scholar]
- Hunter, S. The role of giant petrels in the Southern Ocean ecosystem. In Antarctic Nutrient Cycles and Food Webs; Siegfried, W.R., Condy, P.R., Laws, R.M., Eds.; Springer: Berlin/Heidelberg, Germany, 1985; pp. 534–542. [Google Scholar]
- Reisinger, R.R.; Carpenter-Kling, T.; Connan, M.; Cherel, Y.; Pistorius, P.A. Foraging behaviour and habitat-use drives niche segregation in sibling seabird species. R. Soc. Open Sci. 2020, 7, 200649. [Google Scholar] [CrossRef]
- Obst, B.S. Densities of Antarctic Seabirds at Sea and the Presence of the Krill Euphausia superba. Ornithology 1985, 102, 540–549. [Google Scholar] [CrossRef]
- Harper, P.C. Feeding behaviour and other notes on 20 species of Procellariiformes at sea. Notornis 1987, 34, 169–192. [Google Scholar]
- Rule, J.P.; Adams, J.W.; Marx, F.G.; Evans, A.R.; Tennyson, A.J.D.; Scofield, R.P.; Fitzgerald, E.M.G. First monk seal from the Southern Hemisphere rewrites the evolutionary history of true seals. Proc. R. Soc. B Boil. Sci. 2020, 287, 20202318. [Google Scholar] [CrossRef] [PubMed]
- Thomas, D.B.; Ksepka, D.T.; Holvast, E.J.; Tennyson, A.J.D.; Scofield, P. Re-evaluating New Zealand’s endemic Pliocene penguin genus. N. Z. J. Geol. Geophys. 2020, 63, 324–330. [Google Scholar] [CrossRef]
- Tennyson, A.J.D.; Mannering, A.A. A new species of Pliocene shearwater (Aves: Procellariidae) from New Zealand. Tuhinga 2018, 29, 1–19. [Google Scholar]
- Mayr, G.; Tennyson, A.J.D. A small, narrow-beaked albatross from the Pliocene of New Zealand demonstrates a higher past diversity in the feeding ecology of the Diomedeidae. Ibis 2020, 162, 723–734. [Google Scholar] [CrossRef]
- Tennyson, A.J.D.; Tomotani, B.M. A new fossil species of Procellaria (Aves: Procellariiformes) from the Pliocene of New Zealand. Papéis Avulsos De Zool. 2021, 61, e20216116. [Google Scholar] [CrossRef]
- McKee, J.W.A. A pseudodontorn (Pelecaniformes: Pelagornithidae) from the middle Pliocene of Hawera, Taranaki, New Zealand. N. Z. J. Zool. 1985, 12, 181–184. [Google Scholar] [CrossRef]
- Imber, M.J. Origins, phylogeny and taxonomy of the gadfly petrels Pterodroma spp. Ibis 1985, 127, 197–229. [Google Scholar] [CrossRef]
- Howard, H. Tertiary birds from Laguna Hill, Orange County, California. Los Angeles Cty. Mus. Contrib. Sci. 1968, 142, 1–21. [Google Scholar] [CrossRef]
- Howard, H. Additional avian records from the Miocene of Kern County, California with the description of a new species of fulmar. Bull. South. Calif. Acad. Sci. 1984, 83, 84–89. [Google Scholar]
- Prothero, D.R.; Sanchez, F.; Denke, L.I. Magnetic stratigraphy of the early to middle Miocene Olcese Sand and Round Mountain Silt, Kern County, California. New Mex. Mus. Nat. Hist. Sci. Bull. 2008, 44, 357–363. [Google Scholar]
- Techow, N.; O’Ryan, C.; Phillips, R.; Gales, R.; Marin, M.; Patterson-Fraser, D.; Quintana, F.; Ritz, M.; Thompson, D.; Wanless, R.; et al. Speciation and phylogeography of giant petrels Macronectes. Mol. Phylogenetics Evol. 2010, 54, 472–487. [Google Scholar] [CrossRef] [PubMed]
- Olson, S.L. An early Pliocene marine avifauna from Duinefontein, Cape Province, South Africa. Ann. South Afr. Mus. 1985, 95, 147–164. [Google Scholar]
- Olson, S.L. Early Pliocene Procellariiformes (Aves) from Langebaanweg, south-western Cape Province, South Africa. Ann. South Afr. Mus. 1985, 95, 123–145. [Google Scholar]
- Dickinson, E.C.; Remsen, J.V. The Howard & Moore Complete Checklist of the Birds of the World, 4th ed.; A & C Black: London, UK, 2013; 752p. [Google Scholar]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tennyson, A.J.D.; Salvador, R.B. A New Giant Petrel (Macronectes, Aves: Procellariidae) from the Pliocene of Taranaki, New Zealand. Taxonomy 2023, 3, 57-67. https://doi.org/10.3390/taxonomy3010006
Tennyson AJD, Salvador RB. A New Giant Petrel (Macronectes, Aves: Procellariidae) from the Pliocene of Taranaki, New Zealand. Taxonomy. 2023; 3(1):57-67. https://doi.org/10.3390/taxonomy3010006
Chicago/Turabian StyleTennyson, Alan J. D., and Rodrigo B. Salvador. 2023. "A New Giant Petrel (Macronectes, Aves: Procellariidae) from the Pliocene of Taranaki, New Zealand" Taxonomy 3, no. 1: 57-67. https://doi.org/10.3390/taxonomy3010006