
Metabolites Potentially Determine the High Antioxidant Properties of Limosilactobacillus fermentum U-21
 ,
,  and
and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains
2.2. Method, Conditions, and Media for Strain Propagation
2.3. Phase Separation
2.4. Extracellular Vesicles Preparation and Physicochemical Analysis
2.5. Extraction of Metabolites
2.6. Derivatization of Metabolites
2.7. GC × GC-MS Analysis
2.8. Data analysis
3. Results
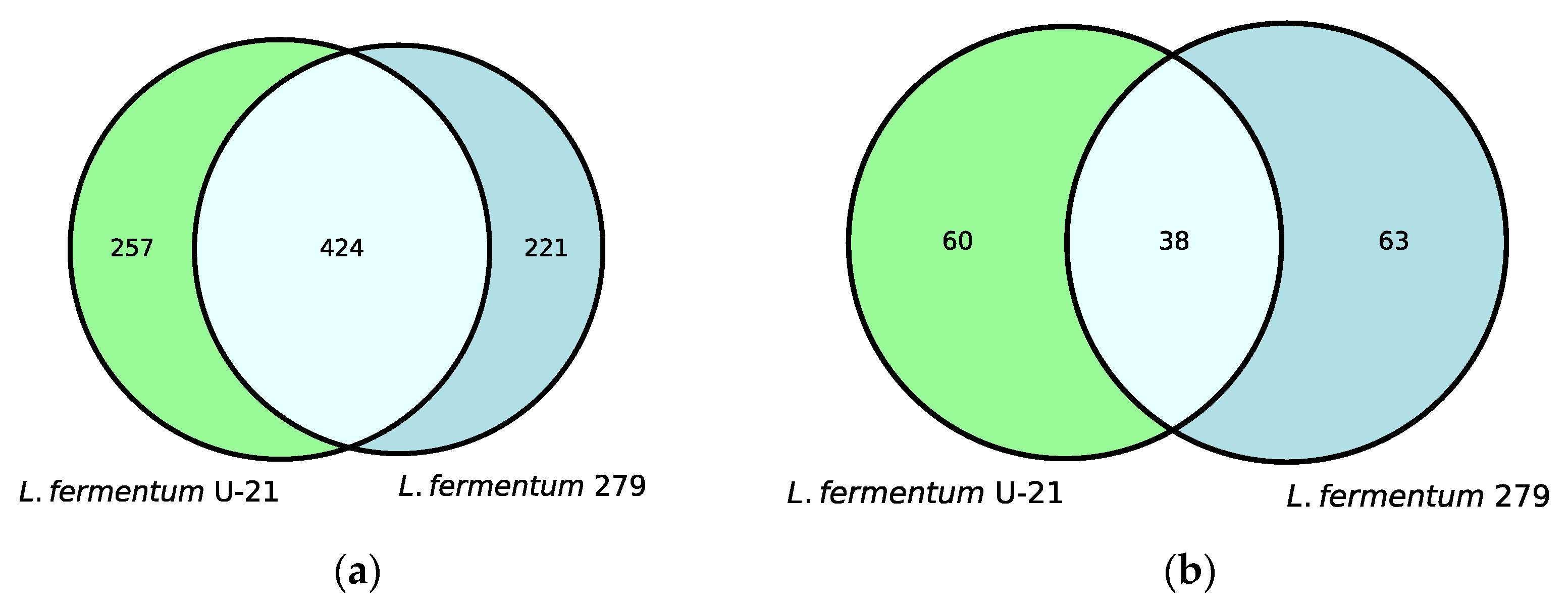
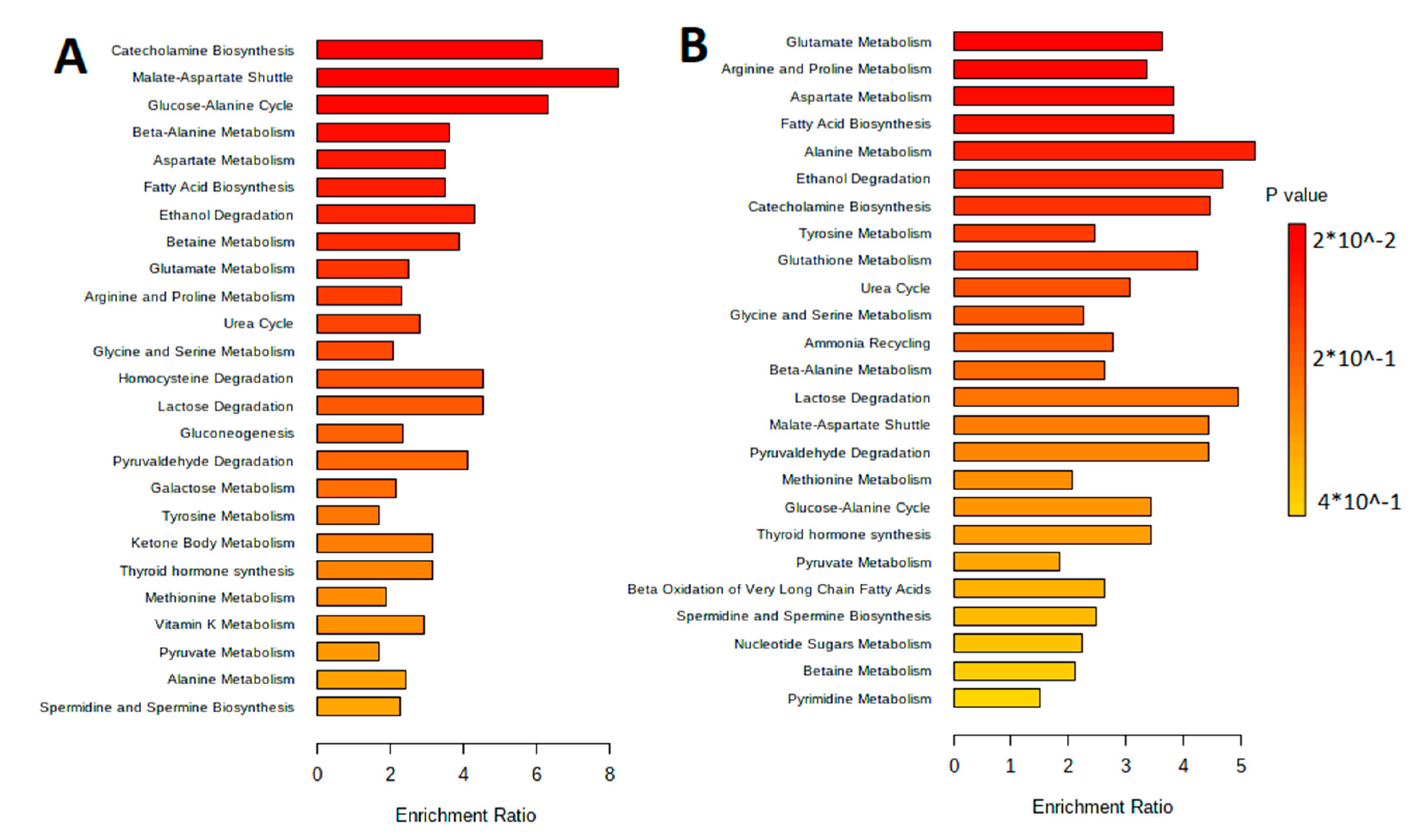
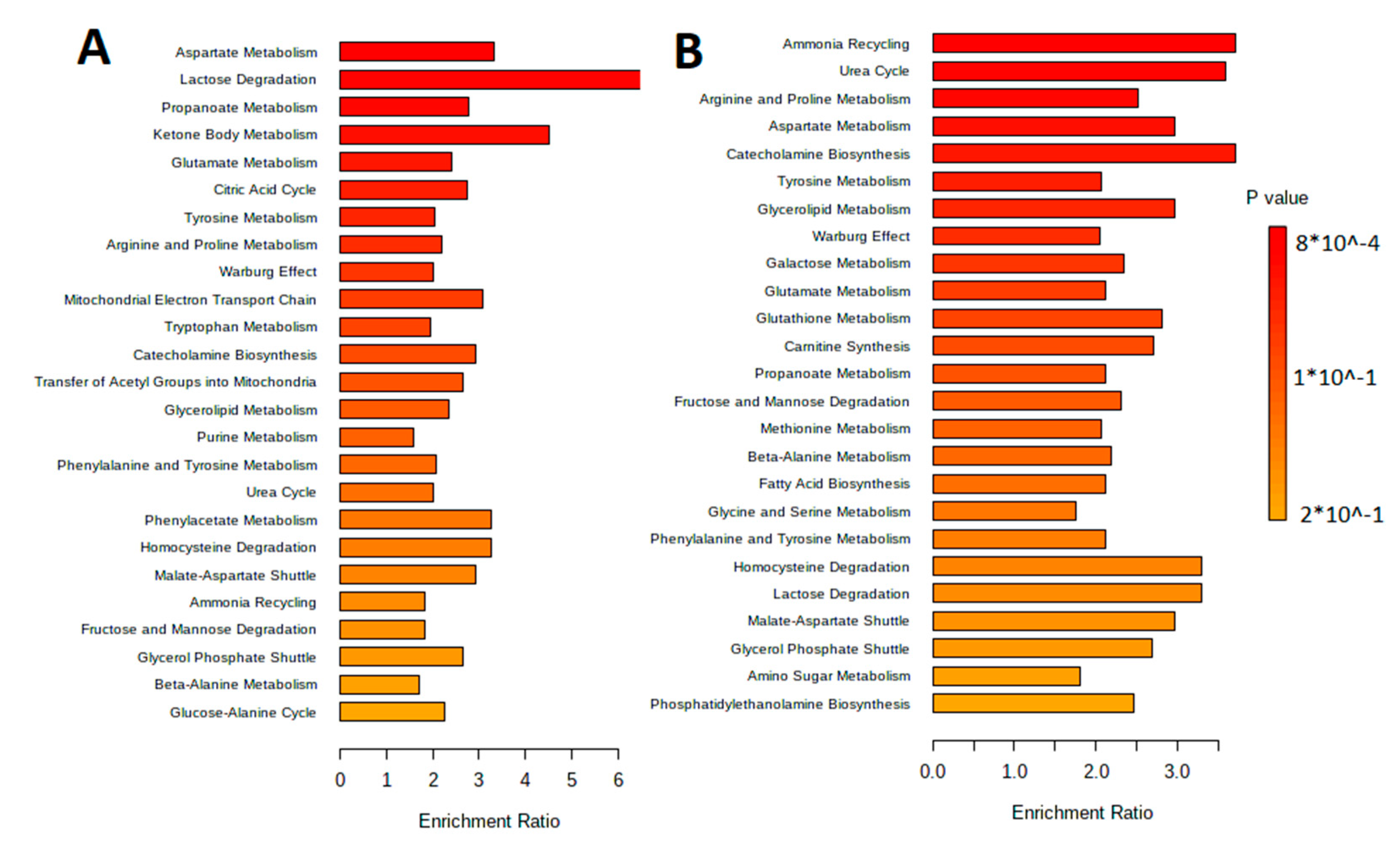
3.1. Characteristic of the Metabolite Profile of the Culture Supernatant
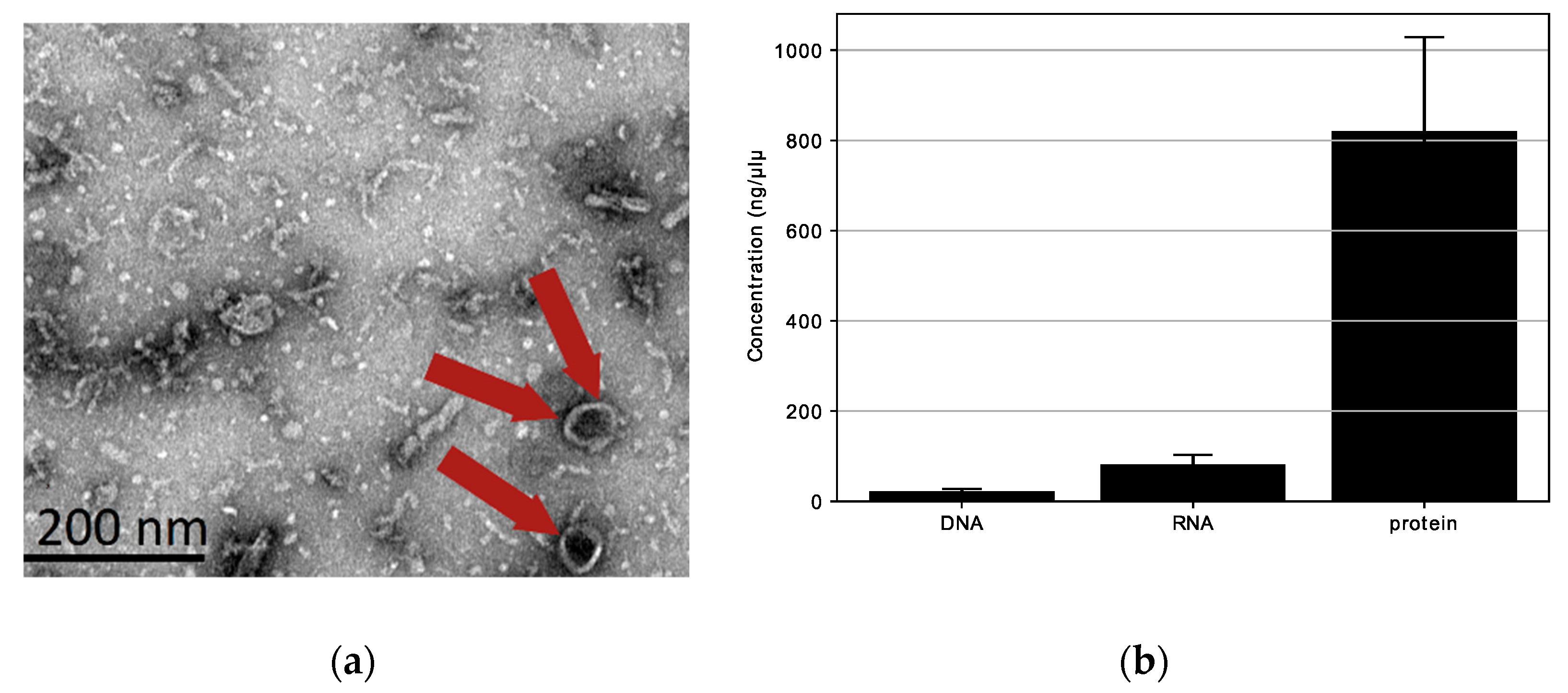
3.2. Physicochemical Characterization of L. fermentum U-21 Extracellular Vesicles
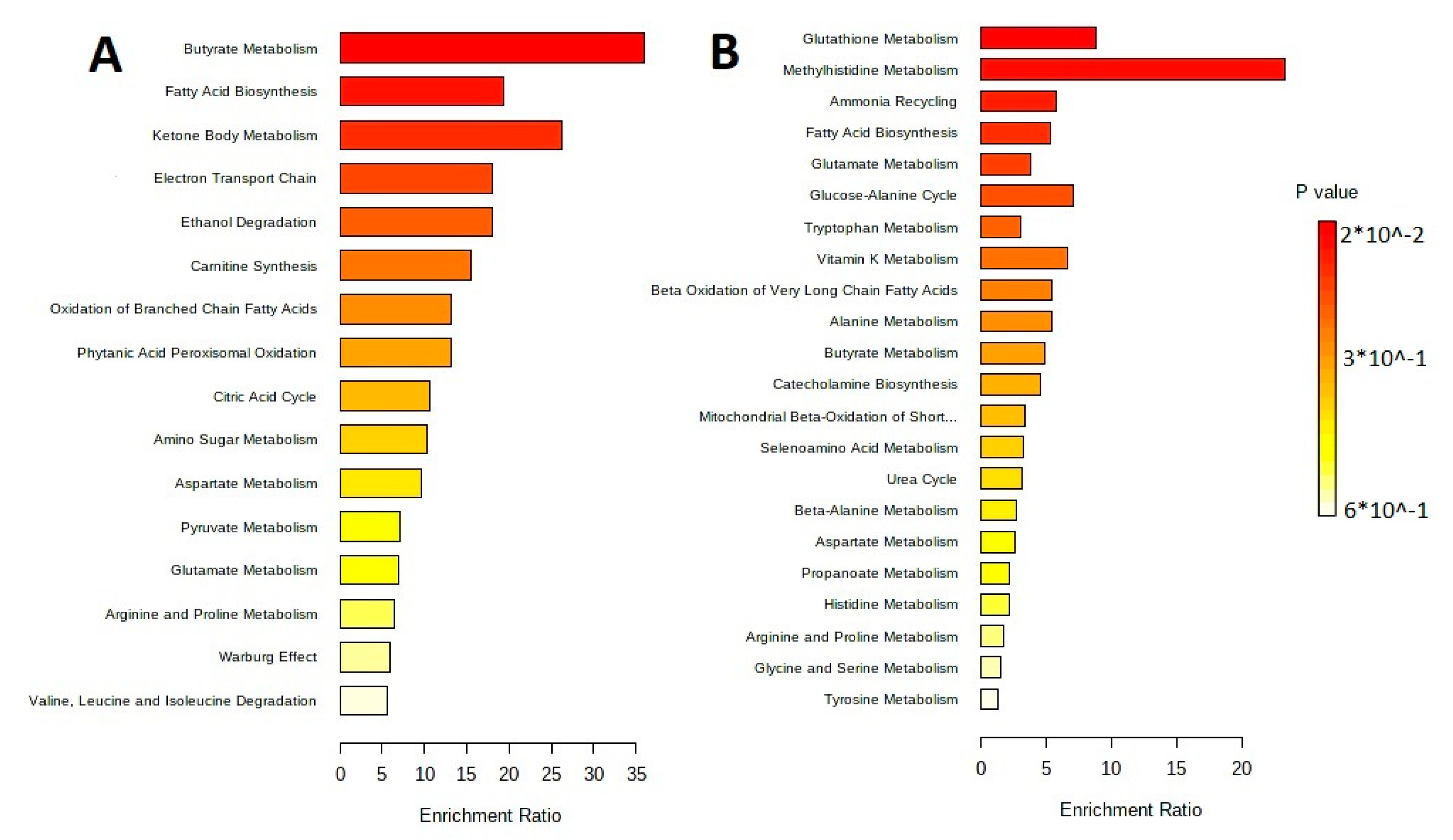
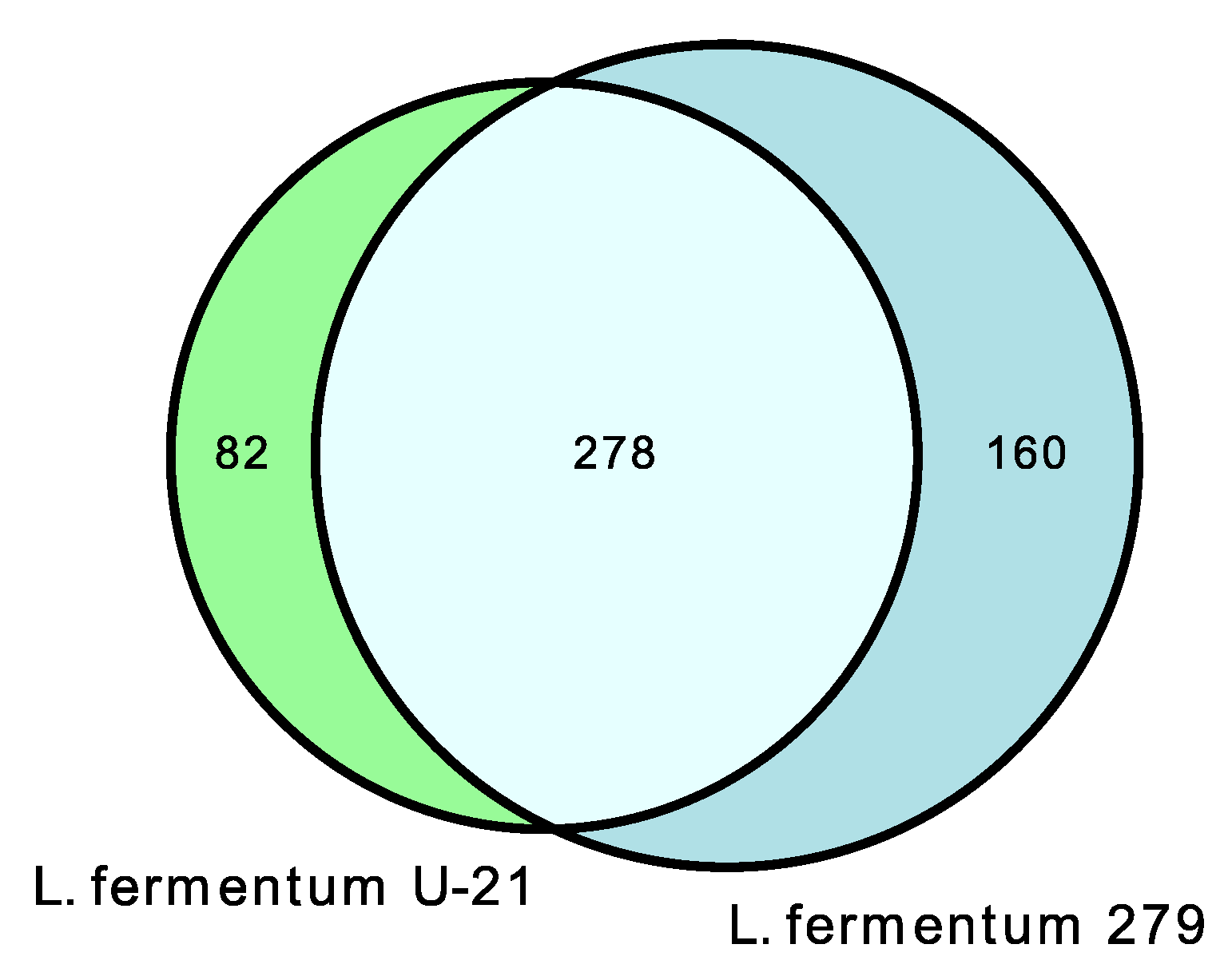
3.3. Isolation and Characteristic of the Metabolite Profile of the Extracellular Vesicles
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dettmer, K.; Aronov, P.A.; Hammock, B.D. Mass spectrometry-based metabolomics. Mass Spectrom. Rev. 2007, 26, 51–78. [Google Scholar] [CrossRef] [PubMed]
- Cambeiro-Pérez, N.; Hidalgo-Cantabrana, C.; Moro-García, M.A.; Alonso-Arias, R.; Simal-Gándara, J.; Sánchez, B.; Martínez-Carballo, E. A metabolomics approach reveals immunomodulatory effects of proteinaceous molecules derived from gut bacteria over human peripheral blood mononuclear cells. Front. Microbiol. 2018, 9, 2701. [Google Scholar] [CrossRef] [PubMed]
- Schell, M.A.; Karmirantzou, M.; Snel, B.; Vilanova, D.; Berger, B.; Pessi, G.; Zwahlen, M.C.; Desiere, F.; Bork, P.; Delley, M.; et al. The genome sequence of Bifidobacterium longum reflects its adaptation to the human gastrointestinal tract. Proc. Natl. Acad. Sci. USA 2002, 99, 14422–14427. [Google Scholar] [CrossRef]
- Duar, R.M.; Lin, X.B.; Zheng, J.; Martino, M.E.; Grenier, T.; Pérez-Muñoz, M.E.; Leulier, F.; Gänzle, M.; Walter, J. Lifestyles in transition: Evolution and natural history of the genus Lactobacillus. FEMS Microbiol. Rev. 2017, 41, S27–S48. [Google Scholar] [CrossRef] [PubMed]
- Dempsey, E.; Corr, S.C. Lactobacillus spp. for gastrointestinal health: Current and future perspectives. Front. Immunol. 2022, 13, 840245. [Google Scholar] [CrossRef]
- Mishra, V.; Shah, C.; Mokashe, N.; Chavan, R.; Yadav, H.; Prajapati, J. Probiotics as potential antioxidants: A systematic review. J. Agric. Food Chem. 2015, 63, 3615–3626. [Google Scholar] [CrossRef]
- Nowak, A.; Paliwoda, A.; Błasiak, J. Anti-proliferative, pro-apoptotic and anti-oxidative activity of Lactobacillus and Bifidobacterium strains: A review of mechanisms and therapeutic perspectives. Crit. Rev. Food Sci. Nutr. 2019, 59, 3456–3467. [Google Scholar] [CrossRef]
- Danilenko, V.N.; Stavrovskaya, A.V.; Voronkov, D.N.; Gushchina, A.S.; Marsova, M.V.; Yamshchikova, N.G.; Ol’shansky, S.; Ivanov, M.; Illarioshkin, S.N. The use of a pharmabiotic based on the Lactobacillus fermentum U-21 strain to modulate the neurodegenerative process in an experimental model of Parkinson disease. Ann. Clin. Exp. Neurol. 2020, 14, 62–69. [Google Scholar]
- Poluektova, E.; Yunes, R.; Danilenko, V. The putative antidepressant mechanisms of probiotic bacteria: Relevant genes and proteins. Nutrients 2021, 13, 1591. [Google Scholar] [CrossRef]
- Zhou, Y.; Gong, W.; Xu, C.; Zhu, Z.; Peng, Y.; Xie, C. Probiotic assessment and antioxidant characterization of Lactobacillus plantarum GXL94 isolated from fermented chili. Front. Microbiol. 2022, 13, 997940. [Google Scholar] [CrossRef]
- Thorakkattu, P.; Khanashyam, A.C.; Shah, K.; Babu, K.S.; Mundanat, A.S.; Deliephan, A.; Deokar, G.S.; Santivarangkna, C.; Nirmal, N.P. Postbiotics: Current trends in food and Pharmaceutical industry. Foods 2022, 11, 3094. [Google Scholar] [CrossRef] [PubMed]
- Averina, O.V.; Poluektova, E.U.; Marsova, M.V.; Danilenko, V.N. Biomarkers and utility of the antioxidant potential of probiotic Lactobacilli and Bifidobacteria as representatives of the human gut microbiota. Biomedicines 2021, 9, 1340. [Google Scholar] [CrossRef] [PubMed]
- Marsova, M.; Abilev, S.; Poluektova, E.; Danilenko, V. A bioluminescent test system reveals valuable antioxidant properties of lactobacillus strains from human microbiota. World J. Microbiol. Biotechnol. 2018, 34, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Marsova, M.; Poluektova, E.; Odorskaya, M.; Ambaryan, A.; Revishchin, A.; Pavlova, G.; Danilenko, V. Protective effects of Lactobacillus fermentum U-21 against paraquat-induced oxidative stress in Caenorhabditis elegans and mouse models. World J. Microbiol. Biotechnol. 2020, 36, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Noureen, S.; Riaz, A.; Arshad, M.; Arshad, N. In vitro selection and in vivo confirmation of the antioxidant ability of Lactobacillus brevis MG000874. J. Appl. Microbiol. 2019, 126, 1221–1232. [Google Scholar] [CrossRef]
- Poluektova, E.; Mavletova, D.; Odorskaya, M.; Marsova, M.; Klimina, K.; Koshenko, T.; Yunes, R.; Danilenko, V. Comparative Genomic, Transcriptomic, and Proteomic Analysis of the Limosilactobacillus fermentum U-21 Strain Promising for the Creation of a Pharmabiotic. Russ. J. Genet. 2022, 58, 1079–1090. [Google Scholar] [CrossRef]
- Schramm, F.D.; Schroeder, K.; Jonas, K. Protein aggregation in bacteria. FEMS Microbiol. Rev. 2020, 44, 54–72. [Google Scholar] [CrossRef]
- Alam, A.; Bröms, J.E.; Kumar, R.; Sjöstedt, A. The role of ClpB in bacterial stress responses and virulence. Front. Mol. Biosci. 2021, 8, 668910. [Google Scholar] [CrossRef]
- Vermassen, A.; Leroy, S.; Talon, R.; Provot, C.; Popowska, M.; Desvaux, M. Cell wall hydrolases in bacteria: Insight on the diversity of cell wall amidases, glycosidases and peptidases toward peptidoglycan. Front. Microbiol. 2019, 10, 331. [Google Scholar] [CrossRef]
- Fiehn, O. Metabolomics by gas chromatography–mass spectrometry: Combined targeted and untargeted profiling. Curr. Protoc. Mol. Biol. 2016, 114, 30–34. [Google Scholar] [CrossRef]
- Choi, J.H.; Moon, C.M.; Shin, T.S.; Kim, E.K.; McDowell, A.; Jo, M.K.; Joo, Y.H.; Kim, S.E.; Jung, H.K.; Shim, K.N.; et al. Lactobacillus paracasei-derived extracellular vesicles attenuate the intestinal inflammatory response by augmenting the endoplasmic reticulum stress pathway. Exp. Mol. Med. 2020, 52, 423–437. [Google Scholar] [CrossRef] [PubMed]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. Expert consensus document: The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [PubMed]
- Chaudhari, A.; Dwivedi, M.K. The concept of probiotics, prebiotics, postbiotics, synbiotics, nutribiotics, and pharmabiotics. In Probiotics in the Prevention and Management of Human Diseases; Elsevier: Amsterdam, The Netherlands, 2022; pp. 1–11. [Google Scholar]
- Chamberlain, M.; O’Flaherty, S.; Cobián, N.; Barrangou, R. Metabolomic Analysis of Lactobacillus acidophilus, L. gasseri, L. crispatus, and Lacticaseibacillus rhamnosus Strains in the Presence of Pomegranate Extract. Front. Microbiol. 2022, 13, 863228. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Miao, K.; Niyaphorn, S.; Qu, X. Production of gamma-aminobutyric acid from lactic acid bacteria: A systematic review. Int. J. Mol. Sci. 2020, 21, 995. [Google Scholar] [CrossRef]
- Stagg, C.J.; Bachtiar, V.; Johansen-Berg, H. The role of GABA in human motor learning. Curr. Biol. 2011, 21, 480–484. [Google Scholar] [CrossRef]
- Nuss, P. Anxiety disorders and GABA neurotransmission: A disturbance of modulation. Neuropsychiatr. Dis. Treat. 2015, 11, 165–175. [Google Scholar]
- Gottesmann, C. GABA mechanisms and sleep. Neuroscience 2002, 111, 231–239. [Google Scholar] [CrossRef]
- Wagner, S.; Castel, M.; Gainer, H.; Yarom, Y. GABA in the mammalian suprachiasmatic nucleus and its role in diurnal rhythmicity. Nature 1997, 387, 598–603. [Google Scholar] [CrossRef]
- Rashmi, D.; Zanan, R.; John, S.; Khandagale, K.; Nadaf, A. γ-aminobutyric acid (GABA): Biosynthesis, role, commercial production, and applications. Stud. Nat. Prod. Chem. 2018, 57, 413–452. [Google Scholar]
- Ovallath, S.; Sulthana, B. Levodopa: History and therapeutic applications. Ann. Indian Acad. Neurol. 2017, 20, 185. [Google Scholar]
- Nogal, A.; Valdes, A.M.; Menni, C. The role of short-chain fatty acids in the interplay between gut microbiota and diet in cardio-metabolic health. Gut Microbes 2021, 13, 1897212. [Google Scholar] [CrossRef] [PubMed]
- Glauben, R.; Siegmund, B. Inhibition of histone deacetylases in inflammatory bowel diseases. Mol. Med. 2011, 17, 426–433. [Google Scholar] [CrossRef] [PubMed]
- Lührs, H.; Gerke, T.; Müller, J.; Melcher, R.; Schauber, J.; Boxberger, F.; Scheppach, W.; Menzel, T. Butyrate inhibits NF-κB activation in lamina propria macrophages of patients with ulcerative colitis. Scand. J. Gastroenterol. 2002, 37, 458–466. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhang, B.; Hu, Y.; Zhao, Y. New insights into gut-bacteria-derived indole and its derivatives in intestinal and liver diseases. Front. Pharmacol. 2021, 12, 3531. [Google Scholar] [CrossRef] [PubMed]
- Gould, R.L.; Pazdro, R. Impact of supplementary amino acids, micronutrients, and overall diet on glutathione homeostasis. Nutrients 2019, 11, 1056. [Google Scholar] [CrossRef]
- Kameli, N.; Borman, R.; López-Iglesias, C.; Savelkoul, P.; Stassen, F.R. Characterization of Feces-Derived Bacterial Membrane Vesicles and the Impact of Their Origin on the Inflammatory Response. Front. Cell. Infect. Microbiol. 2021, 11, 667987. [Google Scholar] [CrossRef]
- Díez-Sainz, E.; Milagro, F.I.; Riezu-Boj, J.I.; Lorente-Cebrián, S. Effects of gut microbiota-derived extracellular vesicles on obesity and diabetes and their potential modulation through diet. J. Physiol. Biochem. 2022, 78, 485–499. [Google Scholar] [CrossRef]
- Ahmadi Badi, S.; Bruno, S.P.; Moshiri, A.; Tarashi, S.; Siadat, S.D.; Masotti, A. Small RNAs in Outer Membrane Vesicles and Their Function in Host-Microbe Interactions. Front. Microbiol. 2020, 11, 1209. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | BioSample ID | GenBank Sequence |
|---|---|---|
| Limosilactobacillus fermentum U-21 | SAMN08290293 | GCA_002869825.2 |
| Limosilactobacillus fermentum 279 | SAMN08014151 | GCA_002794275.1 |
| Metabolite | Class | |
|---|---|---|
| Glycine (Gly) Alanine (Ala) Valine (Val) Leucine (Leu) Isoleucine (Ile) Aspartic acid (Asp) Asparagine (Asn) Glutamic acid (Glu) Glutamine (Gln) Serine (Ser) Threonine (Thr) Methionine (Met) Cysteine (Cys) Lysine (Lys) Histidine (His) Proline (Pro) Phenylalanine (Phe) Tyrosine (Tyr) Tryptophan (Trp) 2-Aminocaprylic acid Norleucine Homocysteine Homoserine Norvaline Ornithine N-Methyl-α-aminoisobutyric acid β-Alanine 5-Hydroxytryptophan Pyroglutamic acid Aceturic acid α-Methyl-l-tyrosine N,N-Dimethylglycine N-Acetyl-l-glutamic acid N-Acetylglutamine N-Acetyl-l-phenylalanine N-Acetyltyrosine | Amino acids and derivatives | |
| Carnitine Propanoic acid Cinnamic acid Succinic acid Formic acid Fumaric acid Maleic acid Malonic acid Phthalic acid Propanedioic acid Oxalic acid 2-Aminobutanoic acid 2-Hydroxybutyric acid 3-Hydroxybutyric acid Glycolic acid Lactic Acid Tartaric acid Methylcitric acid Acetic acid Hydracrylic acid Dodecanedioic acid | Organic acid | |
| Maltose Melibiose Mannobiose Cellobiose Lactose Turanose Xylofuranose 2-Amino-2-deoxyhexose Talose Erythrose Xylose Galactose Glucose Mannose Lyxose Ribose Threose Tagatose Fructose Psicose 2-Keto-gluconic acid Gluconic acid Mannonic acid Ribonic acid Arabinonic acid Galacturonic acid Gulonic acid Xylonic acid D-Erythro-Pentitol Mannitol Glycerol meso-Erythritol 1-Deoxypentitol Threitol Pentitol Arabinitol Erythritol Arabitol Arabinofuranose Dihydroxyacetone Levoglucosan Myo-Inositol Trehalose-6-phosphate Mannopyranose Glucose oxime Mannose oxime Arabinopyranose | Saccharides | |
| 2-Hydroxyisocaproic acid 4-Hydroxybutanoic acid Nonanoic acid Butanoic acid | Fatty acids and analogues | |
| Metabolite | Class |
|---|---|
| Alanine (Ala) Asparagine (Asn) Threonine (Thr) Proline (Pro) Tryptophan (Trp) L-Norvaline 1-Aminocyclopentanecarboxylic Pyroglutamic acid 2-Hydroxy-3-methylbutyric acid 2-Methylalanine N-Methyl-l-glutamic acid | Amino acids and derivatives |
| Oxalic acid Phthalic acid Propanedioic acid Succinic acid Ritalinic acid 3-Hydroxyphenylacetic acid Acetic acid 4-Hydroxybenzoic acid | Organic acids |
| Lyxose Fructopyranose Mannitol meso-Erythritol | Saccharides |
| Heptadecanoic acid Undecanoic acid Butanoic acid 2-Hydroxyisocaproic acid Oleic acid | Fatty acids and analogues |
| Metabolite | Class |
|---|---|
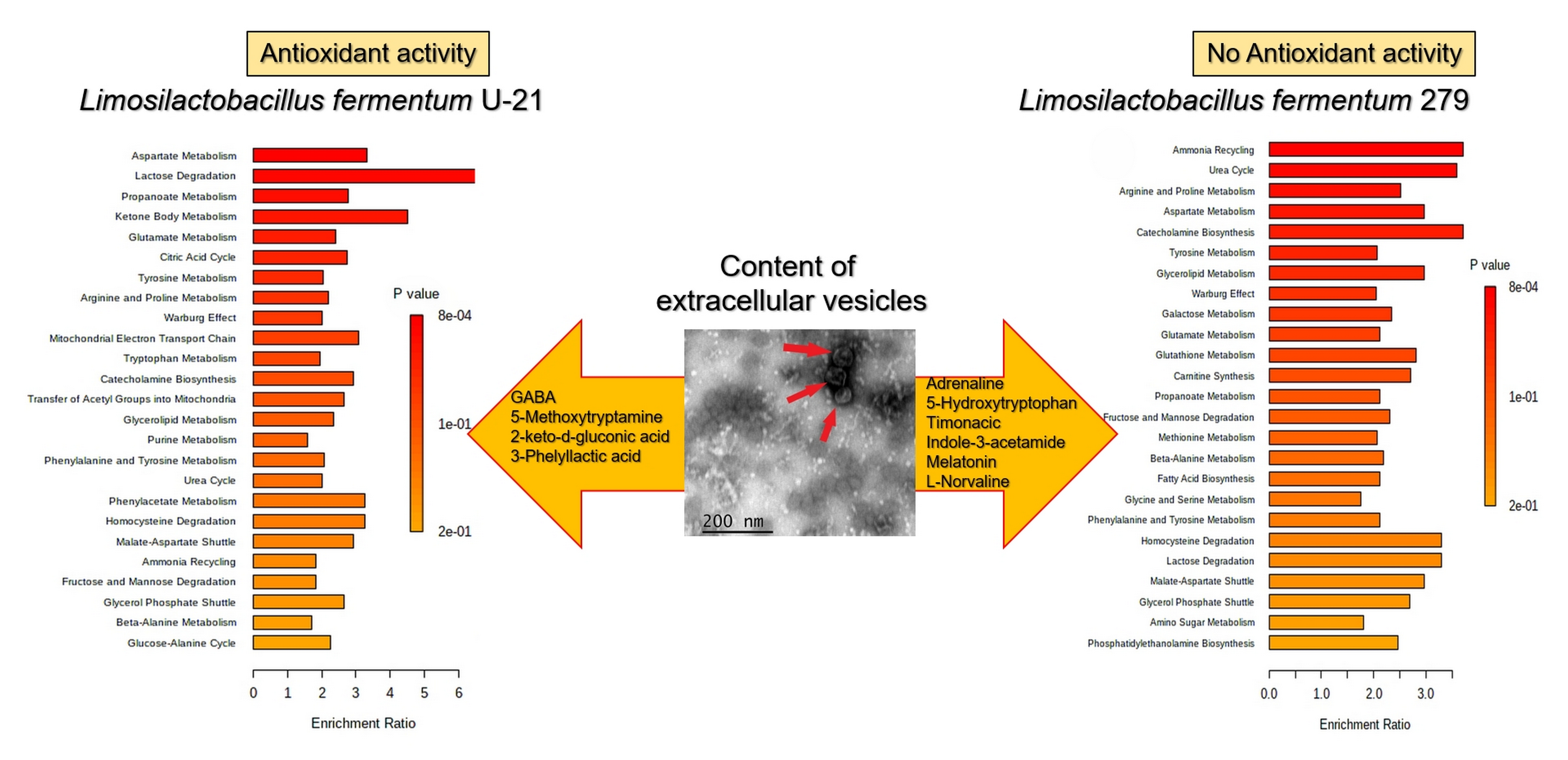
| Dopamine Histamine Noradrenaline Adrenaline GABA l-DOPA Glycine 4-Hydroxybutanoic acid (GHB) | Neurotransmitter |
| Indole-3-acetamide Indole Indolelactate Indole-3-ethanol | Indoles and derivatives |
| Tartaric acid β-Alanine Carnitine | Antioxidant |
| Cinnamic acid L-Norvaline | Anti-inflammatory |
| 5-Hydroxytryptophan | Antidepressant |
| Pyridoxine | Vitamin B6 |
| Niacin Niacinamide | Vitamin B3 |
| Pantothenic acid | Vitamin B5 |
| Acetamide | Antimicrobial |
| Metabolite | Class |
|---|---|
| Adrenaline GABA | Neurotransmitter |
| 5-Hydroxytryptophan | Antidepressant |
| Timonacic 5-Methoxytryptamine (5-MT) | Antioxidant |
| Indole-3-acetamide | Indoles and derivative |
| Melatonin | Hormone |
| 2-keto-D-gluconic acid | Vitamin C precursor |
| L-Norvaline Penicillamine | Anti-inflammatory |
| 3-Phelyllactic acid | Antimicrobial |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grishina, Y.V.; Vatlin, A.A.; Mavletova, D.A.; Odorskaya, M.V.; Senkovenko, A.M.; Ilyasov, R.A.; Danilenko, V.N. Metabolites Potentially Determine the High Antioxidant Properties of Limosilactobacillus fermentum U-21. BioTech 2023, 12, 39. https://doi.org/10.3390/biotech12020039
Grishina YV, Vatlin AA, Mavletova DA, Odorskaya MV, Senkovenko AM, Ilyasov RA, Danilenko VN. Metabolites Potentially Determine the High Antioxidant Properties of Limosilactobacillus fermentum U-21. BioTech. 2023; 12(2):39. https://doi.org/10.3390/biotech12020039
Chicago/Turabian StyleGrishina, Yelena V., Aleksey A. Vatlin, Dilara A. Mavletova, Maya V. Odorskaya, Alexey M. Senkovenko, Rustem A. Ilyasov, and Valeriy N. Danilenko. 2023. "Metabolites Potentially Determine the High Antioxidant Properties of Limosilactobacillus fermentum U-21" BioTech 12, no. 2: 39. https://doi.org/10.3390/biotech12020039