
Protein Delivery to Insect Epithelial Cells In Vivo: Potential Application to Functional Molecular Analysis of Proteins in Butterfly Wing Development


Abstract
:1. Introduction
2. Materials and Methods
2.1. Butterfly Rearing
2.2. Experimental Operations
2.3. Fluorescent Proteins, Antibodies, and Delivery Reagents
2.4. Fluorescent Dyes and Confocal Imaging
2.5. Endosome versus Cytosol Estimates
3. Results
3.1. Protein Delivery without Delivery Reagent
3.1.1. Sandwich Method
3.1.2. Injection Method
3.2. Protein Delivery with L17E Delivery Reagent
3.2.1. Sandwich Method
3.2.2. Injection Method
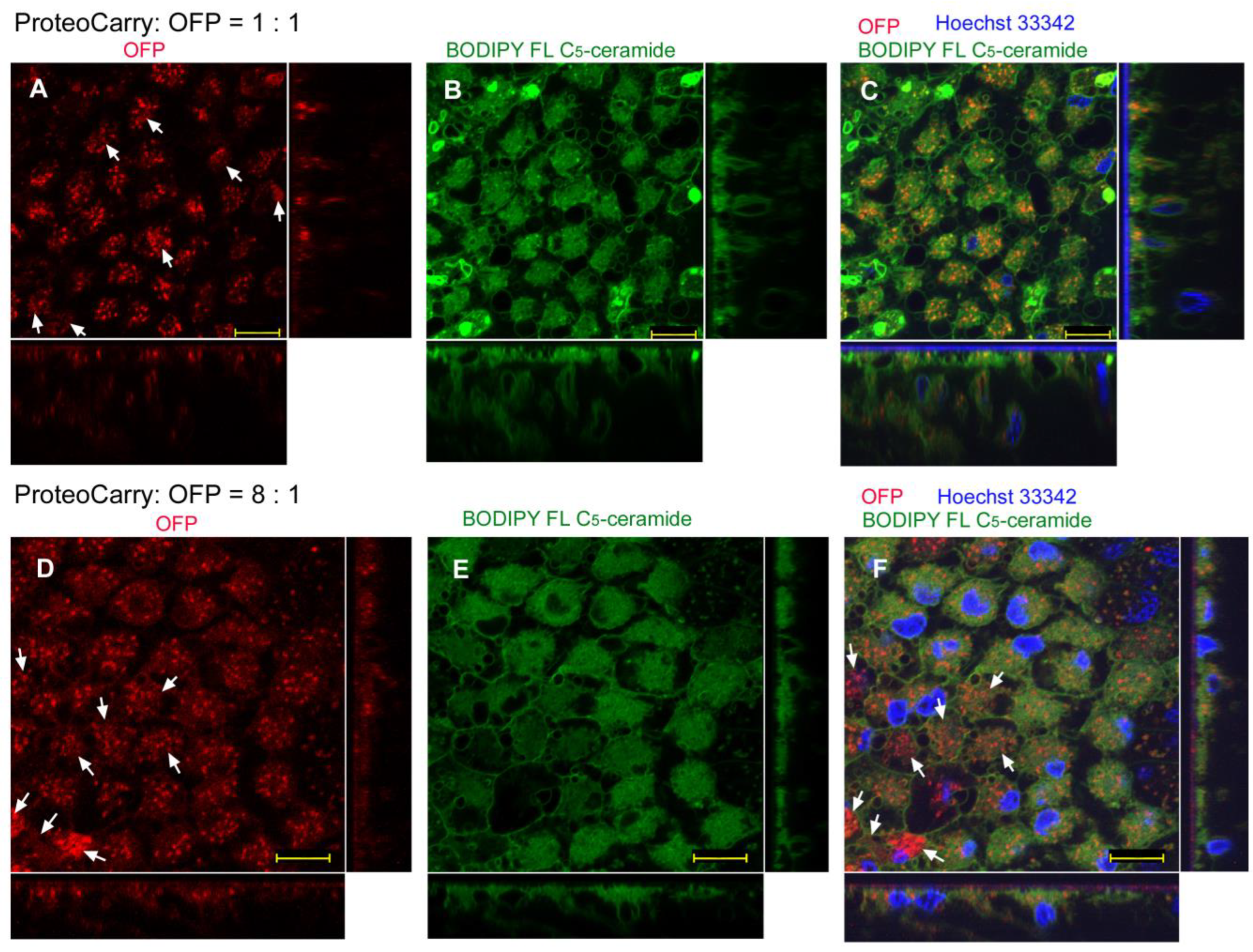
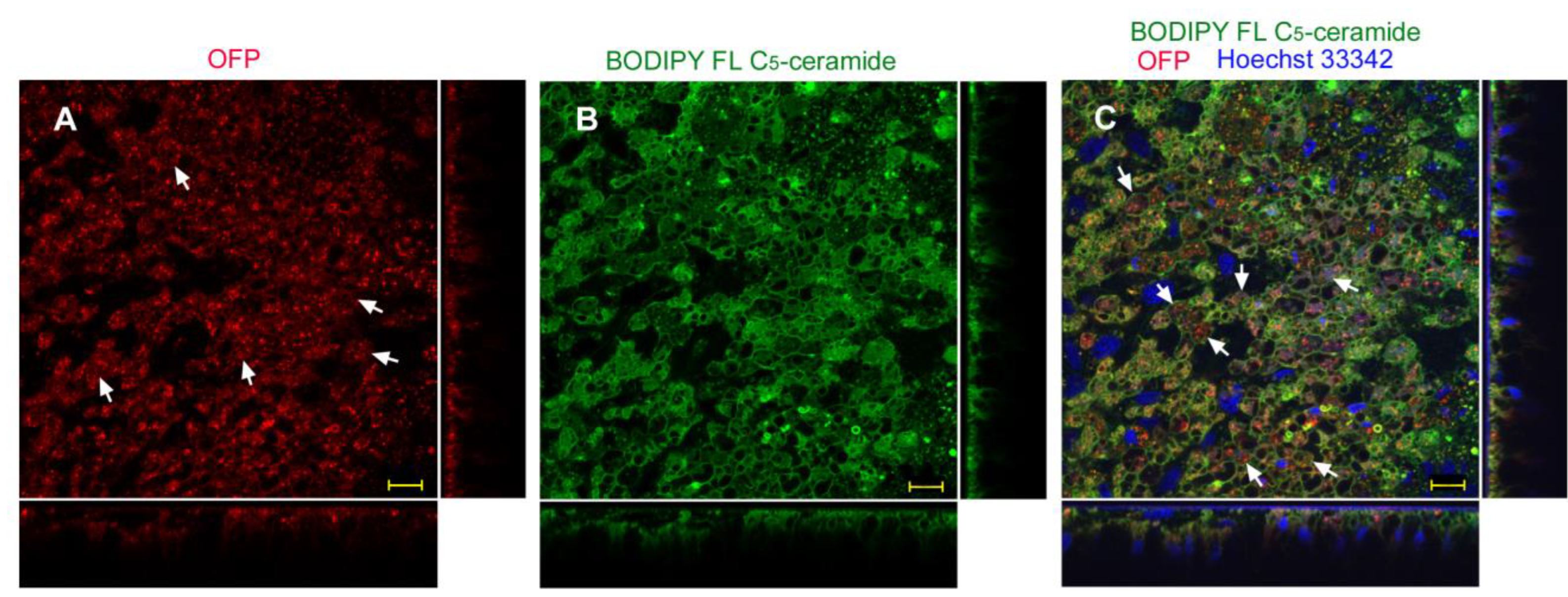
3.3. Protein Delivery with ProteoCarry Delivery Reagent
3.3.1. Sandwich Method
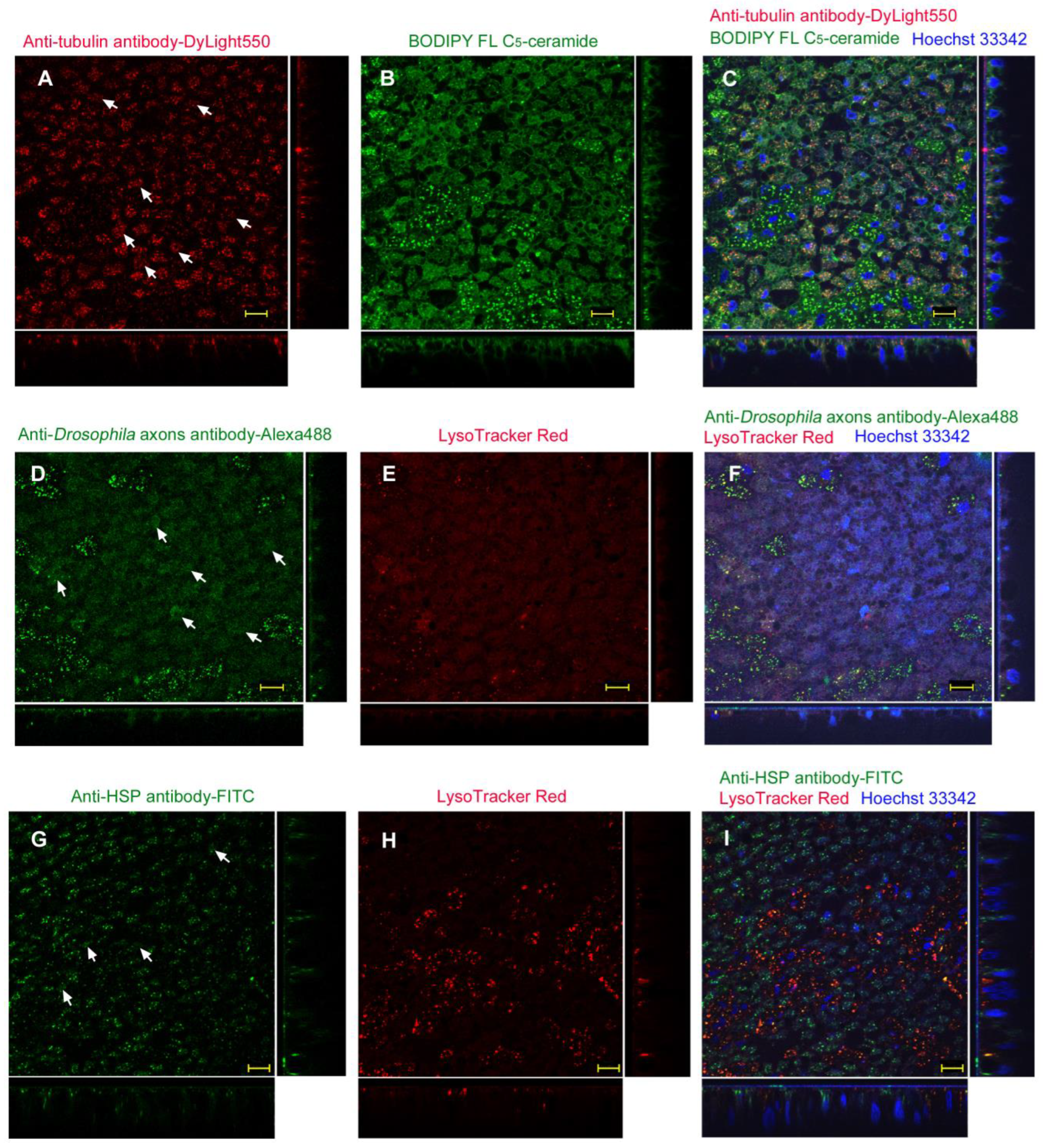
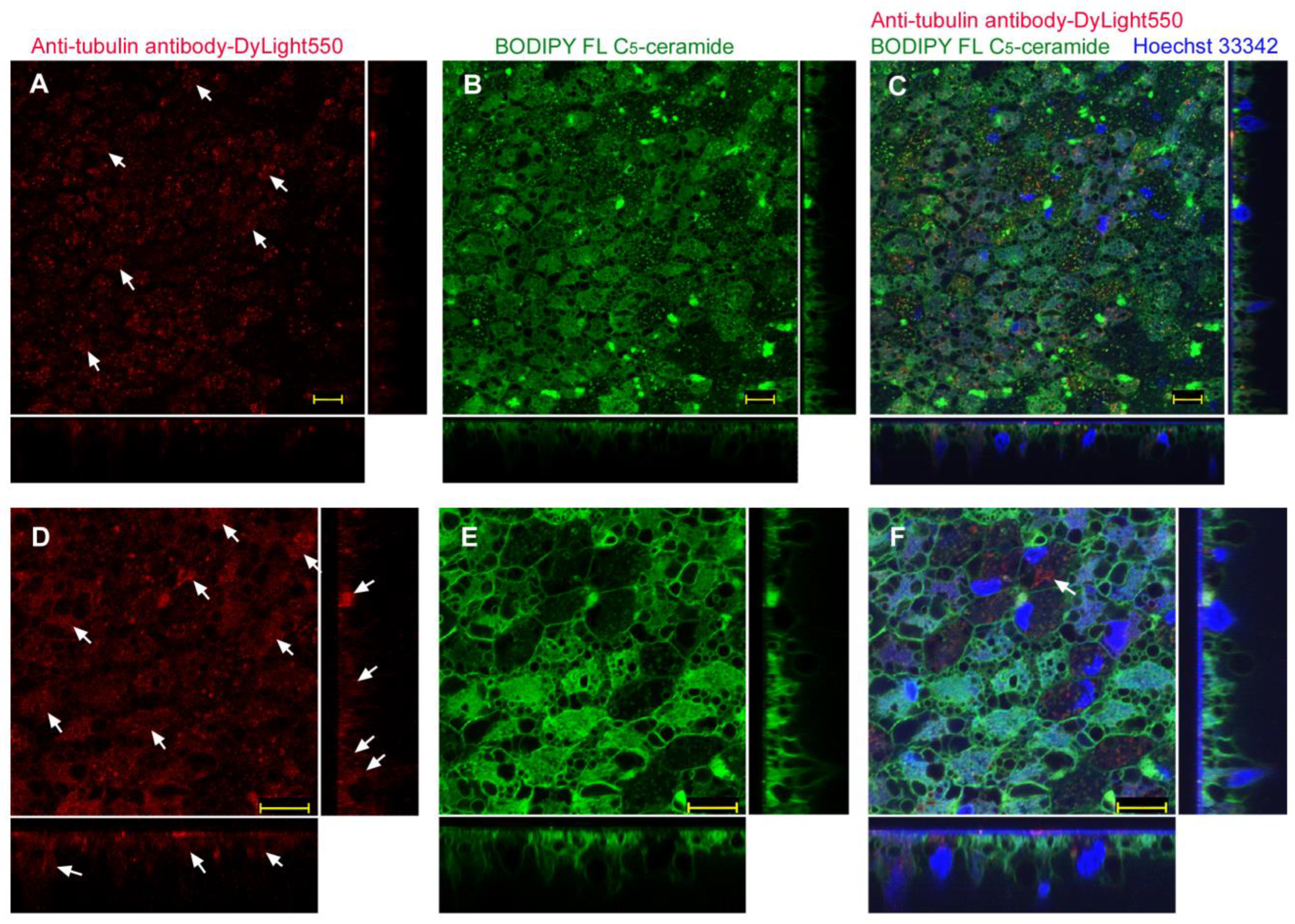
3.3.2. Injection Method
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gantz, V.M.; Akbari, O.S. Gene editing technologies and applications for insects. Curr. Opin. Insect Sci. 2018, 28, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, T.D.; Monteiro, A. CRISPR-Cas9 mediated genome editing in Bicyclus anynana butterflies. Methods Protoc. 2018, 1, 16. [Google Scholar] [CrossRef]
- Ando, T.; Fujiyuki, T.; Kawashima, T.; Morioka, M.; Kudo, T.; Fujiwara, H. In vivo gene transfer into the honeybee using a nucleopolyhedrovirus vector. Biochem. Biophys. Res. Commun. 2007, 352, 335–340. [Google Scholar] [CrossRef]
- Ono, C.; Okamoto, T.; Abe, T.; Matsuura, Y. Baculovirus as a tool for gene delivery and gene therapy. Viruses 2018, 10, 510. [Google Scholar] [CrossRef]
- Naik, N.G.; Lo, Y.-W.; Wu, T.-Y.; Lin, C.-C.; Kuo, S.-C.; Chao, Y.-C. Baculovirus as an efficient vector for gene delivery into mosquitoes. Sci. Rep. 2018, 8, 17778. [Google Scholar] [CrossRef] [PubMed]
- Dhungel, B.; Ohno, Y.; Matayoshi, R.; Otaki, J.M. Baculovirous-mediated gene transfer in butterfly wings in vivo: An efficient expression system with an anti-gb64 antibody. BMC Biotechnol. 2013, 13, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Dhungel, B.; Ohno, Y.; Matayoshi, R.; Iwasaki, M.; Taira, W.; Adhikari, K.; Gurung, R.; Otaki, J.M. Distal-less induces elemental color patterns in Junonia butterfly wings. Zool. Lett. 2016, 2, 4. [Google Scholar] [CrossRef]
- Kaneda, Y. Virosomes: Evolution of the liposome as a targeted drug delivery system. Adv. Drug Deliv. Rev. 2000, 43, 197–205. [Google Scholar] [CrossRef]
- Mastrobattista, E.; Koning, G.A.; van Bloois, L.; Filipe, A.C.; Jiskoot, W.; Storm, G. Functional characterization of an endosome-disruptive peptide and its application in cytosolic delivery of immunoliposome-entrapped proteins. J. Biol. Chem. 2002, 277, 27135–27143. [Google Scholar] [CrossRef] [PubMed]
- Noguchi, H.; Matsushita, M.; Kobayashi, N.; Levy, M.F.; Matsumoto, S. Recent advances in protein transduction technology. Cell Transplant. 2010, 19, 649–654. [Google Scholar] [CrossRef]
- Pae, J.; Pooga, M. Peptide-mediated delivery: An overview of pathways for efficient internalization. Ther. Deliv. 2014, 5, 1203–1222. [Google Scholar] [CrossRef] [PubMed]
- van den Berg, A.; Dowdy, S.F. Protein transduction domain delivery of therapeutic macromolecules. Curr. Opin. Biotechnol. 2011, 22, 888–893. [Google Scholar] [CrossRef]
- Nakase, I.; Kobayashi, S.; Futaki, S. Endosome-disruptive peptides for improving cytosolic delivery of bioactive macromolecules. Biopolymers (Pept. Sci.) 2010, 94, 763–770. [Google Scholar] [CrossRef] [PubMed]
- Varkouhi, A.K.; Scholte, M.; Storm, G.; Haisma, H.J. Endosomal escape pathways for delivery of biologicals. J. Control. Release 2011, 151, 220–228. [Google Scholar] [CrossRef]
- Erazo-Oliveras, A.; Najjar, K.; Dayani, L.; Wang, T.Y.; Johnson, G.A.; Pellois, J.P. Protein delivery into live cells by incubation with an endosomolytic agent. Nat. Methods 2014, 11, 861–867. [Google Scholar] [CrossRef] [PubMed]
- Salerno, J.C.; Ngwa, V.M.; Nowak, S.J.; Chrestensen, C.A.; Healey, A.N.; McMurry, J.L. Novel cell-penetrating peptide-adaptors effect intracellular delivery and endosomal escape of protein cargos. J. Cell Sci. 2016, 129, 893–897. [Google Scholar] [CrossRef]
- Gautam, A.; Nanda, J.S.; Samuel, J.S.; Kumari, M.; Priyanka, P.; Bedi, G.; Nath, S.K.; Mittal, G.; Khatri, N.; Raghava, G.P.S. Topical delivery of protein and peptide using novel cell penetrating peptide IMT-P8. Sci. Rep. 2016, 6, 26278. [Google Scholar] [CrossRef]
- Akishiba, M.; Takeuchi, T.; Kawaguchi, Y.; Sakamoto, K.; Yu, H.; Nakase, I.; Takatani-Nakase, T.; Madani, F.; Gläslund, A.; Futaki, S. Cytosolic antibody delivery by lipid-sensitive endosomolytic peptide. Nat. Chem. 2017, 9, 751–761. [Google Scholar] [CrossRef]
- Verdurmen, W.P.R.; Mazlami, M.; Plückthun, A. A quantitative comparison of cytosolic delivery via different protein uptake systems. Sci. Rep. 2017, 7, 13194. [Google Scholar] [CrossRef]
- Bolhassani, A.; Jafarzade, B.S.; Mardani, G. In vitro and in vivo delivery of therapeutic proteins using cell penetrating peptides. Peptides 2017, 87, 50–63. [Google Scholar] [CrossRef]
- Pei, D.; Buyanova, M. Overcoming endosomal entrapment in drug delivery. Bioconjug. Chem. 2019, 30, 273–283. [Google Scholar] [CrossRef] [PubMed]
- Misao, A.; Futaki, S. Inducible membrane permealization by attenuated lytic peptides: A new concept for accessing cell interiors through ruffled membranes. Mol. Pharmaceutics 2019, 16, 2540–2548. [Google Scholar] [CrossRef]
- Arafiles, J.V.V.; Hirose, H.; Akishiba, M.; Tsuji, S.; Imanishi, M.; Futaki, S. Stimulating macropinocytosis for intracellular nucleic acid and protein delivery: A combined strategy with membrane-lytic peptides to facilitate endosomal escape. Bioconjug. Chem. 2020, 31, 547–553. [Google Scholar] [CrossRef]
- Li, C.; Cao, X.W.; Zhao, J.; Wang, F.J. Effective therapeutic drug delivery by GALA3, an endosomal escape peptide with reduced hydrophobicity. J. Membr. Biol. 2020, 253, 139–152. [Google Scholar] [CrossRef]
- Zhang, K.; Cheng, X.; Zhao, L.; Huang, M.; Tao, Y.; Zhang, H.; Rosenholm, J.M.; Zhuang, M.; Chen, Z.-Y.; Chen, B.; et al. Direct functional protein delivery with a peptide into neonatal and adult mammalian inner ear in vivo. Mol. Ther. Methods Clin. Dev. 2020, 18, 511–519. [Google Scholar] [CrossRef] [PubMed]
- Sahni, A.; Qian, Z.; Pei, D. Cell-penetrating peptides escape the endosome by inducing vesicle budding and collapse. ACS Chem. Biol. 2020, 15, 2485–2492. [Google Scholar] [CrossRef]
- Horn, J.M.; Obermeyer, A.C. Genetic and covalent protein modification strategies to facilitate intracellular delivery. Biomacromolecules 2021, 22, 4883–4904. [Google Scholar] [CrossRef]
- Yu, S.; Yang, H.; Li, T.; Pan, H.; Ren, S.; Luo, G.; Jiang, J.; Yu, L.; Chen, B.; Zhang, Y.; et al. Efficient intracellular delivery of proteins by a multifunctional chimaeric peptide in vitro and in vivo. Nat. Commun. 2021, 12, 5131. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Yang, Y.; Liu, D.; Ji, Y.; Gao, X.; Yin, J.; Yao, W. Cytosolic protein delivery for intracellular antigen targeting using supercharged polypeptide delivery platform. Nano Lett. 2021, 21, 6022–6030. [Google Scholar] [CrossRef]
- Sakamoto, K.; Michibata, J.; Hirai, Y.; Ide, A.; Ikitoh, A.; Takatani-Nakase, T.; Futaki, S. Potentiating the membrane interaction of an attenuated cationic amphiphilic lytic peptide for intracellular protein delivery by anchoring with pyrene moiety. Bioconjug. Chem. 2021, 32, 950–957. [Google Scholar] [CrossRef]
- Gentry, S.B.; Nowak, S.J.; Ni, X.; Hill, S.A.; Wade, L.R.; Clark, W.R.; Keelaghan, A.P.; Morris, D.P.; McMurry, J.L. A real-time assay for cell-penetrating peptide-mediated delivery of molecular cargos. PLoS ONE 2021, 16, e0254468. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, K.; Akishiba, M.; Iwata, T.; Arafiles, J.V.V.; Imanishi, M.; Futaki, S. Use of homoarginine to obtain attenuated cationic membrane lytic peptides. Bioorg. Med. Chem. Lett. 2021, 40, 127925. [Google Scholar] [CrossRef] [PubMed]
- Teo, S.L.Y.; Rennick, J.J.; Yuen, D.; Al-Wassiti, H.; Johnston, A.P.R.; Pouton, C.W. Unravelling cytosolic delivery of cell penetrating peptides with a quantitative endosomal escape assay. Nat. Commun. 2021, 12, 3721. [Google Scholar] [CrossRef]
- Tietz, O.; Cortezon-Tamarit, F.; Chalk, R.; Able, S.; Vallis, K.A. Tricyclic cell-penetrating peptides for efficient delivery of functional antibodies into cancer cells. Nat. Chem. 2022, 14, 284–293. [Google Scholar] [CrossRef]
- Shirai, Y.; Piulachs, M.D.; Belles, X.; Daimon, T. DIPA-CRISPR is a simple and accessible method for insect gene editing. Cell Rep. Methods 2022, 2, 100215. [Google Scholar] [CrossRef]
- Özsu, N.; Chan, Q.Y.; Chen, B.; Gupta, M.D.; Monteiro, A. Wingless is a positive regulator of eyespot color patterns in Bicyclus anynana butterflies. Dev. Biol. 2017, 429, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Reed, R.D. Genome editing in butterflies reveals that spalt promotes and Distal-less represses eyespot colour patterns. Nat. Commun. 2016, 7, 11769. [Google Scholar] [CrossRef] [PubMed]
- Martin, A.; Reed, R.D. Wnt signaling underlies evolution and development of the butterfly wing pattern symmetry systems. Dev. Biol. 2014, 395, 367–378. [Google Scholar] [CrossRef]
- Mazo-Vargas, A.; Concha, C.; Livraghi, L.; Massardo, D.; Wallbank, R.W.R.; Zhang, L.; Papador, J.D.; Martinez-Najera, D.; Jiggins, C.D.; Kronforst, M.R.; et al. Macroevolutionary shifts of WntA function potentiate butterfly wing-pattern diversity. Proc. Natl. Acad. Sci. USA 2017, 114, 10701–10706. [Google Scholar] [CrossRef]
- Nijhout, H.F. The Development and Evolution of Butterfly Wing Patterns; Smithsonian Institution Press: Washington, DC, USA, 1991. [Google Scholar]
- Otaki, J.M. 2008. Physiologically induced color-pattern changes in butterfly wings: Mechanistic and evolutionary implications. J. Insect Physiol. 2008, 54, 1099–1112. [Google Scholar] [CrossRef]
- Nishikawa, H.; Iijima, T.; Kajitani, R.; Yamaguchi, J.; Ando, T.; Suzuki, Y.; Sugano, S.; Fujiyama, A.; Kosugi, S.; Hirakawa, H.; et al. A genetic mechanism for female-limited Batesian mimicry in Papilio butterfly. Nat. Genet. 2015, 47, 405–409. [Google Scholar] [CrossRef] [PubMed]
- Ando, T.; Fujiwara, H. Electroporation-mediated somatic transgenesis for rapid functional analysis in insects. Development 2013, 140, 454–458. [Google Scholar] [CrossRef] [PubMed]
- Kusaba, K.; Otaki, J.M. Positional dependence of scale size and shape in butterfly wings: Wing-wide phenotypic coordination of color-pattern elements and background. J. Insect Physiol. 2009, 55, 174–182. [Google Scholar] [CrossRef] [PubMed]
- Iwata, M.; Ohno, Y.; Otaki, J.M. Real-time in vivo imaging of butterfly wing development: Revealing the cellular dynamics of the pupal wing tissue. PLoS ONE 2014, 9, e89500. [Google Scholar] [CrossRef] [PubMed]
- Ohno, Y.; Otaki, J.M. Live cell imaging of butterfly pupal and larval wings in vivo. PLoS ONE 2015, 10, e0128332. [Google Scholar] [CrossRef]
- Dhungel, B.; Otaki, J.M. Local pharmacological effects of tungstate on the color-pattern determination of butterfly wings: A possible relationship between the eyespot and parafocal element. Zool. Sci. 2009, 26, 758–764. [Google Scholar] [CrossRef]
- Iwasaki, M.; Ohno, Y.; Otaki, J.M. Butterfly eyespot organiser: In Vivo imaging of the prospective focal cells in pupal wing tissues. Sci. Rep. 2017, 7, 40705. [Google Scholar] [CrossRef]
- Hirata, K.; Otaki, J.M. Real-time in vivo imaging of the developing pupal wing tissues in the pale grass blue butterfly Zizeeria maha: Establishing the lycaenid system for multiscale bioimaging. J. Imaging 2019, 5, 42. [Google Scholar] [CrossRef]
- Otaki, J.M. Color-pattern modifications of butterfly wings induced by transfusion and oxyanions. J. Insect Physiol. 1998, 44, 1181–1190. [Google Scholar] [CrossRef]
- Otaki, J.M.; Nakazato, Y. Butterfly wing color patten modification inducers may act on chitin in the apical extracellular site: Implications in morphogenic signals for color pattern determination. Biology 2022, 11, 1620. [Google Scholar] [CrossRef]
- Funakoshi. New Type Protein Transfection Reagent ProteoCarry. Web Page No: 80968. 28 November 2017. Available online: https://www.funakoshi.co.jp/exports_contents/80968 (accessed on 29 November 2022).
- Hiyama, A.; Iwata, M.; Otaki, J.M. Rearing the pale grass blue Zizeeria maha (Lepidoptera: Lycaenidae): Toward the establishment of a lycaenid model system for butterfly physiology and genetics. Entomol. Sci. 2010, 13, 293–302. [Google Scholar] [CrossRef]
- Otaki, J.M.; Hiyama, A.; Iwata, M.; Kudo, T. Phenotypic plasticity in the range-margin population of the lycaenid butterfly Zizeeria maha. BMC Evol. Biol. 2010, 10, 252. [Google Scholar] [CrossRef]
- Muyldermans, S. Nanobodies: Natural single-domain antibodies. Annu. Rev. Biochem. 2013, 82, 775–779. [Google Scholar] [CrossRef]
- Moussian, B.; Schwarz, H.; Bartoszewski, S.; Nüsslein-Volhard, C. Involvement of chitin in exoskeleton morphogenesis in Drosophila melanogaster. J. Morphol. 2005, 264, 117–130. [Google Scholar] [CrossRef] [PubMed]
- Otaki, J.M. Butterfly eyespot color pattern formation requires physical contact of the pupal wing epithelium with extracellular materials for morphogenic signal propagation. BMC Dev. Biol. 2020, 20, 6. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Buchwalter, R.A.; Zheng, C.; Wight, E.M.; Chen, J.V.; Megraw, T.L. A perinuclear microtubule-organizing centre controls nuclear positioning and basement membrane secretion. Nat. Cell Biol. 2020, 22, 297–309. [Google Scholar] [CrossRef]
- Li, J.; Wu, F.; Cheng, L.; Zhang, J.; Cha, C.; Chen, L.; Feng, T.; Zhang, J.; Guo, G. A nuclear localization signal is required for the nuclear translocation of Fign and its microtubule-serving function. Mol. Med. Rep. 2020, 21, 2367–2374. Available online: https://europepmc.org/article/MED/32236575 (accessed on 29 November 2022).













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sandwich Method | Injection Method | |||
|---|---|---|---|---|
| Delivery Reagent | GFP, OFP | Antibodies | GFP, OFP | Antibodies |
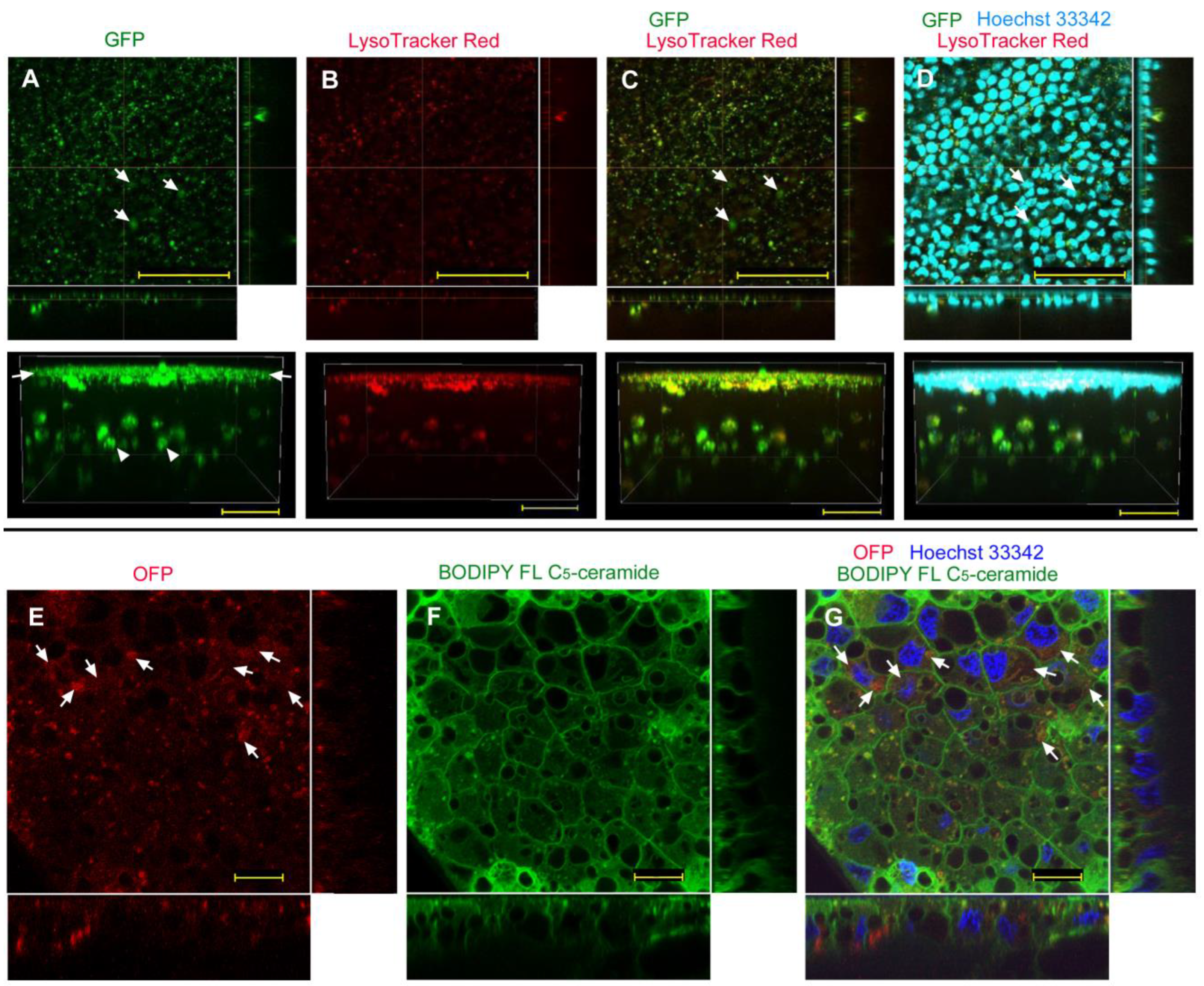
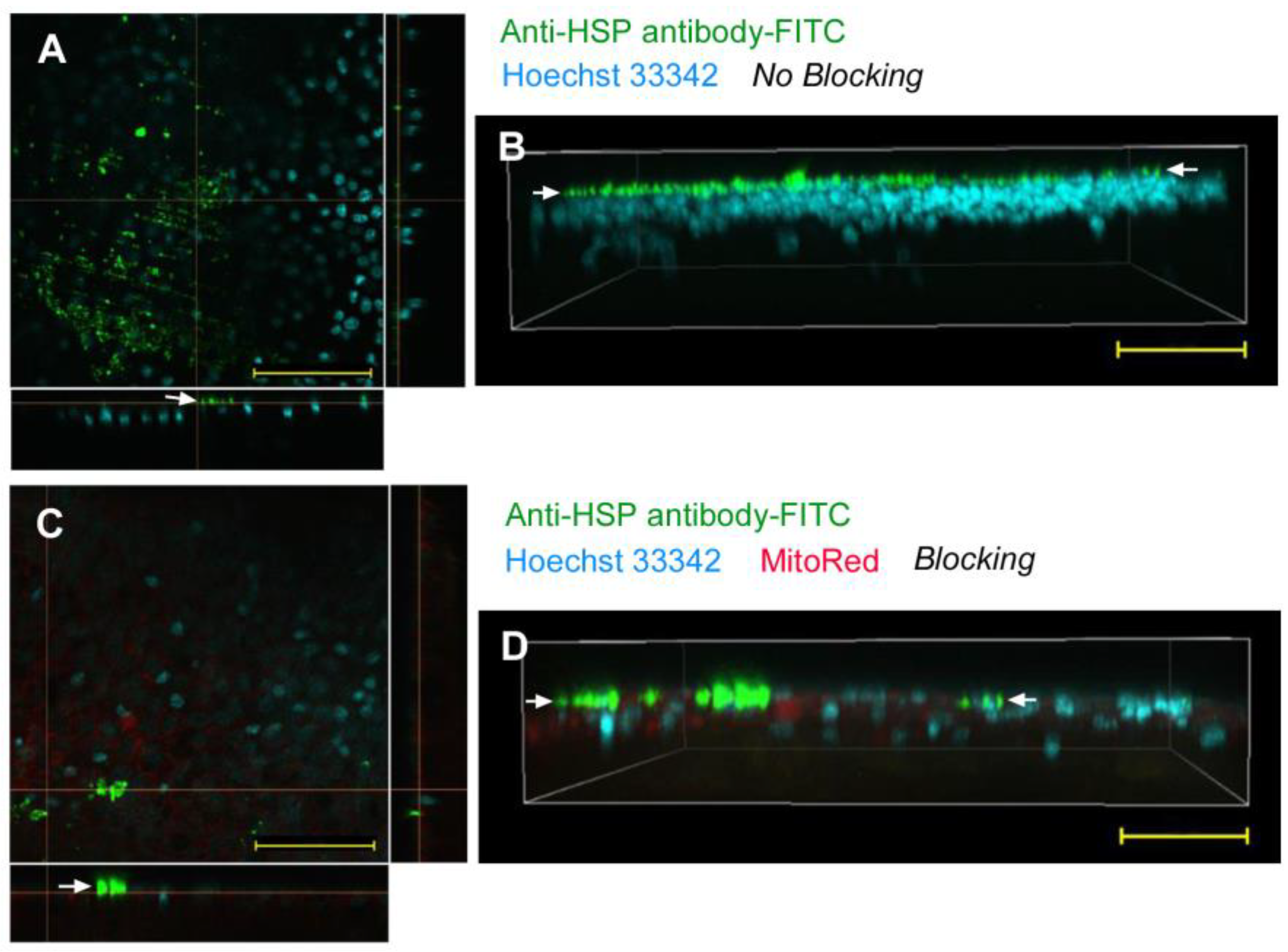
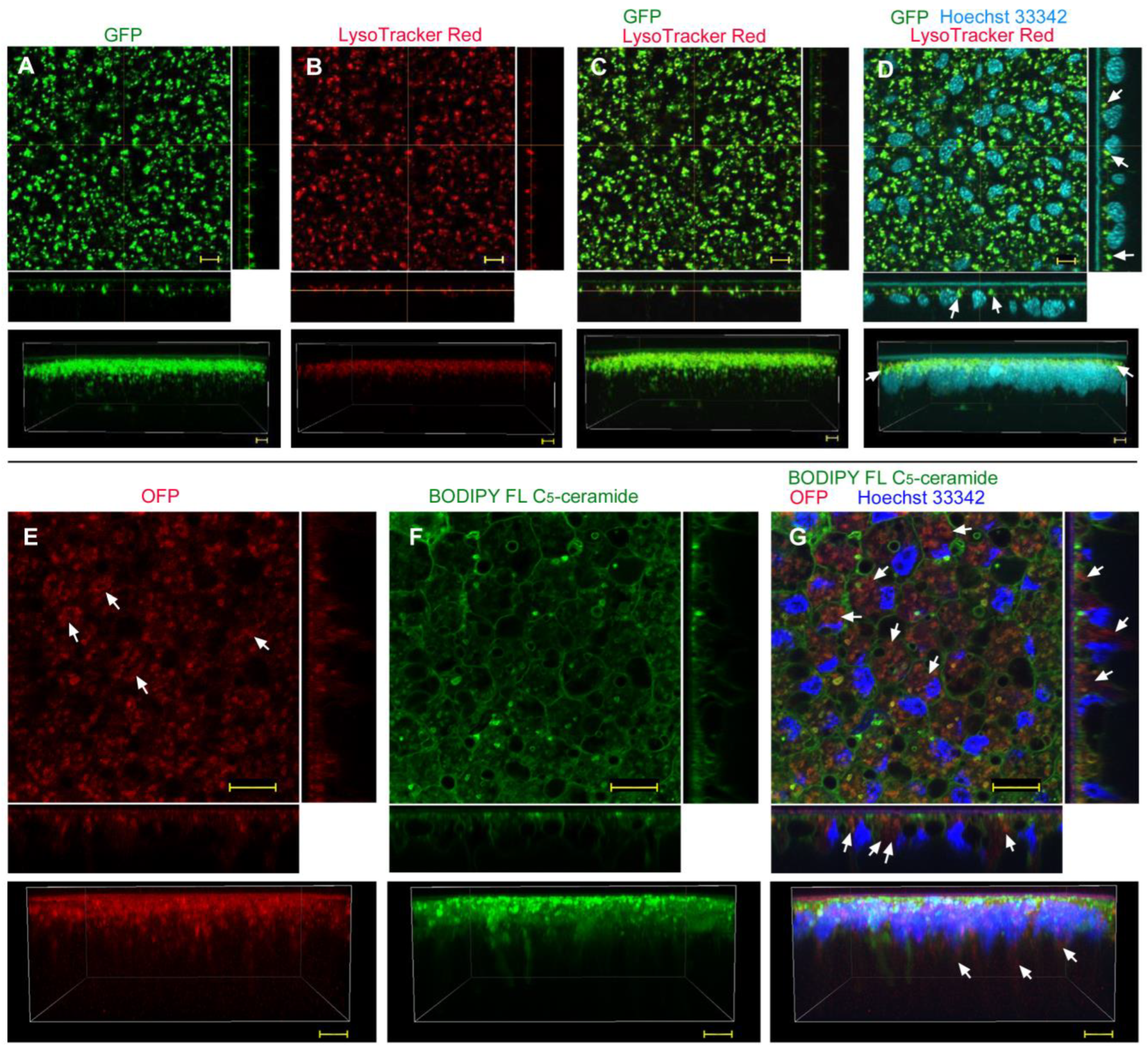
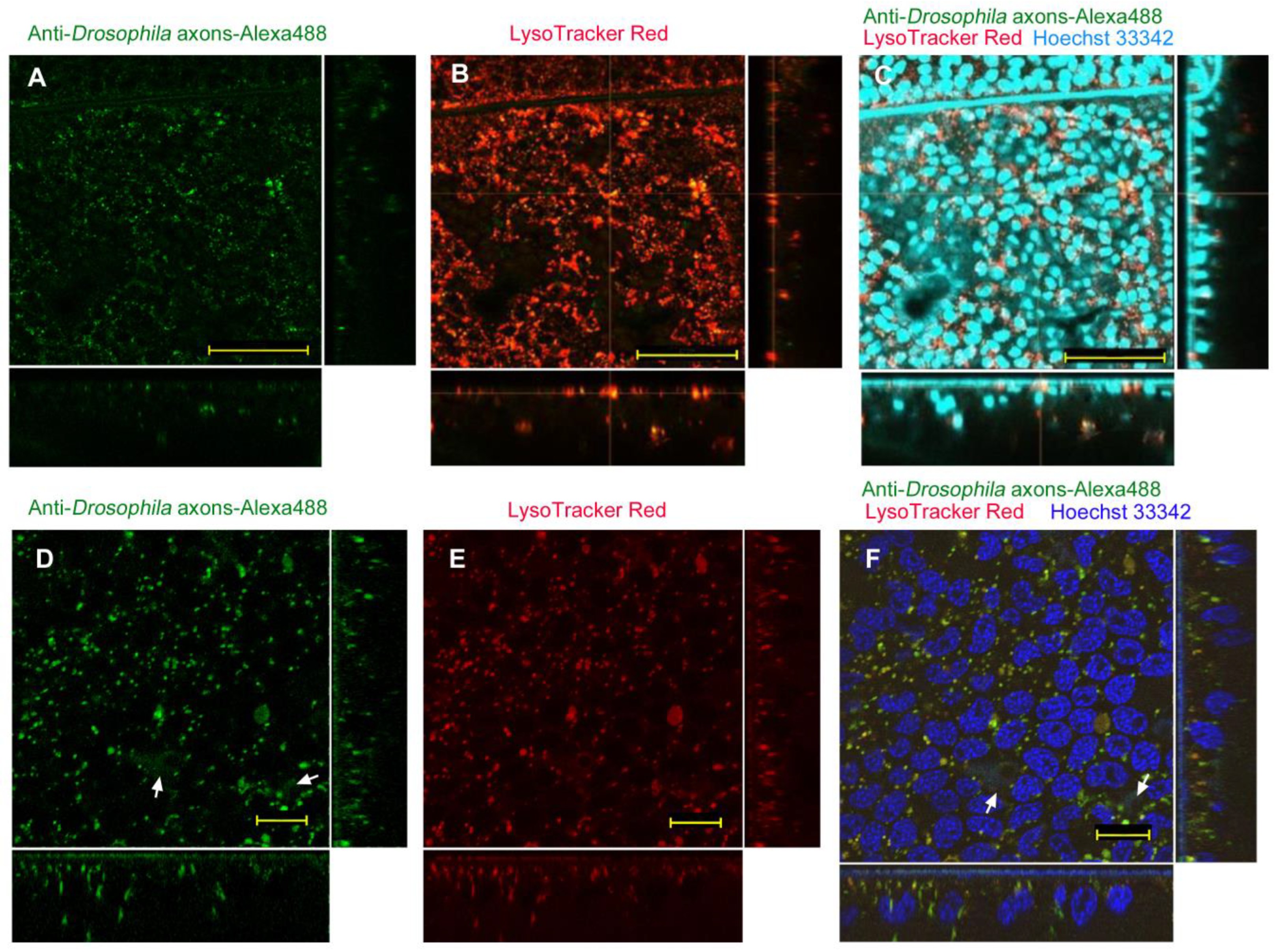
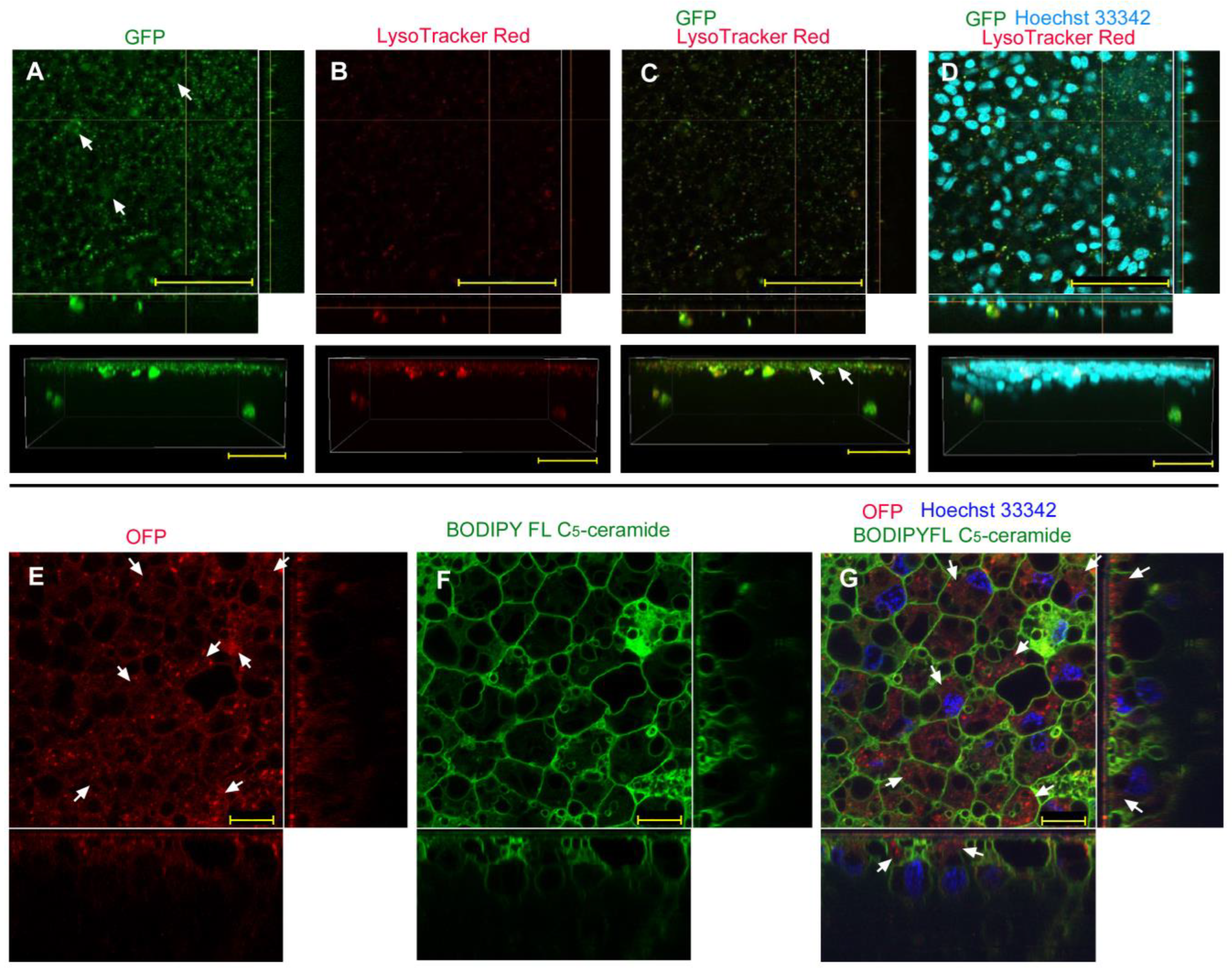
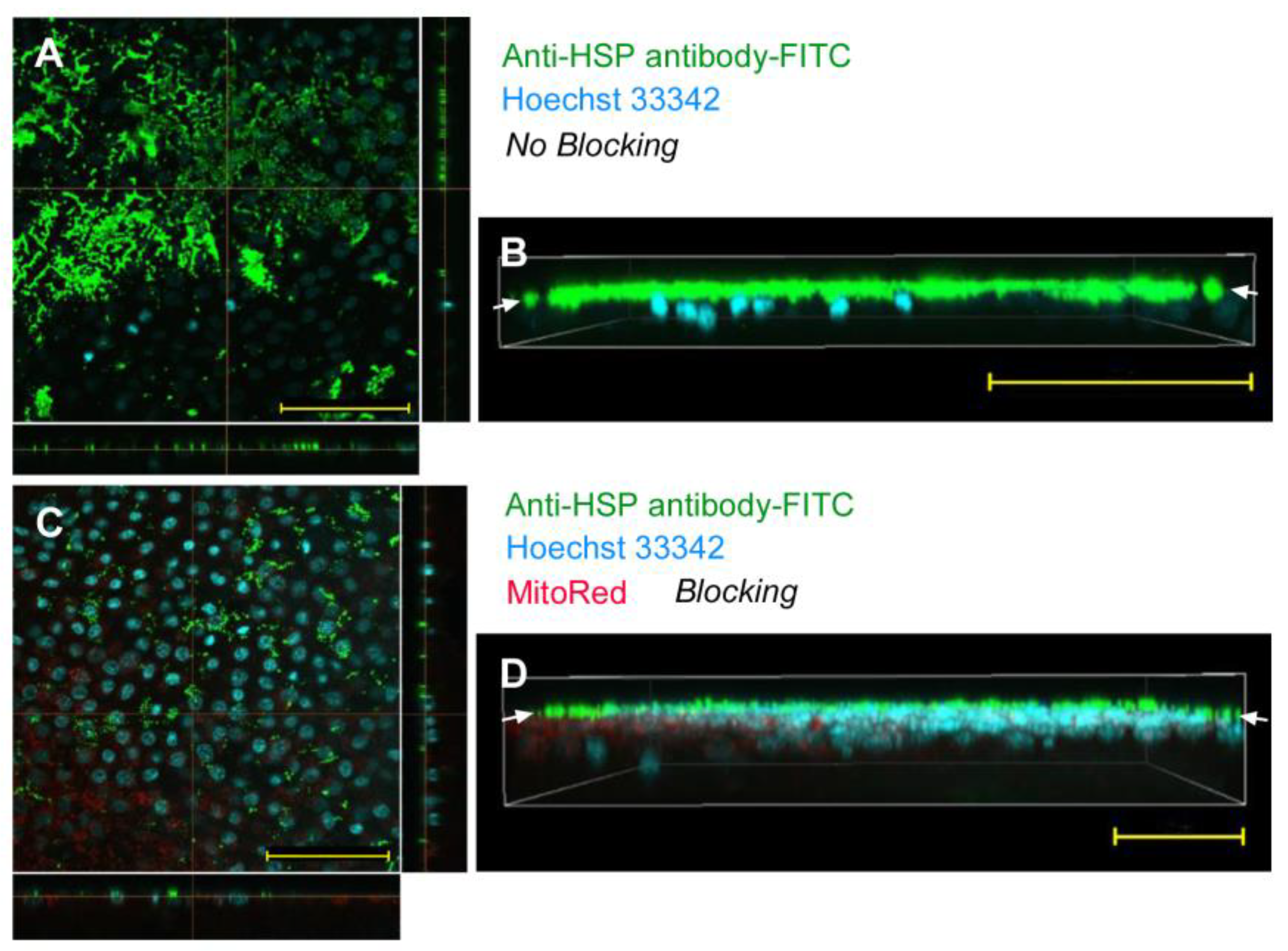
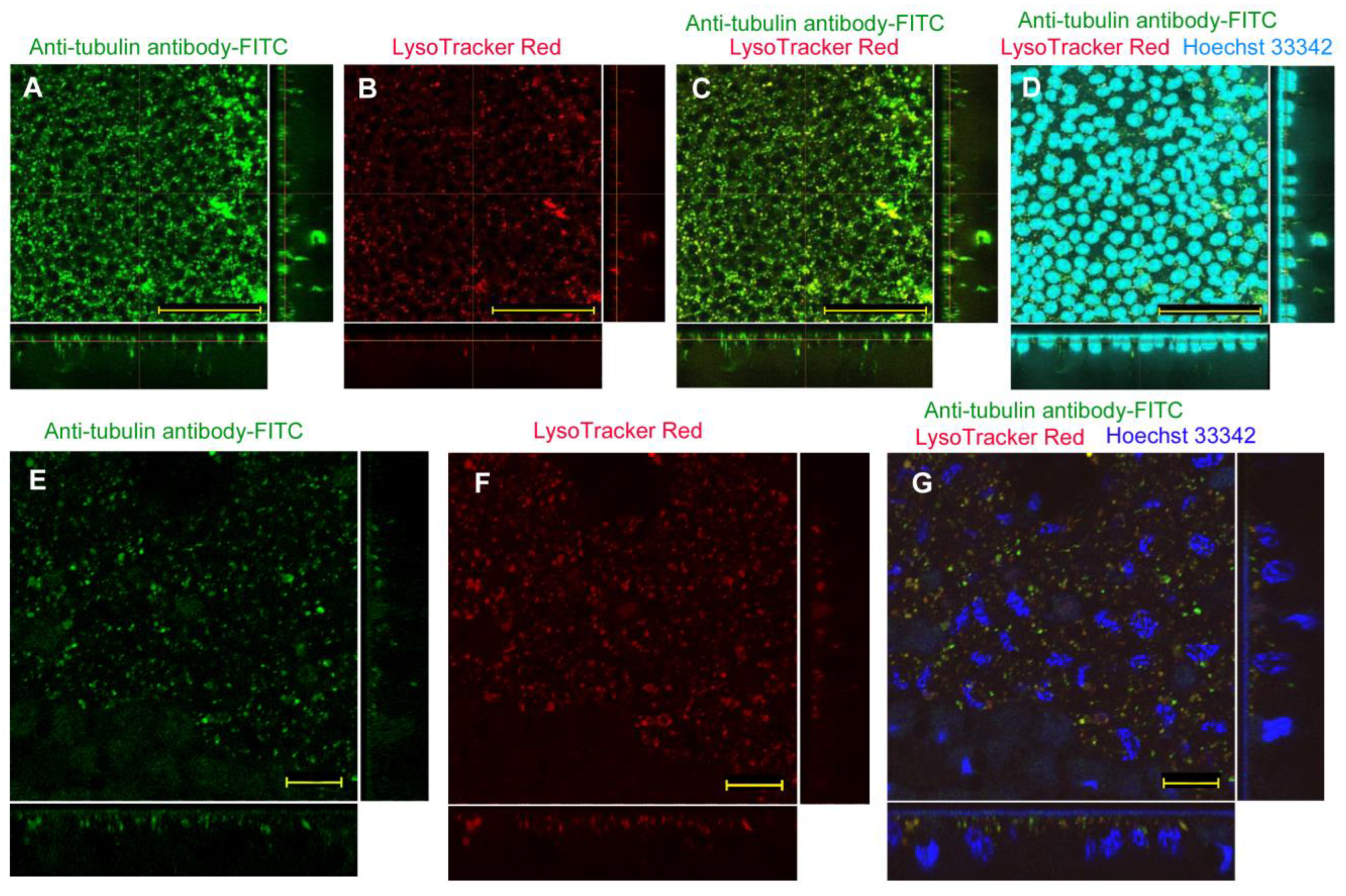
| No reagent | Endosome + Cytosol (Figure 1) | No delivery (Figure 2) | Endosome + Cytosol (Figure 3) | Endosome (Figure 4) |
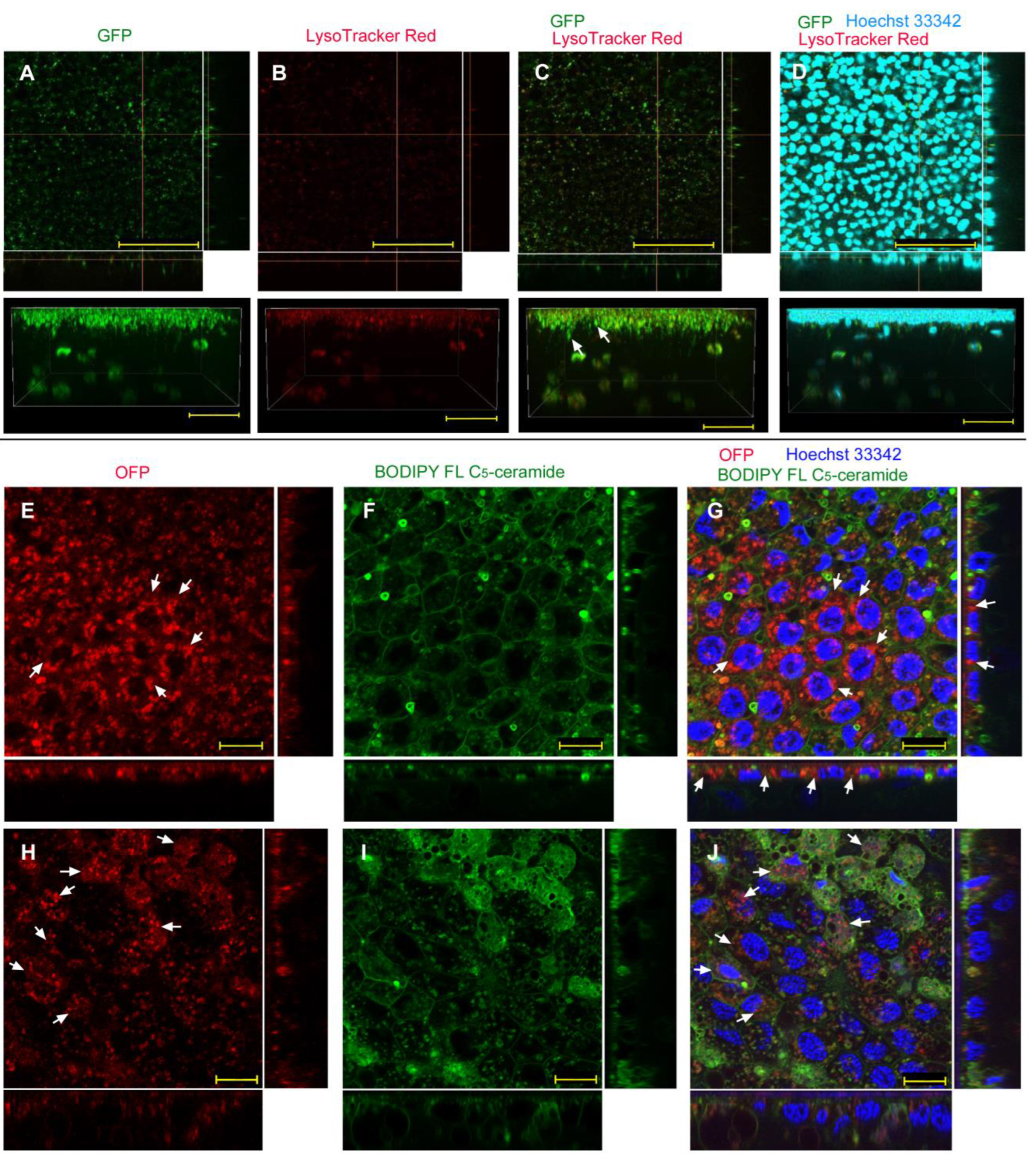
| L17E | Endosome + Cytosol (Figure 5) | No delivery (Figure 6) | Endosome + Cytosol (Figure 7) | Endosome (Figure 8) |
| ProteoCarry | Endosome + Cytosol (Figure 9) | Endosome + Cytosol (Figure 10) | Endosome + Cytosol (Figure 11) | Endosome + Cytosol (Figure 12) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nakazato, Y.; Otaki, J.M. Protein Delivery to Insect Epithelial Cells In Vivo: Potential Application to Functional Molecular Analysis of Proteins in Butterfly Wing Development. BioTech 2023, 12, 28. https://doi.org/10.3390/biotech12020028
Nakazato Y, Otaki JM. Protein Delivery to Insect Epithelial Cells In Vivo: Potential Application to Functional Molecular Analysis of Proteins in Butterfly Wing Development. BioTech. 2023; 12(2):28. https://doi.org/10.3390/biotech12020028
Chicago/Turabian StyleNakazato, Yugo, and Joji M. Otaki. 2023. "Protein Delivery to Insect Epithelial Cells In Vivo: Potential Application to Functional Molecular Analysis of Proteins in Butterfly Wing Development" BioTech 12, no. 2: 28. https://doi.org/10.3390/biotech12020028