
Fruit Color Preference of Frugivorous Birds in an Agroecosystem in Southcentral Mindanao, Philippines
 ,
,  , , , and
, , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
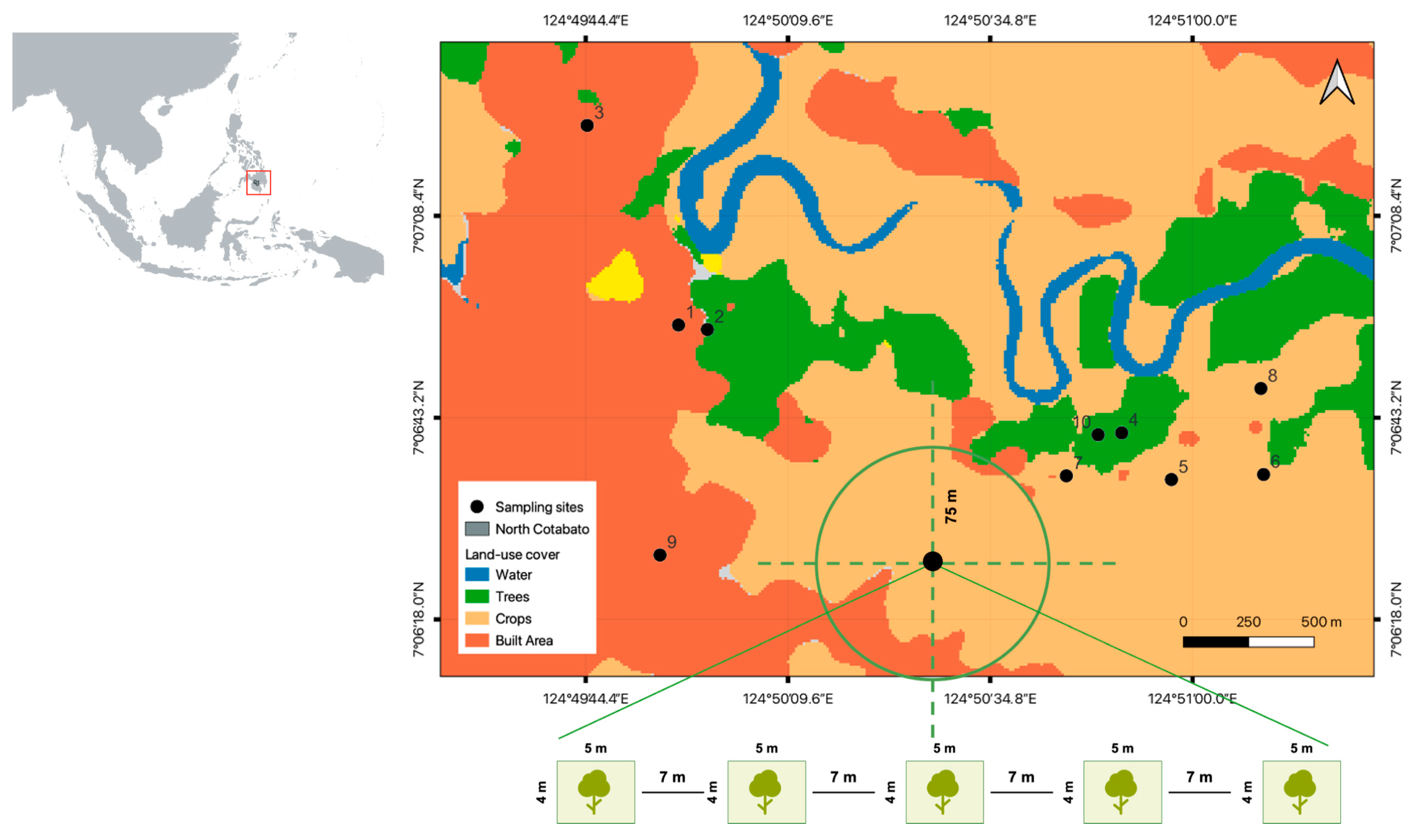
2.1. Study Areas and Sampling Design

2.2. Land Use Cover and Environmental Variables
2.3. Statistical Analyses
3. Results
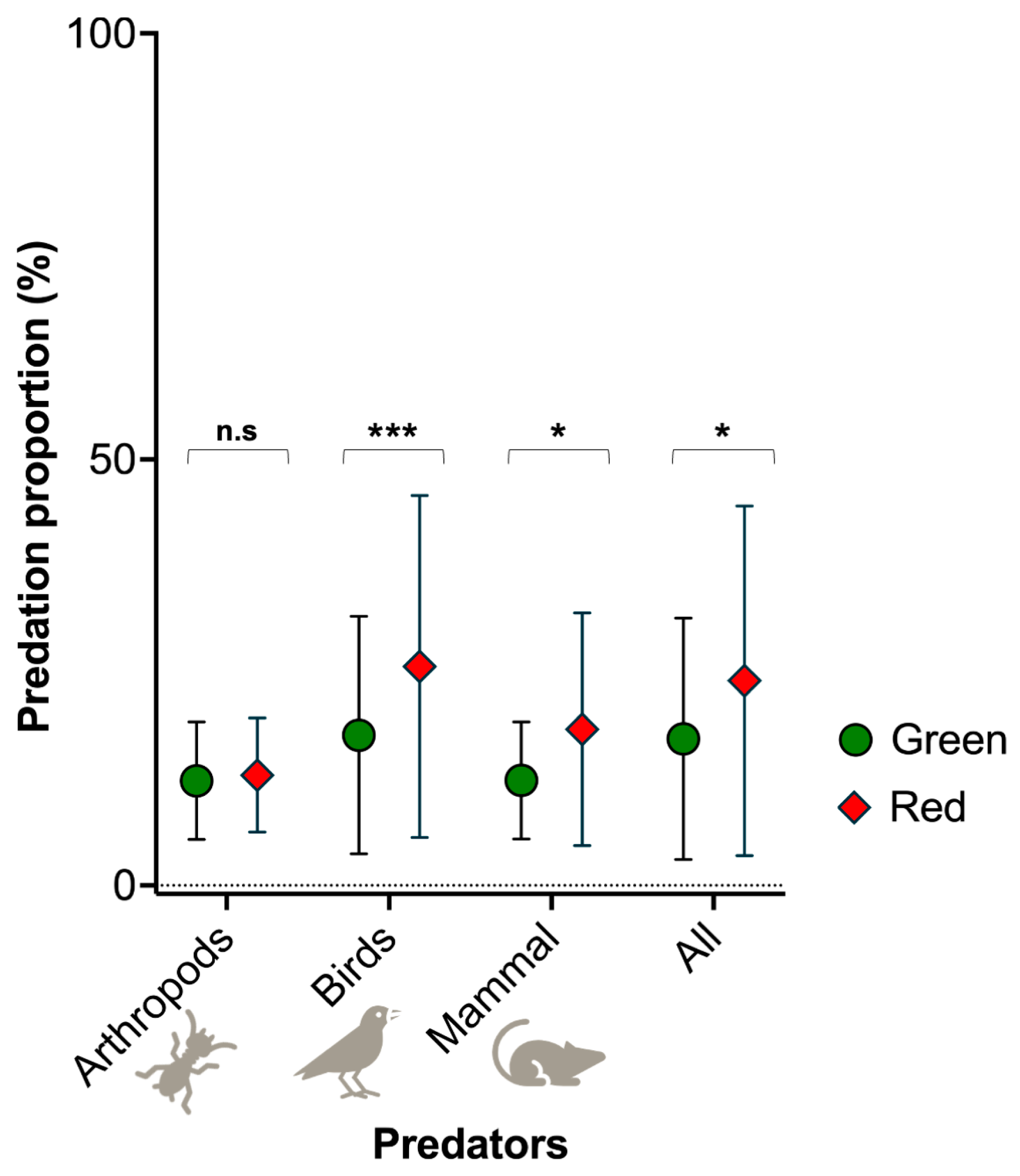
3.1. Fruit Predation Proportion (%) in an Agroecosystem
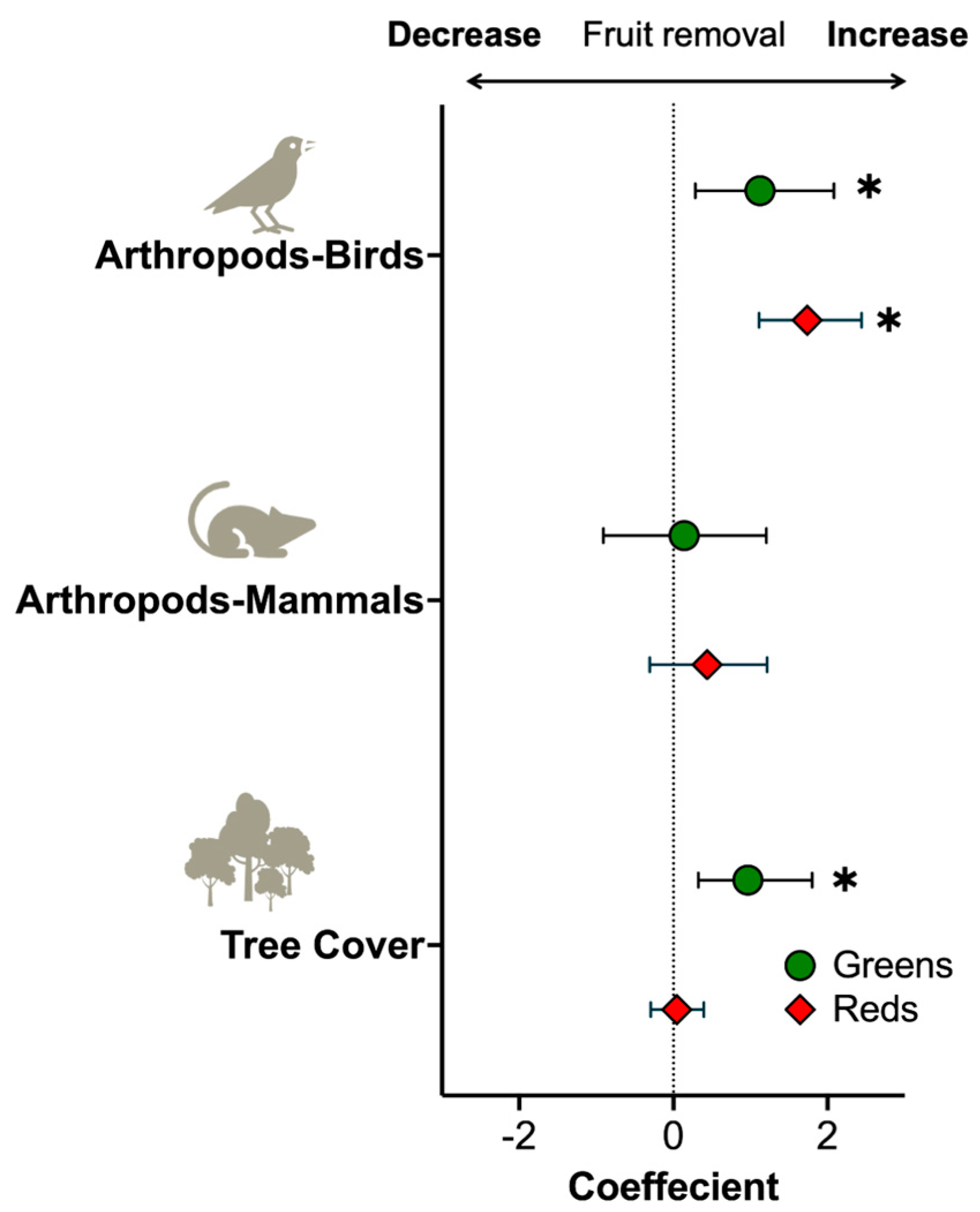
3.2. Risk of Predator Attack
3.3. Risk of Predation by Fruit Color
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aziz, S.A.; McConkey, K.R.; Tanalgo, K.; Sritongchuay, T.; Low, M.-R.; Yong, J.Y.; Mildenstein, T.L.; Nuevo-Diego, C.E.; Lim, V.-C.; Racey, P.A. The Critical Importance of Old World Fruit Bats for Healthy Ecosystems and Economies. Front. Ecol. Evol. 2021, 9, 641411. [Google Scholar] [CrossRef]
- Camargo, P.H.S.A.; Carlo, T.A.; Brancalion, P.H.S.; Pizo, M.A. Frugivore Diversity Increases Evenness in the Seed Rain on Deforested Tropical Landscapes. Oikos 2022, 2022, e08028. [Google Scholar] [CrossRef]
- Monteiro, E.C.S.; Pizo, M.A.; Vancine, M.H.; Ribeiro, M.C. Forest Cover and Connectivity Have Pervasive Effects on the Maintenance of Evolutionary Distinct Interactions in Seed Dispersal Networks. Oikos 2022, 2022, e08028. [Google Scholar] [CrossRef]
- Canterbury, G.E.; Martin, T.E.; Petit, D.R.; Petit, L.J.; Bradford, D.F. Bird Communities and Habitat as Ecological Indicators of Forest Condition in Regional Monitoring. Conserv. Biol. 2000, 14, 544–558. [Google Scholar] [CrossRef]
- Fraixedas, S.; Lindén, A.; Piha, M.; Cabeza, M.; Gregory, R.; Lehikoinen, A. A State-of-the-Art Review on Birds as Indicators of Biodiversity: Advances, Challenges, and Future Directions. Ecol. Indic. 2020, 118, 106728. [Google Scholar] [CrossRef]
- Newbold, T.; Hudson, L.; Hill, S.; Contu, S.; Lysenko, I.; Senior, R.; Börger, L.; Bennett, D.; Choimes, A.; Collen, B.; et al. Global Effects of Land Use on Local Terrestrial Biodiversity. Nature 2015, 520, 45–50. [Google Scholar] [CrossRef]
- Tanalgo, K.C.; Sritongchuay, T.; Hughes, A.C. Seasonal Activity of Fruit Bats in a Monoculture Rubber and Oil Palm Plantation in the Southern Philippines. Conservation 2021, 1, 258–269. [Google Scholar] [CrossRef]
- Tanalgo, K.C.; Achondo, M.J.M.M.; Hughes, A.C. Small Things Matter: The Value of Rapid Biodiversity Surveys to Understanding Local Bird Diversity Patterns in Southcentral Mindanao, Philippines. Trop. Conserv. Sci. 2019, 12, 1940082919869482. [Google Scholar] [CrossRef]
- Gonzales, R.; Ingle, N.; Lagunzad, D.; Nakashizuka, T. Seed Dispersal by Birds and Bats in Lowland Philippine Forest Successional Area. Biotropica 2009, 41, 452–458. [Google Scholar] [CrossRef]
- Ingle, N.R. Seed Dispersal by Wind, Birds, and Bats between Philippine Montane Rainforest and Successional Vegetation. Oecologia 2003, 134, 251–261. [Google Scholar] [CrossRef]
- Heleno, R.H.; Ross, G.; Everard, A.; Memmott, J.; Ramos, J.A. The Role of Avian ‘Seed Predators’ as Seed Dispersers. Ibis 2011, 153, 199–203. [Google Scholar] [CrossRef]
- Sritongchuay, T.; Gale, G.A.; Stewart, A.; Kerdkaew, T.; Bumrungsri, S. Seed Rain in Abandoned Clearings in a Lowland Evergreen Rain Forest in Southern Thailand. Trop. Conserv. Sci. 2014, 7, 572–585. [Google Scholar] [CrossRef]
- Willson, M.F.; Traveset, A. The Ecology of Seed Dispersal. In Seeds: The Ecology of Regeneration in Plant Communities; CABI: Wallingford, UK, 2000; pp. 85–110. [Google Scholar] [CrossRef]
- Wang, B.; Smith, T. Closing the Seed Dispersal Loop. Trends Ecol. Evol. 2002, 17, 379–386. [Google Scholar] [CrossRef]
- Camargo, P.H.S.A.; Pizo, M.A.; Brancalion, P.H.S.; Carlo, T.A. Fruit Traits of Pioneer Trees Structure Seed Dispersal across Distances on Tropical Deforested Landscapes: Implications for Restoration. J. Appl. Ecol. 2020, 57, 2329–2339. [Google Scholar] [CrossRef]
- Pilling, D.; Bélanger, J.; Hoffmann, I. Declining Biodiversity for Food and Agriculture Needs Urgent Global Action. Nat. Food 2020, 1, 144–147. [Google Scholar] [CrossRef]
- Raven, P.H.; Wagner, D.L. Agricultural Intensification and Climate Change Are Rapidly Decreasing Insect Biodiversity. Proc. Natl. Acad. Sci. USA 2021, 118, e2002548117. [Google Scholar] [CrossRef]
- Sun, Z.; Behrens, P.; Tukker, A.; Bruckner, M.; Scherer, L. Global Human Consumption Threatens Key Biodiversity Areas. Environ. Sci. Technol. 2022, 56, 9003–9014. [Google Scholar] [CrossRef]
- Koh, L.P.; Wilcove, D.S. Cashing in Palm Oil for Conservation. Nature 2007, 448, 993–994. [Google Scholar] [CrossRef]
- Sodhi, N.S.; Koh, L.P.; Clements, R.; Wanger, T.C.; Hill, J.K.; Hamer, K.C.; Clough, Y.; Tscharntke, T.; Posa, M.R.C.; Lee, T.M. Conserving Southeast Asian Forest Biodiversity in Human-Modified Landscapes. Biol. Conserv. 2010, 143, 2375–2384. [Google Scholar] [CrossRef]
- Suhonen, J.; Jokimäki, J. Fruit Removal from Rowanberry (Sorbus aucuparia) Trees at Urban and Rural Areas in Finland: A Multi-Scale Study. Landsc. Urban Plan. 2015, 137, 13–19. [Google Scholar] [CrossRef]
- Suhonen, J.; Jokimäki, J.; Lassila, R.; Kaisanlahti-Jokimäki, M.-L.; Carbó-Ramírez, P. Effects of Roads on Fruit Crop and Removal Rate from Rowanberry Trees (Sorbus aucuparia) by Birds in Urban Areas of Finland. Urban For. Urban Green. 2017, 27, 148–154. [Google Scholar] [CrossRef]
- Lomascolo, S.; Schaefer, H. Signal Convergence in Fruits: A Result of Selection by Frugivores? J. Evol. Biol. 2010, 23, 614–624. [Google Scholar] [CrossRef] [PubMed]
- Martin Schaefer, H.; Schaefer, V.; Vorobyev, M. Are Fruit Colors Adapted to Consumer Vision and Birds Equally Efficient in Detecting Colorful Signals? Am. Nat. 2007, 169 (Suppl. 1), S159–S169. [Google Scholar] [CrossRef]
- Cazetta, E.; Schaefer, H.M.; Galetti, M. Why Are Fruits Colorful? The Relative Importance of Achromatic and Chromatic Contrasts for Detection by Birds. Evol. Ecol. 2009, 23, 233–244. [Google Scholar] [CrossRef]
- Galetti, M.; Alves-Costa, C.; Cazetta, E. Effects of Forest Fragmentation, Anthropogenic Edges and Fruit Colour on the Consumption of Ornithocoric Fruits. Biol. Conserv. 2003, 111, 269–273. [Google Scholar] [CrossRef]
- Melo, G.; Penatti, N.; Raizer, J. Fruit of a Contrasting Colour Is More Detectable by Frugivores. J. Trop. Ecol. 2011, 27, 319–322. [Google Scholar] [CrossRef]
- Rey, P.J. Preserving Frugivorous Birds in Agro-Ecosystems: Lessons from Spanish Olive Orchards. J. Appl. Ecol. 2011, 48, 228–237. [Google Scholar] [CrossRef]
- Bieber, A.G.D.; Silva, P.S.D.; Sendoya, S.F.; Oliveira, P.S. Assessing the Impact of Deforestation of the Atlantic Rainforest on Ant-Fruit Interactions: A Field Experiment Using Synthetic Fruits. PLoS ONE 2014, 9, e90369. [Google Scholar] [CrossRef]
- Sritongchuay, T.; Kremen, C.; Bumrungsri, S. Effects of Forest and Cave Proximity on Fruit Set of Tree Crops in Tropical Orchards in Southern Thailand. J. Trop. Ecol. 2016, 32, 269–279. [Google Scholar] [CrossRef]
- Enriquez, T.; Perez-Torres, J.; Ruiz, C.; Suarez, A. Seed Dispersal by Fruit Bats in Colombia Generates Ecosystem Services. Agron. Sustain. Dev. 2020, 40, 1–15. [Google Scholar] [CrossRef]
- Genes, L.; Dirzo, R. Restoration of Plant-Animal Interactions in Terrestrial Ecosystems. Biol. Conserv. 2022, 265, 109393. [Google Scholar] [CrossRef]
- Teixido, A.L.; Fuzessy, L.F.; Souza, C.S.; Gomes, I.N.; Kaminski, L.A.; Oliveira, P.C.; Maruyama, P.K. Anthropogenic Impacts on Plant-Animal Mutualisms: A Global Synthesis for Pollination and Seed Dispersal. Biol. Conserv. 2022, 266, 109461. [Google Scholar] [CrossRef]
- Sritongchuay, T.; Dalsgaard, B.; Wayo, K.; Zou, Y.; Simla, P.; Tanalgo, K.C.; Orr, M.C.; Hughes, A.C. Landscape-Level Effects on Pollination Networks and Fruit-Set of Crops in Tropical Small-Holder Agroecosystems. Agric. Ecosyst. Environ. 2022, 339, 108112. [Google Scholar] [CrossRef]
- Sallabanks, R. Hierarchical Mechanisms of Fruit Selection by an Avian Frugivore. Ecology 1993, 74, 1326. [Google Scholar] [CrossRef]
- Alves-Costa, C.; Lopes, A. Using Artificial Fruits to Evaluate Fruit Selection by Birds in the Field1. Biotropica 2006, 33, 713–717. [Google Scholar] [CrossRef]
- Pena, J.C.; Aoki-Gonçalves, F.; Dáttilo, W.; Ribeiro, M.C.; MacGregor-Fors, I. Caterpillars’ Natural Enemies and Attack Probability in an Urbanization Intensity Gradient across a Neotropical Streetscape. Ecol. Indic. 2021, 128, 107851. [Google Scholar] [CrossRef]
- Karra, K.; Kontgis, C.; Statman-Weil, Z.; Mazzariello, J.C.; Mathis, M.; Brumby, S.P. Global Land Use / Land Cover with Sentinel 2 and Deep Learning. In Proceedings of the 2021 IEEE International Geoscience and Remote Sensing Symposium IGARSS, Brussels, Belgium, 11–16 July 2021; pp. 4704–4707. [Google Scholar]
- Leles, B.; Xiao, X.; Pasion, B.; Nakamura, A.; Tomlinson, K. Does Plant Diversity Increase Top-down Control of Herbivorous Insects in Tropical Forest? Oikos 2017, 126, 1142–1149. [Google Scholar] [CrossRef]
- Low, P.A.; Sam, K.; McArthur, C.; Posa, M.R.C.; Hochuli, D.F. Determining Predator Identity from Attack Marks Left in Model Caterpillars: Guidelines for Best Practice. Entomol. Exp. Appl. 2014, 152, 120–126. [Google Scholar] [CrossRef]
- QGIS Development Team QGIS Geographic Information System. Open Source Geospatial Foundation Project. 2022. Available online: http://qgis.osgeo.org (accessed on 12 December 2022).
- Burnham, K.P.; Anderson, D.R. Multimodel Inference: Understanding AIC and BIC in Model Selection. Sociol. Methods Res. 2004, 33, 261–304. [Google Scholar] [CrossRef]
- The Jamovi Project Jamovi (Version 2.3) [Computer Software]. 2023. Available online: https://www.jamovi.org/ (accessed on 3 January 2023).
- GraphPad Prism; GraphPad Software, Inc.: San Diego, CA, USA, 2022.
- Lim, G.; Burns, K.C. Do Fruit Reflectance Properties Affect Avian Frugivory in New Zealand? New Zealand J. Bot. 2022, 60, 319–329. [Google Scholar] [CrossRef]
- Willson, M.; Graff, D.; Whelan, C. Color Preferences of Frugivorous Birds in Relation to the Colors of Fleshy Fruits. Condor 1990, 92, 545–555. [Google Scholar] [CrossRef]
- Arruda, R.; Rodrigues, D.J.; Izzo, T.J. Rapid Assessment of Fruit-Color Selection by Birds Using Artificial Fruits at Local Scale in Central Amazonia. Acta Amaz. 2008, 38, 291–296. [Google Scholar] [CrossRef]
- Spironello, W.R.; de Tarso Barbosa Sampaio, P.; Ronchi-Teles, B. Produção e predação de frutos em Aniba rosaeodora Ducke var. amazonica Ducke (Lauraceae) em sistema de plantio sob floresta de terra firme na Amazônia Central. Acta Bot. Bras. 2004, 18, 801–807. [Google Scholar] [CrossRef]
- Lomáscolo, S.B.; Speranza, P.; Kimball, R.T. Correlated Evolution of Fig Size and Color Supports the Dispersal Syndromes Hypothesis. Oecologia 2008, 156, 783–796. [Google Scholar] [CrossRef]
- Beauchamp, G. Exploring the Role of Vision in Social Foraging: What Happens to Group Size, Vigilance, Spacing, Aggression and Habitat Use in Birds and Mammals That Forage at Night? Biol. Rev. 2007, 82, 511–525. [Google Scholar] [CrossRef]
- Thompson, C.L.; Bottenberg, K.N.; Lantz, A.W.; de Oliveira, M.A.B.; Melo, L.C.O.; Vinyard, C.J. What Smells? Developing in-Field Methods to Characterize the Chemical Composition of Wild Mammalian Scent Cues. Ecol. Evol. 2020, 10, 4691–4701. [Google Scholar] [CrossRef]
- Flörchinger, M.; Braun, J.; Böhning-Gaese, K.; Schaefer, H.M. Fruit Size, Crop Mass, and Plant Height Explain Differential Fruit Choice of Primates and Birds. Oecologia 2010, 164, 151–161. [Google Scholar] [CrossRef]
- García, D.; Chacoff, N.P. Scale-Dependent Effects of Habitat Fragmentation on Hawthorn Pollination, Frugivory, and Seed Predation. Conserv. Biol. 2007, 21, 400–411. [Google Scholar] [CrossRef]
- Lehouck, V.; Spanhove, T.; Colson, L.; Adringa-Davis, A.; Cordeiro, N.; Lens, L. Habitat Disturbance Reduces Seed Dispersal of a Forest Interior Tree in a Fragmented African Cloud Forest. Oikos 2009, 118, 1023–1034. [Google Scholar] [CrossRef]
- Fahrig, L. Effects of Habitat Fragmentation on Biodiversity. Annu. Rev. Ecol. Evol. Syst. 2003, 34, 487–515. [Google Scholar] [CrossRef]
- Santos, A.S.; Cazetta, E.; Dodonov, P.; Faria, D.; Gaiotto, F.A. Landscape-Scale Deforestation Decreases Gene Flow Distance of a Keystone Tropical Palm, Euterpe Edulis Mart (Arecaceae). Ecol. Evol. 2016, 6, 6586–6598. [Google Scholar] [CrossRef]
- Atwood, T.B.; Valentine, S.A.; Hammill, E.; McCauley, D.J.; Madin, E.M.P.; Beard, K.H.; Pearse, W.D. Herbivores at the Highest Risk of Extinction among Mammals, Birds, and Reptiles. Sci. Adv. 2020, 6, eabb8458. [Google Scholar] [CrossRef]
- Gagetti, B.L.; Piratelli, A.J.; Piña-Rodrigues, F.C.M. Fruit Color Preference by Birds and Applications to Ecological Restoration. Braz. J. Biol. Rev. Brasleira Biol. 2016, 76, 955–966. [Google Scholar] [CrossRef]
- Watson, J.E.M.; Whittaker, R.J.; Dawson, T.P. Habitat Structure and Proximity to Forest Edge Affect the Abundance and Distribution of Forest-Dependent Birds in Tropical Coastal Forests of Southeastern Madagascar. Biol. Conserv. 2004, 120, 311–327. [Google Scholar] [CrossRef]
- Tanalgo, K.C.; Pineda, J.A.F.; Agravante, M.E.; Amerol, Z.M. Bird Diversity and Structure in Different Land-Use Types in Lowland South-Central Mindanao, Philippines. Trop. Life Sci. Res. 2015, 26, 85–103. [Google Scholar]
- Chaves, P.P.; Timóteo, S.; Gomes, S.; Rainho, A. Response of Avian and Mammal Seed Dispersal Networks to Human-Induced Forest Edges in a Sub-Humid Tropical Forest. J. Trop. Ecol. 2022, 38, 199–209. [Google Scholar] [CrossRef]
- Española, C.P.; Collar, N.J.; Mallari, N.A.D.; Marsden, S.J. Large Avian Frugivores in the Philippines Show Linear Responses to Improvements in Forest Quality. For. Ecol. Manag. 2016, 375, 127–133. [Google Scholar] [CrossRef]
- Rey, P.J.; Alcántara, J.M. Effects of Habitat Alteration on the Effectiveness of Plant-Avian Seed Dispersal Mutualisms: Consequences for Plant Regeneration. Perspect. Plant Ecol. Evol. Syst. 2014, 16, 21–31. [Google Scholar] [CrossRef]
- Li, W.; Zhu, C.; Grass, I.; Vázquez, D.P.; Wang, D.; Zhao, Y.; Zeng, D.; Kang, Y.; Ding, P.; Si, X. Plant-Frugivore Network Simplification under Habitat Fragmentation Leaves a Small Core of Interacting Generalists. Commun. Biol. 2022, 5, 1–10. [Google Scholar] [CrossRef]
- Cornell, H.V.; Harrison, S.P. What Are Species Pools and When Are They Important? Annu. Rev. Ecol. Evol. Syst. 2014, 45, 45–67. [Google Scholar] [CrossRef]
- Séguigne, M.; Coutant, O.; Bouton, B.; Picart, L.; Guilbert, É.; Forget, P.-M. Arboreal Camera Trap Reveals the Frequent Occurrence of a Frugivore-Carnivore in Neotropical Nutmeg Trees. Sci. Rep. 2022, 12, 7513. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.; Li, W.; Gregory, T.; Wang, D.; Ren, P.; Zeng, D.; Kang, Y.; Ding, P.; Si, X. Arboreal Camera Trapping: A Reliable Tool to Monitor Plant-Frugivore Interactions in the Trees on Large Scales. Remote Sens. Ecol. Conserv. 2022, 8, 92–104. [Google Scholar] [CrossRef]
- Azhar, B.; Puan, C.L.; Aziz, N.; Sainuddin, M.; Adila, N.; Samsuddin, S.; Asmah, S.; Syafiq, M.; Razak, S.A.; Hafizuddin, A.; et al. Effects of in Situ Habitat Quality and Landscape Characteristics in the Oil Palm Agricultural Matrix on Tropical Understory Birds, Fruit Bats and Butterflies. Biodivers. Conserv. 2015, 24, 3125–3144. [Google Scholar] [CrossRef]
- Agduma, A.R.; Achondo, M.J.M.M.; Bretana, B.L.P.; Bello, V.P.; Remollo, L.L.; Mancao, L.S.; Supremo, J.P.; Salem, J.G.C.; Salvaña, F.R.P. Diversity of Vascular Plant Species in an Agroforest: The Case of a Rubber (Hevea brasiliensis) Plantation in Makilala, North Cotabato. Philipp. J. Crop Sci. 2011, 36, 57–64. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Models | Terms | AICc | ΔAICc | RL | wAICDc |
|---|---|---|---|---|---|
| 1 | Color + Predator + Color × Predator + DBH + Canopy + Tree cover | 747.160 | 0.510 | 0.775 | 0.510 |
| 2 | Color + Predator + Color × Predator + DBH + Canopy | 747.760 | 1.110 | 0.574 | 0.378 |
| 3 | Predator + DBH + Canopy | 765.100 | 18.450 | 0.000 | 0.000 |
| 4 | Predator +Color + Predator × Color | 747.060 | 0.410 | 0.815 | 0.536 |
| 5 | DBH + Canopy + Tree Cover | 808.780 | 62.130 | 0.000 | 0.000 |
| 6 | Color + Predator + Color × Predator + Tree Cover | 746.650 | 0.000 | 1.000 | 0.658 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Balasa, N.K.B.; Roquero, J.O.; Lidasan, A.K.; Casim, L.F.; Agduma, A.R.; Tanalgo, K.C. Fruit Color Preference of Frugivorous Birds in an Agroecosystem in Southcentral Mindanao, Philippines. Birds 2023, 4, 190-201. https://doi.org/10.3390/birds4020016
Balasa NKB, Roquero JO, Lidasan AK, Casim LF, Agduma AR, Tanalgo KC. Fruit Color Preference of Frugivorous Birds in an Agroecosystem in Southcentral Mindanao, Philippines. Birds. 2023; 4(2):190-201. https://doi.org/10.3390/birds4020016
Chicago/Turabian StyleBalasa, Navel Kyla B., Jirriza O. Roquero, Asraf K. Lidasan, Lothy F. Casim, Angelo Rellama Agduma, and Krizler Cejuela Tanalgo. 2023. "Fruit Color Preference of Frugivorous Birds in an Agroecosystem in Southcentral Mindanao, Philippines" Birds 4, no. 2: 190-201. https://doi.org/10.3390/birds4020016