
Arabian Partridge Abundance and Limiting Factors at the Northern Boundary of Its Range
by
 ,
,
Alaaeldin Soultan
1,*,
Moayyed Sher Shah
1,2,
Ahmed Mohammed Almalki
1,2,
Stephen Browne
1 and
Nico Arcilla
3,4 1
The Royal Commission for AlUla, Al Safarat, Riyadh 12512, Saudi Arabia
2
King Salman Royal Nature Reserve, Olaya Towers, Riyadh 12213, Saudi Arabia
3
International Bird Conservation Partnership, Storövägen 13, 14142 Huddinge, Sweden
4
Center for Great Plains Studies, University of Nebraska-Lincoln, 1155 Q St., Lincoln, NE 68508, USA
*
Author to whom correspondence should be addressed.
Birds 2023, 4(2), 179-189; https://doi.org/10.3390/birds4020015
Submission received: 27 February 2023
/
Revised: 14 April 2023
/
Accepted: 17 April 2023
/
Published: 20 April 2023
(This article belongs to the Special Issue Feature Papers of Birds 2022–2023)
Abstract
:Simple Summary
Using hierarchical distance-sampling models, we estimated the population size of the endemic Arabian Partridge at the northern boundary of its range in Saudi Arabia and the factors that influence density and distribution. Overall density in Harrat Uwayrid Biosphere Reserve was estimated at 25.6 (6.16 SE) birds/km2 and abundance at ~118 individuals. We found significant decreases in Arabian Partridge abundance in response to habitat disturbances, including grazing, hunting, and feral donkeys, the latter of which were present at 70% of sites surveyed. We recommend establishing a monitoring program to protect the reserve’s wildlife and habitat from livestock grazing and hunting and a control program to remove feral donkeys. Additionally, we recommend developing a monitoring program for the Arabian Partridge and other endemic species in the reserve and throughout their range and investigating the impact of grazing practices and feral donkeys on the Arabian Partridge and other native species in the region to inform conservation and management plans.
Abstract
The Arabian Partridge (Alectoris melanocephala), is an endemic bird species of the Arabian Desert that lives in groups and inhabits rocky hillsides with patchy vegetation. Throughout their range, Arabian Partridges contend with hunting and habitat destruction, factors that may limit their distribution and abundance. Although the abundance of this species has been assumed to be stable, no actual estimate of its population size has been undertaken. We assessed the distribution and estimated the abundance of the Arabian Partridge at the northern boundary of its range in Saudi Arabia. The estimated density and abundance of the Arabian Partridge in Harrat Uwayrid Biosphere Reserve was 25.6 (6.16 SE) birds/km2 and ~118 individuals, respectively, with higher numbers of individuals in less disturbed sites and near rocky outcrops and hillsides. In sites where hunting occurred, as indicated by the presence of hunting shelters, partridge numbers were extremely low or absent. Our study provides the first quantitative assessment of the Arabian Partridge at the northern limit of its range and highlights the need to reduce threats from hunting, livestock grazing, and feral donkeys and to undertake conservation measures to mitigate factors associated with partridge decline.

1. Introduction
Located in the southwest corner of Asia, the Arabian Desert forms a land bridge between Asia, Europe, and Africa that has allowed for the exchange of biodiversity between continents [1] and resulted in a distinctive wildlife community that is highly adapted to extreme conditions [2]. Fauna and flora in this region comprise many rare, threatened, and endemic species that have unique ecological roles, such as the regionally endangered Eurasian Griffon (Gyps fulvus), and endemic bird species, such as the Arabian Wheatear (Oenanthe lugentoides) and the Arabian Green Bee-eater (Merops cyanophrys). However, due to the challenges inherent in accessing desert terrain and this region’s distinctive harsh climate and topography [3,4], many Arabian Desert species tend to receive relatively little study and conservation attention [5].
Partridges are among those species adapted to harsh climatic and environmental conditions that successfully breed in the Arabian Desert [6]. Robust birds with strong bills and legs, partridges often forage in family or social groups (coveys), especially outside the breeding season, and both sexes have similar plumage [6]. Among the seven partridge species in the Arabian Desert, two are endemic: the Arabian Partridge (Alectoris melanocephala) and Philby’s Partridge (Alectoris philbyi) [6]. Compared to Philby’s Partridge, the Arabian Partridge has a wider range that expands over the eastern and southwestern fringe of the Arabian Peninsula with a designated northern distributional limit at 26° N [6]. Occurring in highlands and rocky slopes in southern Arabia, the Arabian Partridge is found at altitudes from 250–2800 m [7,8].
The global population of Arabian Partridge is believed to number 400,000 pairs [6], although no monitoring program or data-based population estimate has been undertaken. No specific potential threats to Arabian Partridge have been quantified, but this species is considered likely to be affected by habitat degradation and loss caused by livestock grazing, conversion to agricultural land, and drought [9]. In the northern Arabian Desert, declines in congeneric Chukar (A. chuckar) populations are attributed to predation pressure on eggs, chicks, and adults by canid species, such as the golden jackal (Canis aureus) and the red fox (Vulpes vulpes) [10], as well as livestock overgrazing that degrades and destroys their habitat and nesting sites [11]. Although Arabian Partridges are targeted by hunters and egg collectors, the effects of these activities on their populations have not been quantified [9]. The hunting of gamebirds is known to be unsustainable above particular thresholds, such as when it causes mortality of over 15% of a breeding population that produces 2–5 chicks per pair [12]. Partridges may also be targeted by predators such as the African wildcat (Felis lybica), Arabian wolf (Canis lupus), and feral dogs (Canis familiaris).
Recently, the Royal Commission for AlUla (RCU) established the Harrat Uwayrid Biosphere Reserve (HUBR) in northwestern Saudi Arabia [13], which includes the northernmost distribution of the Arabian Partridge. Included in the HUBR mandate is the prohibition of hunting and the development of conservation and/or reintroduction plans for threatened and endemic species, including the Arabian Partridge. While Arabian Partridge numbers have been assumed to be stable throughout their range [9], accurate information about the population trends and conservation status of the Arabian Partridge is not available as there is no monitoring program for this or many other endemic species in this region.
Here, our objectives were to (1) estimate, for the first time, the population size of the Arabian Partridge within the HUBR in Saudi Arabia and (2) assess factors that might influence its distribution, such as landscape features, vegetation cover, and disturbances, including livestock grazing, hunting, and the presence of feral animals, such as donkeys and dogs in the reserve.
We hypothesized that the abundance of the Arabian Partridge would depend on the landscape, topography, and disturbance factors. We predicted that Arabian Partridge numbers would be higher in sites dominated by rocky outcrops and slopes that provide partridges with cover and camouflage. Additionally, we predicted that the presence of livestock grazing and feral animals would adversely affect the abundance of the Arabian Partridge. We tested our hypotheses by modeling the abundance of the Arabian Partridge as a function of landscape- and disturbance-related variables using a hierarchical distance sampling approach.
2. Materials and Methods
2.1. Study Species
The Arabian Partridge is the largest species of its genus [6]. Pale gray in color, it has a black crown that stretches back to its nape, a broad white supercilium and throat, and gray tail feathers that are visible in flight, characteristics that visually distinguish it from its sister species [9,14]. The Arabian Partridge is also phylogenetically distant from other species of Alectoris genus [8]. Males and females have similar plumage, but males (~724 g) are almost 40% larger than females (~522 g) [6,15]. The breeding ecology of the Arabian Partridge is poorly understood, especially in the wild, but it is believed to be a monogamous species [15]. Breeding starts by pair formation in February–March, when males become more vocal and territorial [6]. Females lay eggs between March and May in a scoop nest on the ground, with 5–8 eggs, and chicks have been recorded between April and November, but mainly from June to September [6,15]. Pairs may produce two broods per year in some areas, and in some cases, males and females may each incubate separate broods, with chicks congregatingpost-hatching [6]. Arabian Partridges are gregarious and typically found in coveys of 10–15 individuals, although group numbers as high as 27 have been reported [6]. Their population structure and social behavior are poorly understood [6,15]. The Arabian Partridge is found at high altitudes of up to 2800 m, where it forages on seeds, fresh leaves, and invertebrates, and it does not require open water for drinking, as it obtains its water requirements from food [6,9].
2.2. Study Area
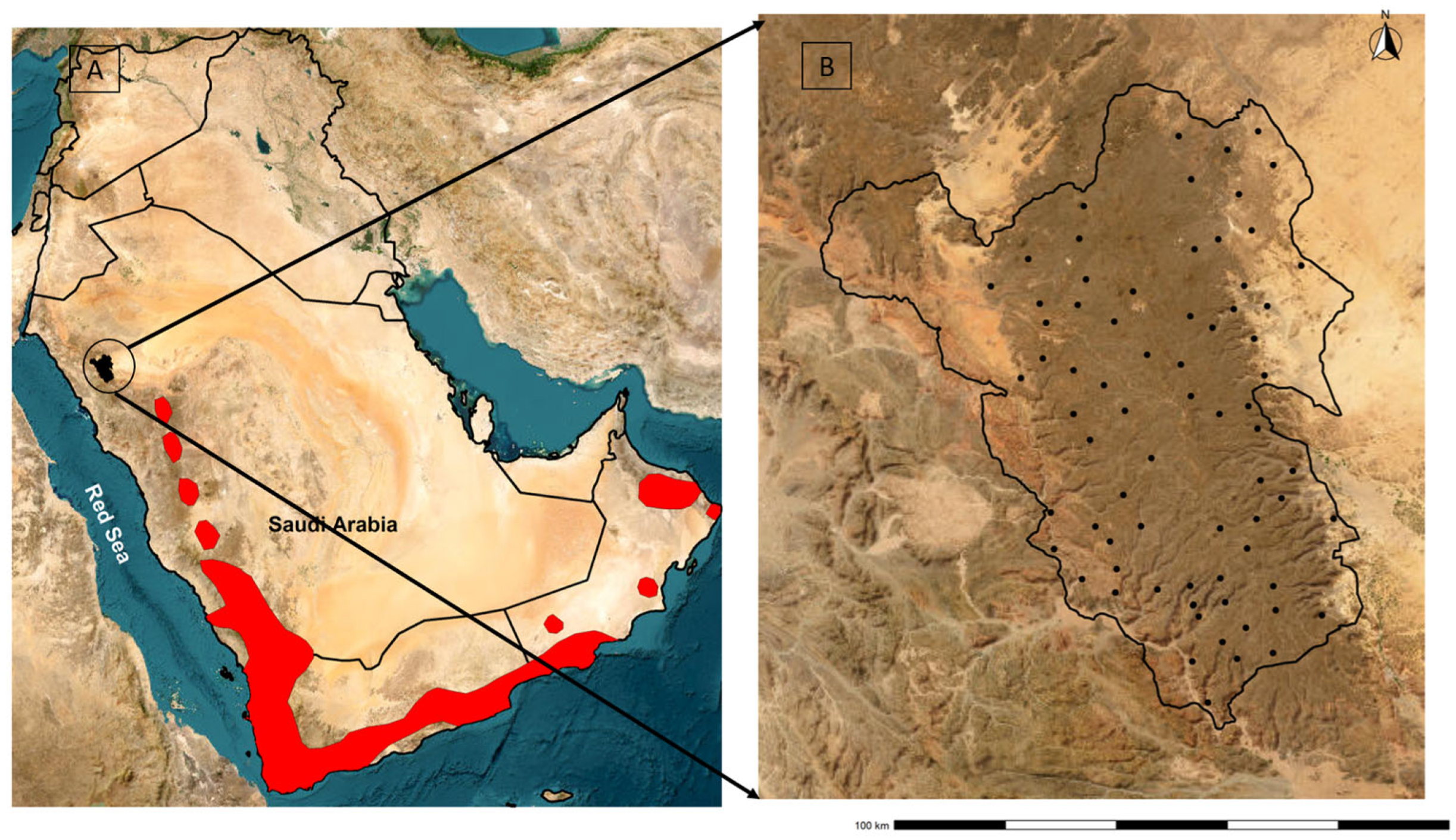
A recently declared UNESCO site, the Harrat Uwayrid Biosphere Reserve (HUBR), is located in the northwest of Saudi Arabia (27.014417° N, 37.517526° E). The largest of five nature reserves in Saudi Arabia’s AlUla region, the HUBR covers 4680 km2 and is characterized by highly variable topography (550–1830 m.a.s.l.) (Figure 1). The HUBR is the first non-marine biosphere reserve in the region [13], and forms part of the Hejaz highlands with a lava-covered sandstone plateau. Approximately 5000 people inhabit the buffer zone and transition areas of HUBR, where they practice traditional farming and livestock grazing. HUBR is located in the most arid region of the Arabian Peninsula, with a maximum precipitation of 29 mm in January [16]. Temperatures range from 45 °C in mid-summer to 14 °C in mid-winter [17]. Part of the Saharo-Arabian region with sparsely vegetated areas [18], HUBR hosts at least 55 higher plant species, with dominant vegetation including Haloxylon salicornicum, Retama raetam, and Acacia woodland. In addition to species already mentioned, the HUBR hosts the regionally vulnerable Striped hyena (Hyaena hyaena), regionally vulnerable Blandford’s fox (Vulpis cana), regional endemic Arabian wolf (Canis lupus arabs), the regionally vulnerable Arabian gazelle (Gazella arabica), and the Nubian ibex (Capra nubiana).
2.3. Field Data Collection
We conducted field surveys for Arabian Partridge between May and September 2022 on 73 line transects distributed across the HUBR. Transects were distributed based on conditional random sampling and were located a minimum of 5 km away from human settlements and 3 km away from the nearest transect in order to avoid pseudo-replication and spatial autocorrelation and ensure that our observations were independent [19,20]. Selected transects were divided among three observers with long-standing experience in desert bird identification. We carried out fieldwork during periods when we expected partridges to be most active, namely after dawn (07:00–09:30) and before dusk (16:00–18:30). Each transect had a predefined length of 1 km and a width of 140 m (70 m on each side). The transect width was determined by the average width of valleys in the study area. We conducted surveys under suitable weather conditions (i.e., not during dust storms or other adverse weather events) to avoid weather effects on partridge activity and detectability. Each transect was traveled by a single observer, with an average speed of 3.5 km/h, who measured the perpendicular distance from their positions along the transect to Arabian Partridges encountered, recording numbers of individuals (i.e., covey/group size). For each transect, landscape covariates that might influence the occurrence and the abundance of the Arabian Partridge were recorded during the survey, including (1) vegetation cover; (2) tree cover; (3) distance to the nearest hillside; (4) wadi (valley) width; (5) distance to vehicle tracks; and (6) disturbance level.
In sparsely vegetated sites, Arabian Partridges may be more easily detected compared to densely vegetated sites. We therefore measured vegetation cover on an ordinal scale in three categories (low, medium, and high). Low vegetation cover was defined as ≤10% of the area of the transect, medium cover as >10% and ≤30%, and high cover as >30% of the area of the transect. Arabian Partridges use trees for roosting and foraging [6], so we expected that tree cover might influence their density and abundance. We documented tree cover by recording the total number of trees (woody plants > 1 m height) in the transect area. In addition to open plains with good vegetation cover, Arabian Partridges are found in rocky habitats [6,15], and we thus measured the Euclidean distance, in meters, between birds encountered and the nearest rocky outcrop or hillside. We also measured wadi (valley) width to allow for an examination of its influence on Arabian Partridge density and abundance; wadi width was defined as the width in meters of the flat area between sloped hillsides on either side.
Site disturbance either by people (e.g., hunting, driving) and domestic or feral animals (e.g., livestock, feral donkeys) significantly affected the density and detectability of Arabian Partridges. We therefore quantified disturbances through two variables: distance to vehicle tracks and site disturbance level. Distance to vehicle tracks was measured as the Euclidean distance between the bird encountered and the nearest vehicle track. All distance measurements were recorded using a laser rangefinder (Bushnell Tour V5). Site disturbance levels were quantified on an ordinal scale with “0” representing no disturbance and “10” representing a high disturbance level (e.g., site overgrazed, both feral donkeys and dogs were recorded, and evidence of hunting was recorded). Disturbances resulted from (1) grazing by livestock; (2) the presence of feral donkeys; (3) the presence of feral dogs; and (4) hunting (as evidenced by, e.g., the presence of bullet cartridges, gunshots, and hunting shelters) and/or camping (e.g., presence of firewood and/or tree cutting) within the transect area. Grazing intensity and hunting evidence were assessed on a scale of four categories, while the apparent presence or absence of feral donkeys and dogs was measured on a binary scale. The scale of grazing was (“0” = no grazing, “1” ≤ 10 grazing animals, “2” ≤ 30 grazing animals, and “3” > 30 grazing animals). The scale for hunting was “0” = no evidence of hunting, “1” = a single indication of hunting (such as a shotgun shell), “2” = two indications of hunting (such as two shotgun shells), and “3” more than two indications of hunting (such as a hunting blind and multiple shotgun shells). The presence or absence of feral donkeys was indicated as “1” or “0”, respectively. The values of these factors were thus used to describe disturbance presence and levels for each transect.
2.4. Statistical Analyses
We tested for correlations between continuous variables (covey size, wadi width, tree cover, distance to vehicle tracks, and distance to nearest hillside). We found a negative correlation (r = −0.8) between the distance to the vehicle track and the distance to the nearest hillside and a positive correlation (r = 0.77) between covey size and distance to vehicle tracks. We therefore avoided combining these correlated variables in the same model.
To estimate Arabian Partridge density, we applied a hierarchical distance-sampling model, following the framework developed by Royle et al. [21] and using the “distsamp” function implemented in “unmarked” R package [22]. This hierarchical approach allows for modeling species density as well as detection probability [21,22,23] and allows for modeling density in response to covariates [23]. We grouped observation data from line transect surveys into 10 m discrete intervals. Partridge detection probability was modeled using two key functions, the half-normal and hazard rates, respectively [22]. In order to identify variables associated with partridge detection probability and density (individual/km2), we modeled partridge detection probability as a function of five variables, including three landscape-related covariates (vegetation cover, tree cover, and wadi width) and two detection-related covariates (observer and covey size). We modeled partridge density as a function of landscape variables and scaled all continuous variables before running models to facilitate model convergence [24]. Our sampling period included the breeding period (May–June) and the main post-breeding period when chicks appear (July–September). To account for temporal variability in species density and abundance, we included season as a binary variable while modeling the density and the abundance of the Arabian Partridge, with a value of “1” indicating that sampling covered the breeding period and a value of “2” indicating that sampling covered the post-breeding period.
We ran all possible model combinations, from the null model, with no covariates incorporated, to the full model, with additive and interaction terms, including linear and quadratic terms. Goodness-of-fit for the global model [25] was evaluated using the “gof ds” function implemented in “Distance” R package [26]. We used Akaike’s information criterion (AIC) to select the best model (lowest AIC) among the rival models [25,27]. Finally, we used a t-test to compare the estimated density of the Arabian Partridge in sites where feral donkeys were recorded to those where they were not recorded. All analyses were performed using the R environment [28].
3. Results
We recorded a total of 91 Arabian Partridges (naïve abundance) with mean and maximum covey sizes of 2.1 and 4, respectively. We found evidence of hunting in almost 25% of sites surveyed and evidence of livestock grazing and the presence of feral donkeys in nearly 70% of surveyed sites. We ran a total of 12 models to investigate Arabian Partridge detection probability and population density (Table S1).
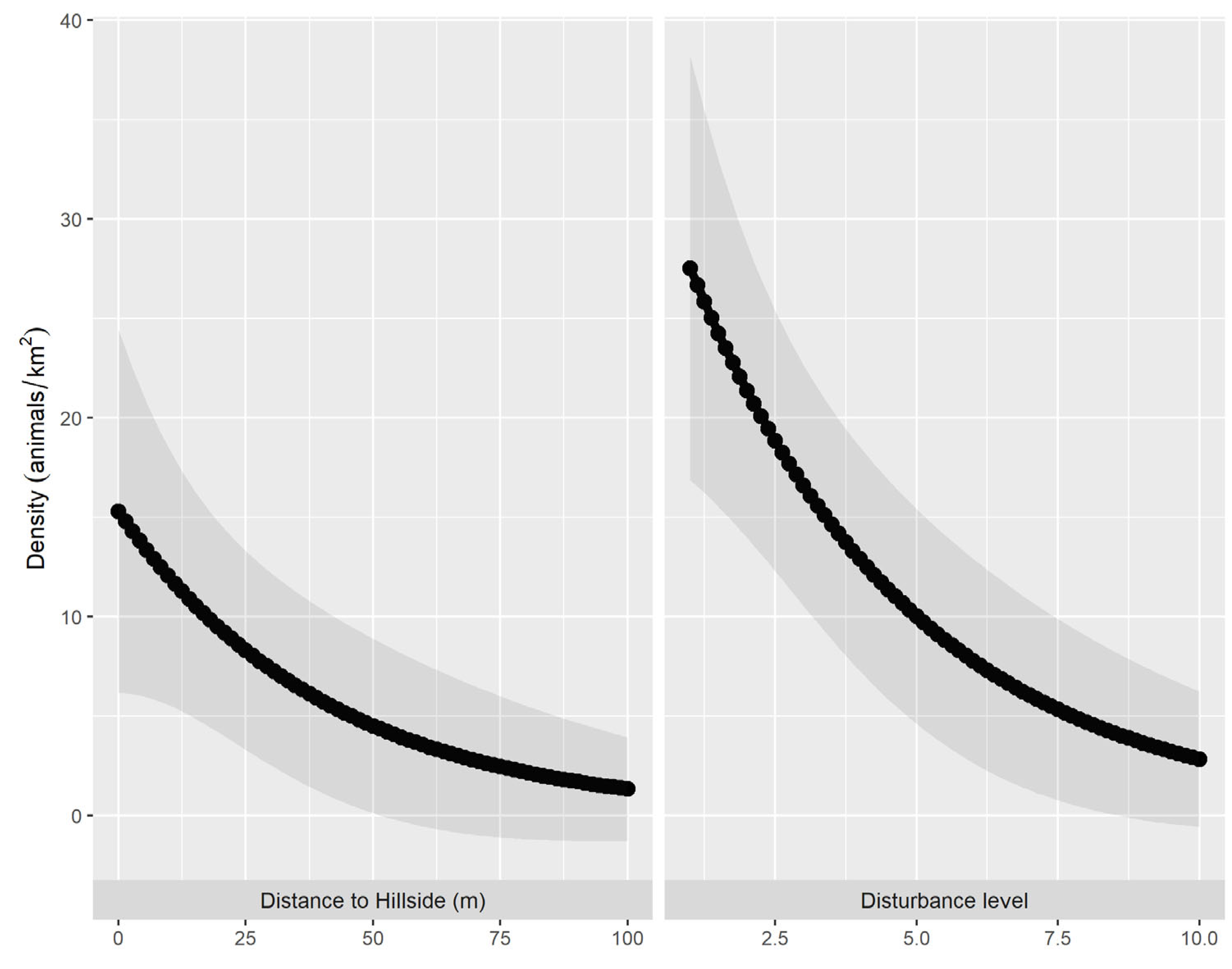
The distance sampling model that best fit of line the data, with an AIC value of 239.06, modeled detection probability as a function of covey size, observer, and vegetation cover. Density was modeled as a function of wadi width, including a quadratic term, and covariates of tree cover, disturbance, and distance to the nearest hillside. We estimated the Arabian Partridge’s overall density as 25.6 (6.16 SE) birds/km2 and abundance in the surveyed area as ~118 individuals. Both covey size and the vegetation cover significantly influenced Arabian Partridge detection probability (Table 1). Our model also demonstrated a strong negative association between the Arabian Partridge density and habitat disturbance levels (Figure 2). The second-best model according to the AIC value included seasons as a factor in order to explain the estimated density and abundance of the Arabian Partridge (Table S1). Meanwhile, there was an insignificant change in the estimated density over the seasons (breeding and post-breeding). The t-test showed a significant difference (t = 9.32, df = 144, p < 0.0001) between the mean estimated density in sites where feral donkeys were present (0.50 birds/km2) and sites where feral donkeys were absent (11.57 birds/km2) (Figure S1).
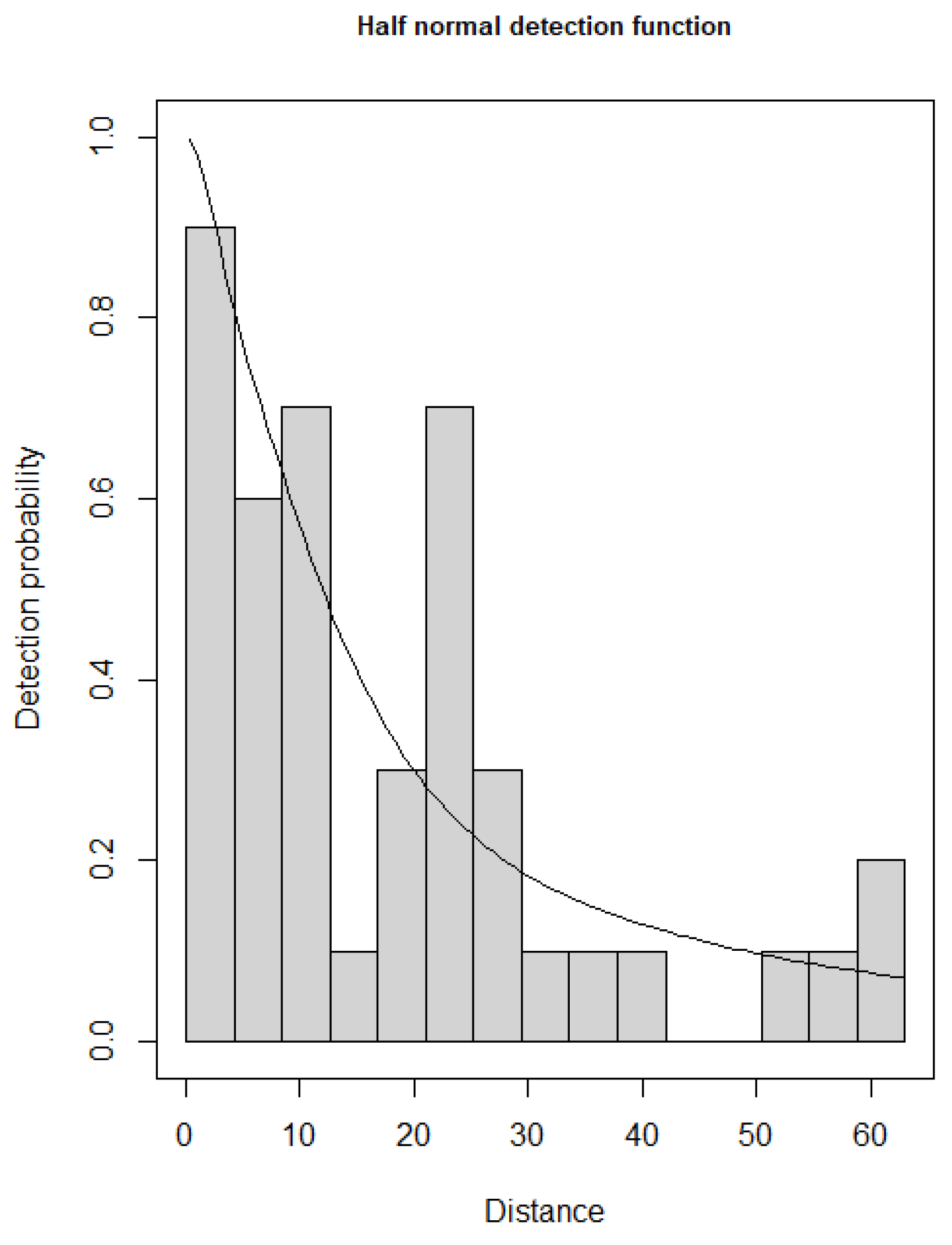
According to the best fit (lowest AIC) model, the detection function was modeled using the half-normal key function (Table 1 and Figure 3). The variation in the detection probability was explained by covey size and the vegetation cover covariates and displayed a good fit (Goodness-of-fit test; χ2 = 0.15, p = 0.37). The detection probability estimated was 0.29 (0.78 SE) with an effective strip width of 18 m (Figure 3).
4. Discussion
This study appears to be the first field-based attempt to estimate a population size for the Arabian Partridge, a species on which previous work has been very limited and focused mostly on phylogeny [8,29]. Our findings show that the current Arabian Partridge population in the HUBR is ~118 individuals, with an estimated overall density of 25.6 (6.16 SE) birds/km2, which was strongly influenced by both disturbance and landscape features. This estimation is prone to yearly fluctuation due to variability in environmental conditions, such as precipitation, which can strongly influence birds’ food availability. For example, lower precipitation levels have been shown to lead to energy stress on related species, such as Chukars and Sand Partridges, in some cases provoking them to leave the area and migrate elsewhere [30]. In our study area, we found that partridge numbers strongly declined in response to disturbances, including livestock grazing, the presence of feral donkeys, and the presence of hunters. We also found a high level of disturbance, with ~70% of the surveyed sites grazed either by livestock (goats, sheep, and/or camels) or feral donkeys and ~25% of sites showing evidence of hunting.
Moderate levels of disturbance resulted in a ~50% decline in the numbers of Arabian Partridges compared to the sites with no disturbance. In this study, rather than assessing the contribution of each factor independently, we classified habitat disturbances into one variable instead of making separate variables for each disturbance type because many of these disturbance types were interrelated. However, our field observations indicated that the impact of feral donkeys on Arabian Partridges is significant and negative. Feral donkeys in the study area traveled in herds averaging 10 animals throughout the area, rather than along tracks or paths, distributing their impact throughout large areas. Previous studies have shown that livestock grazing may strongly affect birds, influencing their activity patterns, distribution, abundance, and/or nest success [31,32,33,34].
Increased grazing intensity may be associated with decreased bird densities, nest success, and recruitment [32,35]. In some respects, free-ranging domestic and feral livestock may partially fill ecological gaps left by the absence of wild grazers that have dwindled or gone extinct in this region (such as the wild oryx and the gazelle), but their effects on habitat and native wildlife may be detrimental or unexpected [32,36]. Feral donkey grazing patterns have been shown to shift the composition of desert vegetation [37,38], negatively influencing habitat structure and function. Feral donkeys may also drive native wildlife declines or extirpations through their interactions with wild carnivores [36] and contribute to habitat degradation, including soil erosion [39] the pollution of water sources and the prevention of wildlife from drinking [40]. Although free-ranging and feral dogs may also have significant negative impacts on wildlife [41], most of the feral dogs we observed during our surveys were either close to human settlements or associated with shepherds, with few free-ranging individuals occurring inside the desert. Thus, we did not find evidence that feral dogs have significant impacts on the Arabian Partridge in the HUBR. Nevertheless, feral dogs should be controlled inside the reserve, as dogs often chase and harass native animals they encounter, forcing them to engage in energetically costly behavior and causing ecological and physiological stress to native animals, driving wildlife decline [41,42,43,44].
Hunting was another source of disturbance in the HUBR. Although the prevalence of this practice was not high (~25% of the covered area), hunting is considered to be among the main threats to the wildlife in Saudi Arabia [1]. For instance, MacQueen’s Bustard (Chlamydotis macqueenii), a gamebird species that once was common in the region, has been extirpated due to the overhunting and prospects regarding the recovery of this species are uncertain [1]. Moreover, previous research has indicated that in a nearby region (Tabuk province), wild-caught Arabian Partridges are among the animals sold at an illegal wildlife market [45]. This implies that the Arabian Partridge is vulnerable to extirpation in this area if hunting is not regulated and controlled.
Landscape features, particularly hillsides, were positively associated with Arabian Partridge abundance. Hillsides provide Arabian Partridges with cover from threats, such as natural predators or hunters. The average Euclidian distance between the observed partridge and the nearest hillside was <10 m, reflecting the importance of this landscape feature for this species. During our surveys, Arabian Partridges typically fled for cover upon detecting people, indicating their fear and vigilance in response to threats or disturbances. The fate of the MacQueen’s Bustard among other species indicates the vulnerability of large-bodied terrestrial birds in this region to extirpation.
To protect the Arabian Partridge and wildlife with similar life histories from decline, we recommend several conservation actions. First, an efficient patrol program to mitigate and prevent disturbance and illegal hunting should be established, ideally using advanced technology, such as cybertracker [46] or the SMART system [47]. Second, a control program for feral animals, including feral donkeys and feral dogs, should be established. Removing feral animals would allow native species occupy additional areas with suitable habitat and decrease their physiological stress [48,49]. Third, we recommend implementing a follow-up program to monitor any changes in the population size of this species. Expanding surveys to cover a greater area of the Arabian Partridge’s range would alert reserve managers to priority sites for conservation and allow the assessment of any changes in Arabian Partridge distribution.
Previous studies have shown that covey size can influence species detectability [50]. Thus, incorporating covey size into detection probability models is important to avoid overestimating density [50,51]. We found that incorporating covey size improved our model performance, and that partridge detectability improved with increasing covey size, due in part to their warning call vocalizations and audible wing flapping to signal to other group members that they feel threatened. Single individuals, on the other hand, may silently hide from potential observers, escaping detection. Experienced observers conducting surveys are less likely to record false absences for this and other reasons.
Previous work [6] reported that the northern distribution of the Arabian Partridge falls below 26° N. However, all our records of the Arabian Partridge were ~100 km north of its known distribution, indicating a northward expansion in the range of the Arabian Partridge. A northward shift has been recorded over the last few decades for many species, both in this region [5] and outside this region [52]. Such movements have been attributed to ongoing changes in climatic conditions, which have been associated with many species shifting either higher in elevation or northward in an apparent effort to adapt to changing climatic conditions [52,53,54]. The impacts of climate change over the last few decades, resulting in warmer temperatures in this region, were not incorporated into our model but might be at least partially responsible for the northward shift in Arabian Partridge distribution.
5. Conclusions
In this study, we estimated for the first time the population size of the Arabian Partridge at its northern limit inside Harrat Uwayrid, the first inland human and biosphere reserve in Saudi Arabia. Our findings indicate a northward expansion in the distribution of this species beyond its previously known range. We also investigated factors that influence the abundance of this species, as a first step towards developing conservation and management plans. We advocate further investigations into the ecological impact of feral animals on the native fauna and flora of this region, as well as other factors that may limit Arabian Partridge populations across its entire range of this species, and its response to climate change. We recommend conservation actions to maintain Arabian Partridge populations, including (1) conducting ranger patrols to prevent activities prohibited in the reserve, including hunting and livestock grazing; (2) establishing a control program for the removal of feral donkeys, dogs, and other non-native animals to mitigate and prevent their destruction of native habitat and wildlife; and (3) monitoring changes in the population size of this and other wildlife species in response to ongoing threats and conservation actions. Conservation actions targeting Arabian Partridges are expected to provide benefits for other native wildlife in this area, including endangered, rare, and endemic species in the Arabian Desert.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/birds4020015/s1, Figure S1: Boxplot showing the estimated density of the Arabian Partridge in sites where feral donkeys were not recorded (absent) and in sites where feral recorded (present) in Harrat Uwayrid Biosphere Reserve, Saudi Arabia. Black lines in boxes represent median values of the estimated density, while the lower and upper box edges represent the 25th and 75th percentile of the density, respectively.; Table S1: A list of all converged models ranked according to their AIC. The model formula is a double right-hand formula describing detection covariates followed by density covariates. ~1 ~1 would be a null model. ΔAIC is the difference in AIC compared to the lowest AIC (the best model).
Author Contributions
Conceptualization, A.S.; methodology, A.S. and M.S.S.; software, A.S.; validation, A.S., M.S.S., S.B. and A.M.A.; formal analysis, A.S.; investigation, A.S. and M.S.S.; resources, A.S. and N.A.; data curation, A.S.; writing—original draft preparation, A.S. and N.A.; writing—review and editing, A.S., M.S.S., A.M.A., S.B. and N.A.; visualization, A.S.; supervision, A.S.; project administration, A.M.A. and S.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Primary data not presented in this study are available on request from the corresponding author.
Acknowledgments
The authors wish to acknowledge their RCU colleagues for facilitating the fieldwork and providing the authors with a detailed description of the site’s accessibility.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Alatawi, A.S. Conservation Action in Saudi Arabia: Challenges and Opportunities. Saudi J. Biol. Sci. 2022, 29, 3466–3472. [Google Scholar] [CrossRef] [PubMed]
- Metallinou, M.; Arnold, E.N.; Crochet, P.-A.; Geniez, P.; Brito, J.C.; Lymberakis, P.; Baha El Din, S.; Sindaco, R.; Robinson, M.; Carranza, S.; et al. Conquering the Sahara and Arabian Deserts: Systematics and Biogeography of Stenodactylus Geckos (Reptilia: Gekkonidae). BMC Evol. Biol. 2012, 12, 258. [Google Scholar] [CrossRef] [PubMed]
- AbuZinada, A.H.; Robinson, E.R.; Nader, I.A.; Al Wetaid, Y.I. First Saudi Arabian National Report on the Convention on Biological Diversity; The National Commission for Wildlife Conservation and Development: Riyadh, Saudi Arabia, 2004. [Google Scholar]
- Vincent, P. Saudi Arabia: An Environmental Overview; CRC Press: Boca Raton, FL, USA, 2008; ISBN 9780429216619. [Google Scholar]
- Soultan, A.; Wikelski, M.; Safi, K. Risk of Biodiversity Collapse under Climate Change in the Afro-Arabian Region. Sci. Rep. 2019, 9, 955. [Google Scholar] [CrossRef] [PubMed]
- Jennings, M.C. Atlas of the Breeding Birds of Arabia. Fauna Arab. 2010, 25, 1–751. [Google Scholar]
- BirdLife International Species Factsheet: Alectoris melanocephala. Available online: http://www.birdlife.org (accessed on 11 December 2022).
- Khan, H.A.; Arif, I.A.; Shobrak, M. DNA Barcodes of Arabian Partridge and Philby’s Rock Partridge: Implications for Phylogeny and Species Identification. Evol. Bioinforma. 2010, 2010, 151–158. [Google Scholar] [CrossRef]
- McGowan, P.J.K.; Kirwan, G.M. Arabian Partridge (Alectoris melanocephala), version 1.0. In Birds of the World; del Hoyo, J., Elliott, A., Sargatal, J., Christie, D., de Juana, E., Eds.; Cornell Lab of Ornithology: Ithaca, NY, USA, 2020. [Google Scholar]
- Yom-Tov, Y.; Benjamini, Y.; Kark, S. Global Warming, Bergmann’s Rule and Body Mass–Are They Related? The Chukar Partridge (Alectoris chukar) Case. J. Zool. 2002, 257, 449–455. [Google Scholar] [CrossRef]
- Gruychev, G.; Dyakov, N.; Dimitrov, D. Habitat Variables Influencing Chukar Partridge Decrease in Southeastern Bulgaria. Folia Zool. 2014, 63, 171–179. [Google Scholar] [CrossRef]
- Sandercock, B.K.; Nilsen, E.B.; Brøseth, H.; Pedersen, H.C. Is Hunting Mortality Additive or Compensatory to Natural Mortality? Effects of Experimental Harvest on the Survival and Cause-Specific Mortality of Willow Ptarmigan. J. Anim. Ecol. 2011, 80, 244–258. [Google Scholar] [CrossRef]
- UNESCO Harrat Uwayrid Biosphere Reserve-Saudi Arabia. Available online: https://www.unesco.org/archives/multimedia/document-5783 (accessed on 21 January 2023).
- Hayes, L. Upland Game Birds: Their Breeding and Care; L.B. Hayes: Valley Center, CA, USA, 1995; ISBN 9780963319623. [Google Scholar]
- McGowan, P.; Madge, S.; D’Silva, C.; Cole, D.; Mead, D.; Cox, J.D.; Franklin, K.; Arlott, N.; Budden, R. Pheasants, Partridges & Grouse: Including Buttonquails, Sandgrouse and Allies; Helm Identification Guides; Bloomsbury Publishing: London, UK, 2010; ISBN 9781408135662. [Google Scholar]
- Obaid, W.A.; Boutraa, T.; Abdel-Hameed, U.K. Floristics and Soil Characteristics of Ohud Mountain, Al-Madinah Al-Munawarah, Western Saudi Arabia. J. Mt. Sci. 2020, 17, 2405–2417. [Google Scholar] [CrossRef]
- Climate Data. Available online: https://en.climate-data.org/ (accessed on 1 March 2023).
- Sayed, O.H.; Masrahi, Y.S. Climatology and Phytogeography of Saudi Arabia. A Review. Arid L. Res. Manag. 2023, 1–58. [Google Scholar] [CrossRef]
- Bataineh, A.L.; Oswald, B.P.; Bataineh, M.; Unger, D.; Hung, I.-K.; Scognamillo, D. Spatial Autocorrelation and Pseudoreplication in Fire Ecology. Fire Ecol. 2006, 2, 107–118. [Google Scholar] [CrossRef]
- Hurlbert, S.H. Pseudoreplication and the Design of Ecological Field Experiments. Ecol. Monogr. 1984, 54, 187–211. [Google Scholar] [CrossRef]
- Royle, J.A.; Dawson, D.K.; Bates, S. Modeling Abundance Eeffects in Distance Sampling. Ecology 2004, 85, 1591–1597. [Google Scholar] [CrossRef]
- Fiske, I.; Chandler, R. Unmarked: An R Package for Fitting Hierarchical Models of Wildlife Occurrence and Abundance. J. Stat. Softw. 2011, 43, 1–23. [Google Scholar] [CrossRef]
- Chandler, R. Distance Sampling Analysis in Unmarked; USGS Patuxent Wildlife Research Center: Laurel, MD, USA, 2019; pp. 1–8. [Google Scholar]
- Ortega-Álvarez, R.; Calderón-Parra, R.; Martínez Molina, U.; Martínez Molina, F.; Martínez Molina, G.; Martínez Molina, Y.; Martínez Villagrán, A.; Martínez Freire, J.D.; Vásquez Robles, R.; García Loaeza, D.; et al. How Many Are Left? Density and Population Size of the Endangered Sierra Madre Sparrow across Central Mexico. J. Ornithol. 2021, 162, 31–41. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach; Springer: New York, NY, USA, 2002. [Google Scholar]
- Miller, D.L.; Rexstad, E.; Thomas, L.; Marshall, L.; Laake, J.L. Distance Sampling in R. J. Stat. Softw. 2019, 89, 1–28. [Google Scholar] [CrossRef]
- Buckland, S.T.; Anderson, D.R.; Burnham, K.P.; Laake, J.L. Distance Sampling. In Encyclopedia of Biostatistics; John Wiley & Sons, Ltd.: Chichester, UK, 2005. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2016; Available online: https://www.R-project.org/ (accessed on 11 December 2022).
- Randi, E. A Mitochondrial Cytochrome B Phylogeny of the Alectoris Partridges. Mol. Phylogenet. Evol. 1996, 6, 214–227. [Google Scholar] [CrossRef]
- Kam, M.; Degen, A.A.; Nagy, K.A. Seasonal Energy, Water, and Food Consumption of Negev Chukars and Sand Partridges. Ecology 1987, 68, 1029–1037. [Google Scholar] [CrossRef]
- Martin, T.G.; Possingham, H.P. Predicting the Impact of Livestock Grazing on Birds Using Foraging Height Data. J. Appl. Ecol. 2005, 42, 400–408. [Google Scholar] [CrossRef]
- Kaplan, R.H.; Rosamond, K.M.; Goded, S.; Soultan, A.; Glass, A.; Kim, D.H.; Arcilla, N. Bobolink (Dolichonyx oryzivorus) Declines Follow Bison (Bison bison) Reintroduction on Private Conservation Grasslands. Animals 2021, 11, 2661. [Google Scholar] [CrossRef]
- Barzan, F.R.; Bellis, L.M.; Dardanelli, S. Livestock Grazing Constrains Bird Abundance and Species Richness: A Global Meta-Analysis. Basic Appl. Ecol. 2021, 56, 289–298. [Google Scholar] [CrossRef]
- Hua, J.; Lu, S.; Song, K.; Wang, J.; Wang, J.; Xu, J. Effects of Livestock Grazing on Spatio-Temporal Patterns and Behaviour of Reeves’s Pheasant Syrmaticus Reevesii. Animals 2022, 12, 2968. [Google Scholar] [CrossRef]
- Tadey, M. Livestock Indirectly Decrease Nest Abundance of Two Shrub-Nesting Species in Patagonian Monte Desert. Rangel. J. 2020, 42, 375. [Google Scholar] [CrossRef]
- Lundgren, E.J.; Ramp, D.; Middleton, O.S.; Wooster, E.I.F.; Kusch, E.; Balisi, M.; Ripple, W.J.; Hasselerharm, C.D.; Sanchez, J.N.; Mills, M.; et al. A Novel Trophic Cascade between Cougars and Feral Donkeys Shapes Desert Wetlands. J. Anim. Ecol. 2022, 91, 2348–2357. [Google Scholar] [CrossRef] [PubMed]
- Beschta, R.L.; Donahue, D.L.; DellaSala, D.A.; Rhodes, J.J.; Karr, J.R.; O’Brien, M.H.; Fleischner, T.L.; Deacon Williams, C. Adapting to Climate Change on Western Public Lands: Addressing the Ecological Effects of Domestic, Wild, and Feral Ungulates. Environ. Manag. 2013, 51, 474–491. [Google Scholar] [CrossRef]
- Danvir, R.E. Multiple-Use Management of Western U.S. Rangelands: Wild Horses, Wildlife, and Livestock. Hum.–Wildl. Interact. 2018, 12, 4. [Google Scholar] [CrossRef]
- Beever, E.A.; Aldridge, C.L. Influences of Free-Roaming Equids on Sagebrush Ecosystems, with a Focus on Greater Sage-Grouse. In Greater Sage-GrouseEcology and Conservation of a Landscape Species and Its Habitats; University of California Press: Oakland, CA, USA, 2011; pp. 272–290. [Google Scholar]
- Attum, O.; El Noby, S.K.; Hassan, I.N. The Influence of Landscape Characteristics and Anthropogenic Factors on Waterhole Use by Vulnerable Nubian Ibex Capra Nubiana. Oryx 2009, 43, 564. [Google Scholar] [CrossRef]
- Young, J.K.; Olson, K.A.; Reading, R.P.; Amgalanbaatar, S.; Berger, J. Is Wildlife Going to the Dogs? Impacts of Feral and Free-Roaming Dogs on Wildlife Populations. Bioscience 2011, 61, 125–132. [Google Scholar] [CrossRef]
- Lenth, B.E.; Knight, R.L.; Brennan, M.E. The Effects of Dogs on Wildlife Communities. Nat. Areas J. 2008, 28, 218–227. [Google Scholar] [CrossRef]
- Manor, R.; Saltz, D. The Impact of Free-Roaming Dogs on Gazelle Kid/Female Ratio in a Fragmented Area. Biol. Conserv. 2004, 119, 231–236. [Google Scholar] [CrossRef]
- Soultan, A.; Attum, O.; Lahue, W. The Relationship between Landscape Features and Domestic Species on the Occupancy of Native Mammals in Urban Forests. Urban Ecosyst. 2021, 24, 1117–1128. [Google Scholar] [CrossRef]
- Aloufi, A.; Eid, E. Conservation Perspectives of Illegal Animal Trade at Markets in Tabuk, Saudi Arabia. Traffic Bull. 2014, 26, 77–80. [Google Scholar]
- Cybertracker. Available online: https://cybertracker.org/ (accessed on 1 March 2023).
- SMART. Available online: https://smartconservationtools.org/ (accessed on 1 March 2023).
- Kingsford, R.T.; West, R.S.; Pedler, R.D.; Keith, D.A.; Moseby, K.E.; Read, J.L.; Letnic, M.; Leggett, K.E.A.; Ryall, S.R. Strategic Adaptive Management Planning—Restoring a Desert Ecosystem by Managing Introduced Species and Native Herbivores and Reintroducing Mammals. Conserv. Sci. Pract. 2021, 3, e268. [Google Scholar] [CrossRef]
- King, R.J.H. Feral Animals and the Restoration of Nature. Between Species Online J. Study Philos. Anim. 2009, 13, 1. [Google Scholar] [CrossRef]
- Drummer, T.D.; Degange, A.R.; Pank, L.L.; McDonald, L.L. Adjusting for Group Size Influence in Line Transect Sampling. J. Wildl. Manag. 1990, 54, 511–514. [Google Scholar] [CrossRef]
- Drummer, T.D.; McDonald, L.L. Size Bias in Line Transect Sampling. Biometrics 1987, 43, 13–21. [Google Scholar] [CrossRef]
- Soultan, A.; Pavón-Jordán, D.; Bradter, U.; Sandercock, B.K.; Hochachka, W.M.; Johnston, A.; Brommer, J.; Gaget, E.; Keller, V.; Knaus, P.; et al. The Future Distribution of Wetland Birds Breeding in Europe Validated against Observed Changes in Distribution. Environ. Res. Lett. 2022, 17, 024025. [Google Scholar] [CrossRef]
- Virkkala, R.; Lehikoinen, A. Patterns of Climate-Induced Density Shifts of Species: Poleward Shifts Faster in Northern Boreal Birds than in Southern Birds. Glob. Chang. Biol. 2014, 20, 2995–3003. [Google Scholar] [CrossRef]
- Hovick, T.J.; Allred, B.W.; McGranahan, D.A.; Palmer, M.W.; Dwayne Elmore, R.; Fuhlendorf, S.D. Informing Conservation by Identifying Range Shift Patterns across Breeding Habitats and Migration Strategies. Biodivers. Conserv. 2016, 25, 345–356. [Google Scholar] [CrossRef]
Figure 1.
Location of the study area (black area inside the circle) and the current distribution of the Arabian Partridge retrieved from IUCN (red areas) [9] (A); location of the transects (black dots; N = 73) within the boundary (black polygon) of Harrat Uwayrid Biosphere Reserve (B).
Figure 1.
Location of the study area (black area inside the circle) and the current distribution of the Arabian Partridge retrieved from IUCN (red areas) [9] (A); location of the transects (black dots; N = 73) within the boundary (black polygon) of Harrat Uwayrid Biosphere Reserve (B).

Figure 2.
The association between the density of the Arabian Partridge and the landscape variables. The black line represents the mean of the estimate, while the shaded area represents the confidence interval.
Figure 2.
The association between the density of the Arabian Partridge and the landscape variables. The black line represents the mean of the estimate, while the shaded area represents the confidence interval.

Figure 3.
The detection probability function (half-normal) for Arabian Partridge in Harrat Uwayrid Biosphere Reserve.
Figure 3.
The detection probability function (half-normal) for Arabian Partridge in Harrat Uwayrid Biosphere Reserve.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
The model parameters for the best model. The first part is the parameters for density estimate and the second part is the parameters for detection probability estimate.
Table 1.
The model parameters for the best model. The first part is the parameters for density estimate and the second part is the parameters for detection probability estimate.
| Estimate | SE | Z Value | p-Value | ||
|---|---|---|---|---|---|
| Density model | (Intercept) | 4.492 | 0.801 | 5.607 | 0.001 |
| Wadi width | −0.085 | 0.377 | −0.227 | 0.820 | |
| Wadi width (quadratic term) | 0.032 | 0.275 | 0.117 | 0.906 | |
| Tree cover | 0.004 | 0.216 | 0.018 | 0.985 | |
| Disturbance | −0.519 | 0.229 | −2.269 | 0.023 | |
| Distance to hillside | 1.849 | 0.979 | 1.887 | 0.059 | |
| Interaction between disturbance and distance to hillside | −0.532 | 0.286 | −1.86 | 0.062 | |
| Detection probability model | (Intercept) | 1.797 | 0.482 | 3.726 | 0.001 |
| Covey size | 0.727 | 0.217 | 3.354 | 0.001 | |
| Second observer | 0.024 | 0.257 | 0.096 | 0.923 | |
| Third observer | 0.224 | 0.321 | 0.699 | 0.484 | |
| Medium vegetation cover | 0.992 | 0.449 | 2.21 | 0.027 | |
| High vegetation cover | 0.437 | 0.455 | 0.962 | 0.336 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Soultan, A.; Shah, M.S.; Almalki, A.M.; Browne, S.; Arcilla, N. Arabian Partridge Abundance and Limiting Factors at the Northern Boundary of Its Range. Birds 2023, 4, 179-189. https://doi.org/10.3390/birds4020015
AMA Style
Soultan A, Shah MS, Almalki AM, Browne S, Arcilla N. Arabian Partridge Abundance and Limiting Factors at the Northern Boundary of Its Range. Birds. 2023; 4(2):179-189. https://doi.org/10.3390/birds4020015
Chicago/Turabian StyleSoultan, Alaaeldin, Moayyed Sher Shah, Ahmed Mohammed Almalki, Stephen Browne, and Nico Arcilla. 2023. "Arabian Partridge Abundance and Limiting Factors at the Northern Boundary of Its Range" Birds 4, no. 2: 179-189. https://doi.org/10.3390/birds4020015