
Activity Patterns, Sex Ratio, and Social Organization of the Bare-Faced Curassow (Crax fasciolata) in the Northern Pantanal, Brazil
 ,
,  , , ,
, , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
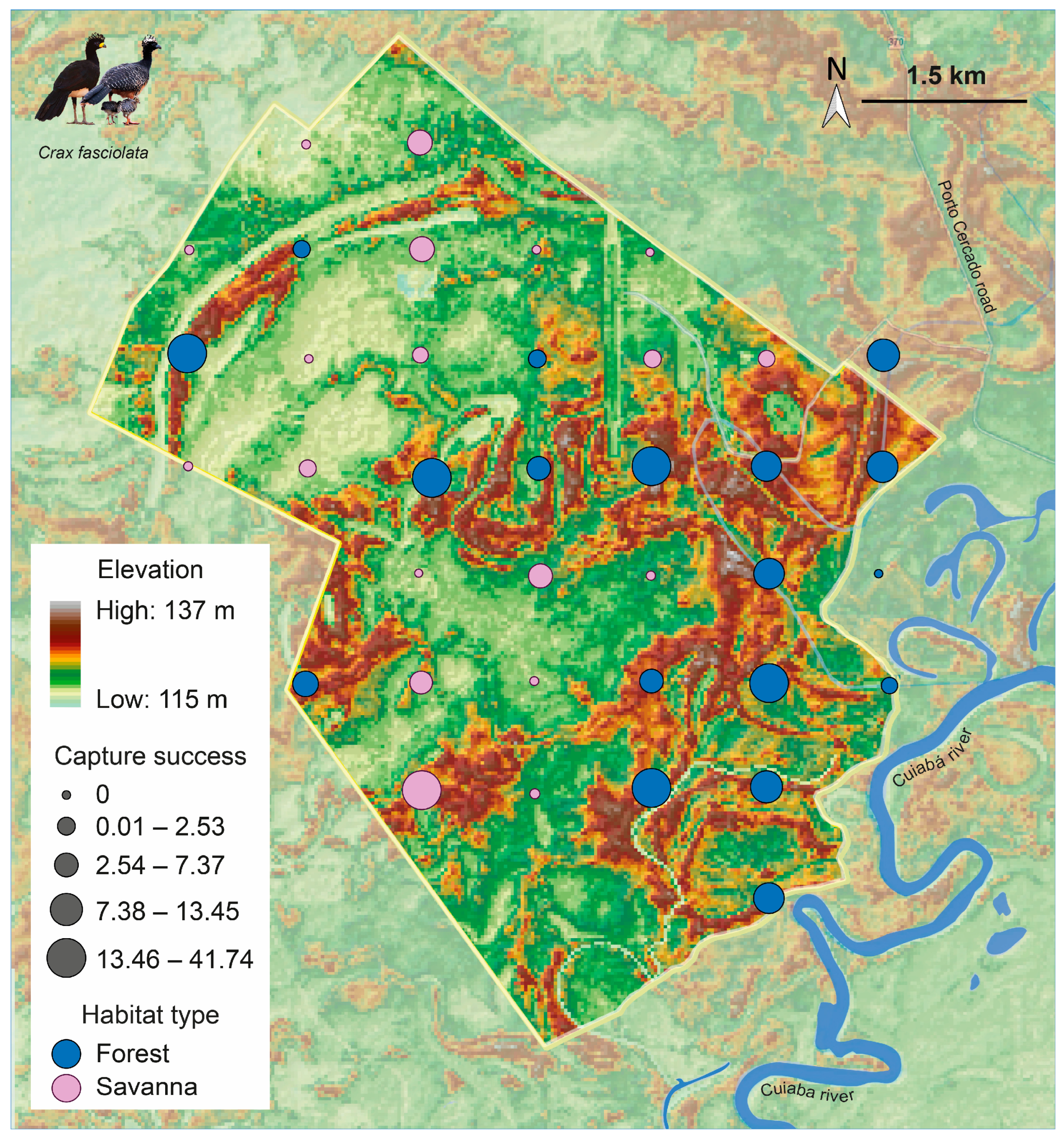
2.1. Study Area
2.2. Data Collection
2.3. Data Preparation
2.4. Statistical Analysis
3. Results
3.1. Camera Trap Records
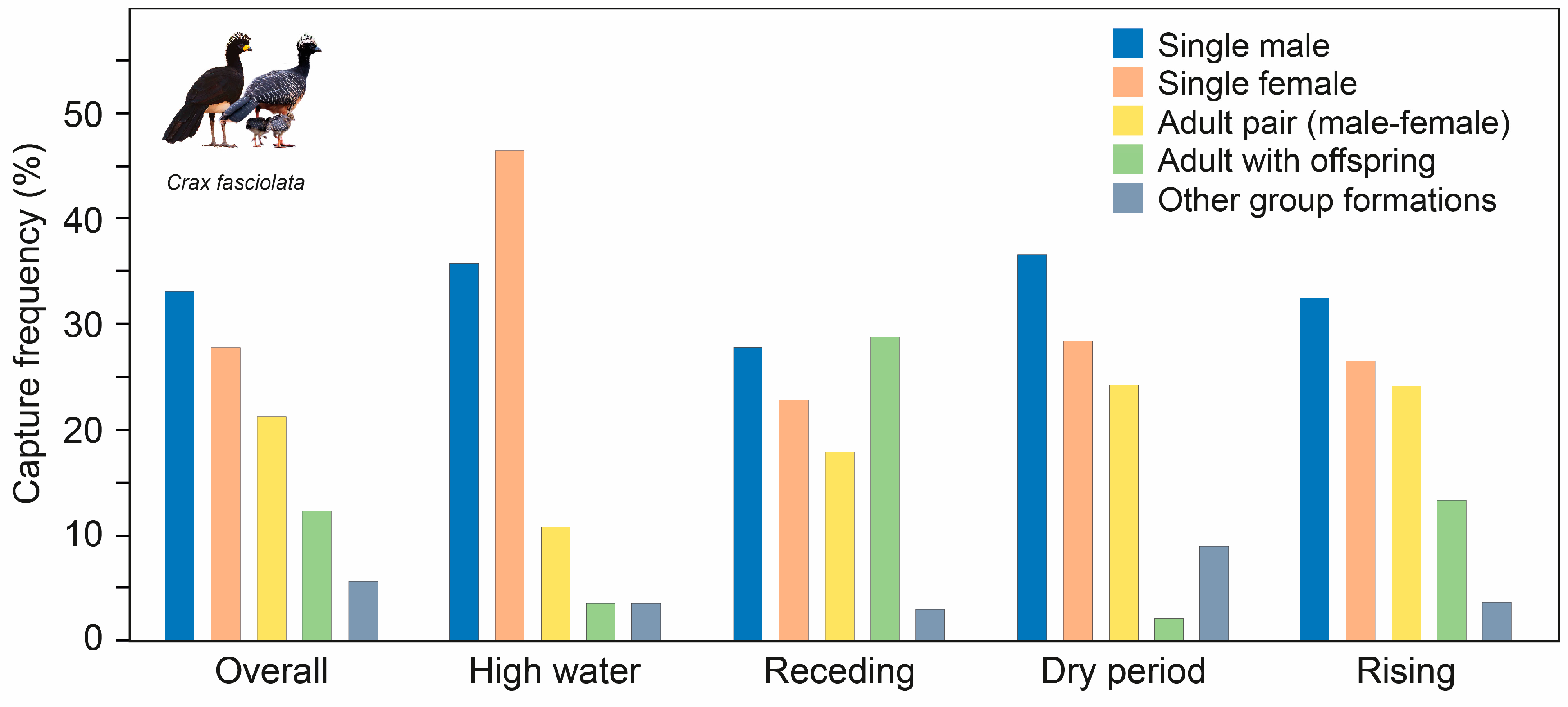
3.2. Capture Success, Seasonal Activity Patterns, and Social Organization
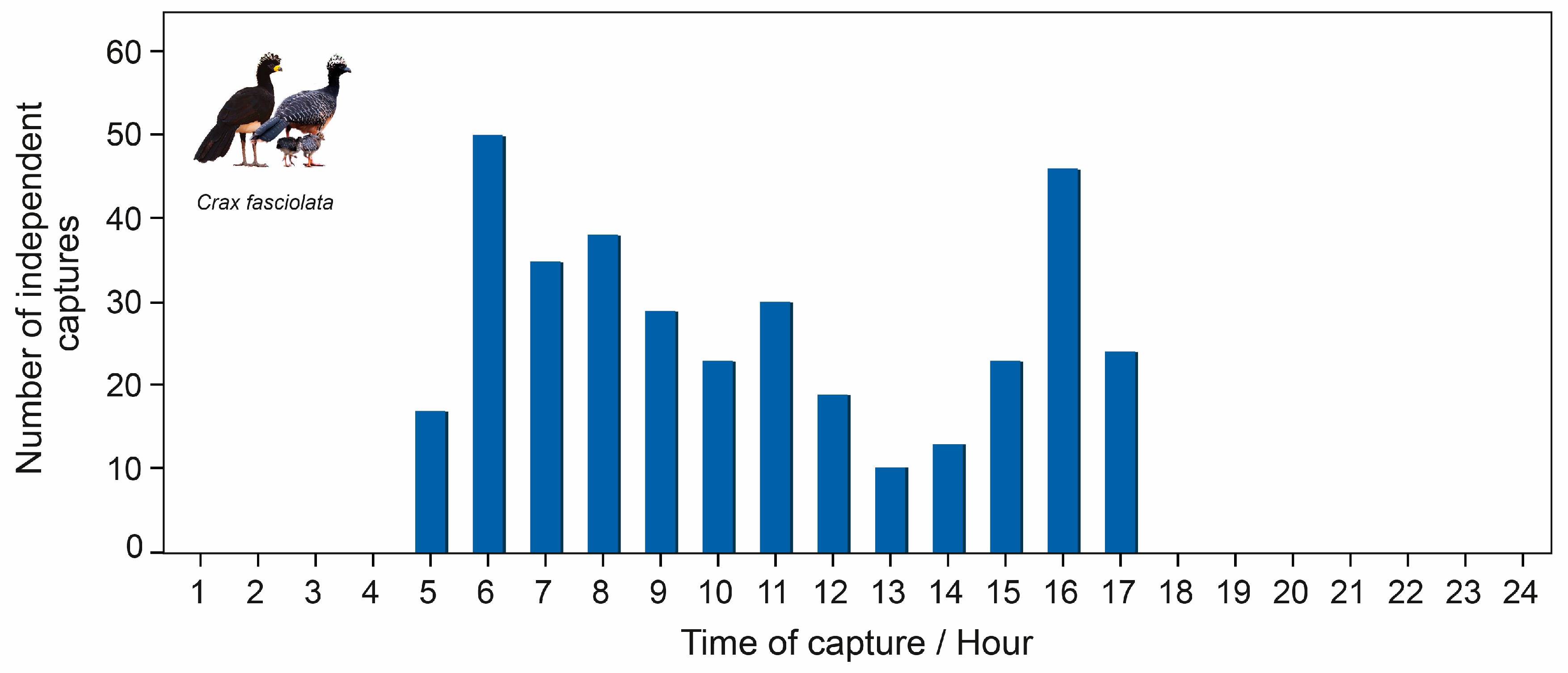
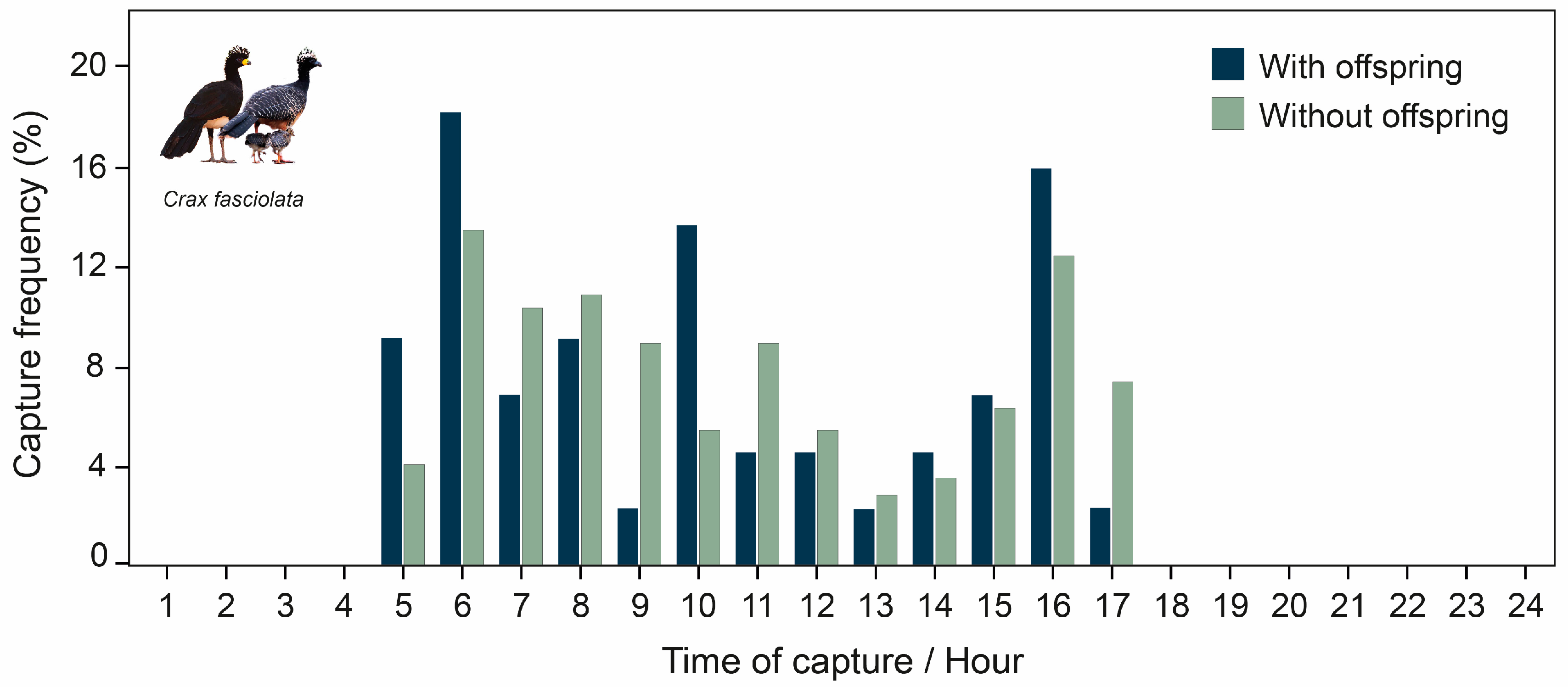
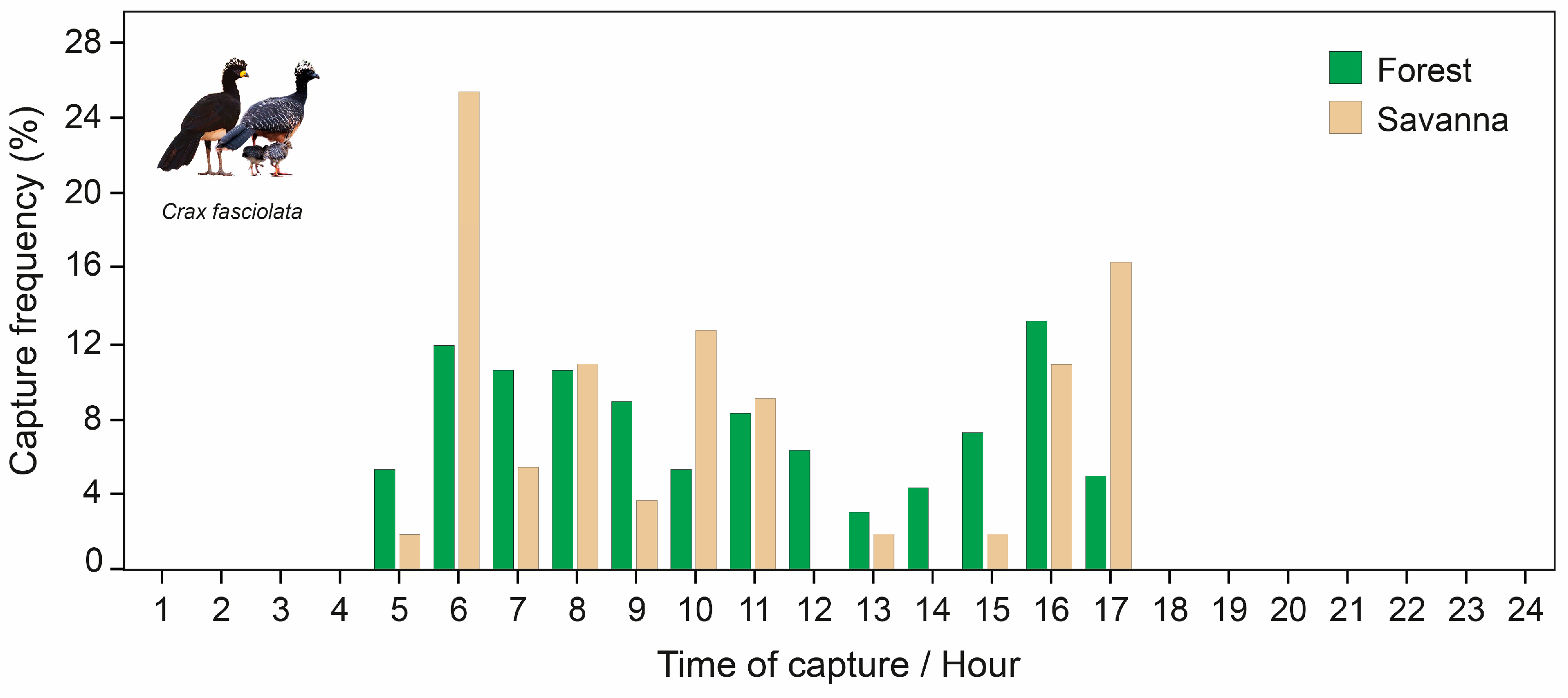
3.3. Daily Activity Patterns
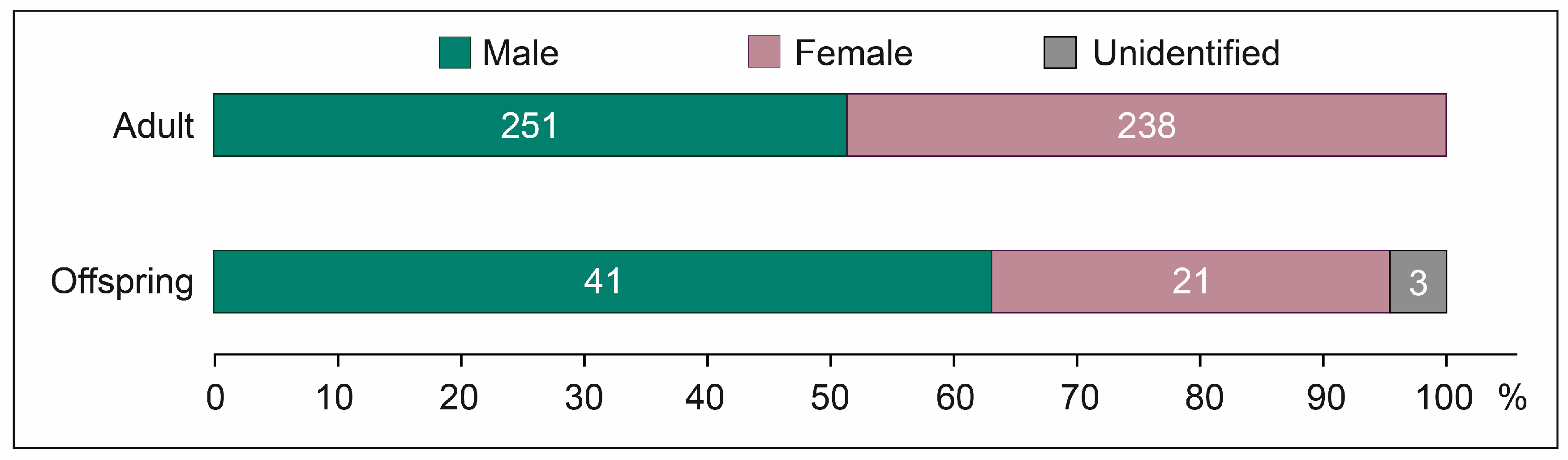
3.4. Sex Ratio: Sexual Structure of the Population
3.5. The Effect of Using Different Data Filters
4. Discussion
4.1. Seasonal Activity Patterns
4.2. Daily Activity Patterns
4.3. Social Organization
4.4. Sex Ratio: Sexual Structure of the Population
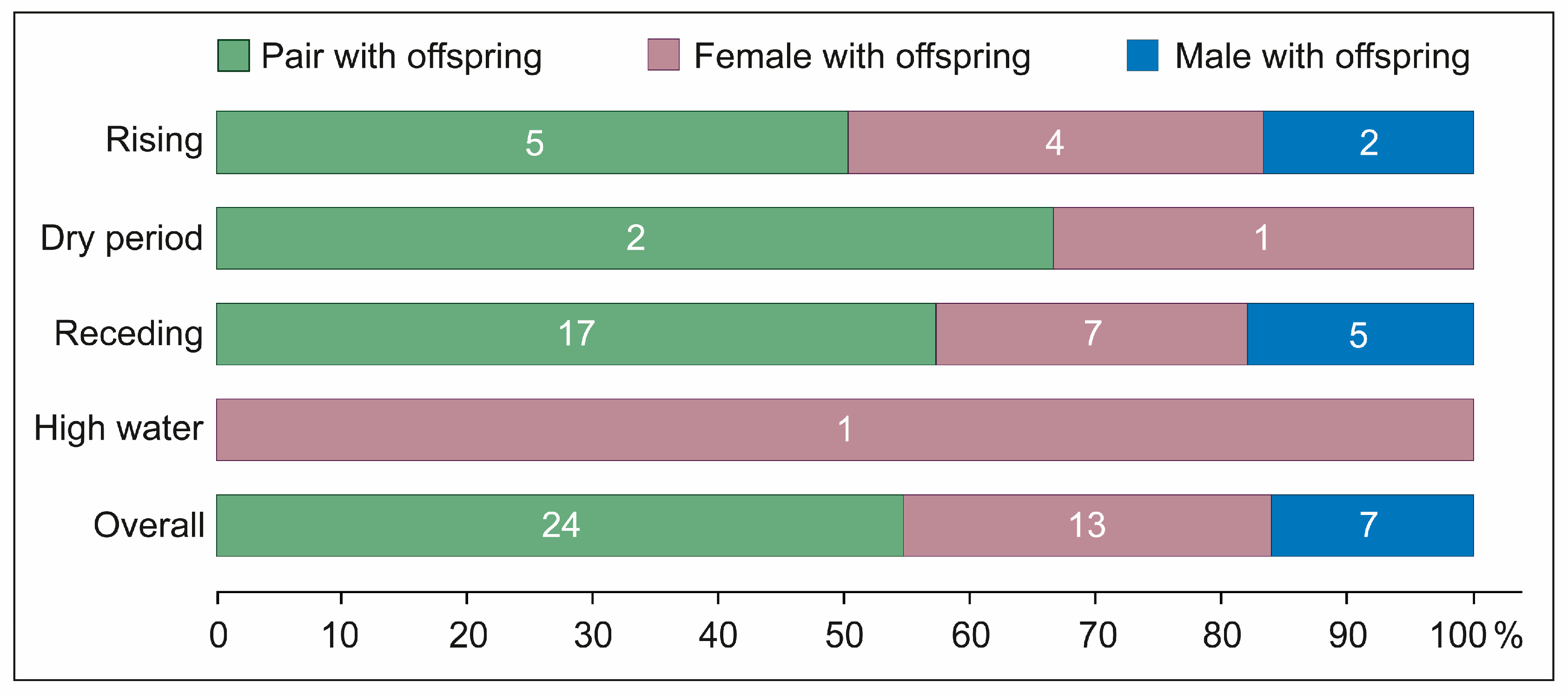
4.5. Parental Care
4.6. Recommendations for Future Studies
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Winkler, D.W.; Billerman, S.M.; Lovette, I.J. Guans, Chachalacas, and Curassows (Cracidae). In Birds of the World, 1st ed.; Billerman, S.M., Keeney, B.K., Rodewald, P.G., Schulenberg, T.S., Eds.; Cornell Lab of Ornithology: Ithaca, NY, USA, 2020. [Google Scholar]
- IUCN 2022. International Union for Conservation of Nature. IUCN Red List of Threatened Species. Available online: https://www.iucnredlist.org/ (accessed on 29 December 2021).
- Brooks, D.M.; Fuller, R.A. Biology and Conservation of Cracids. In Conserving Cracids: The most Threatened Family of Birds in the Americas; Brooks, D.M., Ed.; Miscellaneous Publications of the Houston Museum of Natural Science: Houston, TX, USA, 2006; Volume 6, pp. 10–22. [Google Scholar]
- Del Hoyo, J. Guans, Chachalacas, Curassows (Cracidae). In Handbook of the Birds of the World Alive; Del Hoyo, J., Elliott, A., Sargatal, J., Christie, D.A., Juana, E., Eds.; Lynx Edicions: Barcelona, Spain, 2019. [Google Scholar]
- Brooks, D.M.; Strahl, S.D. Curassows, Guans and Chachalacas. In Status Survey and Conservation Action Plan for Cracids 2000–2004; IUCN/SSC Cracid Specialist Group: Gland, Switzerland; Cambridge, UK, 2000; 182p. [Google Scholar]
- BirdLife International. Species Factsheet: Crax fasciolata. The IUCN Red List of Threatened Species. Available online: http://www.birdlife.org (accessed on 15 December 2022).
- Del Hoyo, J.; Kirwan, G.M.; Sharpe, C.J. Bare-faced Curassow (Crax fasciolata). In Handbook of the Birds of the World Alive; Del Hoyo, J., Elliott, A., Sargatal, J., Christie, D.A., Juana, E., Eds.; Lynx Edicions: Barcelona, Spain, 2019. [Google Scholar]
- Cintra, R.; Yamashita, C. Habitats, Abundância e Ocorrência das Espécies de Aves do Pantanal de Poconé, Mato Grosso, Brasil. Pap. Avulsos Zool. 1990, 37, 1–21. [Google Scholar]
- Del Hoyo, J.; Motis, A. Update Chapter. In Curassows and Related Birds, 2nd ed.; Delacour, J., Amadon, D., Eds.; Lynx Editions and National Museum of Natural History: Barcelona, Spain; New York, NY, USA, 2004. [Google Scholar]
- Desbiez, J.A.L.; Bernardo, S.S.C. Density estimates of the Bare-faced curassow (Crax fasciolata) in the Brazilian Pantanal. Ararajuba 2011, 19, 385–390. [Google Scholar]
- Stotz, D.F.; Fitzpatrick, J.W.; Parker, T.A., III; Moskovits, D.K. Neotropical Birds: Ecology and Conservation; University of Chicago Press: Chicago, IL, USA, 1996. [Google Scholar]
- Delacour, J.; Amadon, D. Curassows and Related Birds, 2nd ed.; Lynx Editions and National Museum of Natural History: Barcelona, Spain; New York, NY, USA, 2004. [Google Scholar]
- Fernández-Duque, F.; Huck, M.; Dávalos, V.; Fernández-Duque, E. Estudio Preliminar sobre la Ecología el Comportamiento y la Demografía del Muitú (Crax fasciolata) en la Selva en Galería del Riacho Pilagá, Formosa, Argentina. Hornero 2013, 28, 65–67. [Google Scholar]
- Laino, R.; Musalem, K.; Caballero-Gini, A.; Bueno-Villafañe, D.; Chaparro, S. Uso de Habitat y Comportamiento de Crax fasciolata en el Chaco Húmedo Paraguayo. Hornero 2018, 33, 121–127. [Google Scholar]
- dos Santos, M.C.R.; Silveira, L.; Jácomo, A.T.D.A.; Alves, G.B.; Ubaid, F.K. Circadian activity patterns and temporal overlap among cracids (Aves: Cracidae) within a vegetation mosaic in the Pantanal of Rio Negro, Brazil. Pap. Avulsos Zool. 2022, 62, e202262011. [Google Scholar] [CrossRef]
- Clay, R.P.; Oren, D.C. Bare-faced Curassow (Crax fasciolata). In Conserving Cracids: The Most Threatened Family of Birds in the Americas; Brooks, D.M., Cancino, L., Pereira, S.L., Eds.; Miscellaneous Publications of the Houston Museum of Natural Science: Houston, TX, USA, 2006; Volume 6, pp. 53–55. [Google Scholar]
- Zalazar, S.; Benitez, A.L.; Di Giacomo, A.S. Determining the factors that influence the occurrence of Bare-faced Curassows (Crax fasciolata) in Humid Chaco, northern Argentina. Avian Conserv. Ecol. 2018, 13, 1. [Google Scholar] [CrossRef] [Green Version]
- Senič, M.; Schuchmann, K.-L.; Marques, M.I. Reproduction Phenology of a High Conservation Priority Cracid—The Bare-faced Curassow (Crax fasciolata; Aves, Galliformes, Cracidae). Pap. Avulsos Zool. 2022, 62, e202262031. [Google Scholar] [CrossRef]
- Fournier, R.; Janik, D. Técnicas de Reproducción en Cautiverio del Pavón (Crax rubra) y su Reintroducción en la Península de Nicoya, Costa Rica. Ornitol. Neotrop. 2008, 19, 353–360. [Google Scholar]
- Hill, D.L.; Arañibar-Rojas, H.; MacLeod, R. Wattled Curassows in Bolivia: Abundance, habitat use, and conservation status. J. Field Ornithol. 2008, 79, 345–351. [Google Scholar] [CrossRef]
- Srbek-Araujo, A.C.; Silveira, L.F.; Chiarello, A.G. The Red Billed Curassow (Crax blumenbachii): Social Organization, and Daily Activity Patterns. Wilson J. Ornitol 2012, 124, 321–327. [Google Scholar] [CrossRef]
- Alves, F.; López-Iborra, G.M.; Silveira, L.F. Population size assessment of the Endangered red-billed curassowCrax blumenbachii: Accounting for variation in detectability and sex-biased estimates. Fauna Flora 2017, 51, 137–145. [Google Scholar] [CrossRef] [Green Version]
- Pardo, L.E.; Lafleur, L.; Spinola, R.M.; Saenz, J.; Cove, M. Camera traps provide valuable data to assess the occurrence of the Great Curassow Crax rubra in northeastern Costa Rica. Neotrop. Biodivers. 2017, 3, 182–188. [Google Scholar] [CrossRef] [Green Version]
- Whitworth, A.; Beirne, C.; Flatt, E.; Huarcaya, R.P.; Diaz, J.C.C.; Forsyth, A.; Molnár, P.K.; Vargas-Soto, J.S. Secondary forest is utilized by Great Curassows (Crax rubra) and Great Tinamous (Tinamus major) in the absence of hunting. Condor 2018, 120, 852–862. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Morales, M.A.; Cruz, P.C.; Cuarón, A.D. Predicted Population Trends for Cozumel Curassows (Crax rubra griscomi): Empirical Evidence and Predictive Models in the Face of Climate Change. J. Field Ornithol. 2009, 80, 317–327. [Google Scholar] [CrossRef]
- Tubelis, D.P.; Tomas, W.M. Bird Species of the Pantanal Wetland, Brazil. Ararajuba 2003, 11, 5–37. [Google Scholar]
- Wetlands International. Waterbird Population Estimates, 4th ed.; Wetlands International: Wageningen, The Netherlands, 2006. [Google Scholar]
- Gwynne, J.A.; Ridgely, R.S.; Tudor, G.; Argel, M. Birds of Brazil. In The Pantanal and Cerrado of Central Brazil; Wildlife Conservation Society; Cornell University Press: Ithaca, NY, USA, 2010. [Google Scholar]
- Nunes, A.P.; Posso, S.R.; da Frota, A.V.B.; Vitorino, B.D.; Laps, R.R.; Donatelli, R.J.; Straube, F.C.; Pivatto, M.A.C.; de Oliveira, D.M.M.; Carlos, B.; et al. Birds of the Pantanal floodplains, Brazil: Historical data, diversity, and conservation. Pap. Avulsos Zool. 2021, 61, e20216182. [Google Scholar] [CrossRef]
- O’Brien, T.G.; Kinnaird, M.F. A picture is worth a thousand words: The application of camera trapping to the study of birds. BirdLife 2008, 18, 144–162. [Google Scholar] [CrossRef] [Green Version]
- Wearn, O.R.; Glover-Kapfer, P. Snap Happy: Camera Traps are an Effective Sampling Tool when compared with Alternative Methods. R. Soc. Open Sci. 2019, 6, 181748. [Google Scholar] [CrossRef] [Green Version]
- Gomes, A.P.N.; Fornitano, L.; Costa, R.T.; Angeli, T.; De Morais, K.D.R.; Olifiers, N.; Bianchi, R.D.C. The importance of protected areas for conservation of bare-faced curassow (Crax fasciolata spix, 1825) (Galliformes: Cracidae) in the São Paulo State, Brazil. Biota Neotrop. 2018, 18, e20180524. [Google Scholar] [CrossRef]
- Lafleur, L.; Pardo, L.; Spínola, R.M.; Saénz, J.; Cove, M.V. Notes on Plumage Patterns and Activity of the Great Curassow (Crax rubra) in Northeastern Costa Rica. Bull. Cracid Group 2014, 36, 17–19. [Google Scholar]
- Pérez-Irineo, G.; Santos-Moreno, A. Occupancy, Relative Abundance, and Activity Patterns of Great Curassow (Crax rubra) in Southeastern Mexico. Ornitol. Neotrop. 2017, 28, 313–320. [Google Scholar] [CrossRef]
- Phalan, B.; Barbosa, A.E.A.; Bernardo, C.S.S.; Bosso, P.; Chaves, F.G.; Corrêa, L.L.C.; Croukamp, C.; Develey, P.; Fernandes, K.C.; Girão, W.; et al. Ex Situ Assessment for Integrated Conservation Planning for Galliformes and Tinamiformes in Brazil; IUCN SSC Conservation Planning Specialist Group: Foz do Iguaçu, Brazil, 2020. [Google Scholar]
- Heckman, C.W. The Pantanal of Poconé. In Biota and Ecology in the Northern Section the World’s Largest Pristine Wetland; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1998; 622p. [Google Scholar]
- Kattan, G.H.; Muñoz, M.C.; Kikuchi, D.W. Population densities of curassows, guans, and chachalacas (Cracidae): Effects of body size, habitat, season, and hunting. Condor 2016, 118, 24–32. [Google Scholar] [CrossRef] [Green Version]
- Thielen, D.; Ramoni-Perazzi, P.; Puche, M.L.; Márquez, M.; Quintero, J.I.; Rojas, W.; Soto-Werschitz, A.; Thielen, K.; Nunes, A.; Libonati, R. The Pantanal under Siege—On the Origin, Dynamics and Forecast of the Megadrought Severely Affecting the Largest Wetland in the World. Water 2021, 13, 3034. [Google Scholar] [CrossRef]
- Seidl, A.F.; De Silva, J.D.S.V.; Moraes, A.S. Cattle ranching and deforestation in the Brazilian Pantanal. Ecol. Econ. 2001, 36, 413–425. [Google Scholar] [CrossRef]
- Abreu, U.G.P.; McManus, C.; Santos, S.A. Cattle ranching, conservation and transhumance in the Brazilian Pantanal. Pastoralism 2010, 1, 99–114. [Google Scholar]
- Alho, C.J.R.; Silva, J.S.V. Effects of Severe Floods and Droughts on Wildlife of the Pantanal Wetland (Brazil)—A Review. Animals 2012, 2, 591–610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Padovani, C.R.; Dacruz, M.L.L.; Padovani, S.L.A.G. Desmatamento do Pantanal Brasileiro para o Ano IV 2000. In Simpósio Sobre Recursos Naturais e Sócio-Econômicos do Pantanal; Embrapa Pantanal: Corumbá, Brazil, 2004. [Google Scholar]
- Bergier, I. Effects of highland land-use over lowlands of the Brazilian Pantanal. Sci. Total Environ. 2013, 463–464, 1060–1066. [Google Scholar] [CrossRef] [PubMed]
- Araujo, A.G.J.; Obregón, G.O.; Sampaio, G.; Monteiro, A.M.V.; da Silva, L.T.; Soriano, B.; Padovani, C.; Rodriguez, D.A.; Maksic, J.; Farias, J.F.S. Relationships between variability in precipitation, river levels, and beef cattle production in the Brazilian Pantanal. Wetl. Ecol. Manag. 2018, 26, 829–848. [Google Scholar] [CrossRef]
- Thielen, D.; Schuchmann, K.-L.; Ramoni-Perazzi, P.; Marquez, M.; Rojas, W.; Quintero, J.I.; Marques, M.I. Quo vadis Pantanal? Expected precipitation extremes and drought dynamics from changing sea surface temperature. PLoS ONE 2020, 15, e0227437. [Google Scholar] [CrossRef] [Green Version]
- Junk, W.J.; Cunha, C.N. Pantanal: A large South American wetland at a crossroads. Ecol. Eng. 2005, 24, 391–401. [Google Scholar] [CrossRef]
- Junk, W.J.; Nunes-da-Cunha, C.; Wantzen, K.M.; Petermann, P.; Strüssmann, C.; Marques, M.I.; Adis, J. Biodiversity and its conservation in the Pantanal of Mato Grosso, Brazil. Aquat. Sci. 2006, 68, 278–309. [Google Scholar] [CrossRef]
- Junk, W.J.; Nunes-da-Cunha, C.; Da Silva, C.J.; Wantzen, K.M. The Pantanal: A large South American Wetland and its Position in Limnological Theory. In The Pantanal: Ecology, Biodiversity and Sustainable Management of a Large Neotropical Seasonal Wetland; Junk, W.J., da Silva, C.J., Nunes da Cunha, C., Wantzen, K.M., Eds.; Pensoft Publishers: Sofia, Bulgaria, 2011; pp. 23–44. [Google Scholar]
- Geofolio. Create a Free Factsheet. Available online: https://geofolio.org/ (accessed on 1 December 2022).
- Copernicus WorldDEM-30 © DLR e.V. 2010–2014 and © Airbus Defence and Space GmbH 2014–2018 Provided under Copernicus by the European Union and ESA; All Rights Reserved. Available online: https://spacedata.copernicus.eu/ (accessed on 15 December 2022). [CrossRef]
- Srbek-Araujo, A.K.; Chiarello, A.G. Is Camera-Trapping an Efficient Method for Surveying Mammals in Neotropical Forests? A case Study in South-Eastern Brazil. J. Trop. Ecol. 2005, 21, 121–125. [Google Scholar] [CrossRef]
- Nunes da Cunha, C.; Junk, W.J.; Leitao-Filho, H.F. Woody vegetation in the Pantanal of Mato Grosso, Brazil: A preliminary typology. Amazoniana 2007, 19, 159–184. [Google Scholar]
- Peral, C.; Landman, M.; Kerley, G.I.H. The inappropriate use of time-to-independence biases estimates of activity patterns of free-ranging mammals derived from camera traps. Ecol. Evol. 2022, 12, e9408. [Google Scholar] [CrossRef] [PubMed]
- Zar, J.H. Biostatistical Analysis, 3rd ed.; Prentice-Hall: Upper Saddle River, NJ, USA, 1996. [Google Scholar]
- McHugh, M.L. Lessons in Biostatistics. The Chi-square test of independence. Biochem. Med. 2013, 23, 143–149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ridout, M.S.; Linkie, M. Estimating overlap of daily activity patterns from camera trap data. J. Agric. Biol. Environ. Stat. 2009, 14, 322–337. [Google Scholar] [CrossRef]
- Meredith, M.; Ridout, M. Overview of the Overlap Package. R Project. 2014. Available online: https://cran.r-project.org/web/packages/overlap/vignettes/overlap.pdf (accessed on 30 October 2019).
- Lund, U.; Agostinelli, C. CircStats: Circular Statistics, from “Topics in Circular Statistics”. R package Version 0.2-6. 2018. Available online: https://CRAN.R-project.org/package=CircStats (accessed on 22 December 2019).
- R core team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- Palencia, P.; Vicente, J.; Soriguer, R.C.; Acevedo, P. Towards a best-practices guide for camera trapping: Assessing differences among camera trap models and settings under field conditions. J. Zoöl. 2022, 316, 179–208. [Google Scholar] [CrossRef]
- MAyDS y AA (Ministerio de Ambiente y Desarrollo Sustentable y Aves Argentina). Categorización de las Aves de la Argentina. In Informe del Ministerio de Ambiente y Desarrollo Sustentable de la Nación y de Aves Argentinas; Ministerio de Ambiente y Desarrollo Sustentable: Buenos Aires, Argentina, 2017; 98p. [Google Scholar]
- Kirwan, G.M.; del Hoyo, J.; Collar, N.; Christie, D.A.; Sharpe, C.J. Bare-faced Curassow (Crax fasciolata). In Birds of the World, version 1.1; Billerman, S.M., Keeney, B.K., Rodewald, P.G., Schulenberg, T.S., Eds.; Cornell Lab of Ornithology: Ithaca, TX, USA, 2020. [Google Scholar]
- de Deus, F.F.; Schuchmann, K.-L.; Marques, M.I. Seasonality in the Brazilian Pantanal influences avian functional diversity. Stud. Neotrop. Fauna Environ. 2020, 57, 187–197. [Google Scholar] [CrossRef]
- Cândido Júnior, J.F. Aceitação de Alimento por Crax blumenbachii, Crax fasciolata e Penelope superciliaris (Cracidae) em Cativeiro. Ararajuba 1996, 4, 42–47. [Google Scholar]
- Prance, G.T.; Schaller, G.B. Preliminary Study of Some Vegetation Types of the Pantanal, Mato Grosso, Brazil. Brittonia 1982, 34, 228–251. [Google Scholar] [CrossRef]
- Mamede, S.B.; Alho, C.J.R. Response of wild mammals to seasonal shrinking-and-expansion of habitats due to flooding regime of the Pantanal, Brazil. Braz. J. Biol. 2006, 66, 991–998. [Google Scholar] [CrossRef] [Green Version]
- Hernández-SaintMartín, A.D.; Rosas-Rosas, O.C.; Palacio-Núñez, J.; Tarango-Arámbula, L.A.; Clemente-Sánchez, F.; Hoogesteijn, A.L. Activity Patterns of Jaguar, Puma and Their Potential Prey in San Luis Potosi, Mexico. Acta Zool. Mex. 2013, 29, 520–533. [Google Scholar] [CrossRef]
- Muñoz, M.C.; Kattan, G.H. Diets of Cracids: How Much Do We Know? Ornitol. Neotrop. 2007, 18, 21–36. [Google Scholar]
- Blake, J.G.; Mosquera, D.; Guerra, J.; Loiselle, B.A.; Romo, D.; Swing, K. Mineral Licks as Diversity Hotspots in Lowland Forest of Eastern Ecuador. Diversity 2011, 3, 217–234. [Google Scholar] [CrossRef] [Green Version]
- Sæbø, J.S. Spatial and Temporal Distributions and Interactions in a Neotropical Ground-Dwelling Animal Community. Master’s Thesis, Norwegian University of Life Sciences, Akershus, Norway, 2016. [Google Scholar]
- Cepeda-Duque, J.C.; Gómez-Valencia, B.; Alvarez, S.; Gutiérrez–Sanabria, D.R.; Lizcano, D.J. Daily Activity Pattern of Pumas (Puma concolor) and their Potential Prey in a Tropical Cloud Forest of Colombia. Anim. Biodivers Conserv. 2021, 44, 267–278. [Google Scholar] [CrossRef]
- Beirne, C.; Pillco-Huarcaya, R.; Serrano-Rojas, S.J.; Whitworth, A. Terrestrial camera traps: Essential tool for the detection and future monitoring of the Critically Endangered Sira curassow Pauxi koepckeae. Endanger. Species Res. 2017, 32, 145–152. [Google Scholar] [CrossRef] [Green Version]
- Krieg, H.; Schumacher, E. Beobachtungen von Wildhühnern. Verh. Ornithol. 1936, 21, 1–18. [Google Scholar]
- Sedaghatkish, G.; Brooks, D.M. Retraso Evolutivo en los Crácidos: Cantado para Ser la Cena del Cazador. In Manejo y Conservación de Fauna Silvestre en América Latina; Fang, T.G., Montenegro, O.L., Bodmer, R.E., Eds.; Editorial-Instituto de Ecología: La Paz, Bolivia, 1999; pp. 335–340. [Google Scholar]
- Donald, P.F. Adult sex ratios in wild bird populations. Ibis 2007, 149, 671–692. [Google Scholar] [CrossRef]
- Coupe, M.F. Breeding Sclater’s Crested Curassows (Bare-faced Curassow). News from Chester ZOO. Aviculture 1966, 72, 168–169. [Google Scholar]
- Bruno, S.F.; de Carvalho, R.B.A.; Kirwan, G.M. Bare-faced Curassow Crax fasciolata in Serra da Canastra National Park, Minas Gerais, Brazil. Cotinga 2006, 26, 80–86. [Google Scholar]
- Kirwan, G.M. Notes on the breeding ecology and seasonality of some Brazilian birds. Rev. Bras. De Ornitol. 2009, 17, 121–136. [Google Scholar]
- Remeš, V.; Freckleton, R.P.; Tökölyi, J.; Liker, A.; Székely, T. The evolution of parental cooperation in birds. Proc. Natl. Acad. Sci. USA 2015, 112, 13603–13608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liker, A.; Freckleton, R.P.; Székely, T. Divorce and Infidelity Are Associated with Skewed Adult Sex Ratios in Birds. Curr. Biol. 2014, 24, 880–884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pease, B.S.; Nielsen, C.K.; Holzmueller, E.J. Single-Camera Trap Survey Designs Miss Detections: Impacts on Estimates of Occupancy and Community Metrics. PLoS ONE 2016, 11, e0166689. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Independent Captures | Detected Individuals | Sampling Effort | Capture Success | |
|---|---|---|---|---|---|
| Total | 357 | 554 | 4768 | 7.49 | |
| Season | High water (Jan–Mar) | 28 | 33 | 473 | 5.92 |
| Receding (Apr–Jun) | 101 | 184 | 1076 | 9.39 | |
| Dry period (Jul–Sep) | 145 | 211 | 1989 | 7.29 | |
| Rising (Oct–Dec) | 83 | 126 | 1230 | 6.75 | |
| Habitat | Forest | 302 | 463 | 2261 | 13.36 |
| Savanna | 55 | 91 | 2507 | 2.19 | |
| Time Interval (min) | n | n Loss (%) | Chi2 | p Value | Overlap (Δ4) | 95% CI | Watson’s U2 | p Value | |
|---|---|---|---|---|---|---|---|---|---|
| Independent BFC captures (only first) | 1 | 461 | - | - | - | - | - | - | - |
| 10 | 369 | 19.96 | 3.29 | 0.99 | 0.96 | 0.91–0.99 | 0.06 | >0.10 | |
| 20 | 358 | 22.34 | 3.94 | 0.98 | 0.95 | 0.91–0.99 | 0.06 | >0.10 | |
| 30 | 357 | 22.56 | 4.08 | 0.98 | 0.95 | 0.90–0.99 | 0.07 | >0.10 | |
| 60 | 343 | 25.60 | 4.73 | 0.97 | 0.95 | 0.90–0.98 | 0.08 | >0.10 | |
| 90 | 337 | 26.90 | 5.60 | 0.93 | 0.94 | 0.89–0.98 | 0.10 | >0.10 | |
| All detected BFC individuals | 1 | 686 | - | - | - | - | - | - | - |
| 10 | 569 | 17.06 | 3.65 | 0.99 | 0.96 | 0.92–0.99 | 0.06 | >0.10 | |
| 20 | 556 | 18.95 | 4.05 | 0.98 | 0.96 | 0.93–0.99 | 0.06 | >0.10 | |
| 30 | 554 | 19.24 | 4.86 | 0.96 | 0.95 | 0.91–0.99 | 0.08 | >0.10 | |
| 60 | 536 | 21.87 | 5.26 | 0.95 | 0.95 | 0.91–0.98 | 0.09 | >0.10 | |
| 90 | 529 | 22.89 | 5.95 | 0.92 | 0.95 | 0.91–0.98 | 0.10 | >0.10 | |
| Comparison between | |||||||||
| 1 (first) | 1 (all) | 32.80 | 2.62 | 1.00 | 0.97 | 0.93–1.00 | 0.03 | >0.10 | |
| 10 (first) | 10 (all) | 35.15 | 3.37 | 0.99 | 0.96 | 0.92–1.00 | 0.03 | >0.10 | |
| 20 (first) | 20 (all) | 35.61 | 3.86 | 0.99 | 0.96 | 0.92–1.00 | 0.03 | >0.10 | |
| 30 (first) | 30 (all) | 35.56 | 3.38 | 0.99 | 0.96 | 0.92–1.00 | 0.03 | >0.10 | |
| 60 (first) | 60 (all) | 36.01 | 3.85 | 0.99 | 0.96 | 0.92–1.00 | 0.03 | >0.10 | |
| 90 (first) | 90 (all) | 36.29 | 3.55 | 0.99 | 0.96 | 0.92–1.00 | 0.03 | >0.10 | |
| Independence Interval (min) | 10 | 20 | 30 | 60 | 90 |
|---|---|---|---|---|---|
| Capture success | 7.74 | 7.51 | 7.49 | 7.19 | 7.07 |
| Deviation from the baseline | +3.34% | +0.27% | - | −4.01% | −5.61% |
| Studied Species | Adult Sex Ratio | Offspring Sex Ratio | Source | |
|---|---|---|---|---|
| Great Curassow (Crax rubra) | - | - | 0.59:1.00 c | [19] |
| Wattled Curassow (Crax globulosa) | 1.20:1.00 a | - | - | [20] |
| Great Curassow (Crax rubra griscomi) | 0.56:1.00 a | - | - | [25] |
| Red-billed Curassow (Crax blumenbachii) | - | 1.56:1.00 b | - | [21] |
| Bare-faced Curassow (Crax fasciolata) | 1.60:1.00 a | 1.67:1.00 b | - | [13] |
| Red-billed Curassow (Crax blumenbachii) | - | 1.60:1.00 b | - | [22] |
| Great Curassow (Crax rubra) | - | 0.71:1.00 b | - | [23] |
| Great Curassow (Crax rubra) | - | 1.50:1.00 b | - | [24] |
| Bare-faced Curassow (Crax fasciolata) | - | 1.26:1.00 b | - | [14] |
| Bare-faced Curassow (Crax fasciolata) | - | 1.05:1.00 b | 0.51:1.00 b | This study |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Senič, M.; Schuchmann, K.-L.; Burs, K.; Tissiani, A.S.; de Deus, F.; Marques, M.I. Activity Patterns, Sex Ratio, and Social Organization of the Bare-Faced Curassow (Crax fasciolata) in the Northern Pantanal, Brazil. Birds 2023, 4, 117-137. https://doi.org/10.3390/birds4010010
Senič M, Schuchmann K-L, Burs K, Tissiani AS, de Deus F, Marques MI. Activity Patterns, Sex Ratio, and Social Organization of the Bare-Faced Curassow (Crax fasciolata) in the Northern Pantanal, Brazil. Birds. 2023; 4(1):117-137. https://doi.org/10.3390/birds4010010
Chicago/Turabian StyleSenič, Martin, Karl-L. Schuchmann, Kathrin Burs, Ana Silvia Tissiani, Filipe de Deus, and Marinez I. Marques. 2023. "Activity Patterns, Sex Ratio, and Social Organization of the Bare-Faced Curassow (Crax fasciolata) in the Northern Pantanal, Brazil" Birds 4, no. 1: 117-137. https://doi.org/10.3390/birds4010010