
When to Return to Normal? Temporal Dynamics of Vigilance in Four Situations

School of Biological & Environmental Sciences, Liverpool John Moores University, James Parsons Building, Byrom Street, Liverpool L3 3AF, UK
Birds 2023, 4(1), 1-14; https://doi.org/10.3390/birds4010001
Submission received: 23 November 2022
/
Revised: 16 December 2022
/
Accepted: 19 December 2022
/
Published: 22 December 2022
(This article belongs to the Special Issue Feature Papers of Birds 2022–2023)
Abstract
:Simple Summary
Animals monitor their environment (vigilance) to detect predators and keep an eye on conspecifics. Once a change has been detected, vigilance generally increased to collect more information. However, little is known about the time course of vigilance to return to normal, which can tell us how threatening animals perceive different situations. This was investigated in Gouldian Finches by assessing the frequency of head movements in 15-min blocks across an hour in four situations (familiar environment, two changed environments with novel objects added at different locations and an unfamiliar environment). The time course of vigilance to return to normal differed between situations with consistently high vigilance when a novel object was placed above the feeder that only returned to normal at the end of the hour indicating that this situation was perceived as threatening. In contrast, a novel object placed at a neutral location on a perch did not elicit a strong response and vigilance mirrored the familiar situation with only a slight decline over time. Finally, vigilance plummeted to very low levels in the unfamiliar environment and did not recover within an hour. Birds may have looked in one direction for longer to take in all details.
Abstract
Vigilance is an important behaviour to monitor the environment from detecting predators to tracking conspecifics. However, little is known about how vigilance changes over time either without disturbance (vigilance decrement) or after a change occurred. The time course of vigilance can indicate how animals perceive a situation and the potential mechanism used to deal with it. I investigated the time course of vigilance in Gouldian Finches in four situations (familiar environment, two changed environments–novel object at a neutral location (exploration trial) or above the feeder (neophobia trial), novel environment). The frequency of head movements was assessed in four consecutive 15-min blocks in same sex pairs with a high frequency generally seen as indicative of high vigilance. Vigilance decreased over time in the familiar situation indicating vigilance decrement with a similar time course in the exploration trial. Vigilance was consistently high in the neophobia trial and only returned to normal in the last block. Finally, vigilance plummeted in the novel environment and did not return to normal within an hour. Results suggest that perceived threats affected vigilance and that information gathering reduced uncertainty allowing vigilance to return to normal levels but with different time courses depending on the situation.
1. Introduction
Vigilance is the focussing of attention on the detection of subtle changes in the environment that occur over a long period of time [1]. It is seen as an important antipredator strategy [2] but is also used to keep track of conspecifics’ activities (e.g., [3,4]). Vigilance is a costly behaviour as it often cannot be performed simultaneously with other behaviours such as foraging (e.g., [5]). While numerous studies have investigated how costs of vigilance can be reduced (e.g., group size [6,7], group composition [8]) and how this differs between individuals due to age and sex [9,10,11,12], there is relatively little research about how vigilance is moderated once a change in the environment has been detected. Investigating the time course to return to before disturbance vigilance levels helps to identify how animals perceive threats and the potential mechanisms involved in dealing with threats over time.
Vigilance serves information gathering [13], not only to detect changes in the environment but also to monitor them [14] which over time can result in approach and exploration (predator inspection [9]; approaching observers [7]). However, vigilance has also been linked to fear [15], neophobia and uncertainty [16] resulting in heightened vigilance in unfamiliar or threatening situations [7,17,18,19,20,21]. Sirot and Pays [13] modelled optimal vigilance strategies considering perceived predation risk. After arrival in a new foraging patch, animals should initially show high vigilance with long and frequent scans to assess the situation. Over time, this will reduce uncertainty about any potential predators present and animals will scan for shorter durations and less often, overall reducing vigilance. Likewise, Carbone et al. [22] suggested that additionally to a reduction in perceived risk, increasing resource competition may further decrease vigilance when resources get depleted. In contrast, decreases in hunger levels (or improved body condition) can result in devoting more time to vigilance [23].
Initial higher vigilance in a novel environment has been confirmed in mammals and fish [19,24]. However, Sirot and Pays’ [13] model encompasses not only arrival in a new foraging patch but generally dangerous situations. Increased vigilance after a change in the environment (increased risk perception) has been confirmed in a range of studies (predator detection [24]; human disturbance [18]; conspecific or heterospecific activity: [25,26,27]) with higher vigilance the more threatening a situation was perceived (higher predator activity at closer distance [17]; increased human disturbance [6,7,19]; aversive person vs. unfamiliar person vs. friendly person next to food, or presence of dog vs. human [15]; urban vs. rural environment [11]). However, next to nothing is known about the further time course of vigilance once a change has occurred.
Some studies investigated the effect of repeated exposure to the same stimulus (habituation) on vigilance over longer periods with most studies finding no reduction in vigilance over months or years in response to ecotourism [7,19,21] (but see [27]) indicating persistent costs of higher vigilance when visitors are around. Others studied habituation in the short-term (days) across experimental trials with some finding a reduction in vigilance (marmosets (Callithrix penicillata) towards model predator [24]; dairy cattle (Bos taurus dom.) towards dogs and people [15], whereas others did not find a change in vigilance (marmosets in novel environment [24]; King Penguins (Aptenodytes patagonicus) towards approaching observer [28]).
The only studies addressing the temporal change of vigilance within sessions primarily focus on vigilance decrement in unchanging environments. Vigilance decrement describes the decline in performance over time to detect rare events [29]. Several theories exist to explain this decline, which can broadly be divided into changes at the processing level or the cognitive level [30]. At the processing level, vigilance decrement can result from sensory habituation (due to lack of changes in the environment) [31]. At the cognitive level, a decline in vigilance can be linked to continuous depletion of attentional resources [30], under-stimulation resulting in decreasing arousal and consequently decreasing vigilance [32], inability to maintain cognitive control due to competition from other tasks (goal habituation; [30]) or a change in the threshold to respond (Signal Detection theory; [31]). Particularly research on humans has found that vigilance decreases over time when the environment does not change with negative consequences for monotonous tasks (e.g., [10,29,30,33]). Only recently, has vigilance decrement been investigated in animals. A decline in vigilance in unchanging environments due to vigilance decrement reduces the probability to detect predators [32]. Vigilance decrement has been shown in spiders in a visual task [32,34], harbour seals (Phoca vitulina) when hauled out of water [35] and in gulls (Larus spp.) [36] with the latter study assuming that the decrease in vigilance was linked to an adjustment in perceived risk rather than vigilance decrement. Very little is known about whether age or sex affect vigilance decrement, but Sauter et al. [10] found that the decrement is faster in older people, whereas no sex differences were found.
Here, I investigate the time course of vigilance in the Gouldian Finch (Chloebia gouldiae) in a familiar environment without a change and in three novel situations considering age, sex and group composition. Gouldian Finches inhabit tropical savannah grassland in North Australia [37] and occur in three distinct head colour morphs in the same population [38]. Head colour morphs do not differ in their vigilance, but head colour morph composition affects vigilance with mixed morph groups being more vigilant [8].
Vigilance was recorded as horizontal head movements, which reflects the area scanned [39]. A higher frequency of head movements is consistent with higher vigilance, e.g., once a predator has been detected [40], at the periphery of a group [41] or when in a novel environment [42]. The frequency of horizontal head movements was used to investigate (a) whether vigilance decrement occurred in the familiar situation, (b) the time course of vigilance after a change in the environment and (c) whether the time course of vigilance differed between head colour morphs, morph compositions, ages and sexes.
2. Materials and Methods
Twenty-two Gouldian Finches took part in the experiment consisting of 12 females (6 black-headed, 6 red-headed) and 10 males (4 black-headed, 6 red-headed) between 1 to 8 years old. All birds were acquired over several years from 10 different bird breeders with all birds residing in the Animal Facility at Liverpool John Moors University since at least a year. Birds were kept in mixed age, sex and head colour groups of about 6 birds, each. Holding cages (120 × 80 × 100 cm (length × depth × height)) consisted of three closed walls with a wire mesh front and ceiling. The cages were structured with natural twigs and perches with food provided in feeders at the front. Food consisted of a mixture of Blattner Amadine Zucht Spezial (Gouldamadine), Blattner Astrilden Spezial and Blattner rote Mannahirse (Blattner Heimtierfutter, Ermengerst, Germany). Blattner bird grit was provided separately, as were eggshells. Cages contained water dispensers and a bath. The light regime was 13 h light to 11 h dark.
2.1. Experimental Procedure
Experiments were conducted in a separate room containing six cages (120 × 70 × 100 cm) arranged back-to-back in two rows. Cages consisted of three wooden walls with a wire mesh front and ceiling facilitating that birds could not see but hear each other. Two perches were available left and right running perpendicular to the front. Food and water were provided at the front wire. A camera was permanently mounted on a tripod one metre away from the front of each cage.
For the current experiment only four of the six cages were used due to logistical reasons. Experiments were conducted over a seven-day period testing four groups at the same time (=one batch) with the next batch of birds being moved into the experimental cages the day after the preceding group had finished. Gouldian Finches are highly social; therefore, birds were tested in same sex pairs either in same or mixed head colour pairs (4 black-headed, 4 red-headed, 4 mixed head colour pairs balanced across sexes). As there were only 22 birds, two black-headed males were used a second time to provide a partner for two red-headed males. Vigilance data of the re-used birds were not included in the analysis as they were already represented in the data set with their first testing. Head colour combinations and sexes were balanced within and across batches.
The following experiments were conducted: The time course of vigilance was investigated in four different situations. (1) novel environment (when released into the experimental cage), (2) familiar environment (day 5 in experimental cage), (3) changed environment 1 (day 6) and (4) changed environment 2 (day 7). A change in the environment consisted of either a novel object (metal star, 10.5 × 4.5 cm, height × width) placed at a neutral location in the cage (over a perch but away from food and water = exploration trial; Figure 1) or a novel object (Christmas stocking sock, 11.5 × 5.5 × 2.5 cm; Height × Width × Depth) placed over the feeder (=neophobia trial; Figure 1). The order of presentation (day 6 or 7) was balanced across the objects. The two changed environments represented different types of situations with different levels of risk. In the exploration trial, the star at the neutral location could be ignored as it was away from vital resources causing a lower level of risk. However, when birds approached then this reflected exploration and information gathering [43]. In contrast, the sock over the feeder in the neophobia trial required approach when the birds wanted to feed presenting a much higher threat. In this situation, fear (neophobia) competes with hunger [43]. All experiments started at 10:00 am and lasted for one hour. Data were recorded with digital video cameras using the GeoVision 1480 software (GeoVision Inc., Taipei, Taiwan) for later analysis.
Birds were released into the cage with video recording starting immediately to collect data on the time course of vigilance in the novel environment (situation 1). After one hour, video recording was stopped, and the birds had until day 5 to habituate to the cage. On day 5, the time course of vigilance was recorded again for one hour in the now familiar environment (situation 2). The next day, either the star was positioned over the neutral perch (exploration trial) or the stocking sock over the feeder (neophobia trial; changed environment one—situation 3). In the neophobia trial, birds were food deprived for one hour prior to the start of the experiment to have similar hunger levels. On day 7, the object not used on day 6 was introduced (changed environment two—situation 4). On both days recording lasted for one hour after which the objects were removed. Birds were moved back into their holding cages after the experiment had finished on day 7.
2.2. Data Analysis
Vigilance was measured as horizontal head movements [39] defined as any movement of the head and extracted for each individual and situation. For analysis, the one-hour session was divided into four periods of 15 min, each, to see whether vigilance changed as a function of time. The frequency of head movements per minute was calculated for each period and used for further analyses. This measure is inversely related to the duration of looking into a particular direction [41].
All analyses were conducted with SPSS v. 26. The full data set is available in the supplementary Table S1. For the time course of vigilance across the four different situations, all data were analysed together (n = 22 birds tested in four situations divided into four periods resulting in 352 data points) using General Linear Mixed Models (GLMM). The dependent variable was the frequency of head movements per minute in each period using an identity link function. Situation (4 categories: novel environment, changed environment (exploration and neophobia trial), familiar environment) and period (four 15-min blocks) were entered as repeated measures with n = 22 in each category. A series of models were built to test for effects of different variables and interactions. The basic model consisted of a fixed factor (period) to test for changes in vigilance over time, an interaction term (period × situation) to test for differences in the time course of vigilance in the different situations and a covariate (partner head colour morph; black-headed, red-headed) to account for effects of group composition [8]. More complex models included interaction terms with period × situation. These were age class (3 categories: one- to two years (n = 5), three to four years (n = 8), older than four years (n = 9)), sex, head colour morph (black-headed, red-headed) and partner head colour morph to test whether they have an effect on the time course of vigilance. Sequential Sidak correction accounted for multiple comparisons. Akaike criterion was used to select the best model. As the GLMM provided posthoc tests for the same period between situations but not for different periods within the same situation, separate GLMM were run for each situation with the same variables as in the main analysis. The only exception was the exploration trial where the basic model was used due to convergent issues.
In a second step I tested whether vigilance differed between individuals that approached or did not approach the objects (exploration and neophobia trial) as approach and closer inspection may reduce uncertainty and hence speed up the process to return to before-disturbance vigilance levels. Exploration and neophobia trials were analysed separately. All birds were included (n = 22 resulting in 88 data points, each) and data were analysed with a GLMM using the same dependent vigilance variable (LG10 transformed) and link function as above but with 1000 iterations to improve convergence. The basic model consisted of two fixed factors (period and approach), an interaction term (period × approach) and a covariate (partner head colour morph). Approach had two categories (yes/no). In the exploration trial, approach meant that birds touched the object (n = 9), whereas in the neophobia trial these were birds that approached the feeder in reach or landed on the feeder (n = 8; only one bird ate next to the object). More complex models included interaction terms of age class, sex, head colour morph and partner head colour morph with period. Sequential Sidak correction accounted for multiple comparisons. Akaike criterion was used to select the best model.
2.3. Ethical Note
Experiments were conducted in accordance with The Association for the Study of Animal Behaviour (ASAB) ethical guidelines [44] and were non-invasive. Experiments were approved by the University Ethics Committee.
3. Results
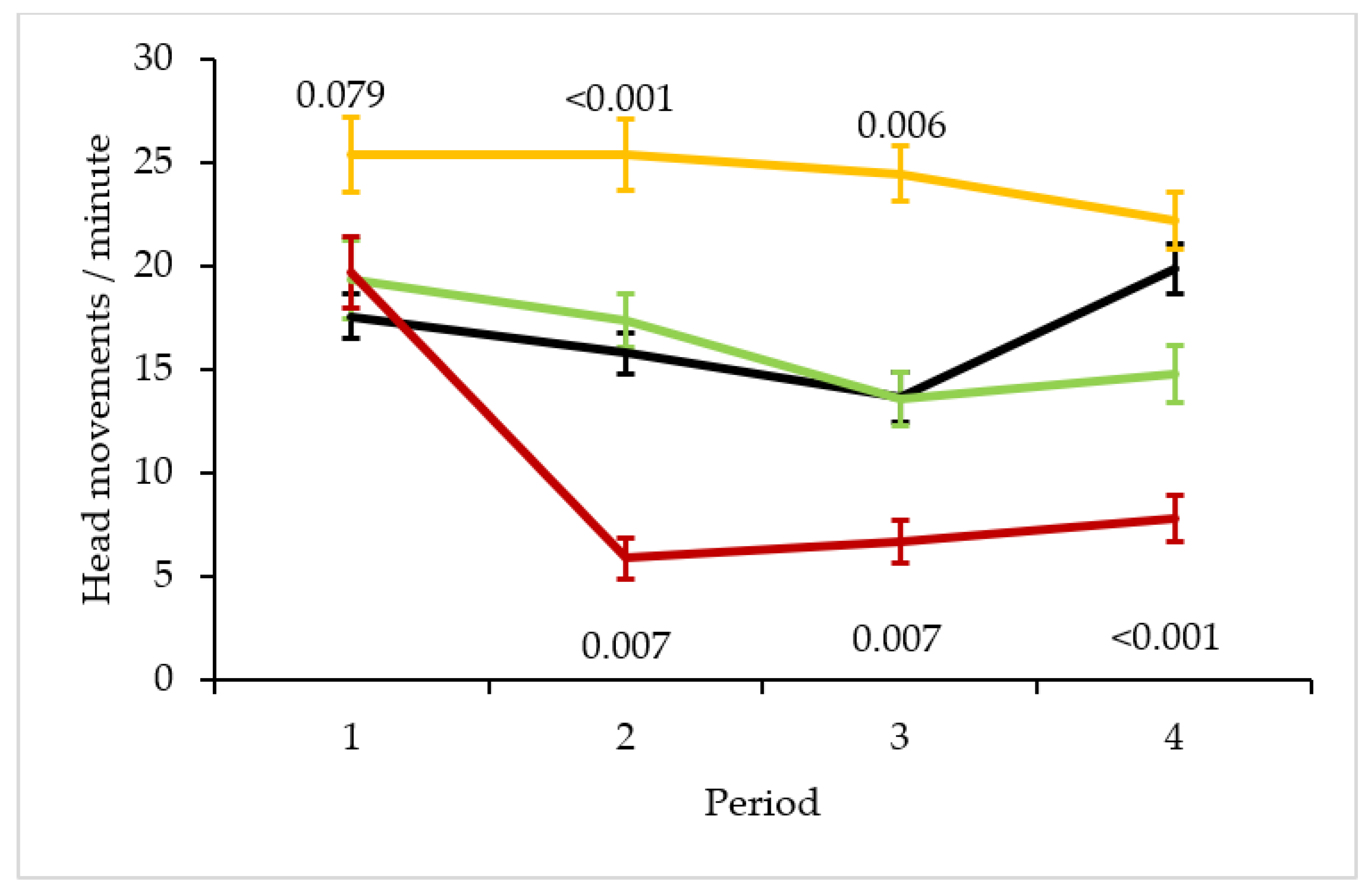
The best model for the time course of vigilance across all situations was the basic model plus the interaction terms between age class × period × situation, sex × period × situation, head colour morph × period × situation and partner head colour morph × period × situation (Table S2a). The main factor period was significant (Table 1) showing a decrease in vigilance over time. However, this varied between situations as the interaction term period × situation was significant (Table 1; Figure 2). Within the first 15 min vigilance was the same across all situations (familiar vs. novel t = −1.049, p = 0.295; familiar vs. changed (star at neutral location t = −0.674, p = 0.501) except for the changed situation with the novel object above the feeder, which showed a trend to be higher than the familiar situation (t = 1.766, p = 0.079). Vigilance in the familiar situation gradually decreased in periods 2 and 3 and then increased in period 4 to levels of the first period (GLMM for familiar situation: period 1 vs. 4 t = −0.927, p = 0.357). Vigilance in period 2 was significantly lower than in period 4 (t = −2.331, p = 0.023) with the same trend in period 3 (period 3 vs. 4 t = −1.721, p = 0.090). In contrast, vigilance in the novel situation plummeted to very low levels in period 2 and remained significantly lower than in the familiar situation throughout the rest of the experiment (familiar vs. novel environment, period 2: t = −2.722, p = 0.007, period 3: t = −2.702, p = 0.007, period 4: t = −4.962, p < 0.001). Period 1 showed a trend to be significantly higher than period 4 (GLMM for novel situation: t = 1.711, p = 0.092) with no differences between the remaining periods (period 2 vs. 4 t = 0.755, p = 0.453, period 3 vs. 4 t = −0.272, p = 0.786). Vigilance in the changed situation with the novel object above the feeder remained significantly higher throughout periods 2 and 3 as compared to the familiar situation and only decreased in period 4 (familiar situation vs. novel object above food, period 2: t = 4.072, p < 0.001, period 3: t = 2.775, p = 0.006, period 4: t = −1.023, p = 0.307). Period 1 to 3 were significantly higher than period 4 (GLMM for neophobia trial: period 1 vs. 4 t = 2.265, p = 0.027, period 2 vs. 4 t = 3.402, p < 0.001, period 3 vs. 4 t = 2.554, p = 0.013). Finally, vigilance in the changed situation with the star at the neutral location followed the same time course as vigilance in the familiar situation showing a gradual decline from period 1 to 3 (familiar situation vs. novel object at neutral location, period 2: t = −0.674, p = 0.501, period 3: t = 0.531, p = 0.596) and remained at this level in period 4 (t = −1.339, p = 0.185). Period 1 was significantly higher than period 4 with no differences between the remaining periods (GLMM for exploration trial: period 1 vs. 4 t = 2.699, p = 0.008, period 2 vs. 4 t = 0.926, p = 0.357, period 3 vs. 4 t = −1.456, p = 0.149).
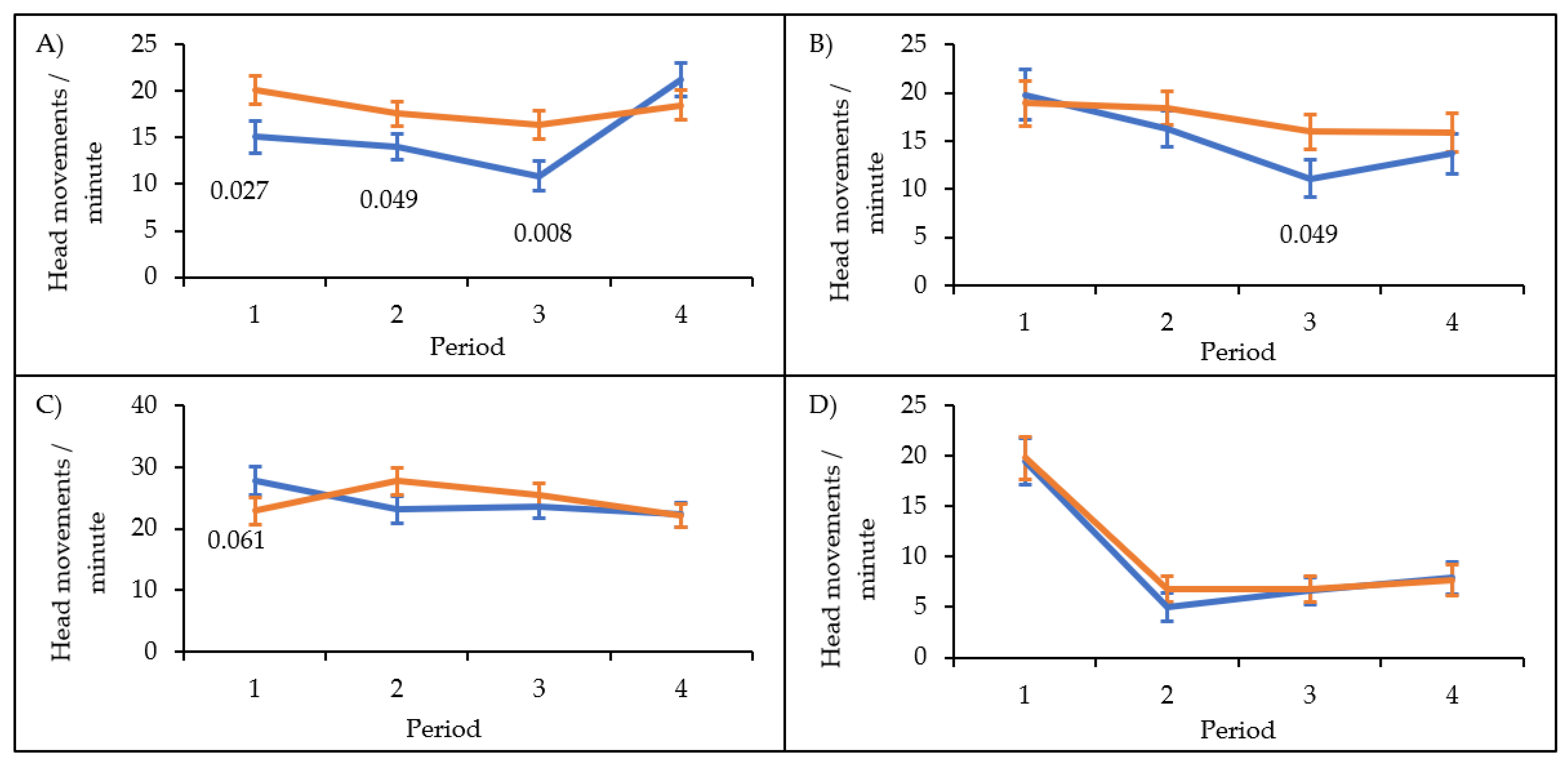
Finally, the interaction between sex × situation × period was significant (Table 1; Figure 3). In the familiar situation, males made fewer head movements than females in period 1 (t = −2.219, p = 0.027), 2 (t = −1.977, p = 0.049) and 3 (t = −2.695, p = 0.008) but not in period 4 (t = 1.179, p = 0.239). Males also made fewer head movements in period 3 in the changed situation with the star at a neutral location (t = −1.976, p = 0.049), whereas in the changed situation with the object above the feeder males tended to make more head movements than females in period 1 (t = 1.881, p = 0.061). Sexes did not differ in any of the other situations and periods (all p > 0.11). The random effect for individuals was not significant (z = 0.488, p = 0.626).
Vigilance was further compared in the exploration trial with the star at a neutral location considering birds that approached the change and those that did not. For the three best models, the Hessian matrix was not positive definite, although all convergence criteria were fulfilled. Results should therefore be treated with caution. The best model was the basic model with the interaction term age class × period (Table S2b). The main factor time period was significant (Table 2) with vigilance following the same time course as described above. There was a trend for birds that touched the object to show more head movements than birds that did not touch the object (Table 2; Figure 4). No other factors and interactions were significant (Table 2). The random effect individual was significant (z = 869.453, p < 0.001).
In the neophobia trial with the novel object above the feeder, vigilance was compared between those birds approaching the feeder and those who did not approach. Two equally best models came out (Table S2c); the basic model was the highest fit with the basic model plus the interaction term sex × period the second-best model, which was less than two AIC worse than the best model. The best model did not produce any significant outcomes (Table 3). However, in the second-best model the interaction term period × approach was significant (Table 3; Figure 5) with birds approaching the feeder making fewer head movements in period 1 than birds that did not approach (t = 2.341, p = 0.022), whereas no differences were found in any of the other periods (all p > 0.10). Furthermore, sex × period was significant (Table 3). Males made significantly more head movements in period 1 (t = 4.421, p < 0.001) but significantly fewer head movements in period 2 as compared to females (t = −2.169, p = 0.033). The random effect individual was significant (z = 2.329, p = 0.020).
4. Discussion
The time course of vigilance was investigated in four situations. Vigilance decreased over time, but the time course was situation specific. In the familiar situation and the changed situation with the star at a neutral location vigilance gradually decreased, whereas with the novel object above the feeder vigilance was consistently high only to decrease at the end. In contrast, in the novel environment vigilance plummeted to very low levels and only slightly recovered over time but never reached normal levels. Moreover, females were more vigilant than males in the familiar situation with sex differences disappearing the more threatening a situation became. Finally, individuals that explored and touched the star at the neutral position tended to be more vigilant than individuals that did not touch the star. In contrast, birds that approached the feeder with the novel object had initially lower vigilance than birds that did not approach the feeder.
Gouldian Finches showed a decrease in vigilance over time taking all situations together. While this result may be primarily driven by the strong decline in head movements in the novel environment, vigilance decreased to some extent in all situations. Vigilance decrement is generally observed in an unchanging environment due to lack of stimulation or decrease in arousal [31,34] and negatively affects the ability of an animal to detect a threat [32]. This was in part observed in the familiar situation with a gradual decline in head movements over time (periods 1–3) but a relapse to vigilance levels in the last period. Alternatively, these changes may reflect adjustment to perceived predation risk [13], although this is an unlikely explanation here as the birds were already several days in this environment and should have adjusted their vigilance to the perceived predation risk. However, the perceived risk may have been slightly increased at the start of the experiment due to the experimenter being in front of the cage when starting the camera, with a subsequent decrease in perceived risk as time passed by leading to an adjustment in vigilance. Whether the decline in vigilance is down to perceived risk adjustment or vigilance decrement needs further investigation. It is currently unclear why vigilance increased in the last period. Potential explanations could be linked to the circadian rhythm with birds becoming active again after a calmer period or systematic disturbance form outside (e.g., noise, which is unlikely as the animal facility is usually very quiet and a consistent disturbance at a particular time is needed to explain this result).
A similar time course was seen with the star positioned at a neutral location indicating that the novel object was not perceived as threatening and once introduced was not monitored more or for longer than any other object already present. This is consistent with [45], who found that scan frequency and duration did not change in marmosets (Callithrix penicillata) when a bear toy was introduced. Similarly, novelty of pictures did not affect vigilance decrement in humans [30]. However, there was a trend that vigilance differed between Gouldian Finches that approached and touched the novel object and those that did not with explorative individuals overall showing more head movements than less explorative individuals. This can be linked to personality. For example, higher vigilance has also been found in bold (more explorative, aggressive and dominant) Eurasian Siskins (Carduelis spinus) off-setting their risk-prone behaviour [46]. Likewise, a higher frequency in eye movements has been observed in humans with high perceptual curiosity, indicating that personality affects how a scene is assessed [47]. The higher frequency of head movements in explorative Gouldian Finches suggests that exploration does incur a cost. Individuals were not only monitoring the environment but were collecting information about the star, which resulted in overall more head movements. Exploration is generally seen as costly as it requires time and energy and deviates attention from the environment [43,48] but this is rarely investigated. Like in the siskin study [46], it seems that explorative individuals off-set the lack of attention by being more vigilant.
While the star at a neutral location only slightly affected vigilance and only those individuals that engaged with the object, the novel object above the feeder had a strong effect on vigilance in all birds. Vigilance overall was much higher than in any other situation and only returned to comparable levels in the familiar situation in the last period. This indicates that the object at this position was perceived as threatening, possibly due to uncertainty whether it is safe to approach the feeder or not [16]. An increase in vigilance in threatening situations has been found in many studies [11,15,17,19,20,21,49,50]. Likewise, a range of studies report that vigilance does not decrease with repeated presentation of a threat [7,28,51] and that constant attention is paid to high threats [52]. Interestingly, in studies investigating vigilance in relation to flight initiation distance (FID) there seems to be a dissociation between vigilance and responses to perceived threats. Here, vigilance is measured as the distance when a threat is detected (alert distance), whereas the actual FID is taken as a reflection of the perceived risk [21]. While vigilance (alert distance) varied with the perceived threat (higher vigilance in more disturbed areas) in marmots (Marmota flaviventer), it did not change over a 15-year period of ecotourism, whereas FID was lower in more disturbed areas and decreased over time indicating habituation to the threat [21]. Similar dissociations have been found in red squirrels (Sciurus vulgaris) [53], fox squirrels (S. niger) [20] and reindeer (Rangifer tarandus) [54]. Vigilance serves to allow detection of a range of threats, which requires consistent vigilance, whereas FID can be context-specific depending on the perceived threat. Samia et al. [55] found that costs of fleeing had the largest effect on escape decisions and questioned whereas selection on adjusting vigilance with perceived risk should be expected [56]. This challenges the model by Sirot and Pays [13], which would assume that vigilance decreases with repeated exposure. Interestingly vigilance was initially highest in the individuals that did not approach the feeder throughout the experiment indicating that those birds found the novel object more threatening than birds that later approached the feeder. This is in line with the concept of a risk-reward trade-off predicting that bold individuals are less vigilant and forgo safety for risk ([57,58,59] but see [46]). Nonetheless, all individuals seemed to have gathered information about the object over time [13,60], either directly though approaching the feeder or indirectly through observing others approaching. This reduced uncertainty and the level of thread perceived eventually leading to a reduction in vigilance.
Vigilance in the novel environment contrasted strongly with the response to a threat in the familiar environment as vigilance fell to a very low level after the first 15 min and stayed there throughout the experiment with only a slight recovery at the end. It seems the birds initially oriented themselves in and scanned the environment for any threats [39] with a frequency of head movements matching the one in the familiar environment. They then settled down and moved very little, a behaviour often observed in the wild when they arrive, e.g., at a waterhole to camouflage with the environment. In hamsters (Cricetus cricetus), females reduce vigilance under threat and freeze to preserve energy [12]. The reduced frequency of head movements in the Gouldian Finches suggests that they switched to a tracking strategy whereby they looked in a given direction for longer assessing this part of the environment in detail [8,39]. While vigilance slightly increased over time it consistently stayed way below their usual vigilance level indicating that the hour was not enough for vigilance to return to normal.
Finally, males made fewer head movements than females throughout most of the familiar situation (period 1–3) and in period 3 with the star at the neutral location, whereas there was a trend for males to be more vigilant in period 1 with the object above the feeder. Differences in vigilance between sexes are varied ranging from females being more vigilant [4,11,61] like in this study to no sex differences [10,19,62] to females being less vigilant [12]. Potentially, females were more vigilant as they could hear males showing heightened social vigilance [3,4]. Generally, sex differences seem to decrease with increasing threat (familiar > object at neutral location > object above feeder > novel environment). This could indicate that attention is diverted from social vigilance to the environment eliminating any sex differences.
5. Conclusions
In conclusion, the time course of vigilance is situation specific and likely reflects the perceived threat. Vigilance was highest and slow to return to normal when a change occurred close to a resource (object above feeder), little affected by changes away from important resources (object at neutral location) with some decrease over time and plummeted to very low levels without recovery in the novel environment representative of a tracking strategy. Moreover, females were more vigilant than males, but sex differences disappeared the more threatening the situation became. Vigilance seemed to differ between birds that engaged in exploration and those that did not indicating that explorative individuals increased vigilance. Future research may investigate whether vigilance linearly changes over time due to continuous accumulation of information or whether it changes more in increments as a function of behavioural actions, e.g., approaching a novel object.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/birds4010001/s1, Table S1: Data set; Table S2: Model selection outcomes from GLMM using Akaike criterion.
Funding
This research received no external funding.
Institutional Review Board Statement
The study was conducted according to the guidelines of the Declaration of Helsinki, and approved by the Institutional Review Board (or Ethics Committee) of Liverpool John Moores University (29 November 2017).
Data Availability Statement
Data are available as a supplementary to the paper.
Acknowledgments
I would like to thank Anna Temple for collecting some of the data for this study, Georgina Eccles for helping with the data collection, Blattner Heimtierfutter for sponsoring all bird food, Peter McGough and other breeders for donating some of the Gouldian Finches, the animal care technicians for looking after the birds and two anonymous reviewers for the constructive feedback.
Conflicts of Interest
The author declares no conflict of interest.
References
- Paus, T.; Zatorre, R.J.; Hofle, N.; Caramanos, Z.; Gotman, J.; Petrides, M.; Evans, A.C. Time-related changes in neural systems underlying attention and arousal during the performance of an auditory vigilance task. J. Cogn. Neurosci. 1997, 9, 392–408. [Google Scholar] [CrossRef] [PubMed]
- Lea, A.J.; Blumstein, D.T. Age and sex influence marmot antipredator behavior during periods of heightened risk. Behav. Ecol. Sociobiol. 2011, 65, 1525–1533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pecorella, I.; Fattorini, N.; Macchi, E.; Ferretti, F. Sex/age differences in foraging, vigilance and alertness in a social herbivore. Acta Ethol. 2019, 22, 1–8. [Google Scholar] [CrossRef]
- Han, L.; Blank, D.; Wang, M.; Yanget, W. Vigilance behaviour in Siberian ibex (Capra sibirica): Effect of group size, group type, sex and age. Behav Proc. 2020, 170, 104021. [Google Scholar] [CrossRef] [PubMed]
- Baker, D.J.; Stillman, R.A.; Smart, S.L.; Bullock, J.M.; Norris, K.J. Are the costs of routine vigilance avoided by granivorous foragers? Func. Ecol. 2011, 25, 617–627. [Google Scholar] [CrossRef]
- Saltz, D.; Berger-Tal, O.; Motro, U.; Shkedy, Y.; Raanan, N. Conservation implications of habituation in Nubian ibex in response to ecotourism. Anim. Cons 2019, 22, 220–227. [Google Scholar] [CrossRef]
- Scheijen, C.P.J.; van der Merwe, S.; Ganswindt, A.; Deacon, F. Anthropogenic influences on distance travelled and vigilance behavior and stress-related endocrine correlates in free-roaming giraffes. Animals 2021, 11, 1239. [Google Scholar] [CrossRef]
- Mettke-Hofmann, C. Morph Composition Matters in the Gouldian Finch (Chloebia gouldiae): Involvement of Red-Headed Birds Increases Vigilance. Birds 2021, 2, 404–414. [Google Scholar] [CrossRef]
- Monclus, R.; Roedel, H.G.; von Holst, D. Fox odour increases vigilance in European rabbits: A study under semi-natural conditions. Ethology 2006, 112, 1186–1193. [Google Scholar] [CrossRef]
- Sauter, C.; Danker-Hopfe, H.; Loretz, E.; Zeitlhofer, J.; Geisler, P.; Popp, R. The assessment of vigilance: Normative data on the Siesta sustained attention test. Sleep Med. 2013, 14, 542–548. [Google Scholar] [CrossRef]
- Hume, G.; Brunton, E.; Burnett, S. Eastern grey kangaroo (Macropus giganteus) vigilance behaviour varies between human-modified and natural environments. Animals 2019, 9, 494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flamand, A.; Rebout, N.; Bordes, C.; Guinnefollau, L.; Berges, M.; Ajak, F.; Siutz, C.; Millesi, E.; Weber, C.; Petit, O. Hamsters in the city: A study on the behaviour of a population of common hamsters (Cricetus cricetus) in urban environment. PLoS ONE 2019, 14, e0225347. [Google Scholar] [CrossRef] [PubMed]
- Sirot, E.; Pays, O. On the dynamics of predation risk perception for a vigilant forager. J. Theor. Biol. 2011, 276, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandez-Juricic, E. Sensory basis of vigilance behavior in birds: Synthesis and future prospects. Behav. Proc. 2012, 89, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Welp, T.; Rushen, J.; Kramer, D.L.; Festa-Bianchet, M.; de Passille, A.M.B. Vigilance as a measure of fear in dairy cattle. Appl. Anim. Behav. Sci 2004, 87, 1–13. [Google Scholar] [CrossRef]
- Feyten, L.E.A.; Brown, G.E. Ecological uncertainty influences vigilance as a marker of fear. Anim. Sent. 2018, 15, 7. [Google Scholar] [CrossRef]
- Goldenberg, S.U.; Borcherding, J.; Heynen, M. Balancing the response to predation—The effects of shoal size, predation risk and habituation on behaviour of juvenile perch. Behav. Ecol. Sociobiol. 2014, 68, 989–998. [Google Scholar] [CrossRef]
- Poudel, B.S.; Spooner, P.G.; Matthews, A. Behavioural changes in marmots in relation to livestock grazing disturbance: An experimental test. Eur. J. Wildl. Res. 2016, 62, 491–495. [Google Scholar] [CrossRef]
- Montero-Quintana, A.N.; Vazquez-Haikin, J.A.; Merkling, T.; Blanchard, P.B.; Osorio-Beristain, M. Ecotourism impacts on the behaviour of whale sharks: An experimental approach. Oryx 2020, 54, 270–275. [Google Scholar] [CrossRef] [Green Version]
- Kittendorf, A.; Dantzer, B. Urban fox squirrels exhibit tolerance to humans but respond to stimuli from natural predators. Ethology 2021, 127, 697–709. [Google Scholar] [CrossRef]
- Uchida, K.; Blumstein, D.T. Habituation or sensitization? Long-term responses of yellow-bellied marmots to human disturbance. Behav. Ecol. 2021, 32, 668–678. [Google Scholar] [CrossRef]
- Carbone, C.; Thompson, W.A.; Zadorina, L.; Rowcliffe, J.M. Competition, predation risk and patterns of flock expansion in barnacle geese (Branta leucopsis). J. Zool. 2003, 259, 301–308. [Google Scholar] [CrossRef]
- Beale, C.M.; Monaghan, P. Behavioural responses to human disturbance: A matter of choice? Anim. Behav. 2004, 68, 1065–1069. [Google Scholar] [CrossRef]
- Barros, M.; Alencar, C.; de Souza Silva, M.A.; Tomaz, C. Changes in experimental conditions alter anti-predator vigilance and sequence predictability in captive marmosets. Behav. Proc. 2008, 77, 351–356. [Google Scholar] [CrossRef]
- Dupuch, A.; Morris, D.W.; Halliday, W.D. Patch use and vigilance by sympatric lemmings in predator and competitor-driven landscapes of fear. Behav. Ecol. Sociobiol. 2014, 68, 299–308. [Google Scholar] [CrossRef]
- Kautz, T.M.; Beyer, D.E., Jr.; Farley, Z.; Fowler, N.L.; Kellner III, K.F.; Lutto, A.L.; Petroelje, T.R.; Belant, J.L. American martens use vigilance and short-term avoidance to navigate a landscape of fear from fishers at artificial scavenging sites. Sci. Rep. 2021, 11, 12146. [Google Scholar] [CrossRef]
- Vallino, C.; Caprio, E.; Genco, F.; Chamberlain, D.; Palestrini, C.; Roggero, A.; Bocca, M.; Rolando, A. Behavioural responses to human disturbance in an alpine bird. J. Orn. 2019, 160, 763–772. [Google Scholar] [CrossRef] [Green Version]
- Hammer, T.L.; Bize, P.; Saraux, C.; Gineste, B.; Robin, J.-P.; Groscolas, R.; Viblanc, V.A. Repeatability of alert and flight initiation distances in king penguins: Effects of colony, approach speed, and weather. Ethology 2022, 128, 303–316. [Google Scholar] [CrossRef]
- Meeker, T.J.; Emerson, N.M.; Chien, J.-H.; Saffer, M.I.; Bienvenu, O.J.; Korzeniewska, A.; Greenspan, J.D.; Lenz, F.A. During vigilance to painful stimuli: Slower response rate is related to high trait anxiety, whereas faster response rate is related to high state anxiety. J. Neurophysiol. 2021, 125, 305–319. [Google Scholar] [CrossRef]
- Jun, J.; Remington, R.W.; Koutstaal, W.; Jiang, Y.V. Characteristics of sustaining attention in a gradual-onset continuous performance task. J. Exp. Psychol. Human Perc. Perf. 2019, 45, 386–401. [Google Scholar] [CrossRef]
- Siddle, D.A.T. Vigilance decrement and speed of habituation of the GSR component of the orienting response. Br. J. Psychol. 1972, 63, 191–194. [Google Scholar] [CrossRef] [PubMed]
- Humphrey, B.; Helton, W.S.; Bedoya, C.; Dolev, Y.; Nelson, X.J. Psychophysical investigation of vigilance decrement in jumping spiders: Overstimulation or understimulation? Anim. Cogn. 2018, 21, 787–794. [Google Scholar] [CrossRef] [PubMed]
- Martin, J.T.; Whittaker, A.H.; Johnston, S.J. Pupillometry and the vigilance decrement: Task-evoked but not baseline pupil measures reflect declining performance in visual vigilance tasks. Eur. J. Neurosci. 2022, 55, 778–799. [Google Scholar] [CrossRef] [PubMed]
- Melrose, A.; Nelson, X.J.; Dolev, Y.; Helton, W.S. Vigilance all the way down: Vigilance decrement in jumping spiders resembles that of humans. Q. J. Exp. Psychol. 2019, 72, 1530–1538. [Google Scholar] [CrossRef] [PubMed]
- Terhune, J.M.; Brilliant, S.W. Harbour seal vigilance decreases over time since haul out. Anim. Behav. 1996, 51, 757–763. [Google Scholar] [CrossRef] [Green Version]
- Beauchamp, G.; Ruxton, G.D. Vigilance Decreases with Time at Loafing Sites in Gulls (Larus spp.). Ethology 2012, 118, 733–739. [Google Scholar] [CrossRef]
- Dostine, P.L.; Johnson, G.C.; Franklin, D.C.; Zhang, Y.; Hempel, C. Seasonal use of savanna landscapes by the Gouldian finch, Erythrura gouldiae, in the Yinberrie Hills area, Northern Territory. Wildl. Res. 2001, 28, 445–458. [Google Scholar] [CrossRef]
- Brush, A.H.; Seifried, H. Pigmentation and feather structure in genetic variants of the Gouldian finch, Poephila gouldiae. Auk Ornithol. Adv. 1968, 85, 416–430. [Google Scholar] [CrossRef]
- Fernandez-Juricic, E.; Gall, M.D.; Dolan, T.; O’Rourke, C.; Thomas, S.; Lynch, J. Visual systems and vigilance behaviour of two ground-foraging avian prey species: White-crowned sparrows and California towhees. Anim. Behav. 2011, 81, 705–713. [Google Scholar] [CrossRef]
- Jones, K.A.; Krebs, J.R.; Whittingham, M.J. Vigilance in the third dimension: Head movement not scan duration varies in response to different predator models. Anim. Behav. 2007, 74, 1181–1187. [Google Scholar] [CrossRef]
- Fernandez-Juricic, E.; Beauchamp, G.; Treminio, R.; Hoover, M. Making heads turn: Association between head movements during vigilance and perceived predation risk in brown-headed cowbird flocks. Anim. Behav. 2011, 82, 573–577. [Google Scholar] [CrossRef]
- Krebs, H.; Weyers, P.; Macht, M.; Weijers, H.-G.; Janke, W. Scanning behavior of rats during eating under stressful noise. Physiol. Behav. 1997, 62, 151–154. [Google Scholar] [CrossRef] [PubMed]
- Mettke-Hofmann, C.; Winkler, H.; Leisler, B. The significance of ecological factors for exploration and neophobia in parrots. Ethology 2002, 108, 249–272. [Google Scholar] [CrossRef]
- ASAB. Guidelines for the treatment of animals in behavioural research and teaching. Anim. Behav. 2020, 159, i–xi. [Google Scholar] [CrossRef]
- Dacier, A.; Maia, R.; Agustinho, D.P.; Barros, M. Rapid habituation of scan behavior in captive marmosets following brief predator encounters. Behav. Proc. 2006, 71, 66–69. [Google Scholar] [CrossRef]
- Pascal, J.; Senar, J.C. Antipredator behavioural compensation of proactive personality trait in male Eurasian siskins. Anim. Behav. 2014, 90, 297–303. [Google Scholar] [CrossRef]
- Risko, E.F.; Anderson, N.C.; Lanthier, S.; Kingstone, A. Curious eyes: Individual differences in personality predict eye movement behavior in scene-viewing. Cogn. 2012, 122, 86–90. [Google Scholar] [CrossRef]
- Sol, D.; Griffin, A.S.; Bartomeus, I.; Boyce, H. Exploring or Avoiding Novel Food Resources? The Novelty Conflict in an Invasive Bird. PLoS ONE 2011, 6, e19535. [Google Scholar] [CrossRef] [Green Version]
- Barros, M.; de Souza Silva, M.A.; Huston, J.P.; Tomaz, C. Multibehavioral analysis of fear and anxiety before, during, and after experimentally induced predatory stress in Callithrix penicillate. Pharm. Biochem. Behav. 2004, 78, 357–367. [Google Scholar] [CrossRef]
- Atkins, A.; Little, R.B.; Redpath, S.M.; Amar, A. Impact of increased predation risk on vigilance behaviour in a gregarious waterfowl, the Egyptian goose Alopochen aegyptiana. J. Avian Biol. 2019, 50, e02121. [Google Scholar] [CrossRef]
- Biedenweg, T.A.; Parsons, M.H.; Fleming, P.A.; Blumstein, D.T. Sounds scary? Lack of habituation following the presentation of novel sounds. PLoS ONE 2011, 6, e14549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rinck, M.; Becker, E.S. Spider fearful individuals attend to threat, then quickly avoid it: Evidence from eye movements. J. Abnorm. Psychol. 2006, 115, 231–238. [Google Scholar] [CrossRef] [PubMed]
- Uchida, K.; Suzuki, K.K.; Shimamoto, T.; Yanagawa, H.; Koizumi, I. Decreased vigilance or habituation to humans? Mechanisms on increased boldness in urban animals. Behav. Ecol. 2019, 30, 1583–1590. [Google Scholar] [CrossRef]
- Reimers, E.; Loe, L.E.; Eftestol, S.; Colman, J.E.; Dahle, B. Effects of hunting on response behaviors of wild reindeer. J. Wildl. Manag. 2009, 73, 844–851. [Google Scholar] [CrossRef]
- Samia, D.S.M.; Blumstein, D.T.; Stankowich, T.; Cooper, W.E., Jr. Fifty years of chasing lizards: New insights advance optimal escape theory. Biol. Rev. 2016, 91, 349–366. [Google Scholar] [CrossRef]
- Samia, D.S.M.; Blumstein, D.T.; Díaz, M.; Grim, T.; Ibanez-Alamo, J.D.; Jokimäki, J.; Tätte, K.; Marko, G.; Tryjanowski, P.; Moeller, A.P. Rural-urban differences in escape behavior of European birds across a latitudinal gradient. Front. Ecol. Evol. 2017, 5, 66. [Google Scholar] [CrossRef] [Green Version]
- Bergvall, U.A.; Schäpers, A.; Kjellander, P.; Weiss, A. Personality and foraging decisions in fallow deer, Dama dama. Anim. Behav. 2011, 81, 101–112. [Google Scholar] [CrossRef]
- Couchoux, C.; Cresswell, W. Personality constraints versus flexible antipredation behaviors: How important is boldness in risk management of redshanks (Tringa totanus) foraging in a natural system? Behav. Ecol. 2011, 23, 290–301. [Google Scholar] [CrossRef] [Green Version]
- Mazza, V.; Jacob, J.; Dammhahn, M.; Zaccaroni, M.; Eccard, J.A. Individual variation in cognitive style reflects foraging and antipredator strategies in a small mammal. Sci. R. 2019, 9, 10157. [Google Scholar] [CrossRef] [Green Version]
- Perez-Duenas, C.; Acosta, A.; Lupianez, J. Reduced habituation to angry faces: Increased attentional capture as to override inhibition of return. Psychol. Res. 2014, 78, 196–208. [Google Scholar] [CrossRef]
- Childress, M.J.; Lung, M.A. Predation risk, gender and the group size effect: Does elk vigilance depend upon the behaviour of conspecifics? Anim. Behav. 2003, 66, 389–398. [Google Scholar] [CrossRef]
- Giambra, L.M.; Quilter, R.E. Sex differences in sustained attention across the adult life span. J. Appl. Psychol. 1989, 74, 91–95. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Novel objects in the two changed situations. (A) Star at a neutral location above a perch (exploration trial); (B) Stocking sock above the feeder (neophobia trial).
Figure 1.
Novel objects in the two changed situations. (A) Star at a neutral location above a perch (exploration trial); (B) Stocking sock above the feeder (neophobia trial).

Figure 2.
Time course of vigilance across four situations. Mean and standard errors of frequency of head movements are shown across 15-min blocks in the familiar situation (black line), in the changed situations with the novel object at a neutral location (exploration trial; green line) and with the novel object above the feeder (neophobia trial; yellow line) and in a novel environment (red line). Significance levels are in relation to the familiar situation.
Figure 2.
Time course of vigilance across four situations. Mean and standard errors of frequency of head movements are shown across 15-min blocks in the familiar situation (black line), in the changed situations with the novel object at a neutral location (exploration trial; green line) and with the novel object above the feeder (neophobia trial; yellow line) and in a novel environment (red line). Significance levels are in relation to the familiar situation.

Figure 3.
Time course of vigilance for males and females in four situations. Mean and standard errors of frequency of head movements are shown across 15-min blocks for males (blue) and females (orange) in (A) the familiar situation, (B) the changed situation with the novel object at a neutral location (exploration trial), (C) the changed situation with the novel object above the feeder (neophobia trial) and (D) the novel environment.
Figure 3.
Time course of vigilance for males and females in four situations. Mean and standard errors of frequency of head movements are shown across 15-min blocks for males (blue) and females (orange) in (A) the familiar situation, (B) the changed situation with the novel object at a neutral location (exploration trial), (C) the changed situation with the novel object above the feeder (neophobia trial) and (D) the novel environment.

Figure 4.
Time course of vigilance in the exploration trial for birds that touched the object and those that did not. Mean and standard errors of frequency of head movements (transformed) are shown across 15-min blocks for birds that touched the novel object (solid line) and those that did not (dashed line).
Figure 4.
Time course of vigilance in the exploration trial for birds that touched the object and those that did not. Mean and standard errors of frequency of head movements (transformed) are shown across 15-min blocks for birds that touched the novel object (solid line) and those that did not (dashed line).

Figure 5.
Time course of vigilance in the neophobia trial for birds that approached the feeder and those that did not. Mean and standard errors of frequency of head movements (transformed) are shown across 15-min blocks for birds that approached the feeder or landed on it (solid line) and those that did not (dashed line).
Figure 5.
Time course of vigilance in the neophobia trial for birds that approached the feeder and those that did not. Mean and standard errors of frequency of head movements (transformed) are shown across 15-min blocks for birds that approached the feeder or landed on it (solid line) and those that did not (dashed line).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
General Linear Mixed Model outcome of best model for the time course of vigilance over four fixed 15-min periods in four situations.
Table 1.
General Linear Mixed Model outcome of best model for the time course of vigilance over four fixed 15-min periods in four situations.
| Variables | F | Df1 | Df2 | p |
|---|---|---|---|---|
| Corrected model | 10.182 | 95 | 256 | <0.001 |
| Period | 14.066 | 3 | 256 | <0.001 |
| Period × situation | 41.122 | 12 | 256 | <0.001 |
| Age class × period × situation | 1.364 | 32 | 256 | 0.100 |
| Sex × period × situation | 2.090 | 16 | 256 | 0.009 |
| Colour morph × period × situation | 1.128 | 16 | 256 | 0.329 |
| Partner colour morph × period × situation | 0.790 | 16 | 256 | 0.696 |
Table 2.
General Linear Mixed Model outcome of best model for the time course of vigilance over four fixed 15-min periods in the exploration trial comparing birds that touched the object and those that did not.
Table 2.
General Linear Mixed Model outcome of best model for the time course of vigilance over four fixed 15-min periods in the exploration trial comparing birds that touched the object and those that did not.
| Variables | F | Df1 | Df2 | p |
|---|---|---|---|---|
| Corrected model | 1.897 | 15 | 70 | 0.038 |
| Period | 5.847 | 3 | 70 | 0.001 |
| Approach | 3.311 | 1 | 70 | 0.073 |
| Period × approach | 0.044 | 3 | 70 | 0.988 |
| Age class × period | 1.397 | 8 | 70 | 0.213 |
Table 3.
General Linear Mixed Model outcome of the two best models for the time course of vigilance over four fixed 15-min periods in the neophobia trial comparing birds that approached the feeder in reach or landed on it and those that did not. The best and the second-best model were less than two AIC different from each other.
Table 3.
General Linear Mixed Model outcome of the two best models for the time course of vigilance over four fixed 15-min periods in the neophobia trial comparing birds that approached the feeder in reach or landed on it and those that did not. The best and the second-best model were less than two AIC different from each other.
| Best Model | ||||
|---|---|---|---|---|
| Variables | F | Df1 | Df2 | p |
| Corrected model | 1.161 | 6 | 81 | 0.336 |
| Period | 1.325 | 3 | 81 | 0.272 |
| Approach | 0.072 | 1 | 81 | 0.789 |
| Period × approach | 0.873 | 2 | 81 | 0.422 |
| Second best model | ||||
| Corrected model | 4.211 | 10 | 77 | <0.001 |
| Period | 1.642 | 3 | 77 | 0.187 |
| Approach | 0.494 | 1 | 77 | 0.484 |
| Period × approach | 4.087 | 2 | 77 | 0.021 |
| Sex × period | 9.213 | 4 | 77 | <0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Mettke-Hofmann, C. When to Return to Normal? Temporal Dynamics of Vigilance in Four Situations. Birds 2023, 4, 1-14. https://doi.org/10.3390/birds4010001
AMA Style
Mettke-Hofmann C. When to Return to Normal? Temporal Dynamics of Vigilance in Four Situations. Birds. 2023; 4(1):1-14. https://doi.org/10.3390/birds4010001
Chicago/Turabian StyleMettke-Hofmann, Claudia. 2023. "When to Return to Normal? Temporal Dynamics of Vigilance in Four Situations" Birds 4, no. 1: 1-14. https://doi.org/10.3390/birds4010001