
How Safe Is Gadobutrol? Examining the Effect of Gadolinium Deposition on the Nervous System

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
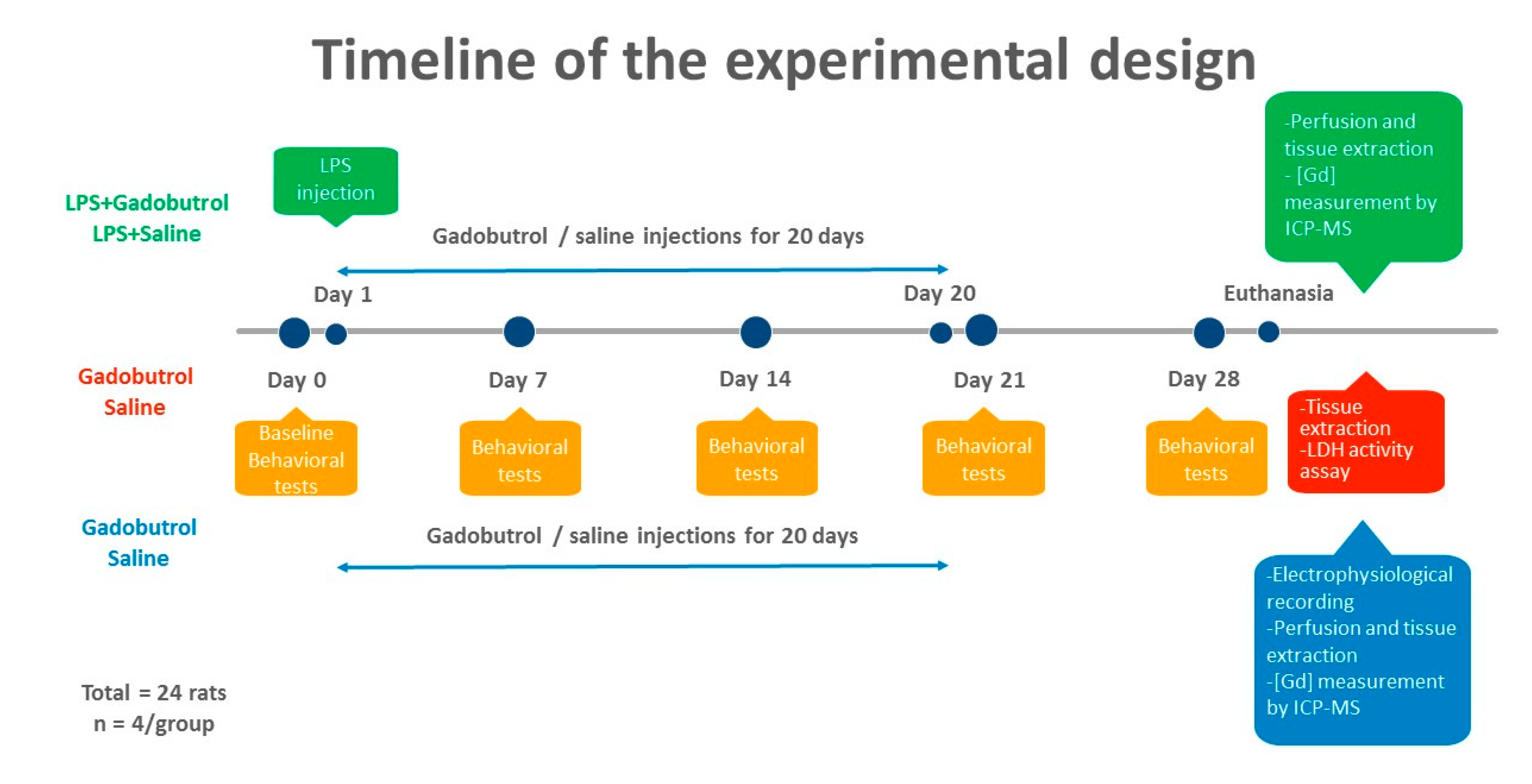
2.2. Treatment Regimen
2.3. Animal Perfusion
2.4. Quantification of Gadolinium Using ICP-MS
2.5. LDH Assay
2.6. Behavioral Tests
2.6.1. Heat Sensitivity Test
2.6.2. Beam Walking Test
2.6.3. Spontaneous Alternation T-Maze Test
2.7. Electrophysiological Assessment
2.8. Statistical Analysis
3. Results
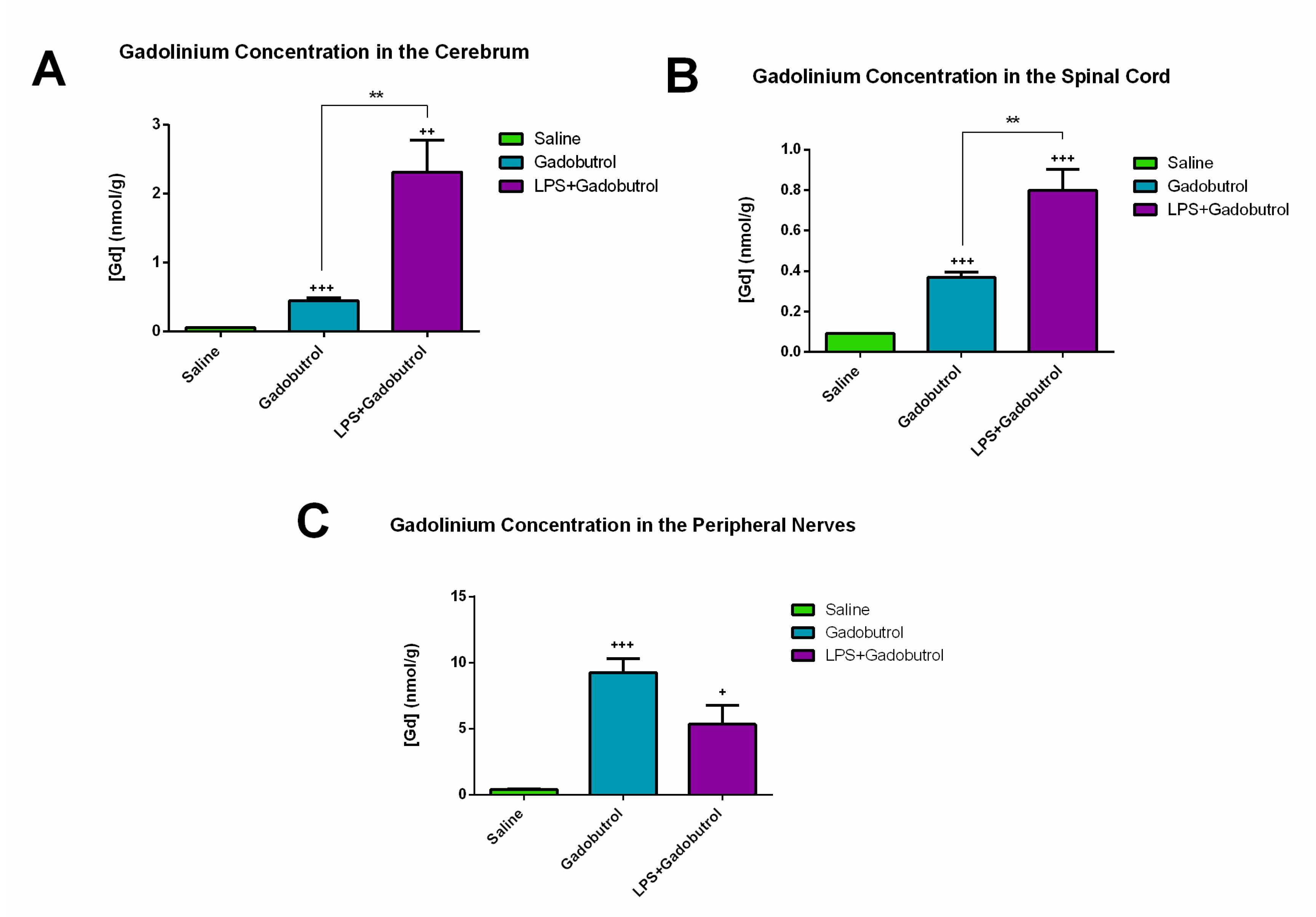
3.1. Quantification of Gadolinium Using ICP-MS
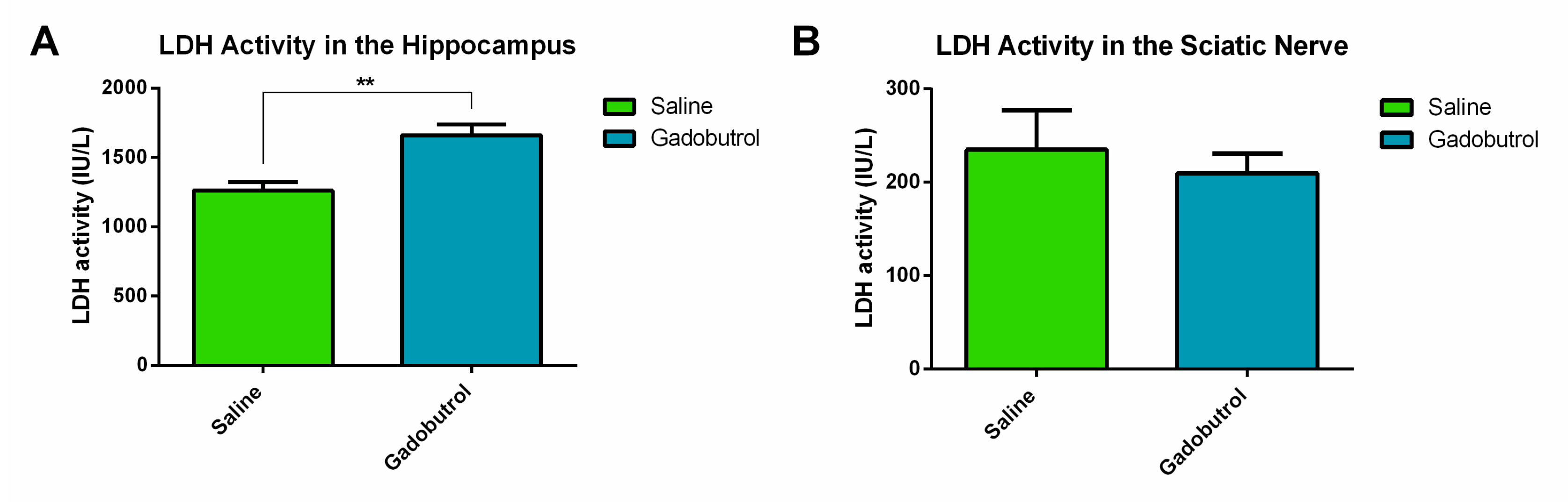
3.2. LDH Assay
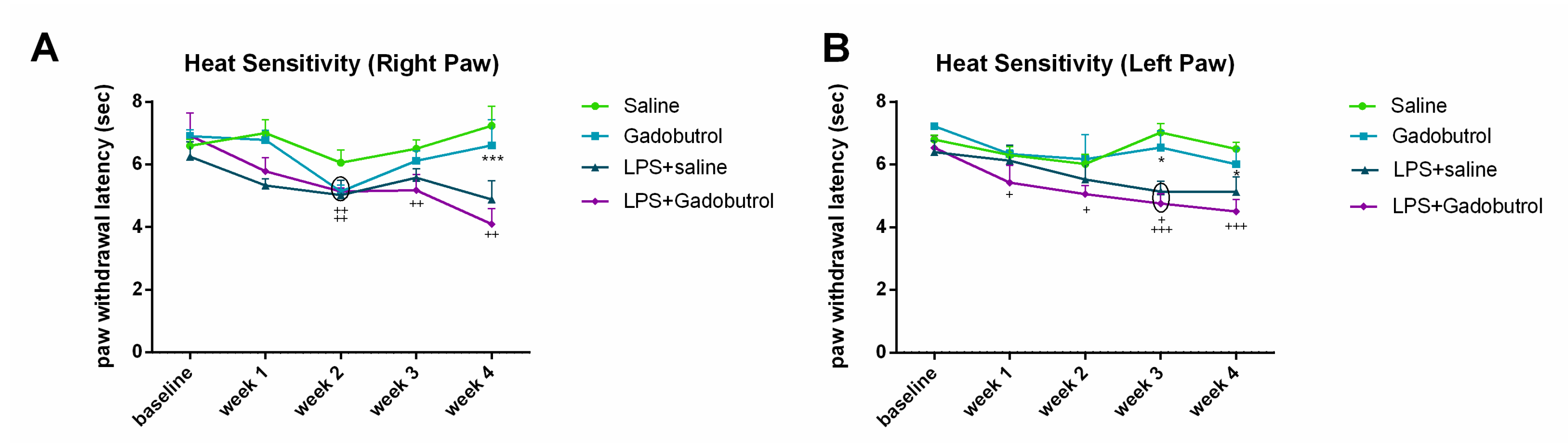
3.3. Heat Sensitivity Test
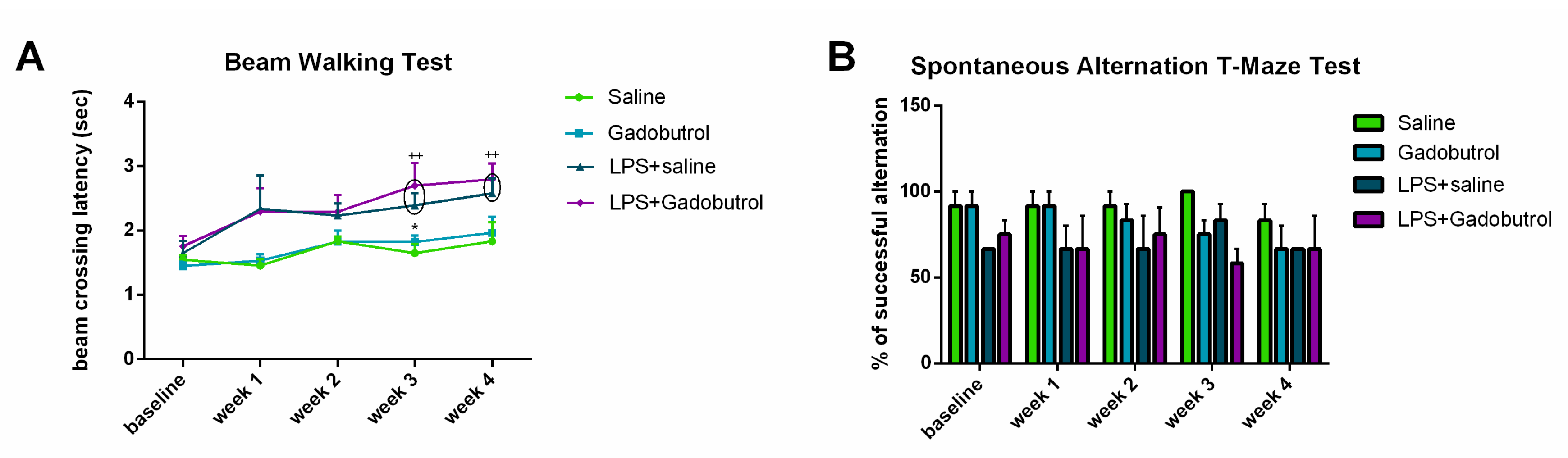
3.4. Beam Walking Test
3.5. Spontaneous Alternation T-Maze Test
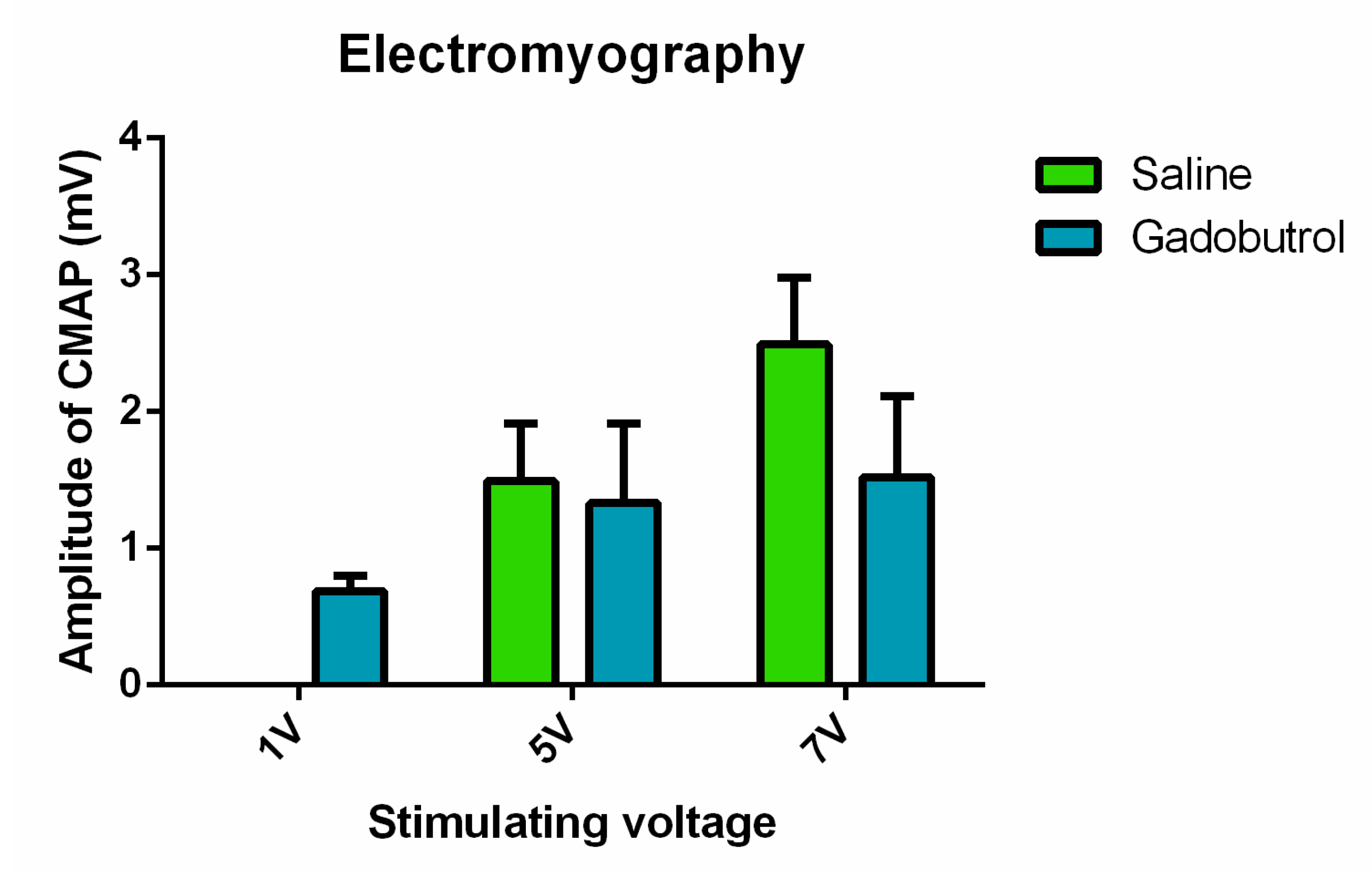
3.6. Electrophysiological Assessment
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Botta, M. Second coordination sphere water molecules and relaxivity of gadolinium (III) complexes: Implications for MRI contrast agents. Eur. J. Inorg. Chem. 2000, 2000, 399–407. [Google Scholar] [CrossRef]
- Davies, J.; Siebenhandl-Wolff, P.; Tranquart, F.; Jones, P.; Evans, P. Gadolinium: Pharmacokinetics and toxicity in humans and laboratory animals following contrast agent administration. Arch. Toxicol. 2022, 96, 403–429. [Google Scholar] [CrossRef] [PubMed]
- McDonald, R.J.; McDonald, J.S.; Dai, D.; Schroeder, D.; Jentoft, M.E.; Murray, D.L.; Kadirvel, R.; Eckel, L.J.; Kallmes, D.F. Comparison of gadolinium concentrations within multiple rat organs after intravenous administration of linear versus macrocyclic gadolinium chelates. Radiology 2017, 285, 536–545. [Google Scholar]
- Alkhunizi, S.M.; Fakhoury, M.; Abou-Kheir, W.; Lawand, N. Gadolinium Retention in the Central and Peripheral Nervous System: Implications for Pain, Cognition, and Neurogenesis. Radiology 2020, 297, 407–416. [Google Scholar] [CrossRef] [PubMed]
- Erdene, K.; Nakajima, T.; Kameo, S.; Khairinisa, M.A.; Lamid-Ochir, O.; Tumenjargal, A.; Koibuchi, N.; Koyama, H.; Tsushima, Y. Organ retention of gadolinium in mother and pup mice: Effect of pregnancy and type of gadolinium-based contrast agents. Jpn. J. Radiol. 2017, 35, 568–573. [Google Scholar] [CrossRef]
- Khairinisa, M.A.; Takatsuru, Y.; Amano, I.; Erdene, K.; Nakajima, T.; Kameo, S.; Koyama, H.; Tsushima, Y.; Koibuchi, N. The Effect of Perinatal Gadolinium-Based Contrast Agents on Adult Mice Behavior. Investig. Radiol. 2018, 53, 110–118. [Google Scholar] [CrossRef]
- Saupe, N.; Zanetti, M.; Pfirrmann, C.W.; Wels, T.; Schwenke, C.; Hodler, J. Pain and other side effects after MR arthrography: Prospective evaluation in 1085 patients. Radiology 2009, 250, 830–838. [Google Scholar] [CrossRef] [Green Version]
- Semelka, R.C.; Commander, C.W.; Jay, M.; Burke, L.M.; Ramalho, M. Presumed gadolinium toxicity in subjects with normal renal function: A report of 4 cases. Investig. Radiol. 2016, 51, 661–665. [Google Scholar] [CrossRef]
- Ayers-Ringler, J.; McDonald, J.S.; Connors, M.A.; Fisher, C.R.; Han, S.; Jakaitis, D.R.; Scherer, B.; Tutor, G.; Wininger, K.M.; Dai, D.; et al. Neurologic Effects of Gadolinium Retention in the Brain after Gadolinium-based Contrast Agent Administration. Radiology 2022, 302, 676–683. [Google Scholar] [CrossRef]
- Scott, L.J. Gadobutrol: A Review in Contrast-Enhanced MRI and MRA. Clin. Drug Investig. 2018, 38, 773–784. [Google Scholar] [CrossRef] [Green Version]
- Kaunzner, U.W.; Gauthier, S.A. MRI in the assessment and monitoring of multiple sclerosis: An update on best practice. Ther. Adv. Neurol. Disord. 2017, 10, 247–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- FDA. Gadavist (Gadobutrol) Injection. Available online: https://www.accessdata.fda.gov/drugsatfda_docs/label/2011/201277s000lbl.pdf (accessed on 1 August 2021).
- Port, M.; Idée, J.-M.; Medina, C.; Robic, C.; Sabatou, M.; Corot, C. Efficiency, thermodynamic and kinetic stability of marketed gadolinium chelates and their possible clinical consequences: A critical review. Biometals 2008, 21, 469–490. [Google Scholar] [CrossRef] [PubMed]
- Glutig, K.; Hahn, G.; Kuvvetli, P.; Endrikat, J. Safety of gadobutrol: Results of a non-interventional study of 3710 patients, including 404 children. Acta Radiol. 2019, 60, 873–879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Endrikat, J.; Schwenke, C.; Prince, M. Gadobutrol for contrast-enhanced magnetic resonance imaging in elderly patients: Review of the safety profile from clinical trial, post-marketing surveillance, and pharmacovigilance data. Clin. Radiol. 2015, 70, 743–751. [Google Scholar] [CrossRef] [Green Version]
- Ozturk, K.; Nascene, D. Effect of at least 10 serial gadobutrol administrations on brain signal intensity ratios on T1-weighted MRI in children: A matched case-control study. Am. J. Roentgenol. 2021, 217, 753–760. [Google Scholar] [CrossRef] [PubMed]
- Mlinar, B.; Enyeart, J. Block of current through T-type calcium channels by trivalent metal cations and nickel in neural rat and human cells. J. Physiol. 1993, 469, 639–652. [Google Scholar] [CrossRef]
- Green, C.; Jost, G.; Frenzel, T.; Boyken, J.; Schwenke, C.; Pietsch, H. The Effect of Gadolinium-Based Contrast Agents on Longitudinal Changes of Magnetic Resonance Imaging Signal Intensities and Relaxation Times in the Aging Rat Brain. Investig. Radiol. 2022, 57, 453–462. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.; Xia, Q.; Yuan, L.; Yang, X.; Wang, K. Impaired mitochondrial function and oxidative stress in rat cortical neurons: Implications for gadolinium-induced neurotoxicity. Neurotoxicology 2010, 31, 391–398. [Google Scholar] [CrossRef]
- Xia, Q.; Feng, X.; Huang, H.; Du, L.; Yang, X.; Wang, K. Gadolinium-induced oxidative stress triggers endoplasmic reticulum stress in rat cortical neurons. J. Neurochem. 2011, 117, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Erdoğan, M.A.; Apaydin, M.; Armagan, G.; Taskiran, D. Evaluation of toxicity of gadolinium-based contrast agents on neuronal cells. Acta Radiol. 2021, 62, 206–214. [Google Scholar] [CrossRef]
- Wermuth, P.J.; Jimenez, S.A. Induction of a type I interferon signature in normal human monocytes by gadolinium-based contrast agents: Comparison of linear and macrocyclic agents. Clin. Exp. Immunol. 2014, 175, 113–125. [Google Scholar] [CrossRef]
- Schmidt-Lauber, C.; Bossaller, L.; Abujudeh, H.H.; Vladimer, G.I.; Christ, A.; Fitzgerald, K.A.; Latz, E.; Gravallese, E.M.; Marshak-Rothstein, A.; Kay, J. Gadolinium-based compounds induce NLRP3-dependent IL-1β production and peritoneal inflammation. Ann. Rheum. Dis. 2015, 74, 2062–2069. [Google Scholar] [CrossRef] [Green Version]
- Weng, T.-I.; Chen, H.J.; Lu, C.-W.; Ho, Y.-C.; Wu, J.-L.; Liu, S.-H.; Hsiao, J.-K. Exposure of macrophages to low-dose gadolinium-based contrast medium: Impact on oxidative stress and cytokines production. Contrast Media Mol. Imaging 2018, 2018, 3535769. [Google Scholar] [CrossRef] [PubMed]
- Bossù, P.; Cutuli, D.; Palladino, I.; Caporali, P.; Angelucci, F.; Laricchiuta, D.; Gelfo, F.; De Bartolo, P.; Caltagirone, C.; Petrosini, L. A single intraperitoneal injection of endotoxin in rats induces long-lasting modifications in behavior and brain protein levels of TNF-α and IL-18. J. Neuroinflammation 2012, 9, 101. [Google Scholar] [CrossRef] [PubMed]
- Muller, E.I.; Souza, J.P.; Muller, C.C.; Muller, A.L.H.; Mello, P.A.; Bizzi, C.A. Microwave-assisted wet digestion with H2O2 at high temperature and pressure using single reaction chamber for elemental determination in milk powder by ICP-OES and ICP-MS. Talanta 2016, 156–157, 232–238. [Google Scholar] [CrossRef]
- Deacon, R.M.; Rawlins, J.N.P. T-maze alternation in the rodent. Nat. Protoc. 2006, 1, 7. [Google Scholar] [CrossRef]
- Aggleton, J.P.; Hunt, P.; Rawlins, J. The effects of hippocampal lesions upon spatial and non-spatial tests of working memory. Behav. Brain Res. 1986, 19, 133–146. [Google Scholar] [CrossRef]
- Varatharaj, A.; Carare, R.O.; Weller, R.O.; Gawne-Cain, M.; Galea, I. Gadolinium enhancement of cranial nerves: Implications for interstitial fluid drainage from brainstem into cranial nerves in humans. Proc. Natl. Acad. Sci. USA 2021, 118, e2106331118. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Hesse, B.; Roman, M.; Stier, D.; Castillo-Michel, H.; Cotte, M.; Suuronen, J.-P.; Lagrange, A.; Radbruch, H.; Paul, F. Increased retention of gadolinium in the inflamed brain after repeated administration of gadopentetate dimeglumine: A proof-of-concept study in mice combining ICP-MS and micro–and Nano–SR-XRF. Investig. Radiol. 2019, 54, 617–626. [Google Scholar] [CrossRef] [PubMed]
- Anderhalten, L.; Silva, R.V.; Morr, A.; Wang, S.; Smorodchenko, A.; Saatz, J.; Traub, H.; Mueller, S.; Boehm-Sturm, P.; Rodriguez-Sillke, Y. Different impact of gadopentetate and gadobutrol on inflammation-promoted retention and toxicity of gadolinium within the mouse brain. Investig. Radiol. 2022, 57, 677–688. [Google Scholar] [CrossRef]
- Fu, H.Q.; Yang, T.; Xiao, W.; Fan, L.; Wu, Y.; Terrando, N.; Wang, T.L. Prolonged neuroinflammation after lipopolysaccharide exposure in aged rats. PLoS ONE 2014, 9, e106331. [Google Scholar] [CrossRef] [Green Version]
- Qin, L.; Wu, X.; Block, M.L.; Liu, Y.; Breese, G.R.; Hong, J.S.; Knapp, D.J.; Crews, F.T. Systemic LPS causes chronic neuroinflammation and progressive neurodegeneration. Glia 2007, 55, 453–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manouchehrian, O.; Ramos, M.; Bachiller, S.; Lundgaard, I.; Deierborg, T. Acute systemic LPS-exposure impairs perivascular CSF distribution in mice. J. Neuroinflammation 2021, 18, 34. [Google Scholar] [CrossRef] [PubMed]
- Galea, I. The blood–brain barrier in systemic infection and inflammation. Cell. Mol. Immunol. 2021, 18, 2489–2501. [Google Scholar] [CrossRef] [PubMed]
- Montague-Cardoso, K.; Malcangio, M. Changes in blood–spinal cord barrier permeability and neuroimmune interactions in the underlying mechanisms of chronic pain. Pain Rep. 2021, 6, e879. [Google Scholar] [CrossRef]
- Lauer, M.; Lauer, A.; You, S.J.; Kluge, S.; Hattingen, E.; Harter, P.N.; Senft, C.; Wagner, M.; Voss, M. Neurotoxicity of subarachnoid Gd-based contrast agent accumulation: A potential complication of intraoperative MRI? Neurosurg. Focus 2021, 50, E12. [Google Scholar] [CrossRef]
- Banks, W.A.; Gray, A.M.; Erickson, M.A.; Salameh, T.S.; Damodarasamy, M.; Sheibani, N.; Meabon, J.S.; Wing, E.E.; Morofuji, Y.; Cook, D.G. Lipopolysaccharide-induced blood-brain barrier disruption: Roles of cyclooxygenase, oxidative stress, neuroinflammation, and elements of the neurovascular unit. J. Neuroinflammation 2015, 12, 223. [Google Scholar] [CrossRef] [Green Version]
- Bassi, G.S.; Kanashiro, A.; Santin, F.M.; de Souza, G.E.; Nobre, M.J.; Coimbra, N.C. Lipopolysaccharide-induced sickness behaviour evaluated in different models of anxiety and innate fear in rats. Basic Clin. Pharmacol. Toxicol. 2012, 110, 359–369. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jammoul, M.; Abou-Kheir, W.; Lawand, N. How Safe Is Gadobutrol? Examining the Effect of Gadolinium Deposition on the Nervous System. Radiation 2023, 3, 75-86. https://doi.org/10.3390/radiation3020007
Jammoul M, Abou-Kheir W, Lawand N. How Safe Is Gadobutrol? Examining the Effect of Gadolinium Deposition on the Nervous System. Radiation. 2023; 3(2):75-86. https://doi.org/10.3390/radiation3020007
Chicago/Turabian StyleJammoul, Maya, Wassim Abou-Kheir, and Nada Lawand. 2023. "How Safe Is Gadobutrol? Examining the Effect of Gadolinium Deposition on the Nervous System" Radiation 3, no. 2: 75-86. https://doi.org/10.3390/radiation3020007