
Quantifying Acute Behavioral Reactions of Bali Mynas (Leucopsar rothschildi) to Environmental and Progressively Challenging Enrichment

Abstract
:
1. Introduction
2. Materials and Methods
2.1. Study Animals and Habitats
2.2. Enrichment and Filming
2.3. Behaviors
2.4. Statistical Analysis
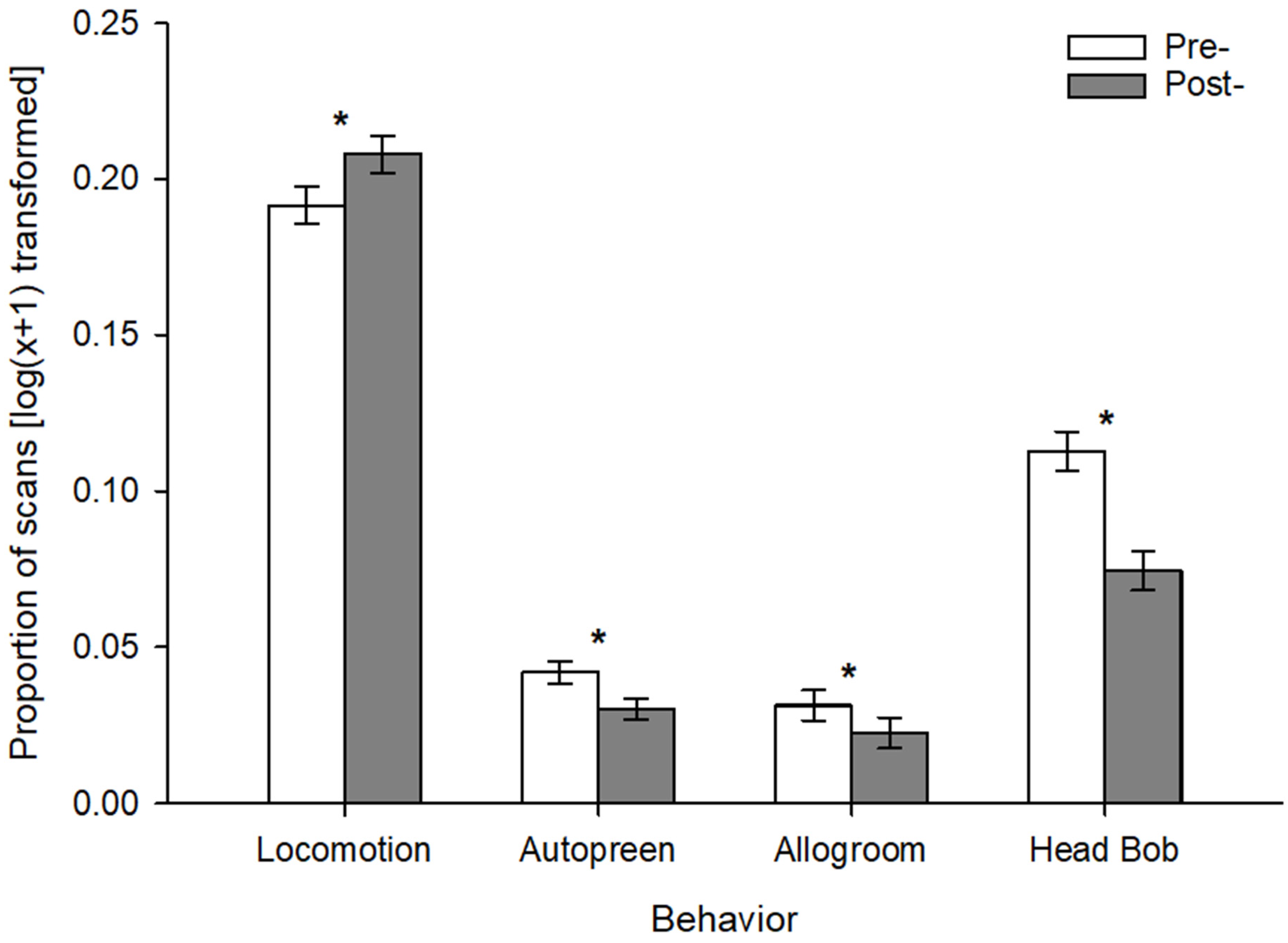
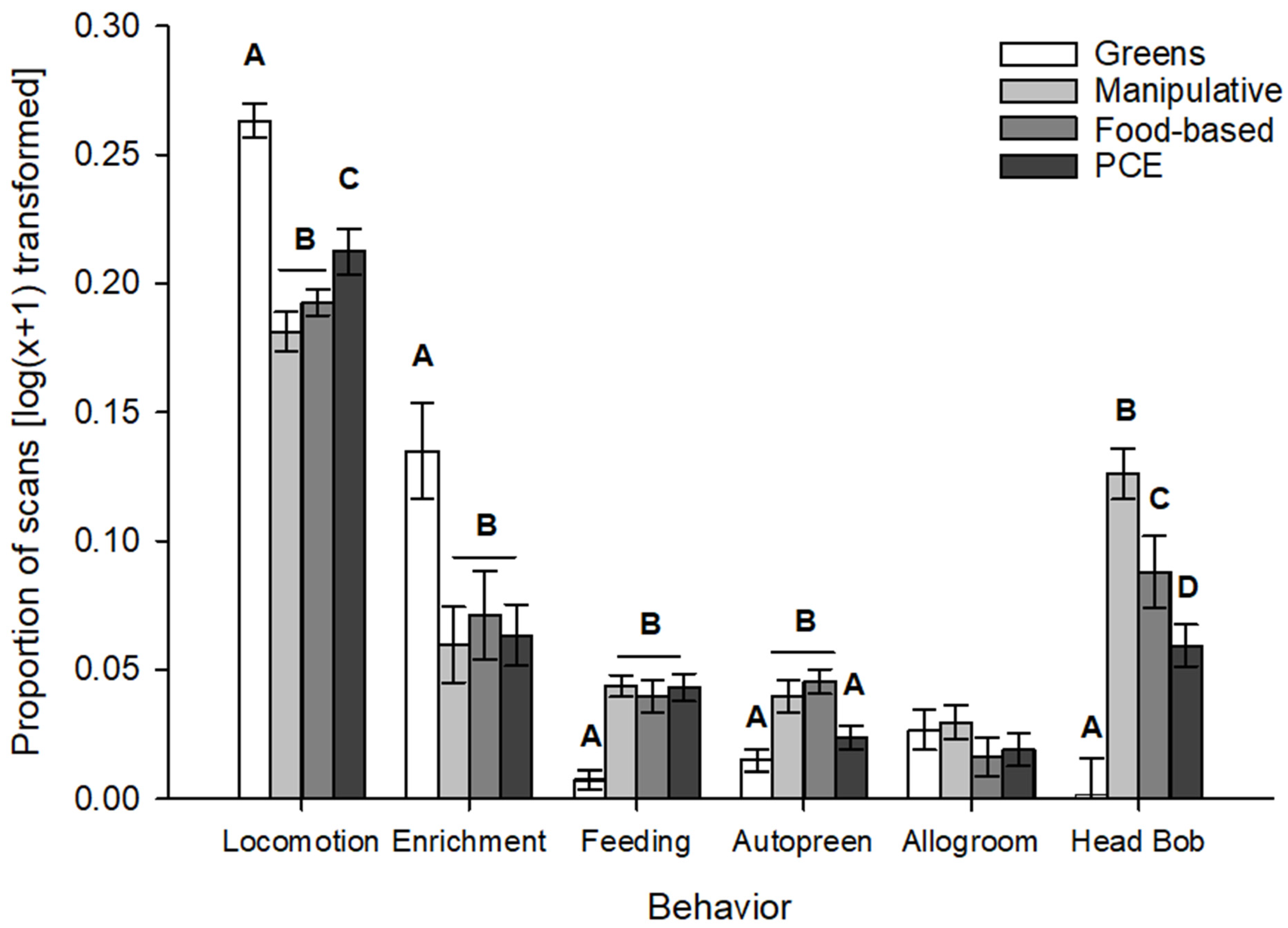
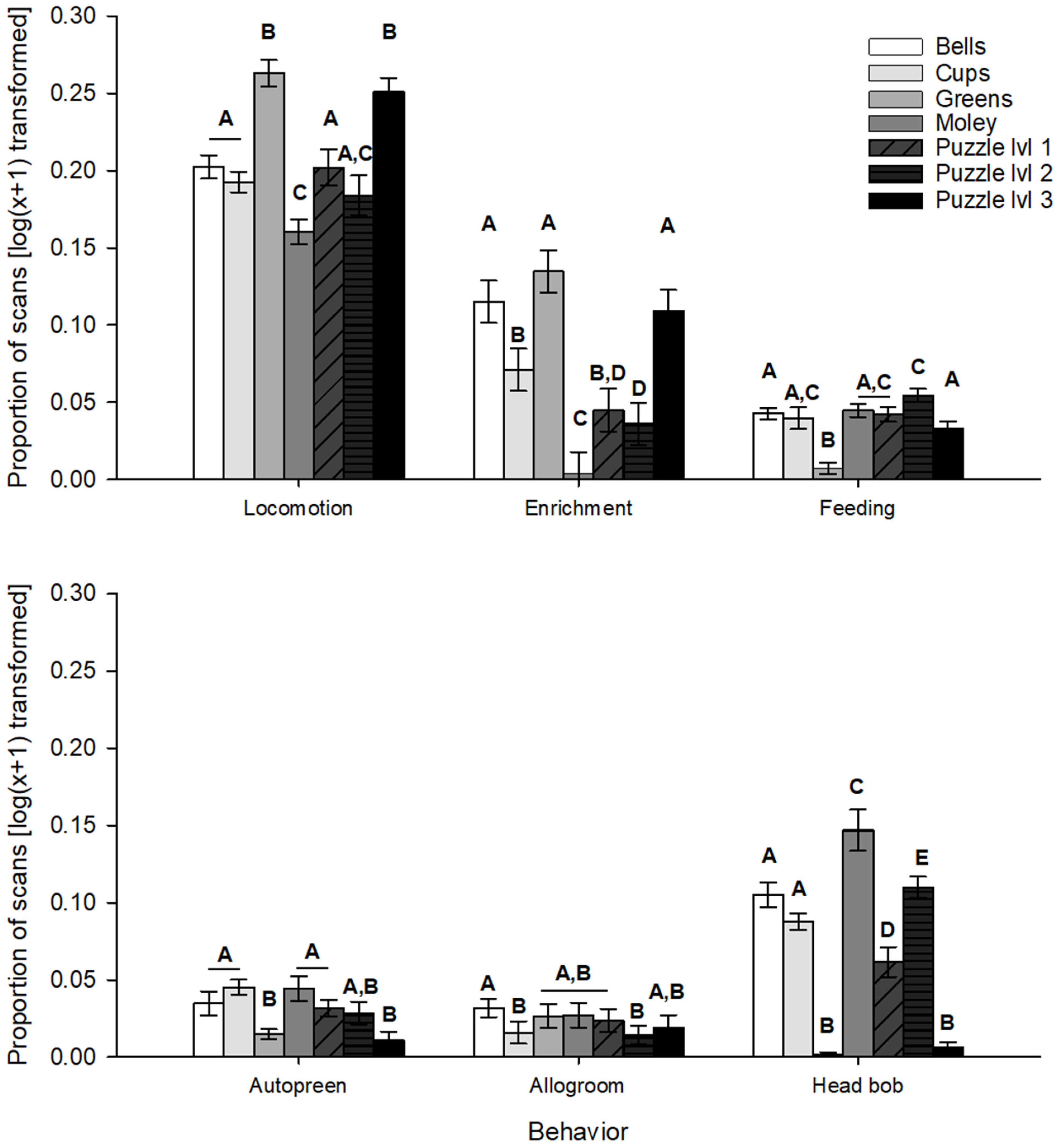
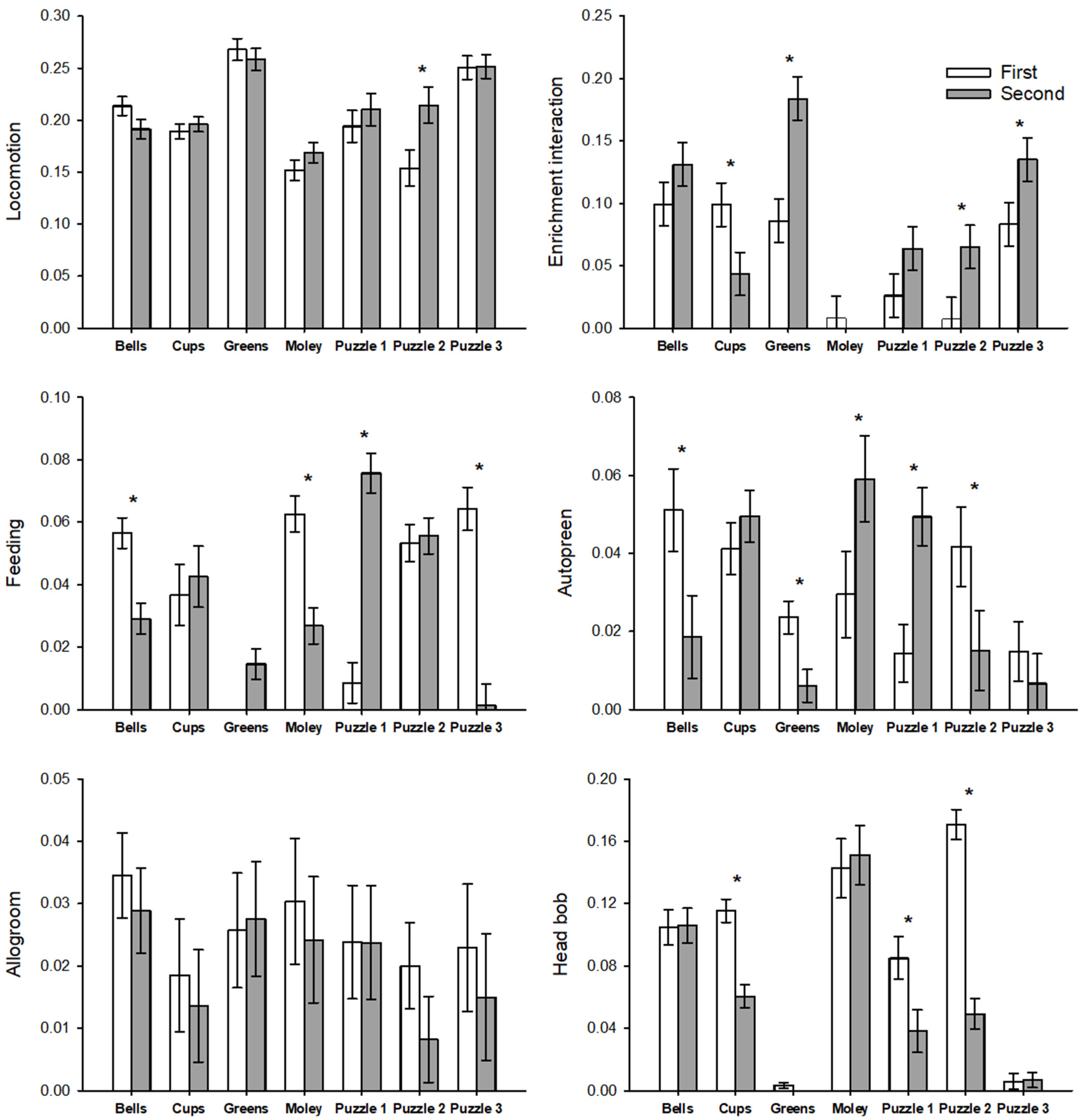
3. Results
4. Discussion
5. Conclusions
- ⮚
- Bali mynas had an acute reaction to the addition of environmental enrichment to their habitats, characterized by interactions with the enrichment, as well as significant differences in locomotion, and a reduction in autopreening, allogrooming, and head bobbing. Partner plucking, however, was unaffected by enrichment.
- ⮚
- Different items elicited stronger or weaker responses, with the offerings of naturalistic greens having reactions that were the most different from the other enrichment types. Greens strongly stimulated locomotion and interaction with the enrichment, reminiscent of natural foliage gleaning behavior.
- ⮚
- Engagement with progressively challenging enrichment increased with each successive presentation of a given difficulty level and increased significantly between the first and third difficulty level, whereas interaction decreased or saw no change during the second presentation of familiar manipulative and food-based enrichment items (e.g., cups, balls, “holey moley”).
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Farina, A.; Belgrano, A. The eco-field hypothesis: Toward a cognitive landscape. Landsc. Ecol. 2006, 21, 5–17. [Google Scholar] [CrossRef]
- Špinka, M.; Wemelsfelder, F. Environmental challenge and animal agency. In Animal Welfare, 3rd ed.; Appleby, M.C., Olsson, I.A.S., Galindo, F., Eds.; CAB International: Wallingford, UK, 2011; pp. 39–55. [Google Scholar] [CrossRef] [Green Version]
- Langbein, J.; Siebert, K.; Nürnberg, G. On the use of an automated learning device by group-housed dwarf goats: Do goats seek cognitive challenges? Appl. Anim. Behav. Sci. 2009, 120, 150–158. [Google Scholar] [CrossRef]
- McGowan, R.T.S.; Robbins, C.T.; Alldredge, J.R.; Newberry, R.C. Contrafreeloading in grizzly bears: Implications for captive foraging enrichment. Zoo Biol. 2009, 29, 484–502. [Google Scholar] [CrossRef] [PubMed]
- Sasson-Yenor, J.; Powell, D.M. Assessment of contrafreeloading preferences in giraffe (Giraffa Camelopardalis). Zoo Biol. 2019, 38, 414–423. [Google Scholar] [CrossRef] [PubMed]
- Mason, G.; Clubb, R.; Latham, N.; Vickery, S. Why and how should we use environmental enrichment to tackle stereotypic behaviour? Appl. Anim. Behav. Sci. 2007, 102, 163–188. [Google Scholar] [CrossRef] [Green Version]
- Meehan, C.L.; Garner, J.P.; Mench, J.A. Environmental enrichment and development of cage stereotypy in orange-winged Amazon parrots (Amazona amazonica). Dev. Psychobiol. 2004, 44, 209–218. [Google Scholar] [CrossRef]
- Clegg, I.L.K.; Domingues, M.; Ström, E.; Berggren, L. Cognitive foraging enrichment (but not non-cognitive enrichment) improved several longer-term welfare indicators in bottlenose dolphins. Animals 2023, 13, 238. [Google Scholar] [CrossRef]
- Gronqvist, G.; Kingston-Jones, M.; May, A.; Lehmann, J. The effects of three types of environmental enrichment on the behaviour of captive Javan gibbons (Hylobates moloch). Appl. Anim. Behav. Sci. 2013, 147, 214–223. [Google Scholar] [CrossRef]
- Hamilton, J.; Fuller, G.; Allard, S. Evaluation of the impact of behavioral opportunities on four zoo-housed aardvarks (Orycteropus afer). Animals 2020, 10, 1433. [Google Scholar] [CrossRef]
- Shepherdson, D.J. Environmental enrichment: Past, present and future. Int. Zoo Yearb. 2003, 38, 118–124. [Google Scholar] [CrossRef]
- Swaisgood, R.R.; Shepherdson, D.J. Scientific approaches to enrichment and stereotypies in zoo animals: What’s been done and where should we go next? Zoo Biol. 2005, 24, 499–518. [Google Scholar] [CrossRef]
- Douglas, C.; Bateson, M.; Walsh, C.; Bédué, A.; Edwards, S.A. Environmental enrichment induces optimistic cognitive biases in pigs. Appl. Anim. Behav. Sci. 2012, 139, 65–73. [Google Scholar] [CrossRef]
- Rodríguez-López, R. Environmental enrichment for parrot species: Are we squawking up the wrong tree? Appl. Anim. Behav. Sci. 2016, 180, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Association of Zoos and Aquariums Accreditation Standards & Related Policies 2023. Available online: https://assets.speakcdn.com/assets/2332/aza-accreditation-standards.pdf (accessed on 13 January 2023).
- Grimberg-Henrici, C.G.E.; Vermaak, P.; Bolhuis, J.E.; Nordquist, R.E.; van der Staay, F.J. Effects of environmental enrichment on cognitive performance of pigs in a spatial holeboard discrimination task. Anim. Cogn. 2016, 19, 271–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newberry, R.C. Environmental enrichment: Increasing the biological relevance of captive environments. Appl. Anim. Behav. Sci. 1995, 44, 229–243. [Google Scholar] [CrossRef]
- Hoy, J.M.; Murray, P.J.; Tribe, A. Thirty years later: Enrichment practices for captive mammals. Zoo Biol. 2009, 29, 303–316. [Google Scholar] [CrossRef] [PubMed]
- Burn, C.C. Bestial boredom: A biological perspective on animal boredom and suggestions for its scientific investigation. Anim. Behav. 2017, 130, 141–151. [Google Scholar] [CrossRef] [Green Version]
- Clark, F.E.; Smith, L.J. Effect of a cognitive challenge device containing food and non-food rewards on chimpanzee well-being: Chimpanzee cognitive challenge. Am. J. Primatol. 2013, 75, 807–816. [Google Scholar] [CrossRef]
- Meagher, R.K.; Mason, G.J. Environmental enrichment reduces signs of boredom in caged mink. PLoS ONE 2012, 7, e49180. [Google Scholar] [CrossRef] [Green Version]
- Greenberg, R.; Mettke-hofmann, C. Ecological aspects of neophobia and neophilia in Birds. In Current Ornithology; vol Nolan, V., Thompson, C.F., Eds.; Springer: Boston, MA, USA, 2001; pp. 119–178. [Google Scholar] [CrossRef]
- Meehan, C.L.; Mench, J.A. The challenge of challenge: Can problem solving opportunities enhance animal welfare? Appl. Anim. Behav. Sci. 2007, 102, 246–261. [Google Scholar] [CrossRef]
- Oesterwind, S.; Nürnberg, G.; Puppe, B.; Langbein, J. Impact of structural and cognitive enrichment on the learning performance, behavior and physiology of dwarf goats (Capra aegagrus hircus). Appl. Anim. Behav. Sci. 2016, 177, 34–41. [Google Scholar] [CrossRef]
- Keen, H.A.; Nelson, O.L.; Robbins, C.T.; Evans, M.; Shepherdson, D.J.; Newberry, R.C. Validation of a novel cognitive bias task based on difference in quantity of reinforcement for assessing environmental enrichment. Anim. Cogn. 2014, 17, 529–541. [Google Scholar] [CrossRef] [PubMed]
- Meade, T.M.; Hutchinson, E.; Krall, C.; Watson, J. Use of an aquarium as a novel enrichment item for singly housed rhesus macaques (Macaca mulatta). J. Am. Assoc. Lab. Anim. Sci. 2014, 53, 472–477. [Google Scholar] [PubMed]
- Tarou, L.R.; Bashaw, M.J. Maximizing the effectiveness of environmental enrichment: Suggestions from the experimental analysis of behavior. Appl. Anim. Behav. Sci. 2007, 102, 189–204. [Google Scholar] [CrossRef]
- Podturkin, A.A. In search of the optimal enrichment program for zoo-housed animals. Zoo Biol. 2021, 40, 527–540. [Google Scholar] [CrossRef] [PubMed]
- Tully, E.; Felsher, C. A progressively challenging enrichment project for a sloth bear (Melursus ursinus) at the Saint Louis Zoo. In Proceedings of the 40th National Conference of the American Association of Zoo Keepers, Inc., Greensboro, NC, USA, 24 September 2013. [Google Scholar]
- Hoppe, P.; Baskir, E. Swamp monkeys solve kerplunk: A progressively challenging enrichment experiment. Anim. Keep. Forum 2016, 43, 312–316. [Google Scholar]
- Boinski, S.; Swing, S.P.; Gross, T.S.; Davis, J.K. Environmental enrichment of brown capuchins (Cebus apella): Behavioral and plasma and fecal cortisol measures of effectiveness. Am. J. Primatol. 1999, 48, 49–68. [Google Scholar] [CrossRef]
- Caselli, M.; Messeri, P.; Dessì-Fulgheri, F.; Bandoli, F. Enriching zoo-housed ring-tailed lemurs (Lemur catta): Assessing the influence of three types of environmental enrichment on behavior. Animals 2022, 12, 2836. [Google Scholar] [CrossRef]
- Laméris, D.W.; Verspeek, J.; Depoortere, A.; Plessers, L.; Salas, M. Effects of enclosure and environmental enrichment on the behaviour of ring-tailed lemurs (Lemur catta). J. Zool. Bot. Gard. 2021, 2, 164–173. [Google Scholar] [CrossRef]
- Renner, M.J.; Lussier, J.P. Environmental enrichment for the captive spectacled bear (Tremarctos ornatus). Pharmacol. Biochem. Behav. 2002, 73, 279–283. [Google Scholar] [CrossRef]
- Rooney, M.B.; Sleeman, J. Effects of selected behavioral enrichment devices on behavior of western lowland gorillas (Gorilla gorilla gorilla). J. Appl. Anim. Welf. Sci. 1998, 1, 339–351. [Google Scholar] [CrossRef] [PubMed]
- Soriano, A.I.; Ensenyat, C.; Serrat, S.; Maté, C. Introducing a semi-naturalistic exhibit as structural enrichment for two brown bears (Ursus arctos). Does this ensure their captive well-being? J. Appl. Anim. Welf. Sci. 2006, 9, 299–314. [Google Scholar] [CrossRef] [PubMed]
- Szokalski, M.S.; Litchfield, C.A.; Foster, W.K. Enrichment for captive tigers (Panthera tigris): Current knowledge and future directions. Appl. Anim. Behav. Sci. 2012, 139, 1–9. [Google Scholar] [CrossRef]
- Rose, P.E.; Brereton, J.E.; Rowden, L.J.; de Figueiredo, R.L.; Riley, L.M. What’s new from the zoo? An analysis of ten years of zoo-themed research output. Palgrave Commun. 2019, 5, 128. [Google Scholar] [CrossRef] [Green Version]
- Azevedo, C.S.; Caldeira, J.R.; Faggioli, Â.B.; Cipreste, C.F. Effects of different environmental enrichment items on the behavior of the endangered Lear’s Macaw (Anodorhynchus leari, Psittacidae) at Belo Horizonte Zoo, Brazil. Rev. Bras. Ornitol. 2016, 24, 204–210. [Google Scholar] [CrossRef]
- Brereton, J.E.; Myhill, M.N.G.; Shora, J.A. Investigating the effect of enrichment on the behavior of zoo-housed southern ground hornbills. J. Zool. Bot. Gard. 2021, 2, 600–609. [Google Scholar] [CrossRef]
- Coulton, L.E.; Waran, N.K.; Young, R.J. Effects of foraging enrichment on the behavior of parrots. Anim. Welf. 1997, 6, 357–363. [Google Scholar] [CrossRef]
- Fangmeier, M.L.; Burns, A.L.; Melfi, V.A.; Meade, J. Foraging enrichment alleviates oral repetitive behaviors in captive red-tailed black cockatoos (Calyptorhynchus banksii). Zoo Biol. 2020, 39, 3–12. [Google Scholar] [CrossRef]
- Field, D.A.; Thomas, R. Environmental enrichment for psittacines at Edinburgh Zoo. Int. Zoo Yearb. 2000, 37, 232–237. [Google Scholar] [CrossRef]
- Jepson, P.R. Saving a species threatened by trade: A network study of Bali starling Leucopsar rothschildi conservation. Oryx 2016, 50, 480–488. [Google Scholar] [CrossRef] [Green Version]
- Zuccon, D.; Pasquet, E.; Ericson, P.G.P. Phylogenetic relationships among Palearctic-Oriental starlings and mynas (genera Sturnus and Acridotheres: Sturnidae). Zool. Scr. 2008, 37, 469–481. [Google Scholar] [CrossRef]
- Higgins, P.J.; Peter, J.M.; Cowling, S.J. (Eds.) Handbook of Australian, New Zealand & Antarctic Birds; Volume Oxford University Press: Melbourne, Australia, 2006. [Google Scholar]
- Nikolov, S.; Kanakov, D. Influencing factors leading to damaging behavior—Feather pecking and cannibalism in game birds. Trakia J. Sci. 2020, 18, 377–387. [Google Scholar] [CrossRef]
- Kinkaid, H.Y.M.; Mills, D.S.; Nichols, S.G.; Meagher, R.K.; Mason, G.J. Feather-damaging behaviour in companion parrots: An initial analysis of potential demographic risk factors. Avian Biol. Res. 2013, 6, 289–296. [Google Scholar] [CrossRef]
- van Zeeland, Y.R.A.; Spruit, B.M.; Rodenburg, T.B.; Riedstra, B.; van Hierden, Y.M.; Buitenhuis, B.; Korte, S.M.; Lumeij, J.T. Feather damaging behaviour in parrots: A review with consideration of comparative aspects. Appl. Anim. Behav. Sci. 2009, 121, 75–95. [Google Scholar] [CrossRef]
- Costa, P.; Macchi, E.; Valle, E.; De Marco, M.; Nucera, D.M.; Gasco, L.; Schiavone, A. An association between feather damaging behavior and corticosterone metabolite excretion in captive African grey parrots (Psittacus erithacus). PeerJ 2016, 4, e2462. [Google Scholar] [CrossRef] [Green Version]
- Heinsius, J.; van Staaveren, N.; Kwon, I.Y.; Li, A.; Kjaer, J.B.; Harlander-Matauschek, A. Chickens selected for feather pecking can inhibit prepotent motor responses in a Go/No-Go task. Sci. Rep. 2020, 10, 6485. [Google Scholar] [CrossRef] [Green Version]
- Mens, A.J.W.; van Krimpen, M.M.; Kwakkel, R.P. Nutritional approaches to reduce or prevent feather pecking in laying hens: Any potential to intervene during rearing? World’s Poult. Sci. J. 2020, 76, 591–610. [Google Scholar] [CrossRef]
- Costa, P.; Macchi, E.; Tomassone, L.; Ricceri, F.; Bollo, E.; Scaglione, F.E.; Tarantola, M.; De Marco, M.; Prola, L.; Bergero, D.; et al. Feather picking in pet parrots: Sensitive species, risk factor and ethological evidence. Ital. J. Anim. Sci. 2016, 15, 473–480. [Google Scholar] [CrossRef] [Green Version]
- Lumeij, J.T.; Hommers, C.J. Foraging ‘enrichment’ as treatment for pterotillomania. Appl. Anim. Behav. Sci. 2008, 111, 85–94. [Google Scholar] [CrossRef]
- Van Staaveren, N.; Ellis, J.; Baes, C.F.; Harlander-Matauschek, A. A meta analysis on the effect of environmental enrichment on feather pecking and feather damage in laying hens. Poult. Sci. 2021, 100, 397–411. [Google Scholar] [CrossRef]
- Martin, P.; Bateson, P. Measuring Behaviour: An Introductory Guide, 3rd ed.; Cambridge University Press: Cambridge, UK, 2007. [Google Scholar] [CrossRef]
- Schielzeth, H.; Dingemanse, N.J.; Nakagawa, S.; Westneat, D.F.; Allegue, H.; Teplitsky, C.; Réale, D.; Dochtermann, N.A.; Garamszegi, L.Z.; Araya-Ajoy, Y.G. Robustness of linear mixed-effects models to violations of distributional assumptions. Methods Ecol. Evol. 2020, 11, 1141–1152. [Google Scholar] [CrossRef]
- Arnau, J.; Bendayan, R.; Blanca, M.J.; Bono, R. The effect of skewness and kurtosis on the robustness of linear mixed models. Behav. Res. 2013, 45, 873–879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rudkin, C. Feather pecking and foraging uncorrelated—The redirection hypothesis revisited. Br. Poult. Sci. 2022, 63, 265–273. [Google Scholar] [CrossRef]
- Shi, H.; Li, B.; Tong, Q.; Zheng, W.; Zeng, D.; Feng, G. Effects of LED Light Color and Intensity on Feather Pecking and Fear Responses of Layer Breeders in Natural Mating Colony Cages. Animals 2019, 9, 814. [Google Scholar] [CrossRef] [Green Version]
- Schwarzer, A.; Plattner, C.; Bergmann, S.; Rauch, E.; Erhard, M.; Reese, S.; Louton, H. Feather pecking in non-beak-trimmed and beak-trimmed laying hens on commercial farms with aviaries. Animals 2021, 11, 3085. [Google Scholar] [CrossRef] [PubMed]
- Lima, M.F.F.; de Azevedo, C.S.; Young, R.J.; Viau, P. Impacts of food-based enrichment on behaviour and physiology of male greater rheas (Rhea Americana, Rheidae, Aves). Papéis Avulsos Zool. 2019, 59, e20195911. [Google Scholar] [CrossRef]
- de Almeida, A.C.; Palme, R.; Moreira, N. How environmental enrichment affects behavioral and glucocorticoid responses in captive blue-and-yellow macaws (Ara ararauna). Appl. Anim. Behav. Sci. 2018, 201, 125–135. [Google Scholar] [CrossRef]
- Miglioli, A.; da Silva Vasconcellos, A. Can behavioural management improve behaviour and reproduction in captive blue-and-yellow macaws (Ara ararauna)? Appl. Anim. Behav. Sci. 2021, 241, 105386. [Google Scholar] [CrossRef]
- Congdon, S. Starlings and mynas. AFA Watchb. 1999, 26, 17–19. [Google Scholar]
- Hernowo, J.B.; Haquesta, S. Evaluation on Bali Mynah (Leucopsar rothschildi Stresemann, 1912) population, result of release process 1998–2015 in Bali Barat National Park, Indonesia. Biodiversitas 2021, 22, 2699–2710. [Google Scholar] [CrossRef]
- Miller, R.; Lambert, M.L.; Frohnwieser, A.; Brecht, K.F.; Bugnyar, T.; Crampton, I.; Garcia-Pelegrin, E.; Gould, K.; Greggor, A.L.; Izawa, E.; et al. Socio-ecological correlates of neophobia in corvids. Curr. Biol. 2022, 32, 74–85. [Google Scholar] [CrossRef] [PubMed]
- Miller, R.; Garcia-Pelegrin, E.; Danby, E. Neophobia and innovation in critically endangered Bali myna, Leucopsar rothschildi. R. Soc. Open Sci. 2022, 9, 211781. [Google Scholar] [CrossRef] [PubMed]
- Ghavamian, Y.; Minier, D.E.; Jaffe, K.E. Effects of complex feeding enrichment on the behavior of captive Malayan sun bears (Helarctos malayanus). J. Appl. Anim. Welf. Sci. 2022, 24, 1–15. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category Name | Description | Examples |
|---|---|---|
| Manipulative | Physical objects with no food reward | hanging bells; “holey moley” hollow rubber dog toy sphere with hexagonal cut-outs filled with hay |
| Greens | Between 2 and 4 pieces of whole romaine lettuce leaf, skewered or staked onto branches | |
| Food-based | Physical objects filled with non-greens food, regardless of how offered | food in hanging cups |
| Progressively challenging enrichment (PCE) | Modified suet feeder (see below) |
| PCE Difficulty Level | Description |
|---|---|
| 1 | Bamboo tubes measure 2.54 +/− 0.32 cm above feeder mesh |
| 2 | Bamboo tubes measure 5.08 +/− 0.32 cm above feeder mesh |
| 3 | Bamboo tubes measure 5.08 +/− 0.32 cm above feeder mesh and are filled with wood shaving substrate |
| Item | Category | Dates Offered and Analyzed |
|---|---|---|
| bells | manipulative | 12 and 20 May 2019 |
| diet served in hanging cup clusters | food-based | 25 May and 8 June 2019 |
| greens | greens | 28 May and 18 June 2019 |
| “holey moley” dog toy filled with plastic toy (i.e., rubber duck) or hay | manipulative | 21 and 23 May 2019 |
| Puzzle, level 1 | PCE | 24 and 30 May 2019 |
| Puzzle, level 2 | PCE | 7 and 10 June 2019 |
| Puzzle, level 3 | PCE | 15 and 19 June 2019 |
| Behavior | Sampling Method | Description |
|---|---|---|
| Locomotion | 1-0 | Any movement, flying, walking, or hopping, at any speed that displaces distance |
| Feeding | 1-0 | Subject is actively removing food from a non-enrichment source, e.g., food dish |
| Enrichment interaction | 1-0 | Subject is within one wing length of an enrichment device or object and either directly touching or directly staring with head oriented at the enrichment item |
| Autopreen | 1-0 | Subject grooms themself, using beak to clean and smooth feathers |
| Allogroom | 1-0 | Subject makes continued beak contact with conspecific’s feathers, body, head, etc. Based on duration of contact and whether withdrawal of initiator’s head is forceful, Pluck (see below) may occur during this behavior but is not considered an inherent part of it |
| Head bob | 1-0 | With head feathers raised, bird vigorously and repeatedly tosses its head up and down, with or without open mouth and vocalization |
| Out of sight | 1-0 | This behavior is only scored if a subject is absent from camera for an entire interval |
| Pluck | continuous | Brief beak contact with conspecific’s neck/throat area that ends with forceful jerk of initiator’s beak away from recipient. Score a pluck event each time contact is forcefully broken with beak withdrawing in closed position |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baskir, E.; Parsons, A.; Elden, M.; Powell, D.M. Quantifying Acute Behavioral Reactions of Bali Mynas (Leucopsar rothschildi) to Environmental and Progressively Challenging Enrichment. J. Zool. Bot. Gard. 2023, 4, 176-190. https://doi.org/10.3390/jzbg4010017
Baskir E, Parsons A, Elden M, Powell DM. Quantifying Acute Behavioral Reactions of Bali Mynas (Leucopsar rothschildi) to Environmental and Progressively Challenging Enrichment. Journal of Zoological and Botanical Gardens. 2023; 4(1):176-190. https://doi.org/10.3390/jzbg4010017
Chicago/Turabian StyleBaskir, Eli, Alayna Parsons, Marija Elden, and David M. Powell. 2023. "Quantifying Acute Behavioral Reactions of Bali Mynas (Leucopsar rothschildi) to Environmental and Progressively Challenging Enrichment" Journal of Zoological and Botanical Gardens 4, no. 1: 176-190. https://doi.org/10.3390/jzbg4010017