1. Introduction
Amphibian populations have been declining at accelerated rates in the past several decades due in part to habitat destruction, diseases, and invasive species [
1,
2]. With this decline, it is imperative to maintain the health of amphibians in human care and facilitate breeding programs with free-ranging reintroduction goals for critically endangered species. Nutrition is an important factor that promotes the health and reproduction of amphibians [
3].
Vitamin A is an important antioxidant in amphibian diets and is essential for vision, immune system function, growth, and reproduction [
4,
5]. Many amphibian species are thought to be dependent on the carotenoids in the diet for vitamin A [
6] since they are dietary carnivores and likely cannot synthesize either vitamin A or preformed vitamin A carotenoids, such as retinol [
5,
7]. However, there are questions regarding the appropriate amount and dietary form, especially since reports of vitamin A deficiencies have been documented even in supplemented animals [
5]. Amphibian vitamin A deficiency concerns include short tongue syndrome, periocular and conjunctival swelling, growth deficiency, skin ulcers, gastric bloat, septicemia, and death [
8]. Vitamin A biochemistry in amphibian species is poorly understood and establishing model species for vitamin A dietary recommendations and establishing reference ranges is needed [
5]. However, with nearly 8000 amphibian species existing [
9] worldwide, there is likely considerable variation among species making this vitamin A dietary requirement concept quite complex. While there is potential importance in supplementing vitamin A to prevent deficiencies, there also is the concern of over-supplementing and causing toxicity due to the lack of an established reference range of normal concentration [
5].
Vitamin E is an antioxidant that helps decrease susceptibility to disease and has anti-inflammatory properties [
10,
11]. There are a variety of diseases and symptoms that can result from vitamin E deficiency in animals including cardiomyopathy, anemia, encephalomalacia, nerve degeneration, fat necrosis, and anorexia [
4]. However, there is little information regarding the vitamin E deficiency symptoms in amphibians [
12]. The lack of information and the importance of vitamin E in reproduction warrants further investigation regarding its effects, especially in the population of amphibians in their natural habitat and human-managed populations intended for reintroduction.
Marine toads (
Rhinella marina) are an invasive species that were originally introduced in the United States in the early to mid-1900′s to try to control insects and agricultural pests, particularly in cane fields in the southeast [
13]. Wild populations that have become established have potential negative ecological and health effects, such as native species population reduction and toxicity to companion animals [
13,
14]. Marine toads are often annually euthanized or removed via government or community-sanctioned population control programs and have proven to be very useful when they were utilized for veterinary health and nutrition research projects to help progress our understanding of amphibian biology [
12,
15,
16,
17]. Although, as previously noted, there are thousands of species of amphibians within the Amphibia class, nutrition research with species within this class is sparse and therefore marina toad research could provide more precise data due to their unique diet strategies than using species within other taxonomic classes [
3].
The objective of this research study was to analyze the kinetics of vitamin A, vitamin E, and carotenoids in marine toads as they transitioned from a free-ranging diet to a diet commonly fed to amphibians under human care with and without vitamin A and E supplementation.
2. Materials and Methods
This trial was conducted under the approval of the NC State University Institutional Animal Care and Use Committee (IACUC) (IACUC #20-207), the Zoo Miami Animal Care and Use Committee (#2020-6), and the North Carolina Zoo (NC Zoo) research review board.
2.1. Animals
Sixty-six free-ranging (wild) marine toads (Rhinella marina) were collected from the grounds of Zoo Miami in Miami, FL, USA on the night of 19 August 2020, during a routine invasive species population control program event. Selected toads had a minimum weight of 35 g for consistency and to avoid cannibalism of very small individuals. The first 10 toads collected (average weight of 128.7 ± 15.31 g) were immediately anesthetized and euthanized individually by a veterinarian via overdosing in a bath of MS-222 (10 g/L, buffered with sodium bicarbonate) (Tricaine methanesulfonate, Syndel, Ferndale, Washington, DC, USA); death was verified by using a Doppler ultrasound probe. The remaining toads were individually identified with a subcutaneous passive integrated transponder (Biomark®, Boise, ID, USA).
2.2. Housing and Diet
The remaining 56 toads were examined by a Zoo Miami veterinarian and shipped overnight via International Air Transport Association (IATA) amphibian guidelines to the NC Zoo in Asheboro, NC, USA via the fastest route possible [
18]. The toads were health inspected upon arrival and housed at the Hanes Veterinary Hospital under established housing according to protocols developed for amphibians and were quarantined at the NC Zoo [
19]. All toads were individually and visually assessed for health daily. Four days after arrival, all toads were examined by the veterinary staff, weighed, and dewormed with ivermectin at 0.2 mg/kg PO [
20]. All 56 toads were blocked by weight and then randomly assigned to either Diet 1 or 2 (average weights of 95.6 g and 95.5 g, respectively) and sorted within housing tubs by Microsoft Excel’s shuffle function (Microsoft Office 2013, Redmond, WA, USA).
Housing tubs consisted of six 511 L tubs (178.8 cm × 81.3 cm × 35.6 cm). Each tub had a hiding area and a pool of reconstituted reverse osmosis (RO) water that was slightly salinated (NaCl) at 2 ppt and a dry area lined with a damp towel as substrate [
12]. Each morning, water was drained and replaced with fresh RO water, a fresh towel was placed in the dry area, and tubs were disinfected weekly with chlorhexidine gluconate 2% solution diluted 1 oz per gallon of water. Animals were kept at ambient temperature with a light cycle from 0800–1700 h to correspond with the schedule of the animal husbandry staff with no necessary seasonal supplemental heating or cooling. The temperature and humidity of the room were recorded daily to ensure appropriate conditions using an AcuRite
® (#219CA, Lake Geneva, WI, USA) indoor temperature and humidity monitor.
The toads were fed live 0.75-inch adult brown house crickets (Acheta domestica) (Catawaba Cricket Hatchery Inc. (Charlotte, NC, USA)). The crickets utilized throughout this study were shipped bi-weekly overnight to the NC Zoo with small pieces of potato in their carrier. The crickets were then gut loaded for a minimum of 72 h using Mazuri® (St. Louis, MO, USA) Cricket Diet 5M38 diet plus small amounts of sweet potato and carrot before being fed to the toads. Half of the toads were fed the control diet (Diet 1) which consisted of the gut-loaded crickets alone. The other half of the toads were fed a treatment diet, which was identical to the control diet with the exception that the crickets were additionally dusted with Repashy® Superfoods Vitamin A Plus (La Jolla, CA, USA) immediately before being provided as food to the tubs of toads (Diet 2). Toads were offered approximately 12 crickets per toad at 4 PM daily, with the uneaten crickets disposed of by 11 AM the next day, 7 days a week for the first 4 weeks. Based on the number of crickets left uneaten, feeding frequency was decreased to 5 days a week for week 5. After week 6, toads were offered only nine crickets per toad for the remainder of the study.
Seven toads from each diet treatment were euthanized at different time points (9, 15, 32, and 62 days). Each toad was anesthetized and euthanized individually by a veterinarian via overdosing in a bath of MS-222 (10 g/L, buffered with sodium bicarbonate) (Tricaine methanesulfonate) followed by pithing, with death verified via Doppler ultrasound probe.
2.3. Liver and Diet Collection and Analysis Methods
Entire livers were removed immediately after euthanasia, with a 1 g subsample taken and wrapped in aluminum foil to protect from light, and frozen in a −80 °C freezer. The subsamples were shipped to Arizona State University (Tempe, AZ, USA) using dry ice at the end of the trial. Livers were analyzed for vitamin A (retinol), vitamin E (α-tocopherol), and the carotenoids: apocarotenoid, β-carotene, β-carotene ester, β-cryptoxanthin, β-cryptoxanthin ester, lutein, lutein ester, zeaxanthin, and zeaxanthin ester using the HPLC method [
21].
Similarly, samples of the Mazuri
® cricket Diet 5M38 and Repashy
® Vitamin A Plus were analyzed for the same nutrients. Brown house crickets after consuming the 5M38 diet for 72 h with and without the Repashy
® Vitamin A Plus dusting supplement, indicative of each diet treatment, were euthanized using an overdose of Isoflurane followed by freezing at −80 °C [
22,
23] and then shipped overnight with the liver samples on dry ice for the same analyses.
2.4. Cricket Feed and Cricket Analysis Methods
Samples of the Mazuri® cricket Diet 5M38 and Repashy® Vitamin A Plus were sent to Zooquarius® Laboratory Services (Ithaca, NY, USA) for their Feed 1 profile (#190). Crickets that had been gut loaded for 72 h and indicative of each diet treatment (euthanized, stored, and shipped as discussed above) were also sent to Zooquarius® for their Insect 1 (#191) profile.
Diet samples were analyzed for moisture, dry matter (DM), crude protein (CP), nitrogen-free extract (NFE), acid detergent fiber (ADF), neutral detergent fiber (and), crude fiber (CF), water-soluble carbohydrates (WSC), starch, fat (ether extract), ash, calcium (Ca), phosphorus (P), Ca:P ratio, magnesium (Mg), potassium (K), sodium (Na), chloride (Cl), sulfur (S), iron (Fe), zinc (Zn), copper (Cu), manganese (Mn), molybdenum (Mo), and cobalt (Co).
Crickets were analyzed for: DM, CP, ADF, fat (acid hydrolysis fat, (AHF)), ash, Ca, P, Ca:P ratio, Mg, K, Na, S, Fe, Zn, Cu, Mn, and Mo. The following analytical methods were used: moisture (AOAC 934.01), DM (AOAC 950.46), CP (AOAC 992.15), ADF (AOAC 973.18), aNDF [
24] CF (AOAC 978.10), WSC [
25], starch (AACC 76-13), fat (EE (AOAC 2003.05), AHF (AOAC 954.02), ash (AOAC 942.05), and minerals analyzed by inductively coupled plasma (ICP) (AOAC 985.01) (AACC, 2000; AOAC, 2006). NFE was calculated (100- (Moisture + Ash + Protein + Fiber + Fat)).
2.5. Histology
A complete necropsy was performed, and tissue sets were collected and fixed in neutral buffered formalin (H + E stain) from wild-caught toads and those kept in managed care for 15, 32, and 62 days [
26,
27]. The majority of the liver for the toads euthanized on Day 9 was utilized in Cabot et al.’s study characterizing enzyme activities in the marine toad (
Rhinella marina), and therefore were only sent for nutrient liver analysis and not necropsy in this current study [
17]. After fixation, two sections of the liver, as well as sections of the kidney, the gastrointestinal tract, the urogenital tract, the lung, and the liver from each toad were trimmed and processed routinely for evaluation by light microscopy by a board-certified veterinary pathologist. The sex of individual toads was histologically verified and staged. Hepatic cytoplasmic vacuolation was scored following standard best practices for toxicologic pathology based on modified criteria from the Larval Amphibian Growth and Development Assay: Grade 0 = 25% of hepatocytes show vacuolation; Grade 1 = 25–50% of hepatocytes show vacuolation; Grade 2 = 25–50% of hepatocytes show vacuolation; Grade 3 = 50–75% of hepatocytes show vacuolation; and Grade 4 = 75% of hepatocytes show vacuolation [
26,
27].
2.6. Statistical Analysis
Statistical analysis was performed using Proc GLM (general linear models) procedure model statements in SAS 9.4 (Cary, NC, USA). Data were normally distributed within a day and differences were determined with p < 0.05 considered to be statistically significant and (p = 0.05 to 0.10) as significant tendencies. The least square (LS) means and standard error of the mean (SEM) were calculated. Analysis of variance (ANOVA) was used to determine if there were significant differences among the toad’s free-ranging weight and nutrient concentrations (Day 0) and all other time points (Days 9, 15, 32, and 62) on both diets (diet * day comparisons).
3. Results
All marine toads remained visually healthy throughout the duration of the research study. The area housing the study animals was not climate-controlled and indoor temperatures and humidity fluctuated daily and throughout the study period. The temperature and humidity range for the 62-day duration study was: 21–28 °C and 42–88%, respectively.
The free-ranging toads (FR) consisted of five females and five males. The 14 toads euthanized on Day 9 of managed care consisted of four females, eight males, and two that were unknown. The 14 toads euthanized on day 15 consisted of five females and nine males. The 14 toads euthanized on day 32 consisted of five females and nine males. Lastly, the 14 toads euthanized on Day 62 consisted of three females and 11 males.
Toad weight was determined upon euthanasia and did not differ by euthanasia day (
p = 0.10) or by treatment (
p = 0.60). The free-ranging toads at Day 0 weighed on average 128.7 ± 15.31 g. The 14 toads euthanized on Day 9 weighed on average 121.0 ± 10.99 g. The 14 toads on Day 15 weighed on average 140.5 ± 9.86 g. The 14 toads on Day 32 weighed on average 152.4 ± 15.27 g. Lastly, the 14 toads on Day 62 weighed an average of 174.0 ± 15.08 g. Weight by treatment group on euthanasia day is shown in
Table 1.
3.1. Nutrient Analyses of Cricket Diet, Dusting supplement, Crickets, and Crickets with Dusting Supplement
Analysis of the cricket diet and dusting supplement are shown in
Table 1. The supplement had the expected higher levels of α-carotene, α-tocopherol, β-carotene, lutein, retinol, and zeaxanthin when compared to the cricket diet. Analysis of the crickets themselves after consuming a cricket diet with and without supplement dusting for three days is shown in
Table 2. Crickets that were dusted also showed the expected higher levels of α-carotene, β-carotene, lutein, retinol, and zeaxanthin when compared to the crickets without the dusting.
3.2. Hepatic Vitamin A, Vitamin E, and Carotenoids Concentrations
All measured nutrients decreased numerically over the totality of the two-month period in human care regardless of dietary treatment with numerous significant differences among the diet by day in human management statistical analyses (
Table 3). Alpha-tocopherol, apocarotenoid, β-carotene ester, and lutein concentrations in the liver were all significantly highest at Day 0 (
p < 0.05) for both Diet 1 and Diet 2 compared to Day 62. Apocarotenoids were highest for both diets starting on Day 15.
There were several decreasing tendencies from Day 0 with liver retinol, lutein ester, and zeaxanthin when comparing concentrations among the diet treatment by day in human management. Most notably, Diet 1 and Diet 2 for retinol and lutein ester tended to decrease by Day 62, and zeaxanthin tended to decrease for both diets by Day 32. There were no other tendencies between diet treatments for any other nutrients (
Table 3). β-carotene, β-cryptoxanthin, and Zeaxanthin esters visually decreased but no statistical differences or tendencies were noted among the comparisons.
When looking at Diet 1 vs. Diet 2, differences specifically by date in human management, only β-carotene esters differed between the two on Day 9. Thus, while nutrients measured in human management had numerous differences and tendencies over time, Diets 1 and 2 remained almost completely consistent statistically.
Table 1.
Laboratory nutrient analyses (dry matter basis) of the exact batches of commercial cricket diet 1 and nutrient dusting supplement 2 that were fed to Marine toads (Rhinella marina) housed at the NC Zoo.
Table 1.
Laboratory nutrient analyses (dry matter basis) of the exact batches of commercial cricket diet 1 and nutrient dusting supplement 2 that were fed to Marine toads (Rhinella marina) housed at the NC Zoo.
| Nutrient | Unit | Cricket Diet | Supplement |
|---|
| Moisture | % | 7.7 | 2.2 |
| Dry Matter | % | 92.3 | 97.8 |
| Crude Protein | % | 20.2 | 20.5 |
| Nitrogen Free Extract | % | 39.5 | NT |
| Acid Detergent Fiber | % | 12.4 | NT |
| Neutral Detergent Fiber | % | 13.8 | NT |
| Water Soluble Carbohydrates | % | 3.9 | NT |
| Starch | % | 20.7 | NT |
| Crude Fat (ether extract) | % | 3.8 | NT |
| Crude Fiber | % | 10.4 | NT |
| Ash | % | 23.8 | NT |
| Ca | % | 8.1 | 22.7 |
| P | % | 0.59 | 0.29 |
| Ca:P Ratio | 1 to | 13.8 | 78.2 |
| Mg | % | 0.27 | 0.18 |
| K | % | 0.91 | 0.47 |
| Na | % | 0.20 | 0.36 |
| Cl | % | NT | 0.31 |
| S | % | 0.29 | 0.52 |
| Fe | ppm | 297 | 247 |
| Zn | ppm | 74 | 10 |
| Cu | ppm | 16 | NT |
| Mn | ppm | 99 | 55 |
| Mo | ppm | 1.7 | 1.7 |
| Co | ppm | NT | 0.48 |
| α-carotene | ug/g | 0.02 | 57.3 |
| α-tocopherol | ug/g | 8.1 | 30.8 |
| β-carotene | ug/g | 0.03 | 218.9 |
| β-cryptoxanthin | ug/g | ND | ND |
| Lutein | ug/g | 0.8 | 7.4 |
| Retinol | ug/g | 12.3 | 286.6 |
| Zeaxanthin | ug/g | 0.3 | 227.7 |
Table 2.
Adult brown house cricket 1 (Acheta domestica) proximate analyses (dry matter basis) after consuming cricket diet 2 for three days with and without supplement 3 dusting.
Table 2.
Adult brown house cricket 1 (Acheta domestica) proximate analyses (dry matter basis) after consuming cricket diet 2 for three days with and without supplement 3 dusting.
| Nutrient | Unit | Crickets Alone | Crickets + Supplement |
|---|
| Moisture | % | 70.5 | 69.0 |
| Dry Matter | % | 29.5 | 31.0 |
| Crude Protein | % | 67.3 | 63.3 |
| Acid Detergent Fiber | % | 35.2 | 44.2 |
| Crude Fat (Acid Hydrolysis) | % | 16.4 | 17.0 |
| Ash | % | 6.2 | 9.6 |
| Ca | % | 0.80 | 2.25 |
| P | % | 0.99 | 0.92 |
| Ca:P Ratio | 1 to | 0.81 | 2.45 |
| Mg | % | 0.13 | 0.13 |
| K | % | 1.16 | 1.07 |
| Na | % | 0.41 | 0.39 |
| S | % | 0.58 | 0.57 |
| Fe | ppm | 61 | 73 |
| Zn | ppm | 184 | 168 |
| Cu | ppm | 21 | 20 |
| Mn | ppm | 39 | 39 |
| Mo | ppm | 1.7 | 1.3 |
| α-carotene | ug/g | 0.02 | 1.04 |
| α-tocopherol | ug/g | 14.20 | 12.3 |
| β-carotene | ug/g | 1.1 | 13.7 |
| β-cryptoxanthin | ug/g | ND | ND |
| Lutein | ug/g | 0.19 | 0.62 |
| Retinol | ug/g | 17.1 | 60.3 |
| Unknown | ug/g | 0.94 | 3.28 |
| Zeaxanthin | ug/g | 0.09 | 10.70 |
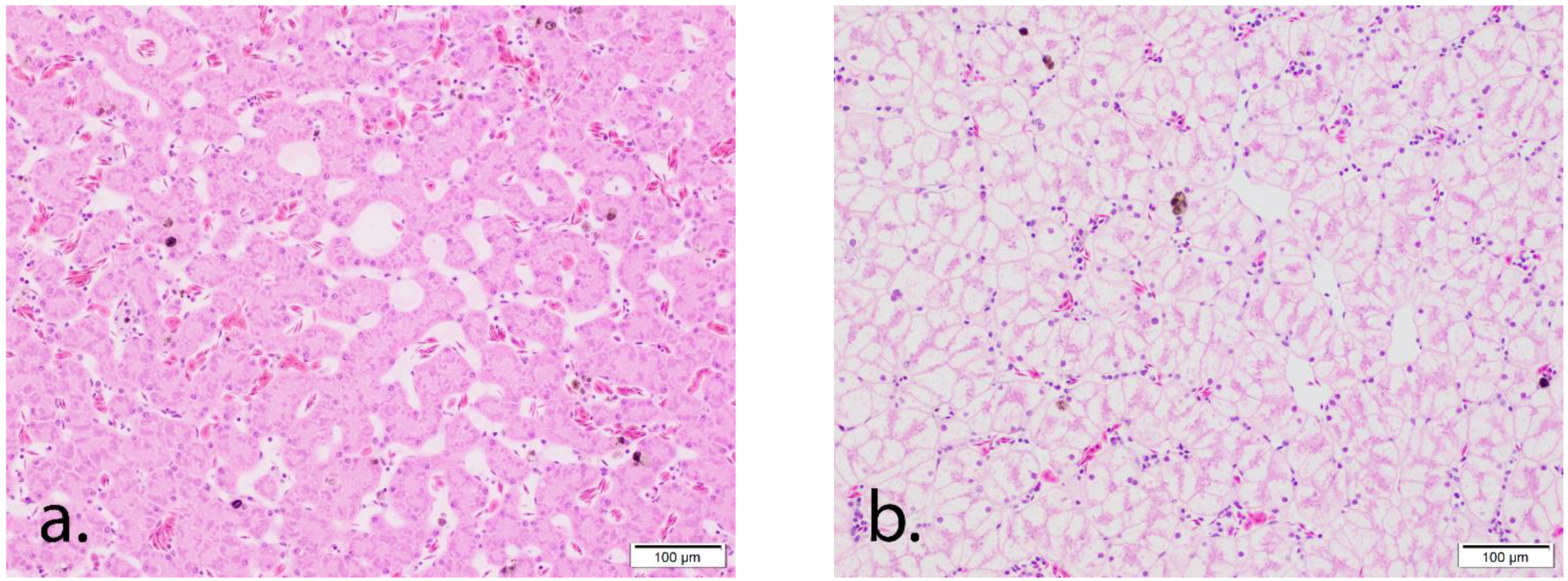
3.3. Histology Results
The sexes of the toads that were wild-caught and kept in care were histologically confirmed. Liver vacuolization was characterized by the expansion of the hepatocyte cytoplasm by indistinct vacuoles (microvesicular vacuolization) within compression and loss of the hepatic sinusoids. Vacuolation in this study increased in a time-dependent manner: Grade 0–2 (average 0.9 ± 0.55) in wild- caught toads; Grade 1–4 (average 3.4 ± 0.64) at Day 15; Grade 2–4 (average 3.9 ± 0.56) at Day 32; and Grade 3–4 (average 3.9 ± 0.13) at Day 62 (
Figure 1) [
26,
27]. This increase in hepatic vacuolation was not associated with any other histologic lesions such as necrosis, hemorrhage, or biliary duplication. Lesions in other tissues were limited to intestinal nematodiasis associated with minimal to mild inflammation which was observed in toads from all groups.
Figure 1.
Histologic images of liver vacuolation in Marine toads (Rhinella marina). (a) Grade 0 vacuolation in a liver from a wild-caught Marine toad. The hepatocytes are thin with no visible intracytoplasmic vacuoles and prominent sinusoidal spaces. (b) Grade 4 vacuolation in a liver from a Marine toad in human care for 62 days. Hepatocytes are diffusely, markedly expanded by cytoplasmic vacuoles with loss of sinusoids.
Figure 1.
Histologic images of liver vacuolation in Marine toads (Rhinella marina). (a) Grade 0 vacuolation in a liver from a wild-caught Marine toad. The hepatocytes are thin with no visible intracytoplasmic vacuoles and prominent sinusoidal spaces. (b) Grade 4 vacuolation in a liver from a Marine toad in human care for 62 days. Hepatocytes are diffusely, markedly expanded by cytoplasmic vacuoles with loss of sinusoids.
Table 3.
Marine Toad (Rhinella marina) Liver Vitamin A (Retinol), Vitamin E (α-tocopherol), and Carotenoid Concentrations (LS means ± SEM) as the Toads Adjusted from the Wild (Day 0) thru Day 62 ‡ in Human Care Consuming Two Different Diets †.
Table 3.
Marine Toad (Rhinella marina) Liver Vitamin A (Retinol), Vitamin E (α-tocopherol), and Carotenoid Concentrations (LS means ± SEM) as the Toads Adjusted from the Wild (Day 0) thru Day 62 ‡ in Human Care Consuming Two Different Diets †.
| | Day 0
Wild
(n = 10) | Day 9
Diet 1 †
(n = 7) | Day 9
Diet 2 †
(n = 7) | Day 15
Diet 1
(n = 7) | Day 15
Diet 2
(n = 7) | Day 32
Diet 1
(n = 7) | Day 32
Diet 2
(n = 7) | Day 62
Diet 1
(n = 7) | Day 62
Diet 2
(n = 7) |
|---|
Weight
(g) | 128.7 ± 15.31 | 121.3 ± 17.40 | 120.6 ± 20.68 | 135.9 ± 20.68 | 144.0 ± 17.40 | 150.4 ± 17.40 | 154.9 ± 20.68 | 171.1 ± 20.68 | 176.3 ± 17.40 |
| Nutrients |
Retinol
(µg/g) | 108 ± 21.3 x | 55 ± 23.8 x,y | 93 ± 27.4 x,z | 82 ± 27.4 x,y | 49 ± 23.8 x,y | 36 ± 23.8 y | 47 ± 27.4 x,y | 12 ± 27.4 y | 28 ± 23.8 y,z |
α-tocopherol
(µg/g) | 8.8 ± 0.5 a,d | 8.4 ± 0.59 a,c,d | 9.6 ± 0.69 a | 8.1 ± 0.69 a,d,e | 7.6 ± 0.59 d,e | 8.7 ± 0.59 a,d,e | 8.2 ± 0.69 a,d,e | 7.1 ± 0.69 b,c,e | 5.5 ± 0.59 b |
Apocarotenoid
(µg/g) | 1.8 ± 0.30 a | 0.9 ± 0.34 b | 0.9 ± 0.39 a,b | 0.7 ± 0.39 b | 0.6 ± 0.34 b | 0.4 ± 0.34 b | 0.6 ± 0.39 b | 0.1 ± 0.39 b | 0.1 ± 0.34 b |
β-carotene
(µg/g) | 4.0 ± 2.73 | 11.4 ± 3.05 | 3.4 ± 3.52 | 2.7 ± 3.52 | 2.3 ± 3.05 | 1.7 ± 3.05 | 1.8 ± 3.52 | 0.5 ± 3.52 | 1.2 ± 3.05 |
β-carotene ester
(µg/g) | 1.4 ± 0.28 a,b | 2.1 ± 0.31 a | 1.1 ± 0.36 b,c | 0.6 ± 0.36 b,c | 0.8 ± 0.31 b,c | 0.7 ± 0.31 b,c | 1.0 ± 0.36 b,c | 0.3 ± 0.36 c | 0.3 ± 0.31 c |
β-cryptoxanthin
(µg/g) | 2.2 ± 0.49 | 0.7 ± 0.54 | 0.5 ± 0.63 | 1.0 ± 0.63 | 1.0 ± 0.54 | 0.4 ± 0.54 | 0.5 ± 0.63 | 0.4 ± 0.63 | 0.5 ± 0.54 |
Lutein
(µg/g) | 9.4 ± 1.56 a | 6.7 ± 1.74 a,b | 2.7 ± 2.01 b,c | 3.6 ± 2.01 a,c | 3.6 ± 1.74 a,c | 2.0 ± 1.74 a,c | 2.4 ± 2.01 a,c | 0.2 ± 2.01 c | 1.4 ± 1.74 c |
Lutein ester
(µg/g) | 1.4 ± 0.30 x | 1.3 ± 0.34 x | 1.6 ± 0.39 x | 0.9 ± 0.39 x,y | 0.8 ± 0.34 x,y | 0.7 ± 0.34 x,y | 0.7 ± 0.39 x,y | 0.1 ± 0.39 y | 0.3 ± 0.34 y |
Zeaxanthin
(µg/g) | 5.6 ± 1.17 x | 4.7 ± 1.31 x,y | 3.1 ± 1.51 x,y,z | 2.4 ± 1.51 x,y,z | 2.8 ± 1.31 x,y,z | 1.9 ± 1.31 y | 1.5 ± 1.51 y | 0.1 ± 1.51 z | 0.7 ± 1.31 z |
| Zeaxanthin ester (µg/g) | 2.2 ± 0.55 | 1.6 ± 0.61 | 2.7 ± 0.70 | 1.4 ± 0.70 | 1.1 ± 0.61 | 1.2 ± 0.61 | 0.9 ± 0.70 | 0.1 ± 0.70 | 0.5 ± 0.61 |
4. Discussion
Despite the addition of Repashy dusting supplementation to crickets in Diet 2, there were still significant decreases in all marine toad nutrient concentrations analyzed over the 62-day study period, when comparing those consuming both treatment Diet 1 and 2 to the free-ranging marine toad wild diet nutrient values (Day 0). These findings warrant concern for maintaining free-ranging vitamin A, vitamin E, and carotenoid concentrations via the consumption of commonly fed commercial diets in marine toads and likely other amphibian species.
Hepatic concentrations of Vitamin A are currently thought to be the ideal way to evaluate concentrations in amphibians since approximately 90% is stored in the liver [
5,
12,
28,
29]. Retinol values for Day 0 of free-ranging toads in this study averaged 108 ± 21.3 μg/g. By the end of the 62 days of this study, marine toads on Diet 1 had an average of 12 ± 27.4 μg/g and on Diet 2 had an average of 28 ± 23.8 μg/g. The decrease in retinol is similar to Freel et al.’s significant decrease [
12]. Freel et al.’s study was also conducted with Zoo Miami culled marine toads in the summer season, and the hepatic retinol values for Day 0 for free-ranging marine toads averaged 87.7 ± 16.12 μg/g. By Day 50 of their study, marine toads with Diet 1 (crickets gut-loaded with Mazuri Hi Calcium Gut Loading diet with supplemental carrots and sweet potatoes) averaged at 18.0 ± 3.68 μg/g, and marine toads on Diet 2 (crickets gut-loaded with Maruzi Hi Calcium Gut Loading Diet without any supplementation) averaged at 11.0 ± 1.72 μg/g. Although there is a similarity with the decrease in retinol, there is still a higher level of retinol concentration in the marine toads in the current study at Day 0 than the other time points. The reason for this difference, however, is unclear; possibilities could include yearly environmental changes, stress, diet availability, age, sex, or other factors.
There were higher concentrations of β-carotene, α-tocopherol, β-carotene, retinol, and zeaxanthin in the nutrient Repashy dusting supplement compared to the concentrations found in the livers of the marine toads. The crickets alone had significantly less concentrations of these nutrients than both the Repashy dusting supplement and the liver values. It was also noted that α-tocopherol values decreased when the crickets that were dusted were analyzed after three days (
Table 2). The crickets that were dusted had increased levels of β-carotene and retinol, but decreased levels of α-tocopherol when fed just the cricket diet for three days. General vitamin A mechanisms have been sequenced and it is believed that some amphibian species may be able to metabolize Vitamin A, similarly to many mammal species, although much is unknown and unstudied [
3,
5]. It also may be that not all the nutritional supplements utilized are actually being ingested by the marine toads in the current study. Different forms of carotenoids, vitamin A, and vitamin E included within the cricket diets and dusting supplements themselves may provide varying results via altered digestion and absorptions as shown in other species, although the mechanisms are not clear in amphibians [
5,
6,
30,
31]. Regardless, the decrease in vitamins and carotenoids in the liver while consuming high-quality and supplemented diets is concerning and fuels the need for more research.
The vitamin A decreases during human management noted within this study indicate that there may need to be changes to the diet and supplementation strategies used within many institutions. While the animals in this study did not demonstrate any health concerns visually or histologically, the lower concentrations of the vitamin A, vitamin E, and carotenoid noted could lead to various health concerns such as squamous metaplasia, stunted growth, gastric bloat, [
8] and decreased reproductive success and susceptibility to diseases [
4]. These clinical symptoms may not have been shown due to the short time frame of this study, which may be an area of study to investigate due to the concerns of nutritional deficiency of toads in human care. These clinical manifestations of vitamin A deficiency are concerning to the well-being of amphibians, especially with the noted decline in worldwide populations.
With there being little information about vitamin E deficiency in amphibians [
12], it is difficult to assess the minimum concentrations of vitamin E required for their well-being. The α-tocopherol levels of the free-ranging toads in this study for both Diets 1 and 2 decreased despite the additional vitamin E supplementation. In comparison to Freel et al.’s results, the marine toads in this study had higher hepatic α-tocopherol levels on Day 0. Freel et al.’s average on Day 0 for free-ranging toads were 9.2 ± 0.54 μg/g, with concentrations decreasing to 7.7 ± 0.44 μg/g for Diet 1 and 7.7 ± 0.54 μg/g for Diet 2. Although the concentrations of α-tocopherol started off higher at Day 0 for this study, it decreased to a similar if not lower amount for both diets by Day 62 when compared to the results by Freel et al. [
12]. It may be possible that more Repashy needs to be supplemented or ingested since it has more α-tocopherol than just the crickets.
When comparing the two diet treatments in this study, hepatic retinol concentrations tended to decrease more slowly with Diet 2 at Day 32 compared to Diet 1, which indicates supplementation with Repashy may allow the marine toads to have vitamin A concentrations closer to wild marine toads.
Animals fed Diet 2 were expected to have higher-concentration of the analyzed vitamins and carotenoids due to the additional supplementation with Repashy. Analysis of Repashy supplementation revealed 30.8 μg/g of α-tocopherol, and 286.6 μg/g of retinol as shown in
Table 1. It is possible that the human-managed environment and digestion of the crickets could factor into the lower hepatic concentrations of these nutrients, or formulation of supplementation is allowing for excretion prior to being stored in the liver. It has been shown that live crickets dusted with supplementation can clean over 50% of the supplement off their bodies within a couple of minutes of the dusting method [
6,
32]. With the results, further research may be warranted regarding using a different form of vitamin A supplementation or technique in feeding amphibians.
For all toads, there was a time-dependent increase in hepatic vacuolation. The significance of this finding is unclear. In general, hepatic vacuolization is understood as the result of metabolic derangement and has been cited as the cause of subcutaneous edema and morbidity/mortality in other amphibian species [
33,
34]. In the absence of subcutaneous edema and other hepatic lesions such as necrosis or hemorrhage, this change was unlikely to be clinically relevant at the time of death. Additionally, it should be kept in mind that these toads were fed more than they would normally consume in the wild or in other zoological species exhibits to prevent group housing-related cannibalism as noted within this species.
The Day 0 toads were not dewormed, shipped, or housed and fed within a managed care setting. Thus, stress may be a factor that affected Day 0 toads differently across all analyzed parameters compared to the other time points. However, several co-authors from the current paper previously studied urinary corticosterone as a measure of potential stress within marine toads used for nutrition and veterinary care research [
35]. The toads from the prior paper and the current paper were captured, transported, housed, and euthanized in a nearly identical manner [
35]. Data indicated that managed care animals had similar yet lower urinary corticosterone and therefore potentially lower stress than free-ranging toads (potentially due to the ease of ad libitum diets and easy access to mates) [
12,
35]. Thus, the scientific community has much to learn about stress (eustress and distresss) within amphibian species.
It should be noted that this study only fed crickets to the toads. This was conducted in order to compare to the Freel et al. paper [
12] and to better understand the diet nutrients provided. However, both the ad libitum quantity of crickets and only feeding one insect species are not currently recommended amphibian husbandry methods (although still common-place strategies in the pet trade and some zoological facilities). Additionally, our results may only be valid for the invasive marine toad utilized in this study [
3]. While this species has been increasingly used as an amphibian model [
12,
15,
16,
17], it may not reflect all anuran or all amphibian species [
3,
5]. Different diets for amphibians and the vitamin sources and concentrations within the system of amphibians may be the next step for investigation. Adult anurans are carnivorous, and their natural diet includes other invertebrates such as various species of worms and roaches [
3]. Additional supplementation with these other invertebrates may also be another area of further research, due to the dusting method being cleaned off of crickets [
6,
27], or looking into different cricket diets, based on the results of nutritional cricket analyses. Vitamin A and vitamin E homeostasis are poorly understood in toads (and in all amphibians) and therefore research into deficiencies and toxicities of these nutrients is paramount for human management conservation programs [
5].

 ,
, 
{kind=link}
