1. Introduction
Encouraging positive welfare states for animals under human care is a central mission for zoos and aquariums around the world. One method for assessing welfare is through behavioral observation. Generally, positive welfare is associated with the exhibition of species-specific behaviors, while stereotypic behaviors [
1,
2] are considered an indication of diminished welfare [
1,
3,
4]. Stereotypic behaviors can reduce the amount of time engaged in species-specific behaviors [
5,
6]. Enrichment, exhibit additions, and alteration in husbandry routines can be used to provide opportunities for choice and control and the mitigation of the occurrence of stereotypy [
2,
5,
6,
7,
8].
Providing choices for an animal within their environment can add complexity and present challenges and opportunities to exhibit species-specific behaviors [
9,
10]. Automatic feeders to disperse food widely and at set or random intervals [
7], ice blocks containing food items [
11], or unpredictability in schedules [
9,
12] have all been employed as forms of environmental enrichment [
10] to introduce variety in an animal’s daily routine. Another method of providing choice is through offering access to various enclosure areas, including those away from guest view [
2,
13,
14]. Presenting options for space use may reduce stress-related behaviors [
13,
14,
15,
16,
17]. For example, eastern black-and-white colobus monkeys (
Colobus guereza) and chimpanzees (
Pan troglodytes) with access to both indoor and outdoor areas were reported to be more active and interacted with enrichment objects more often, suggesting improved welfare [eastern black-and-white colobus monkeys: [
8]; chimpanzees: [
18]. In addition, multiple studies have shown that when animals were given free access to more than one enclosure area, signs of behavioral agitation lessened; this was determined to most likely be due to increased choice rather than increased stimulation from additional areas [giant pandas,
Ailuropoda melanoleuca: [
5,
13]; eastern black-and-white colobus monkeys: [
8]; Asian elephants,
Elaphus maximus: [
19]].
Bears are commonly housed within facilities accredited by the Association of Zoos and Aquariums [
20] but are prone to exhibiting stereotypic behaviors [
13,
14,
21,
22]. Pacing is the most frequently observed stereotypy in bear species under human care [
5], which is thought to be the result of a combination of factors, including space restriction, monotony, and the inability to complete the idiosyncratic expression of some natural behaviors [
4,
5,
23,
24]. Several studies have demonstrated a reduction of aberrant behaviors through enrichment, but few have examined how access affects behavior. One study reported that offering a honey-filled log and scatter feeding reduced the pacing of a sloth bear (
Melursus ursinus) and an American black bear (
U. americanus) [
7]. Various food items within ice blocks were found to increase activity levels and decrease the occurrence of stereotypic behaviors in a Kodiak bear (
U. arctos middendorffi) and a polar bear (
U. maritimus) [
11]. While there is limited research examining how increasing access to a variety of locations impacts behavior among bears, one study found that polar bears with access to off-habitat areas reduced engagement in stereotypy and increased time spent in species-typical behaviors [
14]. In another study, giant pandas (
A. melanoleuca) exhibited a reduction in the occurrence of stress-related behaviors when given the free choice of habitat location [
13].
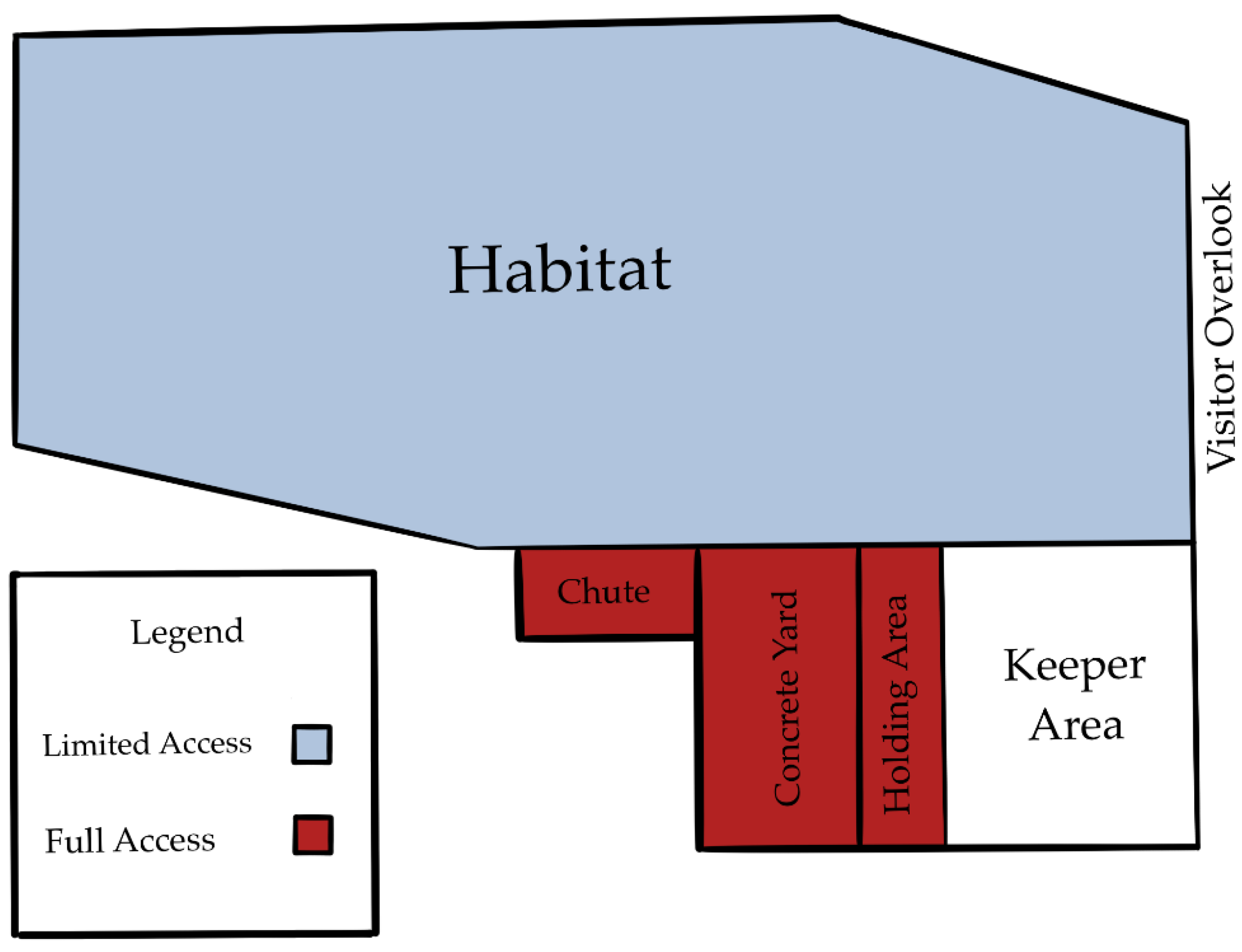
Our study aims to assess how greater choice, in the form of access to multiple areas within a habitat, effects the welfare of American black bears. We quantified and compared behaviors exhibited by two American black bears housed at the North Carolina Zoo across days where access to alternate spaces was limited and those where access was available. We predicted that giving the bears choice and control over their space use via access to various habitats would reduce stereotypic behaviors (pacing) and increase species-specific behaviors (foraging). While some level of stress is expected and species-appropriate, we are focused on stereotypic patterns, specifically pacing, which are considered aberrant and undesired [
1,
4]. Because visibility for the guest view is a common concern across zoos [
25,
26,
27,
28], we also assessed the effect of access on guest visibility. We believe that this study will supplement our understanding of how to promote good welfare among zoo-housed carnivores and inspire further research into the effects of choice and control.
4. Discussion
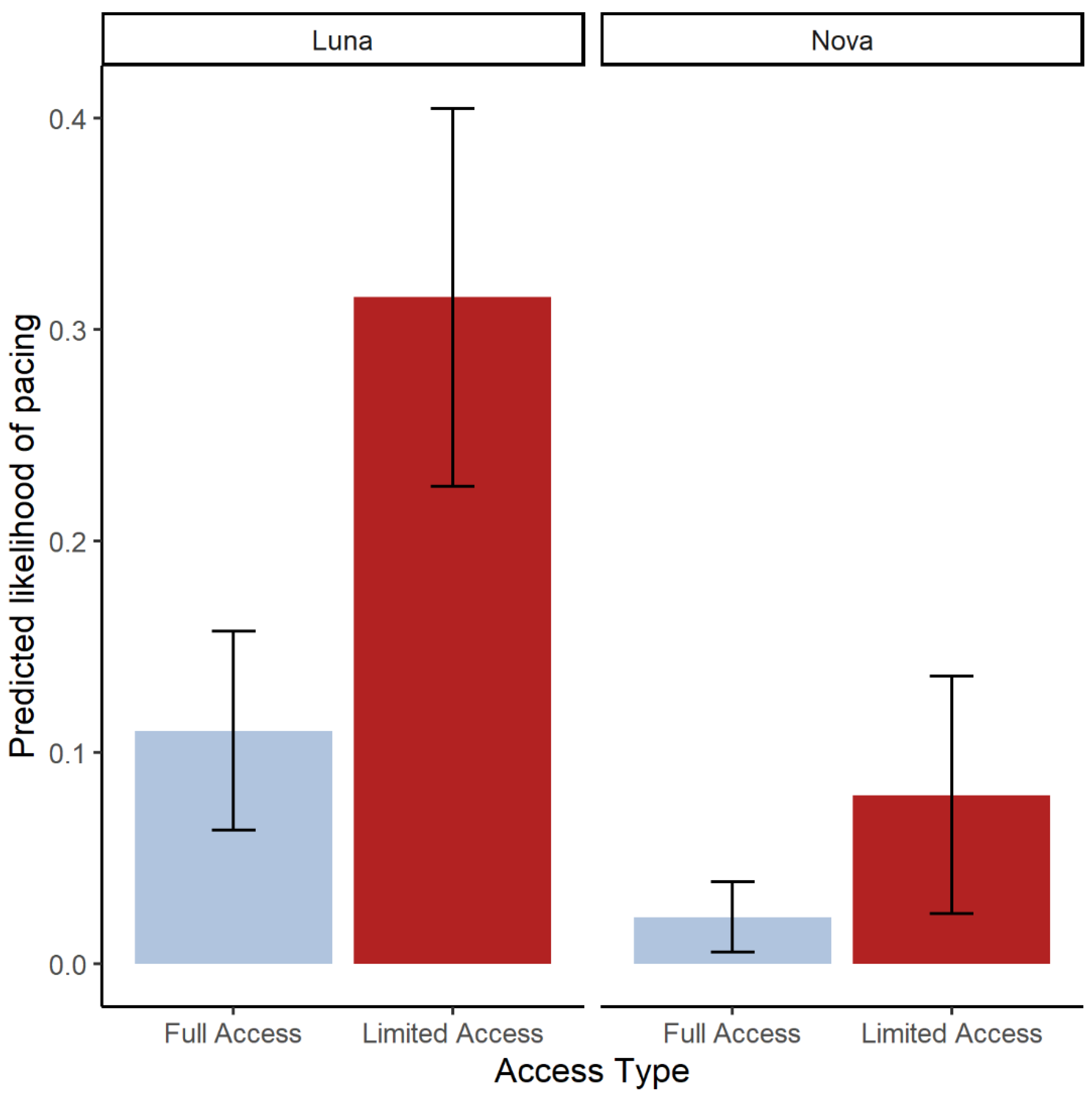
We report that access to multiple areas within a habitat significantly reduced stereotypy and increased foraging among two zoo-housed black bears. In addition, full access only reduced the likelihood of visibility for one bear, while the other remained in guest view, suggesting that this method of enrichment may not always affect the guest experience. Overall, this case study provides evidence that environmental enrichment in the form of access improves welfare while having a limited impact on visibility to visitors.
This case study builds on the current understanding of the efficacy of providing choice and control to animals under human management [
13,
14,
18,
22]. In particular, choice and control over space use was found to improve welfare, even when the additional space was not utilized or highly enriched [
13,
14,
18,
22]. When a pair of male and female sibling polar bears were given the choice to access indoor dens, pacing was reduced, and social play increased [
14]. For carnivores and other species that require larger home range sizes, artificial housing can constrain locomotion, which may be associated with an increase in stereotypical behaviors [
45,
46,
47]. Providing access to more spaces, even if unused, can serve as a form of choice and control for the animal, enabling options for space use [
13,
14,
21]. The benefits of providing access can lead to a decrease in stress and time spent in stereotypic behaviors, allowing for more time to be spent performing species-typical behaviors. [
10,
14,
22,
48].
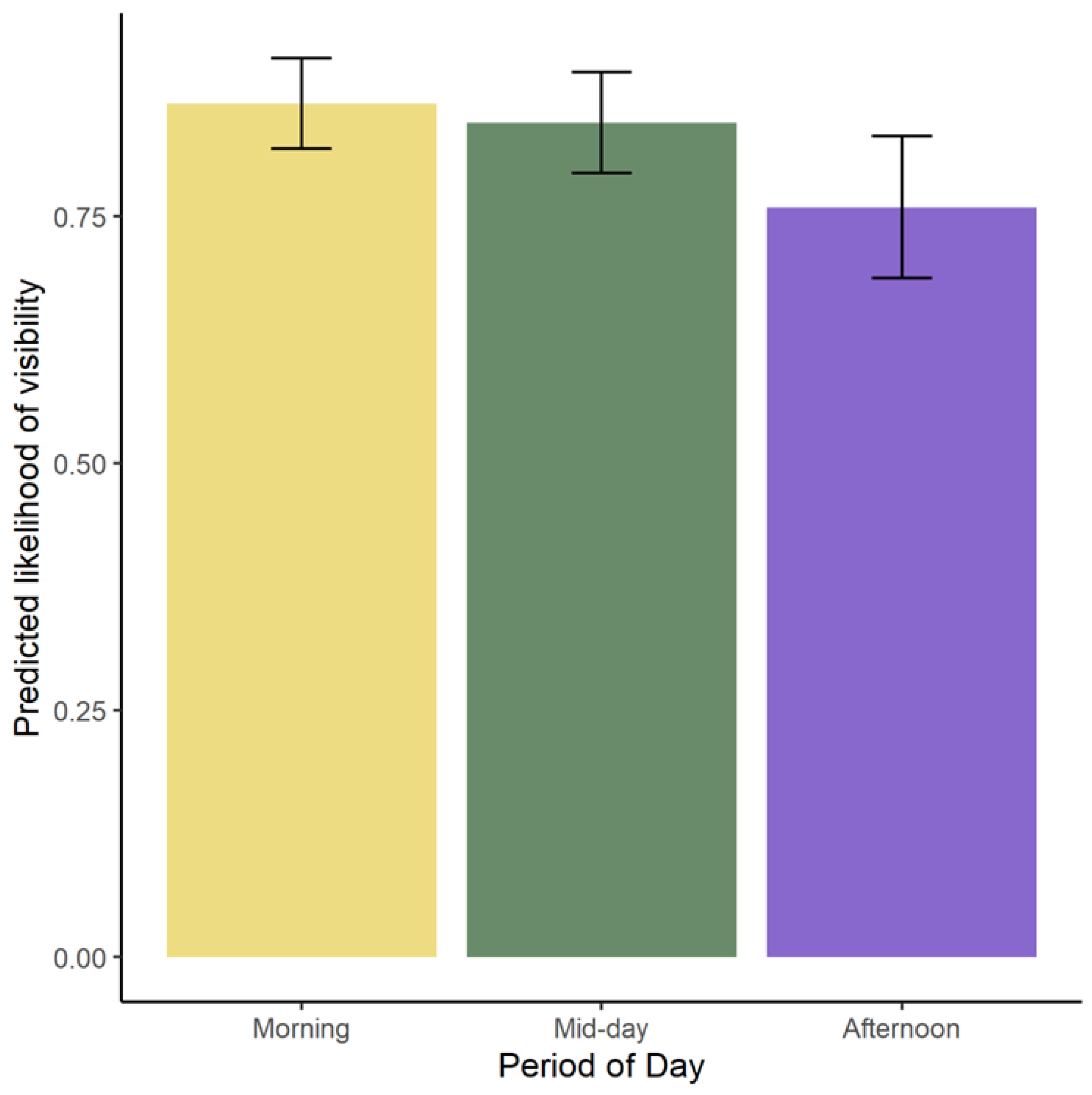
It should be noted that time of day was found to have a significant effect on behavior. We believe that a few factors may have contributed to the increase in Nova’s foraging activity in the mid-day and afternoon. Guest counts wane towards the end of the day and that may reduce the occurrence of stress-related behavior. The bears are fed during mid-day so we can assume that they forage more in the afternoon. An increase in pacing is likely due to husbandry scheduling and routines, where the anticipation of feedings and keeper interactions can influence behavior [
21,
38,
39,
44]. Our findings are consistent with previous studies on carnivores, demonstrating that stereotypic behaviors and activity are closely tied to the time of day due to routine husbandry schedules [
21,
38,
49]. Anticipatory pacing may occur before an expected feeding. Due to feeding occurring over a two-hour period, a distinction between pre-feeding anticipatory behavior and stereotypic behavior was not made; however, if possible, this distinction should be noted [
50]. We believe that the visibility was highest in the morning due to husbandry routines. The keepers service the habitat as early as possible and leave diet and enrichment around the habitat. The bears are more likely to search for enrichment and diet following this. Our paper further highlights the importance of considering the time of day when monitoring welfare and the effects of husbandry changes.
Finally, we explored the guest experience, as it relates to access. Because visitor density and the duration of time spent observing a habitat generally increases when animals are visible and active [
42,
51], animal visibility is a primary concern across zoological institutions [
25,
26,
27,
28]. We report that full access significantly decreased pacing behavior for both bears, but only affected the visibility of one bear, Luna. However, it should be noted that among the two bears, at least one was visible in the habitat 74% of all the observed time, when given full access. As visitor perception of a zoo decreases when stereotypical behaviors are observed [
43], beyond welfare and ethical concerns, it is useful to the success of an institution to mitigate stereotypy from the guest perspective. With the reduction of stereotypic behavior and a minimal effect on visibility, our study suggests the value of providing access to both the individual animal and the guest.
Two important caveats are notable in this study. First, our study was limited to two focal animals. Future research would benefit from exploring the effects of access on various behaviors across carnivores or various ages and sexes.
Second, given the constraints of the daily routine for animal caretakers, we implemented a consistent schedule for limited and full access days. However, staff ensured management procedures, including feeding times, training schedules, habitat servicing routines, and so on, were consistent across days of the week. In addition, guest numbers do not often fluctuate during weekdays, so visitor numbers are not expected to have had an impact. Therefore, this consistent schedule is unlikely to have affected our results. Future work, though, may benefit from exploring this possibility.
Overall, our study suggests that access can reduce stereotypic behaviors (pacing) and increase desired behaviors (foraging) without fully compromising visibility for zoo visitors. Choice and control are more likely responsible for these outcomes than enrichment and stimulus diversity since the added areas of access were not more enriched than the habitat. The individual bears had varying degrees of change in response to access, which we believe are due to personality differences. Building habitats in zoos is very time-consuming and expensive. Opening gates to areas that are already built presents a no cost and simple method to improve welfare under human care. This case study can serve as the foundation for future research to explore the value of choice and control as a method to enrich the lives of zoo-housed carnivores.








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}