
Immune Modulatory Effects of Ketogenic Diet in Different Disease Conditions
by
 , and
, and
Shivani Srivastava
1,†,
Vishakha Anand Pawar
2,†,
Anuradha Tyagi
3,†,
Kanti Prakash Sharma
4,†,
Vinay Kumar
5,* and
and
Surendra Kumar Shukla
6,* 1
Department of Pathology, School of Medicine, Yale University, New Haven, CT 06520, USA
2
The University of Texas MD Anderson Cancer Center, Houston, TX 77030, USA
3
Department of cBRN, Institute of Nuclear Medicine and Allied Science, Delhi 110054, India
4
Department of Nutrition Biology, Central University of Haryana, Mahendragarh 123029, India
5
Department of Physiology and Cell Biology, The Ohio State University Wexner Medical Center, 473 W 12th Ave, Columbus, OH 43210, USA
6
Department of Oncology Science, OU Health Stephenson Cancer Center, Oklahoma City, OK 73104, USA
*
Authors to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Immuno 2023, 3(1), 1-15; https://doi.org/10.3390/immuno3010001
Submission received: 29 November 2022
/
Revised: 21 December 2022
/
Accepted: 22 December 2022
/
Published: 25 December 2022
(This article belongs to the Section Autoimmunity and Immunoregulation)
Abstract
:Interceding nutrients have been acquiring increased attention and prominence in the field of healing and deterrence of various disorders. In this light, the present article encompasses several facets of ketogenic diet as an immunomodulator with respect to its expansive clinical applications. Accordingly, several scientific records, models, and case histories, including viral infections, cancer, chronic diseases, e.g., cardiovascular diseases, epilepsy, as well as numerous other neuro-disorders, are assembled, revealing a profound influence of KD in favor of improvement in the patient’s condition. We accentuate possible manifold mechanisms of KD that require further exploration.
1. Introduction
Lifestyle usually plays an important role in human beings’ health. Various categories of diets, e.g., “low carbohydrate diets”, “traditional regional diets”, and “plant forward diets”, are known, followed, and prescribed in the interest of improving and/or maintaining the well being of respective individuals. An extensive survey and research is being conducted in almost all the corners of the world to study the impact of these diets on human beings. Ketogenic diets (KD) encompass a lower consumption of carbohydrates, adequate protein, and a high fat regimen which induces ketone body production via mimicking the metabolism of the fasting state without significant calorie deprivation.
The long-chain triglyceride (LCT) diet, the modified Atkins diet (MAD), the low glycemic index procedure, and the medium-chain triglyceride (MCT) diet are four major types of ketogenic diets (KD) that have been shown to be effective in many scientific reports [1,2]. Since 20th century, KD has been used for the treatment of epilepsy [3]. KD started gaining attention of the clinical and research community in the past decade due to its wide ranging therapeutic functions in several diseases, e.g., polycystic ovarian syndrome, obesity, dyslipidemia, type 2 diabetes, hypertension, Alzheimer’s disease, cancer, respiratory compromise, cardiovascular health, cancer, intestinal disorders, Parkinson’s disease, and malignancies (Figure 1) [1,4,5].
The genetic variation within the gut microbiome of an individual could be improved by following KD. KD could alter the diversity of the microbiome by increasing the ratio of Bacteroidetes to Firmicutes. KD might affect epigenome by stimulating a signaling molecule called β-hydroxybutyrate (BHB). As an adjuvant therapy, KD can be used to starve cancer cells by making them more defenseless towards radiation and chemotherapy, owing to its Warburg effect [4]. KD can be modified to fit a vegan or vegetarian practice, with plant-based proteins, fats, etc. or can be adapted to consume animal products, giving flexibility to choose dietary preference to patients of diabetes and obesity [6]. According to recent reports, low-carbohydrate KDs could result in weight loss as well as favorable alterations in high-density lipoprotein cholesterol and serum triglycerides [7].
Similar metabolic patterns are seen in fasting studies. In one of the rheumatoid arthritis case study, patients’ β-hydroxybutyrate (BHB) levels were found to be elevated upon following intermittent fasting. This was correlated to diminishing disease symptoms. In this regard, KD also holds a great potential as a therapeutic alternative in treating arthritis. However, more evidence will be helpful to support this outcome [8]. A low-calorie ketogenic diet could be considered as a promising therapy to improve psoriasis-related dysmetabolism, upon extensive reformation of the inflammatory as well as metabolic state of the subject [9]. Moreover, in a recent study, KD was also shown to inhibit colitis and protect intestinal barrier decay [10].
2. Immune Modulation by Ketogenic Diet in Viral Infection
The COVID-19 pandemic has contributed to the deaths of more than 6 million people. Studies have provided the cellular mechanisms for understanding the composite SARS-CoV-2 access route on the cell’s surface in the host. SARS-CoV-2 binds to ACE2 receptors and excessively induces the secretion of TNF-α, IL-6, and IL-1 are pro-inflammatory cytokines, promoting the progression of acute respiratory distress syndrome (ARDS) [11]. A eucaloric KD has been proposed to have potential therapeutic role against COVID-19 due to its role in suppressing critical risk complications, such as anti-inflammation, hypertension, type-2 diabetes, obesity, and metabolism modulation [12,13]. An MCTs-rich KD can provoke lipid metabolism switch, could disfavor infection and replication of virus, and can inhibit the cytokine storm [14]. This swapping of the host lipid metabolism can also be achieved by consumption of coconut-rich medium-chain fatty acids along with olive oil, followed by fasting for 8–12 h and a dinner rich with vegetables and fruits, resulting in activation of the ketogenic pathway [14].
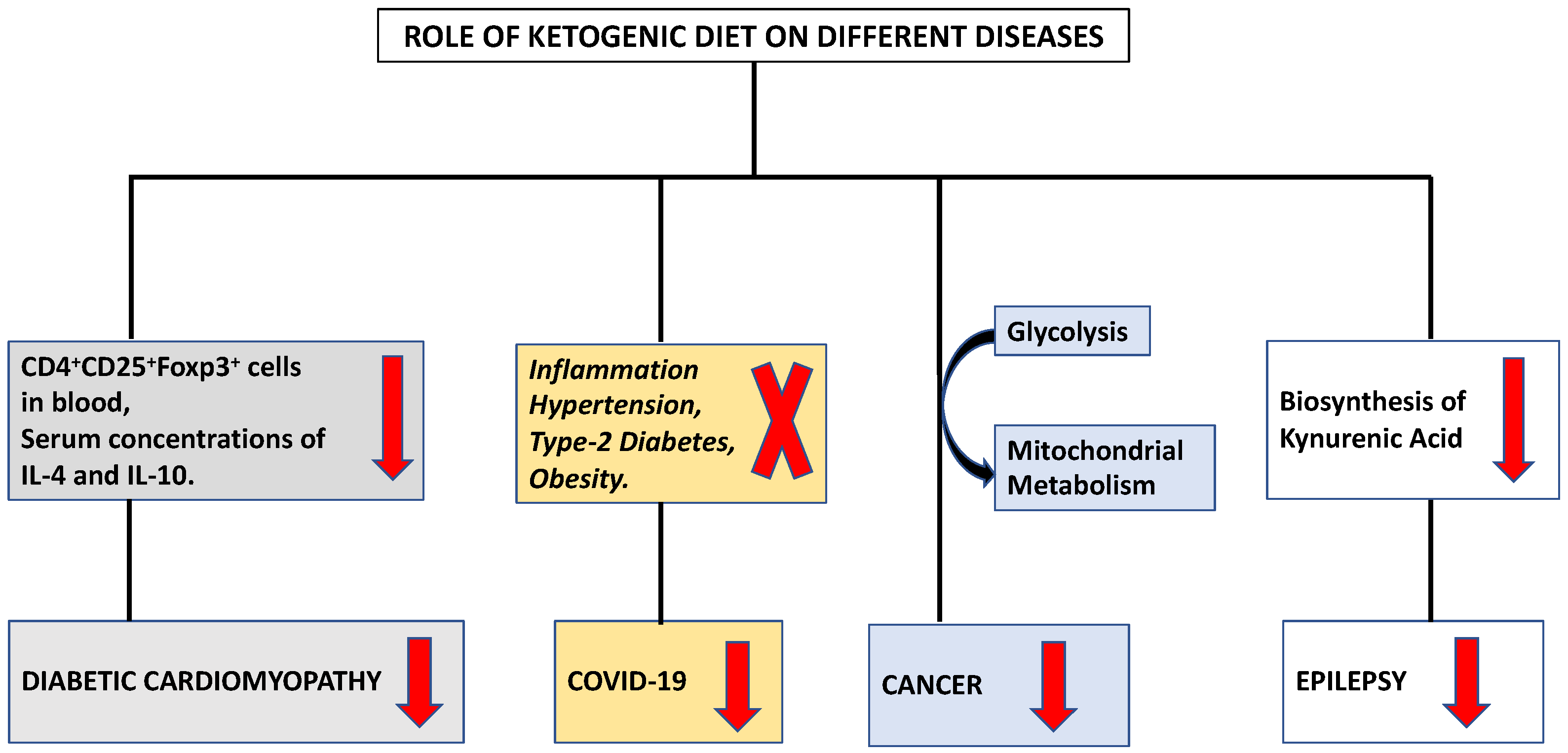
Amongst the 68 COVID-19 patients who received a eucaloric standard diet, 34 patients receiving KD were observed to have a lower risk of mortality [15]. Ketone bodies like β-hydroxybutyrate (BHB) maintain the redox balance by providing an alternative carbon source for oxidative phosphorylation (OXPHOS) and the synthesis of bioenergetic amino acids and glutathione. KD induces levels of interferon-γ by CD4+. Under a SARS-CoV-2 stimulated ARDS stress, the exhausted and glycolysis skewed T cells can be reprogrammed metabolically by BHB to perform OXPHOS [16]. Activation of ketogenesis reduces pathogenic monocytes in the lungs of aged mice infected with mCoV-A59 (murine beta coronavirus infected with mouse hepatitis virus strain-A59), inactivates the NLRP3 inflammasome, and increases the tissue-protective γδ T cells [3,17,18]. There are many proposed molecular mechanisms that explain the therapeutic role of the exogenous ketone-based metabolic therapy in combination with a moderately high-fat diet against the cytokine storm induced by severe SARS-CoV-2 infection. By reducing glucose uptake into ILC2s, KD reduces lung inflammation. A study in mice proved that KD potentially activated a γδ T cell response, leading to decrease IAV mortality. KD is also known to normalize the disease induced upregulation of Th17/Treg Ratio [19]. In COVID-19 patients, KD has been proven to provide superior energy by directing human CD8+ T cells towards aerobic mitochondrial metabolism [20].
3. Ketogenic Diet Mediated Immune Regulation in Cancer
The ketogenic diet (KD) stimulates a metabolic switch from glycolysis into mitochondrial metabolism, the differential stress resistance phenomenon with high tumor control ability and lower normal-tissue complications, making it an intriguing dietary approach for cancer patients who are under the supervision and follow-up of a healthcare provider (Figure 2) [21]. Initial findings regarding the role of food consumption curtailment in tumor growth was reported by Rous in 1914 [22]. Later, Tisdale et al. (1987), [23] reported the anti-tumor effect of ketogenic diet. Afterwards, many researchers supported the utilization of the ketogenic diet in various animal models through numerous mechanisms [24,25,26].
The ketogenic diet was utilized for the management of human malignant brain tumor [27]. Researchers utilized an immunocompetent mouse model of malignant glioma and observed that a ketogenic diet led to blood glucose reduction, blood ketones elevation, and overall life extension [28,29]. CTLA-4 (cytotoxic T lymphocytes associated antigen 4) and PD-1 (Programmed cell death protein-1) are the immune checkpoints utilized as targets for treatment of multiple tumor types [30]. By lowering PD-L1 protein levels and enhancing the expression of type I interferon and antigen presentation genes, the introduction of the ketogenic diet increases the effectiveness of anti-CTLA-4 immunotherapy [31] resulting in anti-proliferative effects due to cell cycle extension.
IFN induced chemokines, such as CXCL-9, CXCL-10, and CXCL-11, can upregulate T-cell infiltration in melanoma [32] and CT26+ mouse model [33], thus enhancing immunity against tumor. Moreover, 3-hydroxybutyrate, a principal ketone body generated via ketogenic diet consumption was reported to induce T-cell dependent tumor growth interruption in belligerent tumor models [34]. In a glioblastoma mouse model, the ketogenic diet enhanced anti-tumor primary and acquired immune response by promoting cytolysis mediated by CD8+ T cells and increased CD4+ T-cells infiltration together with T cell killing activity [35]. Further, Sun et al. 2022 [36] also demonstrated the inhibitory role of ketogenic diet on tumor growth by enhancing the Th1 cells and cellular immune function. An increase in CD8+ T-cells and decrease in CD4+ FOX P3+ T-cells in tumor tissue, attenuation of PD-L1, and CTL4 immunosuppression were the other observed mechanisms of action of the ketogenic diet in a tumor model [33].
Although PI3K enhanced the downstream of both insulin receptor and IGF-1R, PI3K/Akt dysregulation is directly linked to neoplasmic development and increased resistance to cancer therapy [37]. There are many different factors and micro-environments where mTOR signaling is modulated [38]. Growth factors, mitogens, PI3K, activated AMP kinase, and hormones such as insulin all operate as stimulants for mTOR signaling [38]. AMP-activated protein kinase (AMPK), phosphatidylinositide 3-kinase (PI3K), and mTOR are all adversely impacted by decreased nutritional patterns [39]. The tumor suppressor activity is increased by a ketogenic diet, which inhibits mTOR signaling by activating the AMPK signaling pathway [39]. Calorie restriction via the ketogenic diet decreased the expression of pro-inflammatory markers, including cyclooxygenase 2, nuclear factor-k, and macrophage inflammatory protein 2, in a mouse model of astrocytoma [40].
Macrophages are known to have regulatory functions in modulating tumor immune response. Studies reported that ketogenic diet utilization led to switch of tumor associated macrophage from M2 to M1 phenotype which inhibited tumor progression [41]. M1 macrophage can promote and amplify the Th1 type response inhibiting tumorigenesis [42]. Equilibrium between Th1/Th2 responses is crucial for cancer development as Th2 cytokines (IL-4, IL-5, IL-13) promote tumor growth and progression. An enhanced Th1/Th2 ratio was demonstrated by implementation of the ketogenic diet in a colon tumor allograft mouse model by up regulating the Th1 driven immune response and inflammatory response provided beneficial effects against tumor [33]. Although the ketogenic diet has several positive effects, it may not be able to stop tumors from developing, but it can postpone their growth and increase survival rates [43]. Furthermore, when used in conjunction with conventional radiotherapy or chemotherapy, KD exhibits a synergistic effect on the treatment of cancer [29,44].
4. Ketogenic Diet Mediated Immune Regulation in Cardiovascular Diseases
The four categories of cardiovascular disease, frequently referred as heart events, are as follows: Aortic atherosclerosis, cerebrovascular disease, peripheral artery disease, and coronary artery disease (CAD) [45,46,47]. Immune cells also play important role in heart failure, particularly to pathological CD4+ T-lymphocytes during ischemic heart failure and heart remodeling [48]. Humans practice fasting for various reasons including religious, ethical, health reasons etc. since ancient times. Visioli et al. (2022) concluded that dietary intake manipulation to reduce calorie intake, intermittent fasting, and prolonged fasting are included in human culture possibly because of their positive effect on health [49]. Evidence showing the relationship between the differentiation and functioning of immune cells with reference to nutrient metabolism is accumulating in the scientific literature. To induce ketosis without limiting fat intake, the keto diet, also known as the ketogenic diet, consists of a low-carbohydrate diet with a moderate amount of protein restriction [50].
A low carbohydrate diet for longer period depletes the glycogen store of the body and stimulate keto-genesis, making ketone bodies the only source of energy. In addition to serving as energy sources, these ketone bodies function as significant signaling molecules that influence the expression and activity of transcription factors including PGC-1 and sirtuins (SIRTs) [51,52], poly-adenosine diphosphate [ADP]-ribose polymerase 1 (PARP1), and ADP ribosyl cyclase [53] fibroblast growth factor 21 and nicotinamide adenine dinucleotide (NAD+) [54]. Additionally, calorie restriction inhibits the PI3K/Akt/mTOR axis while simultaneously activating the adenosine monophosphate-activated protein kinase (AMPK) and sirtuin family proteins [51,55,56].
Acetyl-coenzyme A carboxylase 1 (ACC1) activity is inhibited by AMPK, which prevents the production of fatty acids. ACC1 induces vascular endothelial cell impairment leading to increase in disease severity in acute ischemic stroke patients [57,58]. It also increases plasma triglyceride levels, which will lead to an enlargement of atherosclerotic plaque and vascular occlusion, and finally increases disease severity [59]. This alteration in lipid metabolism is linked to an increase in T regulatory cells and a decrease in Th17 cells, which together ameliorate brain ischemia [60]. Th17 cells influence blood pressure by producing IL-17 and IL-22. IL-17 may have an impact on the sodium transport system, which includes sodium chloride cotransporter, epithelial sodium channels, and the sodium-hydrogen exchanger, in the renal proximal and distal tubular epithelial cells 3 [61]. On the other hand, IL-22 may affect the cyclooxygenase of the cells of the vascular wall and increase endothelial dysfunction, in turn increasing resistance in blood flow [62]. Additionally, mice lacking Ƴδ T cells were reported to be protected from endothelial damage and hypertension caused by angiotensin II [63]. Fat-related illness symptoms have been linked to decreased pro-inflammatory Th17 cell numbers in the gut and adipose tissue [64].
The individual role of B cells was poorly understood in hypertension. It might occur due to the activation of B cells that need co-stimulatory signals from T cells. However, anti CD 20 therapy [65] and Taylor et al. (2018) [66] have shown an association in preventing angiotensin II- related hypertension in mice. IgG was accumulated in the aortic adventitia during Angiotensin II- dependent hypertension in mice. It is regarded as a significant site of collagen and macrophage accumulation [67]. Endothelial cell nitric oxide synthase relaxes vessels during hypertension. Mice fed on high fat diet expresses Fcγ receptors on endothelial cells. IgG after associating with antigen can target these Fcγ receptors of endothelial cells thus have a negative effect of vasorelaxing activity [68]. These findings indicate a role for B lymphocytes in the endothelial dysfunction that primes vascular stiffening and elevated blood pressure as well as vascular remodeling.
It has been established that a ketogenic diet is linked to enhanced cardiac function, cardiomyocyte survival, and decreased cardiac fibrosis [69,70]. According to investigation, the activation of cardiac fibroblasts by ketone bodies was increased by the activation of transforming growth factor-β1 [71]. The differentiation of group 2 innate lymphoid cells and the T cell subset is regulated by the ketogenic diet [72]. Additionally, ketogenesis functions as a novel metabolic pathway in CD8+ Tmem cells, modifying these cells to facilitate the creation of memories through improved mitochondrial performance and substrate metabolism [73]. IL 33 has been found to reduce the symptoms of cardiac fibrosis [74]. Tao et al. (2021) examined the function underlying mechanism of Ketogenic diet in diabetic cardiomyopathy [75]. They found a decrease in the level of both CD4+ CD25+ Foxp3+ cells in blood and serum concentrations of IL-4 and IL-10 (Figure 2). Ketone bodies prevented naive CD4+ T cells from differentiating into Tregs. In the presence of ketone bodies, ST2L ligand synthesis, the proportion of ST2L+ cells in Tregs, and IL-33 production all decreased. The NLRP3 inflammasome regulates the release of the pro-inflammatory cytokines IL-1 and IL-18 and caspase-1 activation in macrophages [76]. It is a crucial innate immune sensor that may become active in response to atherosclerosis [77]. Therefore, understanding the endogenous mechanisms that regulate the NLRP3 inflammasome’s deactivation may help in the management of a number of chronic disorders. The ketogenic diet reduces inflammation, and these anti-inflammatory effects may be associated with BHB facilitated inhibition of the NLRP3 inflammasome [78]. The immunomodulatory effects of KD on various immune cells are summarized in Table 1.
5. Ketogenic Diet Mediated Immune Modulation in Central Nervous System Diseases
To sustain homeostasis, the human body comprising the encephalon amends its metabolism with respect to modified sustenance or disorder. This brings dynamic challanges for the energy metabolism of the diverse brain cell types. Many studies have shown significant role of KD in affecting the metabolous cross-talk between CNS cells as well as the periphery and the brain to manage neurological diseases.
Globally, more than 50 million people suffer from epilepsy [84]. Epilepsy is a chronic brain ailment categorized via recurring seizures: The short occurrences of uncontrolled movements capable of distressing either a portion or an entire body that may trigger unconsciousness or uncontrolled bladder or bowel function [85]. Cumulatively, the prognosis of epilepsy leads to suboptimal movement, deteriorated quality of health, and increased probability of death. Infections coupled with head injury, brain defects paired with prenatal or perinatal wounds, and brain tumors are the chief causes of epilepsy [86,87]. Approximately, 40% of epilepsy is triggered by genetic predisposition [88]. Several genes/variants have been correlated in many epileptic forms [89,90,91]. KD aims the downregulated adaptive and innate immunity existent in drug-resilient and refractory epileptic conditions [18]. Due to its extensive anti-inflammatory and neuroprotective activity, KD is being proven to be a safe and an efficient cure in patients with numerous pediatric and neurological disorders. A comprehensive antiepileptic approach of the KD is still indefinite [92].
The central nervous system (CNS), which is known for its privileged immune establishment, has been reported to exhibit immune and inflammatory responses [93]. Since the CNS tumors are vastly reliant on glucose, KD is being investigated as a potential immunomodulatory therapy for its treatment. Targeting the crucial components in immune pathways could facilitate the immunostimulatory approach against epilepsy [94]. KD is a rich source of ketone bodies, which constrain glutamate, in turn decreasing the biosynthesis of kynurenic acid, a metabolite responsible for epilepsy, in a kynurenine pathway (Figure 2). KD causes elevation of fatty acids, intonation of glycemia, and caloric restriction [95]. KD reduces astrocytic adenosine kinase (ADK) expression to increase the extracellular levels of adenosine [96]. Adenosine is an anti-inflammatory molecule that reduces the concentrations of TNF-α, IL-6, and CXCL2/3 and synchronizes an LPS-derived migration of the polymorphonuclear cells. Reduction of ADK downgrades inflammation in central and peripheral system [97]. It also elicits repressive adenosine A1 receptor (A1AR) [98]. Masino et al. (2012) reported a decrease in electrographic seizure activity while studying the stimulus of the adenosine in KD [99]. On other hand, mitochondrial respiration upregulates the levels of uncoupling proteins that decrease the ROS and increase resistance to seizures [100].
Many refractory epilepsy cases in children have reported the efficiency of KD in modifying the concentrations of serotonin as well as dopamine in the cerebrospinal fluid [101]. These reports have also indicated a substantial escalation in agmatine and γ-aminobutyric acid (GABA), exclusive of fluctuations in glutamate intensities [102]. KD works efficiently for children aged <3 years with West syndrome (WS) [103]. In WS, Patient experiences spasms and hypsarrhythmia [104]. Previous findings have reported 90% cognitive deficiency and 10–30% display signs of autistic spectrum disorder [105]. Sadly, many cases with WS were found to develop into a destructive epileptic brain disorder, Lennox-Gastaut syndrome (LGS) [106,107,108]. However, when hormonal therapy was merged with KD, treated children were less prone to developing LGS. This report predicted the efficacy of KD in causing obstructions in the extreme neuroinflammation in WS, consequently, terminating a precarious progression to LGS [109,110,111]. KD is being proven to successfully manage FIRES (febrile infection-related epilepsy syndrome) [112] and NORSE (new onset refractory status epilepticus) [109,113,114,115]. KD’s influence on neuroinflammation has also been found to be fruitful in treating myoclonic atonic epilepsy (MAE) with corticosteroids [116,117].
One of the CNS autoimmune inflammatory disorders is multiple sclerosis (MS). MS causes motor impediment and cerebral deficit. Pro-inflammatory eicosanoids are linked in the pathological process of MS due to their roles in enhancing vascular permeability and encouraging leukocyte migration activity across the brain [118,119,120]. A study showed the influence of KD on upregulation of anti-inflammatory ALOX15 and enzymes COX1, COX2, and ALOX5 eicosanoids in subjects with relapsing-remitting MS [121,122]. Kim et al. (2012) showed the favorable effect of KD correlated with decreased inflammation markers of brain in an experimental autoimmune encephalomyelitis (EAE) murine model [119]. KD could rescript crucial characteristics of MS pathogenesis through enriched CNS bioenergetics, and ketosis supported the expression of antioxidant pathways and lowered effector cell related protection [123]. KD is allied with a neuroprotective activity imparted by an enhanced sirtuin 1, (SIRT1- a participant of class III HDACs), expression in numerous cell types in the CNS [124,125,126]. KD is also reported to provide better neural consequences in a cuprizone-induced demyelination mouse model [127]. The downregulation of peroxisome proliferator- activated receptor γ (PPARγ) mRNA was observed in the monocytes of relapsing-remitting MS subjects [128]. PPARγ might overpower Th17 cell differentiation [129]. Initiation of PPARγ in different cell types of the brain leads to remyelination and shields demyelination as well as neuroinflammation [130,131].
A pilot study evaluated the safety and permissibility of a modified Atkins diet (KDMAD), a form of KD, in 20 patients with relapsing MS. This study presented anthropometrical enhancements on KDMAD, with a decrease in body mass index and total fat mass as well as low levels of proinflammatory adipokines at three months on KD intake [132]. Bahr et al. (2020) reported a decline in disease rigorousness, promising immunomodulatory effect of KD on 111 relapsing–remitting MS patients [133].
Spinal cord injury causes either short term or permanent damage to motor and sensory system [134,135,136]. Streijger et al. (2013) mentioned a full recovery of a spinal cord injury in 3rd week of treatment with KD which was initiated immediately after four hours of injury in mice [135]. In this case, inhibition of the NF-κB pathway followed by KD moderated a proinflammatory cytokines upsurge in the blood and spinal cord [137,138]. KD not only averts the diminution of superoxide dismutase produced by the spinal trauma, but also motivates a significant controller of the oxidative stress response known as nuclear factor-E2-related factor 2 [134,139,140].
Parkinson’s disease (PD) is a neurodegenerative ailment [141,142,143]. Clinically, microglial triggering and neuroinflammation may have a function in the succession of the disease [144,145]. Its symptoms comprise of bradykinesia, tremor in legs, hands, and head, stiffness in muscles, and slowness in movement [146,147]. Yang et al. (2010) reported dysregulated levels of IL-1 β, IL-6 [148], and TNF-α, alleviated motor dysfunction, as well as decreased stimuli of microglia and neuronic forfeiture in the SN upon treating with KD nourished MPTP injection [149,150,151]. Several studies provide information on microbiota-immune-brain axis [152]. Olson et al. (2018) described an acquired seizure protection associated with KD stimulated GABA upsurge in the hippocampus in murine models upon successful transplantation of the KD gut microbiome [153]. Sampson et al. (2020) exposed the association of the overexpressed α-synuclein of a PD murine model with neuroinflammation, cerebral deficit, and gut microbiome [154,155].
6. Keto Diet Complications:
Although KD is efficient in curing epilepsy, several studies have found frequent side effects like nausea, acidosis, and hypoglycemia etc. in patients [156]. However, these complications could be managed effortlessly by vigilant supervision of blood counts, hepatic enzymes, urinalysis, profiles depicting levels of minerals, vitamin, lipid, and serum carnitine [157]. Studies have also shown specific atypical and grave complications of KD, e.g., inflammation of pancreas, impairment or irritation and soreness to the small intestine, heart rhythm disorder, cardiomyopathy, and Parkinsonism. In these cases, immediate termination of the KD, proper examination and advance care should be applied to minimalize unfavorable consequences [158]. Patients on KD require special attention throughout surgical practices as various medical intervention procedures involve exposure of carbohydrates into the blood circulation. The epilepsy should be managed by excluding drugs such as propofol, a carbohydrate-comprising remedy to counteract post-surgical seizure incidents [159,160]. A disturbing effect of KDs on micronutrient levels was found in a case of a refractory epilepsy female patient who developed scurvy due to an insufficient level of vitamin C while on a KD [161,162,163].
Some antiseizure Medicines are also known to interact negatively with KD [159,164]. One of the examples is of Phenobarbital, which was reported to grow by 100% in patients on a KD. It was found to migrate to the blood–brain barrier, inducing distorted cerebral status at the usual prescription doses [165]. Thus, phenobarbital doses should be altered by observing the AED levels [165]. A prominently negative neural after-effect was reported by Erickson et al. (2003) [166] in an infant diagnosed with cryptogenic epileptic encephalopathy that evolved chorea and ataxia three weeks after starting a classical KD. One of the GI effects of the KD is cholelithiasis induced by metabolic changes turning into amplified gallbladder activity. Two such cases are mentioned, among which one required surgical attention [167]. Obese patients are more prone to these complications, having a high risk of dyslipidemia. The high fat intake in KD could have consequences of dyslipidemia and atherosclerosis. Kapetanakis et al. (2017) assessed 26 KD treated epileptic infants who exhibited declined carotid distensibility and deteriorated lipid profiles upon three months to a year post initiation of KD [168]. The same group evaluated the changes steadied by second year. Based on this evidence, patients with underlying dysregulations in their metabolism should be subject to very careful monitoring during KD.
7. Conclusions
Overall, several preclinical studies show or indicate that the KD modifies the immune system to combat different disease conditions by impacting the immune microenvironment of the host. Furthermore, molecular and mechanistic studies are required to establish the concrete role of ketogenic diets in immune alterations.
Author Contributions
Conceptualization, V.K. and S.K.S.; writing—original draft preparation, S.S., V.A.P., A.T., K.P.S., V.K. and S.K.S.; writing—review and editing, S.S., V.A.P., A.T., K.P.S., V.K. and S.K.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Zhu, H.; Bi, D.; Zhang, Y.; Kong, C.; Du, J.; Wu, X.; Wei, Q.; Qin, H. Ketogenic diet for human diseases: The underlying mechanisms and potential for clinical implementations. Signal Transduct. Target. Ther. 2022, 7, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Joanne Carroll, M.R.D.K. The Ketogenic Diet: A Practical Guide for Caregivers. J. Am. Diet. Assoc. 1998, 98, 316–321. [Google Scholar] [CrossRef] [PubMed]
- Pardo, A.C. Ketogenic Diet: A Role in Immunity? Pediatr. Neurol. Briefs 2020, 34, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dowis, K.; Banga, S. The Potential Health Benefits of the Ketogenic Diet: A Narrative Review. Nutrients 2021, 13, 1654. [Google Scholar] [CrossRef] [PubMed]
- Kakodkar, P.; Arnold, A.; Ali, A.; Kaka, N. Ketogenic Diet: Biochemistry, Weight Loss and Clinical Applications. Nutr. Food Sci. Int. J. 2020, 10, 1–9. [Google Scholar] [CrossRef]
- Ludwig, D.S. The Ketogenic Diet: Evidence for Optimism but High-Quality Research Needed. J. Nutr. 2019, 150, 1354–1359. [Google Scholar] [CrossRef] [Green Version]
- Westman, E.C.; Mavropoulos, J.; Yancy, W.S.; Volek, J.S. A review of low-carbohydrate ketogenic diets. Curr. Atheroscler. Rep. 2003, 5, 476–483. [Google Scholar] [CrossRef]
- Ciaffi, J.; Mitselman, D.; Mancarella, L.; Brusi, V.; Lisi, L.; Ruscitti, P.; Cipriani, P.; Meliconi, R.; Giacomelli, R.; Borghi, C.; et al. The Effect of Ketogenic Diet on Inflammatory Arthritis and Cardiovascular Health in Rheumatic Conditions: A Mini Review. Front. Med. 2021, 8. [Google Scholar] [CrossRef]
- Castaldo, G.; Pagano, I.; Grimaldi, M.; Marino, C.; Molettieri, P.; Santoro, A.; Stillitano, I.; Romano, R.; Montoro, P.; D’Ursi, A.M.; et al. Effect of Very-Low-Calorie Ketogenic Diet on Psoriasis Patients: A Nuclear Magnetic Resonance-Based Metabolomic Study. J. Proteome Res. 2020, 20, 1509–1521. [Google Scholar] [CrossRef]
- Kong, C.; Yan, X.; Liu, Y.; Huang, L.; Zhu, Y.; He, J.; Gao, R.; Kalady, M.F.; Goel, A.; Qin, H.; et al. Ketogenic diet alleviates colitis by reduction of colonic group 3 innate lymphoid cells through altering gut microbiome. Signal Transduct. Target. Ther. 2021, 6, 1–12. [Google Scholar] [CrossRef]
- Charlot, A.; Boumiza, R.; Roux, M.; Zoll, J. Obésité, inflammation et COVID-19: Intérêt préventif de l’alimentation cétogène? Biol. Aujourd’hui 2021, 215, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Paoli, A.; Gorini, S.; Caprio, M. The dark side of the spoon—glucose, ketones and COVID-19: A possible role for ketogenic diet? J. Transl. Med. 2020, 18, 441. [Google Scholar] [CrossRef] [PubMed]
- Gangitano, E.; Tozzi, R.; Gandini, O.; Watanabe, M.; Basciani, S.; Mariani, S.; Lenzi, A.; Gnessi, L.; Lubrano, C. Ketogenic Diet as a Preventive and Supportive Care for COVID-19 Patients. Nutrients 2021, 13, 1004. [Google Scholar] [CrossRef] [PubMed]
- Soliman, S.; Faris, M.E.; Ratemi, Z.; Halwani, R. Switching Host Metabolism as an Approach to Dampen SARS-CoV-2 Infection. Ann. Nutr. Metab. 2020, 76, 297–303. [Google Scholar] [CrossRef] [PubMed]
- Sukkar, S.G.; Cogorno, L.; Pisciotta, L.; Pasta, A.; Vena, A.; Gradaschi, R.; Dentone, C.; Guiddo, E.; Martino, E.; Beltramini, S.; et al. Clinical efficacy of eucaloric ketogenic nutrition in the COVID-19 cytokine storm: A retrospective analysis of mortality and intensive care unit admission. Nutrition 2021, 89, 111236. [Google Scholar] [CrossRef] [PubMed]
- Karagiannis, F.; Peukert, K.; Surace, L.; Michla, M.; Nikolka, F.; Fox, M.; Weiss, P.; Feuerborn, C.; Maier, P.; Schulz, S.; et al. Impaired ketogenesis ties metabolism to T cell dysfunction in COVID-19. Nature 2022, 609, 801–807. [Google Scholar] [CrossRef]
- Ryu, S.; Shchukina, I.; Youm, Y.-H.; Qing, H.; Hilliard, B.; Dlugos, T.; Zhang, X.; Yasumoto, Y.; Booth, C.J.; Fernández-Hernando, C.; et al. Ketogenic diet restrains aging-induced exacerbation of coronavirus infection in mice. Elife 2021, 10. [Google Scholar] [CrossRef]
- Goldberg, E.L.; Molony, R.D.; Kudo, E.; Sidorov, S.; Kong, Y.; Dixit, V.D.; Iwasaki, A. Ketogenic diet activates protective γδ T cell responses against influenza virus infection. Sci. Immunol. 2019, 4. [Google Scholar] [CrossRef]
- Bradshaw, P.C.; Seeds, W.A.; Miller, A.C.; Mahajan, V.R.; Curtis, W.M. COVID-19: Proposing a Ketone-Based Metabolic Therapy as a Treatment to Blunt the Cytokine Storm. Oxidative Med. Cell. Longev. 2020, 2020, 6401341. [Google Scholar] [CrossRef]
- Hirschberger, S.; Gellert, L.; Effinger, D.; Muenchhoff, M.; Herrmann, M.; Briegel, J.-M.; Zwißler, B.; Kreth, S. Ketone Bodies Improve Human CD8+ Cytotoxic T-Cell Immune Response During COVID-19 Infection. Front. Med. 2022, 9. [Google Scholar] [CrossRef]
- Klement, R.J. Fasting, Fats, and Physics: Combining Ketogenic and Radiation Therapy against Cancer. Complement. Med. Res. 2017, 25, 102–113. [Google Scholar] [CrossRef]
- Rous, P. The influence of diet on transplanted and spontaneous mouse tumors. J. Exp. Med. 1914, 20, 433–451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tisdale, M.; Brennan, R.; Fearon, K. Reduction of weight loss and tumour size in a cachexia model by a high fat diet. Br. J. Cancer 1987, 56, 39–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klement, R.J.; Champ, C.E.; Otto, C.; Kämmerer, U. Anti-Tumor Effects of Ketogenic Diets in Mice: A Meta-Analysis. PLOS ONE 2016, 11, e0155050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrere, G.; Alou, M.T.; Liu, P.; Goubet, A.-G.; Fidelle, M.; Kepp, O.; Durand, S.; Iebba, V.; Fluckiger, A.; Daillère, R.; et al. Ketogenic diet and ketone bodies enhance the anticancer effects of PD-1 blockade. J. Clin. Investig. 2021, 6. [Google Scholar] [CrossRef] [PubMed]
- Hao, G.-W.; Chen, Y.-S.; He, D.-M.; Wang, H.-Y.; Wu, G.-H.; Zhang, B. Growth of Human Colon Cancer Cells in Nude Mice is Delayed by Ketogenic Diet With or Without Omega-3 Fatty Acids and Medium-chain Triglycerides. Asian Pac. J. Cancer Prev. 2015, 16, 2061–2068. [Google Scholar] [CrossRef] [Green Version]
- Nebeling, L.C.; Miraldi, F.; Shurin, S.B.; Lerner, E. Effects of a ketogenic diet on tumor metabolism and nutritional status in pediatric oncology patients: Two case reports. J. Am. Coll. Nutr. 1995, 14, 202–208. [Google Scholar] [CrossRef]
- Seyfried, T.N.; Shelton, L.M. Cancer as a metabolic disease. Nutr. Metab. 2010, 7, 7. [Google Scholar] [CrossRef] [Green Version]
- Abdelwahab, M.G.; Fenton, K.E.; Preul, M.C.; Rho, J.M.; Lynch, A.; Stafford, P.; Scheck, A.C. The Ketogenic Diet Is an Effective Adjuvant to Radiation Therapy for the Treatment of Malignant Glioma. PLoS ONE 2012, 7, e36197. [Google Scholar] [CrossRef]
- Sharpe, A.; Pauken, K. The diverse functions of the PD1 inhibitory pathway. Nat. Rev. Immunol. 2018, 18, 153–167. [Google Scholar] [CrossRef]
- Dai, X.; Bu, X.; Gao, Y.; Guo, J.; Hu, J.; Jiang, C.; Zhang, Z.; Xu, K.; Duan, J.; He, S.; et al. Energy status dictates PD-L1 protein abundance and anti-tumor immunity to enable checkpoint blockade. Mol. Cell 2021, 81, 2317–2331.e6. [Google Scholar] [CrossRef]
- Mauldin, I.S.; Wages, N.A.; Stowman, A.M.; Wang, E.; Smolkin, M.E.; Olson, W.C.; Deacon, D.H.; Smith, K.T.; Galeassi, N.V.; Chianese-Bullock, K.A.; et al. Intratumoral interferon-gamma increases chemokine production but fails to increase T cell infiltration of human melanoma metastases. Cancer Immunol. Immunother. 2016, 65, 1189–1199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, W.; Yang, J.; Liu, B.; Liu, Q.; Wang, T.; Wang, Q.; Liu, M.; Li, L.; Wang, Z.; Li, S.; et al. Ketogenic diet inhibits tumor growth by enhancing immune response, attenuating immunosuppression, inhibiting angiogenesis and EMT in CT26 colon tumor allografts mouse model. J. Funct. Foods 2022, 92, 105067. [Google Scholar] [CrossRef]
- Meade, T. Hypercoagulability and ischaemic heart disease. Blood Rev. 1987, 1, 2–8. [Google Scholar] [CrossRef] [PubMed]
- Lussier, D.M.; Woolf, E.C.; Johnson, J.L.; Brooks, K.S.; Blattman, J.N.; Scheck, A.C. Enhanced immunity in a mouse model of malignant glioma is mediated by a therapeutic ketogenic diet. BMC Cancer 2016, 16, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Sun, H.Y.; Sebaratnam, D.F. Prurigo pigmentosa following a ketogenic diet: A case report. Eur. J. Clin. Nutr. 2021, 76, 624–625. [Google Scholar] [CrossRef]
- Talib, W.H.; Mahmod, A.I.; Kamal, A.; Rashid, H.M.; Alashqar, A.; Khater, S.; Jamal, D.; Waly, M. Ketogenic Diet in Cancer Prevention and Therapy: Molecular Targets and Therapeutic Opportunities. Curr. Issues Mol. Biol. 2021, 43, 558–589. [Google Scholar] [CrossRef]
- Meynet, O.; Ricci, J.-E. Caloric restriction and cancer: Molecular mechanisms and clinical implications. Trends Mol. Med. 2014, 20, 419–427. [Google Scholar] [CrossRef]
- Hursting, S.D.; Ford, N.A.; Dunlap, S.M.; Hursting, M.J.; Lashinger, L.M. Calorie Restriction and Cancer Prevention: Established and Emerging Mechanisms. In Obesity, Inflammation and Cancer; Springer: New York, NY, USA, 2013; pp. 363–379. [Google Scholar] [CrossRef]
- Mulrooney, T.J.; Marsh, J.; Urits, I.; Seyfried, T.; Mukherjee, P. Influence of Caloric Restriction on Constitutive Expression of NF-κB in an Experimental Mouse Astrocytoma. PLoS ONE 2011, 6, e18085. [Google Scholar] [CrossRef]
- Zhang, N.; Liu, C.; Zhang, R.; Jin, L.; Yin, X.; Zheng, X.; Siebert, H.-C.; Li, Y.; Wang, Z.; Loers, G.; et al. Amelioration of clinical course and demyelination in the cuprizone mouse model in relation to ketogenic diet. Food Funct. 2020, 11, 5647–5663. [Google Scholar] [CrossRef]
- Murray, P.J.; Wynn, T.A. Protective and pathogenic functions of macrophage subsets. Nat. Rev. Immunol. 2011, 11, 723–737. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.-W.; Hua, T.N. Impact of Lifestyle Behaviors on Cancer Risk and Prevention. J. Lifestyle Med. 2021, 11, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Zou, Y.; Fineberg, S.; Pearlman, A.; Feinman, R.D.; Fine, E.J. The effect of a ketogenic diet and synergy with rapamycin in a mouse model of breast cancer. PLOS ONE 2020, 15, e0233662. [Google Scholar] [CrossRef] [PubMed]
- Rosenzweig, R.; Kumar, V.; Gupta, S.; Bermeo-Blanco, O.; Stratton, M.S.; Gumina, R.J.; Bansal, S.S. Estrogen Receptor-β Agonists Modulate T-Lymphocyte Activation and Ameliorate Left Ventricular Remodeling During Chronic Heart Failure. Circ. Hear. Fail. 2022, 15. [Google Scholar] [CrossRef] [PubMed]
- Rosenzweig, R.; Gupta, S.; Kumar, V.; Gumina, R.J.; Bansal, S.S. Estrogenic bias in T-Lymphocyte biology: Implications for cardiovascular disease. Pharmacol. Res. 2021, 170, 105606. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Prabhu, S.D.; Bansal, S.S. CD4+ T-lymphocytes exhibit biphasic kinetics post-myocardial infarction. Front. Cardiovasc. Med. 2022, 9. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Rosenzweig, R.; Asalla, S.; Nehra, S.; Prabhu, S.D.; Bansal, S.S. TNFR1 Contributes to Activation-Induced Cell Death of Pathological CD4+ T Lymphocytes During Ischemic Heart Failure. JACC: Basic Transl. Sci. 2022, 7, 1038–1049. [Google Scholar] [CrossRef]
- Visioli, F.; Mucignat-Caretta, C.; Anile, F.; Panaite, S.-A. Traditional and Medical Applications of Fasting. Nutrients 2022, 14, 433. [Google Scholar] [CrossRef]
- Kirkpatrick, C.F.; Bolick, J.P.; Kris-Etherton, P.M.; Sikand, G.; Aspry, K.E.; Soffer, D.E.; Willard, K.-E.; Maki, K.C. Review of current evidence and clinical recommendations on the effects of low-carbohydrate and very-low-carbohydrate (including ketogenic) diets for the management of body weight and other cardiometabolic risk factors: A scientific statement from the National Lipid Association Nutrition and Lifestyle Task Force. J. Clin. Lipidol. 2019, 13, 689–711.e1. [Google Scholar] [CrossRef]
- Guarente, L. Calorie restriction and sirtuins revisited. Genes Dev. 2013, 27, 2072–2085. [Google Scholar] [CrossRef] [Green Version]
- Imai, S.-I.; Guarente, L. It takes two to tango: NAD+ and sirtuins in aging/longevity control. npj Aging Mech. Dis. 2016, 2, 16017. [Google Scholar] [CrossRef] [Green Version]
- De Toledo, F.W.; Grundler, F.; Sirtori, C.R.; Ruscica, M. Unravelling the health effects of fasting: A long road from obesity treatment to healthy life span increase and improved cognition. Ann. Med. 2020, 52, 147–161. [Google Scholar] [CrossRef] [PubMed]
- Osataphan, S.; Macchi, C.; Singhal, G.; Chimene-Weiss, J.; Sales, V.; Kozuka, C.; Dreyfuss, J.M.; Pan, H.; Tangcharoenpaisan, Y.; Morningstar, J.; et al. SGLT2 inhibition reprograms systemic metabolism via FGF21-dependent and -independent mechanisms. J. Clin. Investig. 2019, 4, e123130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cantó, C.; Auwerx, J. Calorie Restriction: Is AMPK a Key Sensor and Effector? Physiology 2011, 26, 214–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mercken, E.M.; Crosby, S.D.; Lamming, D.W.; JeBailey, L.; Krzysik-Walker, S.; Villareal, D.T.; Capri, M.; Franceschi, C.; Zhang, Y.; Becker, K.; et al. Calorie restriction in humans inhibits the PI 3 K / AKT pathway and induces a younger transcription profile. Aging Cell 2013, 12, 645–651. [Google Scholar] [CrossRef] [Green Version]
- Glatzel, D.K.; Koeberle, A.; Pein, H.; Loeser, K.; Stark, A.; Keksel, N.; Werz, O.; Mueller, R.; Bischoff, I.; Fürst, R. Acetyl-CoA carboxylase 1 regulates endothelial cell migration by shifting the phospholipid composition. J. Lipid Res. 2018, 59, 298–311. [Google Scholar] [CrossRef] [Green Version]
- Niu, C.; Chen, Z.; Kim, K.T.; Sun, J.; Xue, M.; Chen, G.; Li, S.; Shen, Y.; Zhu, Z.; Wang, X.; et al. Metformin alleviates hyperglycemia-induced endothelial impairment by downregulating autophagy via the Hedgehog pathway. Autophagy 2019, 15, 843–870. [Google Scholar] [CrossRef]
- Chen, L.; Duan, Y.; Wei, H.; Ning, H.; Bi, C.; Zhao, Y.; Qin, Y.; Li, Y. Acetyl-CoA carboxylase (ACC) as a therapeutic target for metabolic syndrome and recent developments in ACC1/2 inhibitors. Expert Opin. Investig. Drugs 2019, 28, 917–930. [Google Scholar] [CrossRef]
- Wang, X.; Zhou, Y.; Tang, D.; Zhu, Z.; Li, Y.; Huang, T.; Müller, R.; Yu, W.; Li, P. ACC1 (Acetyl Coenzyme A Carboxylase 1) Is a Potential Immune Modulatory Target of Cerebral Ischemic Stroke. Stroke 2019, 50, 1869–1878. [Google Scholar] [CrossRef]
- Norlander, A.E.; Saleh, M.A.; Kamat, N.V.; Ko, B.; Gnecco, J.; Zhu, L.; Dale, B.L.; Iwakura, Y.; Hoover, R.S.; McDonough, A.A.; et al. Interleukin-17A Regulates Renal Sodium Transporters and Renal Injury in Angiotensin II–Induced Hypertension. Hypertension 2016, 68, 167–174. [Google Scholar] [CrossRef] [Green Version]
- Ye, J.; Ji, Q.; Liu, J.; Liu, L.; Huang, Y.; Shi, Y.; Shi, L.; Wang, M.; Liu, M.; Feng, Y.; et al. Interleukin 22 Promotes Blood Pressure Elevation and Endothelial Dysfunction in Angiotensin II–Treated Mice. J. Am. Hear. Assoc. 2017, 6. [Google Scholar] [CrossRef] [Green Version]
- Caillon, A.; Mian, M.O.R.; Fraulob-Aquino, J.C.; Huo, K.-G.; Barhoumi, T.; Ouerd, S.; Sinnaeve, P.R.; Paradis, P.; Schiffrin, E.L. γδ T Cells Mediate Angiotensin II-Induced Hypertension and Vascular Injury. Circulation 2017, 135, 2155–2162. [Google Scholar] [CrossRef] [PubMed]
- Kiran, S.; Rakib, A.; Kodidela, S.; Kumar, S.; Singh, U.P. High-Fat Diet-Induced Dysregulation of Immune Cells Correlates with Macrophage Phenotypes and Chronic Inflammation in Adipose Tissue. Cells 2022, 11, 1327. [Google Scholar] [CrossRef] [PubMed]
- Mathis, K.W.; Wallace, K.; Flynn, E.R.; Maric-Bilkan, C.; Lamarca, B.; Ryan, M.J. Preventing Autoimmunity Protects Against the Development of Hypertension and Renal Injury. Hypertension 2014, 64, 792–800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, E.B.; Barati, M.T.; Powell, D.W.; Turbeville, H.R.; Ryan, M.J. Plasma Cell Depletion Attenuates Hypertension in an Experimental Model of Autoimmune Disease. Hypertension 2018, 71, 719–728. [Google Scholar] [CrossRef]
- Chan, C.T.; Sobey, C.G.; Lieu, M.; Ferens, D.; Kett, M.M.; Diep, H.; Kim, H.A.; Krishnan, S.M.; Lewis, C.; Salimova, E.; et al. Obligatory Role for B Cells in the Development of Angiotensin II–Dependent Hypertension. Hypertension 2015, 66, 1023–1033. [Google Scholar] [CrossRef]
- Sundgren, N.C.; Vongpatanasin, W.; Boggan, B.-M.D.; Tanigaki, K.; Yuhanna, I.S.; Chambliss, K.L.; Mineo, C.; Shaul, P.W. IgG Receptor FcγRIIB Plays a Key Role in Obesity-Induced Hypertension. Hypertension 2015, 65, 456–462. [Google Scholar] [CrossRef]
- Guo, Y.; Zhang, C.; Shang, F.-F.; Luo, M.; You, Y.; Zhai, Q.; Xia, Y.; Suxin, L. Ketogenic Diet Ameliorates Cardiac Dysfunction via Balancing Mitochondrial Dynamics and Inhibiting Apoptosis in Type 2 Diabetic Mice. Aging Dis. 2020, 11, 229–240. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.; Wang, F.; Wang, J.; Zhang, D.; Zhao, X. Ketogenic diet attenuates aging-associated myocardial remodeling and dysfunction in mice. Exp. Gerontol. 2020, 140, 111058. [Google Scholar] [CrossRef]
- Ma, Y.; Zou, H.; Zhu, X.-X.; Pang, J.; Xu, Q.; Jin, Q.-Y.; Ding, Y.-H.; Zhou, B.; Huang, D.-S. Transforming growth factor β: A potential biomarker and therapeutic target of ventricular remodeling. Oncotarget 2017, 8, 53780–53790. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Tang, K.; Ma, J.; Zhou, L.; Liu, J.; Zeng, L.; Zhu, L.; Xu, P.; Chen, J.; Wei, K.; et al. Ketogenesis-generated β-hydroxybutyrate is an epigenetic regulator of CD8+ T-cell memory development. Nature 2019, 22, 18–25. [Google Scholar] [CrossRef]
- Mey, J.T.; Erickson, M.L.; Axelrod, C.L.; King, W.T.; A Flask, C.; McCullough, A.J.; Kirwan, J.P. β-Hydroxybutyrate is reduced in humans with obesity-related NAFLD and displays a dose-dependent effect on skeletal muscle mitochondrial respiration in vitro. Am. J. Physiol. Metab. 2020, 319, E187–E195. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Wang, S.; Xie, Y.; Chen, Z.; Guo, Q.; Yuan, W.; Guan, C.; Xu, C.; Huang, Y.; Wang, J.; et al. Interleukin-33 alleviates diabetic cardiomyopathy through regulation of endoplasmic reticulum stress and autophagy via insulin-like growth factor-binding protein 3. J. Cell. Physiol. 2020, 236, 4403–4419. [Google Scholar] [CrossRef] [PubMed]
- Tao, J.; Chen, H.; Wang, Y.-J.; Qiu, J.-X.; Meng, Q.-Q.; Zou, R.-J.; Li, L.; Huang, J.-G.; Zhao, Z.-K.; Huang, Y.-L.; et al. Ketogenic Diet Suppressed T-Regulatory Cells and Promoted Cardiac Fibrosis via Reducing Mitochondria-Associated Membranes and Inhibiting Mitochondrial Function. Oxidative Med. Cell. Longev. 2021, 2021, 5512322. [Google Scholar] [CrossRef]
- Franchi, L.; Eigenbrod, T.; Muñoz-Planillo, R.; Nuñez, G. The inflammasome: A caspase-1-activation platform that regulates immune responses and disease pathogenesis. Nat. Immunol. 2009, 10, 241–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duewell, P.; Kono, H.; Rayner, K.J.; Sirois, C.M.; Vladimer, G.; Bauernfeind, F.G.; Abela, G.S.; Franchi, L.; Nuñez, G.; Schnurr, M.; et al. NLRP3 inflammasomes are required for atherogenesis and activated by cholesterol crystals. Nature 2010, 464, 1357–1361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Youm, Y.-H.; Nguyen, K.Y.; Grant, R.W.; Goldberg, E.L.; Bodogai, M.; Kim, D.; D’Agostino, D.; Planavsky, N.; Lupfer, C.; Kanneganti, T.-D.; et al. The ketone metabolite β-hydroxybutyrate blocks NLRP3 inflammasome–mediated inflammatory disease. Nat. Med. 2015, 21, 263–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klement, R.J.; Meyer, D.; Kanzler, S.; Sweeney, R.A. Ketogenic diets consumed during radio-chemotherapy have beneficial effects on quality of life and metabolic health in patients with rectal cancer. Eur. J. Nutr. 2021, 61, 69–84. [Google Scholar] [CrossRef]
- Sukkar, S.G.; Bassetti, M. Induction of ketosis as a potential therapeutic option to limit hyperglycemia and prevent cytokine storm in COVID-19. Nutrition 2020, 79-80, 110967. [Google Scholar] [CrossRef]
- Crujeiras, A.B.; Izquierdo, A.G.; Primo, D.; Milagro, F.I.; Sajoux, I.; Jácome, A.; Fernandez-Quintela, A.; Portillo, M.P.; Martínez, J.; Martinez-Olmos, M.A.; et al. Epigenetic landscape in blood leukocytes following ketosis and weight loss induced by a very low calorie ketogenic diet (VLCKD) in patients with obesity. Clin. Nutr. 2021, 40, 3959–3972. [Google Scholar] [CrossRef]
- Schreck, K.C.; Lwin, M.; Strowd, R.E.; Henry-Barron, B.J.; Blakeley, J.O.; Cervenka, M.C. Effect of ketogenic diets on leukocyte counts in patients with epilepsy. Nutr. Neurosci. 2017, 22, 522–527. [Google Scholar] [CrossRef]
- Wilhelm, C.; Surendar, J.; Karagiannis, F. Enemy or ally? Fasting as an essential regulator of immune responses. Trends Immunol. 2021, 42, 389–400. [Google Scholar] [CrossRef] [PubMed]
- Beghi, E.; Giussani, G.; Nichols, E.; Abd-Allah, F.; Abdela, J.; Abdelalim, A.; Abraha, H.N.; Adib, M.G.; Agrawal, S.; Alahdab, F.; et al. Global, regional, and national burden of epilepsy, 1990–2016: A systematic analysis for the Global Burden of Disease Study 2016. Lancet Neurol. 2019, 18, 357–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leeman, B.; Cole, A. Advancements in the Treatment of Epilepsy. Annu. Rev. Med. 2008, 59, 503–523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scher, M.S.; Kidder, B.M.; Bangert, B.A. Pediatric Epilepsy Evaluations From the Prenatal Perspective. J. Child Neurol. 2007, 22, 396–401. [Google Scholar] [CrossRef] [PubMed]
- Razaz, N.; Tomson, T.; Wikström, A.-K.; Cnattingius, S. Association Between Pregnancy and Perinatal Outcomes Among Women With Epilepsy. JAMA Neurol. 2017, 74, 983–991. [Google Scholar] [CrossRef] [PubMed]
- Perucca, P.; Bahlo, M.; Berkovic, S.F. The Genetics of Epilepsy. Annu. Rev. Genom. Hum. Genet. 2020, 21, 205–230. [Google Scholar] [CrossRef] [PubMed]
- Leu, C.; Balestrini, S.; Maher, B.; Hernández-Hernández, L.; Gormley, P.; Hämäläinen, E.; Heggeli, K.; Schoeler, N.; Novy, J.; Willis, J.; et al. Genome-wide Polygenic Burden of Rare Deleterious Variants in Sudden Unexpected Death in Epilepsy. eBioMedicine 2015, 2, 1063–1070. [Google Scholar] [CrossRef] [Green Version]
- Guerrini, R.; Balestrini, S.; Wirrell, E.C.; Walker, M.C. Monogenic Epilepsies. Neurology 2021, 97, 817–831. [Google Scholar] [CrossRef]
- Wang, J.; Lin, Z.-J.; Liu, L.; Xu, H.-Q.; Shi, Y.-W.; Yi, Y.-H.; He, N.; Liao, W.-P. Epilepsy-associated genes. Seizure 2016, 44, 11–20. [Google Scholar] [CrossRef] [Green Version]
- Koh, S.; Dupuis, N.; Auvin, S. Ketogenic diet and Neuroinflammation. Epilepsy Res. 2020, 167, 106454. [Google Scholar] [CrossRef]
- Panhans, C.M.; Gresham, G.; Amaral, L.J.; Hu, J. Exploring the Feasibility and Effects of a Ketogenic Diet in Patients With CNS Malignancies: A Retrospective Case Series. Front. Neurosci. 2020, 14, 390. [Google Scholar] [CrossRef] [PubMed]
- Lorenzo, P.M.; Sajoux, I.; Izquierdo, A.G.; Gomez-Arbelaez, D.; Zulet, M.A.; Abete, I.; Castro, A.I.; Baltar, J.; Portillo, M.P.; Tinahones, F.J.; et al. Immunomodulatory effect of a very-low-calorie ketogenic diet compared with bariatric surgery and a low-calorie diet in patients with excessive body weight. Clin. Nutr. 2022, 41, 1566–1577. [Google Scholar] [CrossRef] [PubMed]
- Okawa, T.; Nagai, M.; Hase, K. Dietary Intervention Impacts Immune Cell Functions and Dynamics by Inducing Metabolic Rewiring. Front. Immunol. 2021, 11. [Google Scholar] [CrossRef] [PubMed]
- Brunner, B.; Ari, C.; D’Agostino, D.P.; Kovács, Z. Adenosine Receptors Modulate the Exogenous Ketogenic Supplement-Evoked Alleviating Effect on Lipopolysaccharide-Generated Increase in Absence Epileptic Activity in WAG/Rij Rats. Nutrients 2021, 13, 4082. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; He, X.; Luan, G.; Li, T. Role of DNA Methylation and Adenosine in Ketogenic Diet for Pharmacoresistant Epilepsy: Focus on Epileptogenesis and Associated Comorbidities. Front. Neurol. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Kovacs, Z.; Brunner, B.; D’Agostino, D.P.; Ari, C. Inhibition of adenosine A1 receptors abolished the nutritional ketosis-evoked delay in the onset of isoflurane-induced anesthesia in Wistar Albino Glaxo Rijswijk rats. BMC Anesthesiol. 2020, 20, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Masino, S.A.; Rho, J.M. Mechanisms of Ketogenic Diet Action. In Jasper’s Basic Mechanisms of the Epilepsies; National Center for Biotechnology Information (US): Bethesda, MD, USA, 2012. [Google Scholar]
- Watanabe, M.; Tozzi, R.; Risi, R.; Tuccinardi, D.; Mariani, S.; Basciani, S.; Spera, G.; Lubrano, C.; Gnessi, L. Beneficial effects of the ketogenic diet on nonalcoholic fatty liver disease: A comprehensive review of the literature. Obes. Rev. 2020, 21, e13024. [Google Scholar] [CrossRef] [Green Version]
- Żarnowska, I.; Wróbel-Dudzińska, D.; Tulidowicz-Bielak, M.; Kocki, T.; Mitosek-Szewczyk, K.; Gasior, M.; Turski, W.A. Changes in tryptophan and kynurenine pathway metabolites in the blood of children treated with ketogenic diet for refractory epilepsy. Seizure 2019, 69, 265–272. [Google Scholar] [CrossRef]
- Hirschberger, S.; Strauß, G.; Effinger, D.; Marstaller, X.; Ferstl, A.; Müller, M.B.; Wu, T.; Hübner, M.; Rahmel, T.; Mascolo, H.; et al. Very-low-carbohydrate diet enhances human T-cell immunity through immunometabolic reprogramming. EMBO Mol. Med. 2021, 13, e14323. [Google Scholar] [CrossRef]
- Dressler, A.; Benninger, F.; Trimmel-Schwahofer, P.; Gröppel, G.; Porsche, B.; Abraham, K.; Mühlebner, A.; Samueli, S.; Male, C.; Feucht, M. Efficacy and tolerability of the ketogenic diet versus high-dose adrenocorticotropic hormone for infantile spasms: A single-center parallel-cohort randomized controlled trial. Epilepsia 2019, 60, 441–451. [Google Scholar] [CrossRef]
- Olmi, S.; Petkoski, S.; Guye, M.; Bartolomei, F.; Jirsa, V. Controlling seizure propagation in large-scale brain networks. PLoS Comput. Biol. 2019, 15, e1006805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruskin, D.N.; Murphy, M.I.; Slade, S.L.; Masino, S.A. Ketogenic diet improves behaviors in a maternal immune activation model of autism spectrum disorder. PLOS ONE 2017, 12, e0171643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asadi-Pooya, A.A.; Katibeh, P. Lennox-Gastaut Syndrome. In Comprehensive Pharmacology; Elsevier: Amsterdam, The Netherlands, 2021; pp. 105–122. [Google Scholar] [CrossRef]
- Gokten, E.S.; Duman, N. West syndrome and autism: A case report. Eur. Res. J. 2015, 1, 154–156. [Google Scholar] [CrossRef] [Green Version]
- De Alba, G.O.-G.; Valdez, J.M.; Crespo, F.V. West Syndrome Evolving into the Lennox-Gastaut Syndrome. Clin. Electroencephalogr. 1984, 15, 61–68. [Google Scholar] [CrossRef]
- Calvo, A.; Buompadre, M.C.; Gallo, A.; Gutiérrez, R.; Valenzuela, G.R.; Caraballo, R. Electroclinical pattern in the transition from West to Lennox-Gastaut syndrome. Epilepsy Res. 2020, 167, 106446. [Google Scholar] [CrossRef]
- Amador, J.B.; García-Segovia, R.; Boyano, M.G.; Miño-León, G. West Syndrome in Children With Congenital Zika Virus Infection. Pediatr. Infect. Dis. J. 2021, 40, 1108–1110. [Google Scholar] [CrossRef]
- Pavone, P.; Polizzi, A.; Marino, S.D.; Corsello, G.; Falsaperla, R.; Marino, S.; Ruggieri, M. West syndrome: A comprehensive review. Neurol. Sci. 2020, 41, 3547–3562. [Google Scholar] [CrossRef]
- Kramer, U.; Chi, C.-S.; Lin, K.-L.; Specchio, N.; Sahin, M.; Olson, H.; Nabbout, R.; Kluger, G.; Lin, J.-J.; Van Baalen, A. Febrile infection-related epilepsy syndrome (FIRES): Pathogenesis, treatment, and outcome. Epilepsia 2011, 52, 1956–1965. [Google Scholar] [CrossRef]
- Peng, P.; Peng, J.; Yin, F.; Deng, X.; Chen, C.; He, F.; Wang, X.; Guang, S.; Mao, L. Ketogenic Diet as a Treatment for Super-Refractory Status Epilepticus in Febrile Infection-Related Epilepsy Syndrome. Front. Neurol. 2019, 10, 423. [Google Scholar] [CrossRef] [Green Version]
- Obara, K.; Ono, T. Ketogenic Diet for a Young Adult Patient With Chronic-Phase Febrile Infection-Related Epilepsy Syndrome. Cureus 2022, 14, e22099. [Google Scholar] [CrossRef] [PubMed]
- Li, W.-J.; Xue, C.-L.; Zhang, Y.; Wu, L.-H.; Chen, D.-M.; Chen, F.; Xu, J.; Li, Z.; Miao, H.-J. Ketogenic diet (KD) therapy in the acute phase of febrile infection-related epilepsy syndrome (FIRES): A case report. Transl. Pediatr. 2021, 10, 2392–2397. [Google Scholar] [CrossRef] [PubMed]
- Mehta, V.; Ferrie, C.D.; Cross, H.; Vadlamani, G. Corticosteroids including ACTH for childhood epilepsy other than epileptic spasms. Cochrane Database Syst. Rev. 2015, 2015, CD005222. [Google Scholar] [CrossRef]
- A Kelley, S.; Kossoff, E.H. Doose syndrome (myoclonic-astatic epilepsy): 40 years of progress. Dev. Med. Child Neurol. 2010, 52, 988–993. [Google Scholar] [CrossRef] [PubMed]
- Storoni, M.; Plant, G.T. The Therapeutic Potential of the Ketogenic Diet in Treating Progressive Multiple Sclerosis. Mult. Scler. Int. 2015, 2015, 681289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.Y.; Hao, J.; Liu, R.; Turner, G.; Shi, F.-D.; Rho, J.M. Inflammation-Mediated Memory Dysfunction and Effects of a Ketogenic Diet in a Murine Model of Multiple Sclerosis. PLOS ONE 2012, 7, e35476. [Google Scholar] [CrossRef] [Green Version]
- Valburg, C.; Sonti, A.; Stern, J.N.; Najjar, S.; Harel, A. Dietary factors in experimental autoimmune encephalomyelitis and multiple sclerosis: A comprehensive review. Mult. Scler. J. 2020, 27, 494–502. [Google Scholar] [CrossRef]
- Bock, M.; Karber, M.; Kuhn, H. Ketogenic diets attenuate cyclooxygenase and lipoxygenase gene expression in multiple sclerosis. eBioMedicine 2018, 36, 293–303. [Google Scholar] [CrossRef]
- Düking, T.; Spieth, L.; Berghoff, S.A.; Piepkorn, L.; Schmidke, A.M.; Mitkovski, M.; Kannaiyan, N.; Hosang, L.; Scholz, P.; Shaib, A.H.; et al. Ketogenic Diet Uncovers Differential Metabolic Plasticity of Brain Cells. Science 2022, 8, eabo7639. Available online: https://www.science.org (accessed on 28 November 2022). [CrossRef]
- Lin, W.-S.; Lin, S.-J.; Liao, P.-Y.; Suresh, D.; Hsu, T.-R.; Wang, P.-Y. Role of Ketogenic Diets in Multiple Sclerosis and Related Animal Models: An Updated Review. Adv. Nutr. Int. Rev. J. 2022, 13, 2002–2014. [Google Scholar] [CrossRef]
- Piacente, F.; Bottero, M.; Benzi, A.; Vigo, T.; Uccelli, A.; Bruzzone, S.; Ferrara, G. Neuroprotective Potential of Dendritic Cells and Sirtuins in Multiple Sclerosis. Int. J. Mol. Sci. 2022, 23, 4352. [Google Scholar] [CrossRef]
- Göschl, L.; Preglej, T.; Hamminger, P.; Bonelli, M.; Andersen, L.; Boucheron, N.; Gülich, A.F.; Müller, L.; Saferding, V.; Mufazalov, I.A.; et al. A T cell-specific deletion of HDAC1 protects against experimental autoimmune encephalomyelitis. J. Autoimmun. 2017, 86, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Martin, A.; Tegla, C.A.; Cudrici, C.D.; Kruszewski, A.M.; Azimzadeh, P.; Boodhoo, D.; Mekala, A.P.; Rus, V.; Rus, H. Role of SIRT1 in autoimmune demyelination and neurodegeneration. Immunol. Res. 2014, 61, 187–197. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Zhang, N.; Zhang, R.; Jin, L.; Petridis, A.K.; Loers, G.; Zheng, X.; Wang, Z.; Siebert, H.-C. Cuprizone-Induced Demyelination in Mouse Hippocampus Is Alleviated by Ketogenic Diet. J. Agric. Food Chem. 2020, 68. [Google Scholar] [CrossRef] [PubMed]
- Wouters, E.; Grajchen, E.; Jorissen, W.; Dierckx, T.; Wetzels, S.; Loix, M.; Tulleners, M.P.; Staels, B.; Stinissen, P.; Haidar, M.; et al. Altered PPARγ Expression Promotes Myelin-Induced Foam Cell Formation in Macrophages in Multiple Sclerosis. Int. J. Mol. Sci. 2020, 21, 9329. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Guo, Y.; Ge, Z.; Zhang, Z.; Da, Y.; Li, W.; Zhang, Z.; Xue, Z.; Li, Y.; Ren, Y.; et al. Adiponectin Suppresses T Helper 17 Cell Differentiation and Limits Autoimmune CNS Inflammation via the SIRT1/PPARγ/RORγt Pathway. Mol. Neurobiol. 2016, 54, 4908–4920. [Google Scholar] [CrossRef]
- Ai, R.-S.; Xing, K.; Deng, X.; Han, J.-J.; Hao, D.-X.; Qi, W.-H.; Han, B.; Yang, Y.-N.; Li, X.; Zhang, Y. Baicalin Promotes CNS Remyelination via PPARγ Signal Pathway. Neurol.-Neuroimmunol. Neuroinflammation 2022, 9, e1142. [Google Scholar] [CrossRef]
- Kanakasabai, S.; Pestereva, E.; Chearwae, W.; Gupta, S.K.; Ansari, S.; Bright, J.J. PPARγ Agonists Promote Oligodendrocyte Differentiation of Neural Stem Cells by Modulating Stemness and Differentiation Genes. PLoS ONE 2012, 7, e50500. [Google Scholar] [CrossRef]
- Brenton, J.N.; Banwell, B.; Bergqvist, A.C.; Lehner-Gulotta, D.; Gampper, L.; Leytham, E.; Coleman, R.; Goldman, M.D. Pilot study of a ketogenic diet in relapsing-remitting MS. Neurol.-Neuroimmunol. Neuroinflammation 2019, 6, e565. [Google Scholar] [CrossRef] [Green Version]
- Bahr, L.S.; Bock, M.; Liebscher, D.; Bellmann-Strobl, J.; Franz, L.; Prüß, A.; Schumann, D.; Piper, S.K.; Kessler, C.S.; Steckhan, N.; et al. Ketogenic diet and fasting diet as Nutritional Approaches in Multiple Sclerosis (NAMS): Protocol of a randomized controlled study. Trials 2020, 21, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Alizadeh, A.; Dyck, S.M.; Karimi-Abdolrezaee, S. Traumatic spinal cord injury: An overview of pathophysiology, models and acute injury mechanisms. Front. Neurol. 2019, 10, 282. [Google Scholar] [CrossRef] [Green Version]
- Streijger, F.; Plunet, W.T.; Lee, J.H.T.; Liu, J.; Lam, C.K.; Park, S.; Hilton, B.; Fransen, B.L.; Matheson, K.A.J.; Assinck, P.; et al. Ketogenic Diet Improves Forelimb Motor Function after Spinal Cord Injury in Rodents. PLoS ONE 2013, 8, e78765. [Google Scholar] [CrossRef] [PubMed]
- Seira, O.; Kolehmainen, K.; Liu, J.; Streijger, F.; Haegert, A.; Lebihan, S.; Boushel, R.; Tetzlaff, W. Ketogenesis controls mitochondrial gene expression and rescues mitochondrial bioenergetics after cervical spinal cord injury in rats. Sci. Rep. 2021, 11, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.-C. NF-κB signaling in inflammation. Signal Transduct. Target. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef] [Green Version]
- Kracht, M.; Müller-Ladner, U.; Schmitz, M.L. Mutual regulation of metabolic processes and proinflammatory NF-κB signaling. J. Allergy Clin. Immunol. 2020, 146, 694–705. [Google Scholar] [CrossRef] [PubMed]
- Roberti, A.; Chaffey, L.E.; Greaves, D.R. NF-κB Signaling and Inflammation—Drug Repurposing to Treat Inflammatory Disorders? Biology 2022, 11, 372. [Google Scholar] [CrossRef]
- Lawrence, T. The Nuclear Factor NF-kappa B Pathway in Inflammation. Cold Spring Harb. Perspect. Biol. 2009, 1, a001651. [Google Scholar] [CrossRef] [Green Version]
- Poewe, W.; Seppi, K.; Tanner, C.M.; Halliday, G.M.; Brundin, P.; Volkmann, J.; Schrag, A.E.; Lang, A.E. Parkinson disease. Nat. Rev. Dis. Primers 2017, 3, 17013. [Google Scholar] [CrossRef]
- Bloem, B.R.; Okun, M.S.; Klein, C. Parkinson’s disease. Lancet 2021, 397, 2284–2303. [Google Scholar] [CrossRef]
- Jankovic, J. Parkinson’s disease: Clinical features and diagnosis. J. Neurol. Neurosurg. Psychiatry 2008, 79, 368–376. [Google Scholar] [CrossRef] [Green Version]
- Mullin, S.; Stokholm, M.G.; Hughes, D.; Mehta, A.; Parbo, P.; Hinz, R.; Pavese, N.; Brooks, D.J.; Schapira, A.H. Brain Microglial Activation Increased in Glucocerebrosidase ( GBA ) Mutation Carriers without Parkinson’s disease. Mov. Disord. 2020, 36, 774–779. [Google Scholar] [CrossRef]
- Ferreira, S.A.; Romero-Ramos, M. Microglia Response During Parkinson’s Disease: Alpha-Synuclein Intervention. Front. Cell. Neurosci. 2018, 12, 247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Armstrong, M.J.; Okun, M.S. Diagnosis and Treatment of Parkinson Disease. JAMA 2020, 323, 548–560. [Google Scholar] [CrossRef] [PubMed]
- Sveinbjornsdottir, S. The clinical symptoms of Parkinson’s disease. J. Neurochem. 2016, 139, 318–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Cheng, B. Neuroprotective and Anti-inflammatory Activities of Ketogenic Diet on MPTP-induced Neurotoxicity. J. Mol. Neurosci. 2010, 42, 145–153. [Google Scholar] [CrossRef] [PubMed]
- George, S.; Rey, N.L.; Tyson, T.; Esquibel, C.; Meyerdirk, L.; Schulz, E.; Pierce, S.; Burmeister, A.R.; Madaj, Z.; Steiner, J.A.; et al. Microglia affect α-synuclein cell-to-cell transfer in a mouse model of Parkinson’s disease. Mol. Neurodegener. 2019, 14, 1–22. [Google Scholar] [CrossRef]
- Ingelfinger, F.; Gerdes, L.A.; Kavaka, V.; Krishnarajah, S.; Friebel, E.; Galli, E.; Zwicky, P.; Furrer, R.; Peukert, C.; Dutertre, C.-A.; et al. Twin study reveals non-heritable immune perturbations in multiple sclerosis. Nature 2022, 603, 152–158. [Google Scholar] [CrossRef]
- Badanjak, K.; Fixemer, S.; Smajić, S.; Skupin, A.; Grünewald, A. The Contribution of Microglia to Neuroinflammation in Parkinson’s Disease. Int. J. Mol. Sci. 2021, 22, 4676. [Google Scholar] [CrossRef]
- Ma, Z.; Zhang, D.; Li, J. A dedicated collaboration platform for integrated project delivery. Autom. Constr. 2018, 86, 199–209. [Google Scholar] [CrossRef]
- Olson, C.A.; Vuong, H.E.; Yano, J.M.; Liang, Q.Y.; Nusbaum, D.J.; Hsiao, E.Y. The Gut Microbiota Mediates the Anti-Seizure Effects of the Ketogenic Diet. Cell 2018, 173, 1728–1741.e13. [Google Scholar] [CrossRef] [Green Version]
- Sampson, T.R.; Challis, C.; Jain, N.; Moiseyenko, A.; Ladinsky, M.S.; Shastri, G.G.; Thron, T.; Needham, B.D.; Horvath, I.; Debelius, J.W.; et al. A gut bacterial amyloid promotes α-synuclein aggregation and motor impairment in mice. eLife 2020, 9, e53111. [Google Scholar] [CrossRef]
- Kam, T.-I.; Hinkle, J.T.; Dawson, T.M.; Dawson, V.L. Microglia and astrocyte dysfunction in parkinson’s disease. Neurobiol. Dis. 2020, 144, 105028. [Google Scholar] [CrossRef] [PubMed]
- Batch, J.T.; Lamsal, S.P.; Adkins, M.; Sultan, S.; Ramirez, M.N. Advantages and Disadvantages of the Ketogenic Diet: A Review Article. Cureus 2020, 12, e9639. [Google Scholar] [CrossRef] [PubMed]
- Crosby, L.; Davis, B.; Joshi, S.; Jardine, M.; Paul, J.; Neola, M.; Barnard, N.D. Ketogenic Diets and Chronic Disease: Weighing the Benefits Against the Risks. Front. Nutr. 2021, 8. [Google Scholar] [CrossRef] [PubMed]
- Newmaster, K.; Zhu, Z.; Bolt, E.; Chang, R.J.; Day, C.; Mhanna, A.; Paudel, S.; Farooq, O.; Swaminathan, A.; Acharya, P.; et al. A Review of the Multi-Systemic Complications of a Ketogenic Diet in Children and Infants with Epilepsy. Children 2022, 9, 1372. [Google Scholar] [CrossRef]
- Valencia, I.; Pfeifer, H.; Thiele, E.A. General Anesthesia and the Ketogenic Diet: Clinical Experience in Nine Patients. Epilepsia 2002, 43, 525–529. [Google Scholar] [CrossRef]
- Breu, M.; Häfele, C.; Glatter, S.; Trimmel-Schwahofer, P.; Golej, J.; Male, C.; Feucht, M.; Dressler, A. Ketogenic Diet in the Treatment of Super-Refractory Status Epilepticus at a Pediatric Intensive Care Unit: A Single-Center Experience. Front. Neurol. 2021, 12. [Google Scholar] [CrossRef]
- Sondhi, V.; Agarwala, A.; Pandey, R.M.; Chakrabarty, B.; Jauhari, P.; Lodha, R.; Toteja, G.S.; Sharma, S.; Paul, V.K.; Kossoff, E.; et al. Efficacy of Ketogenic Diet, Modified Atkins Diet, and Low Glycemic Index Therapy Diet Among Children With Drug-Resistant Epilepsy. JAMA Pediatr. 2020, 174, 944. [Google Scholar] [CrossRef]
- Alten, E.; Chaturvedi, A.; Cullimore, M.; Fallon, A.; Habben, L.; Hughes, I.; O’Malley, N.; Rahimi, H.; Renodin-Mead, D.; Schmidt, B.; et al. No longer a historical ailment: Two cases of childhood scurvy with recommendations for bone health providers. Osteoporos. Int. 2020, 31, 1001–1005. [Google Scholar] [CrossRef]
- Willmott, N.S.; Bryan, R.A.E. Scurvy in child with epilepsy on a ketogenic diet with oral complications. Eur. Arch. Paediatr. Dent. 2008, 9, 148–152. [Google Scholar] [CrossRef]
- Kidwell, R.; Tobias, J.D. Anesthetic care of a child with receiving a ketogenic diet. Compr. Pharmacol. 2021, 9, 85–93. [Google Scholar] [CrossRef]
- Grinspan, Z.M.; Shellhaas, R.A.; Coryell, J.; Sullivan, J.E.; Wirrell, E.C.; Mytinger, J.R.; Gaillard, W.D.; Kossoff, E.H.; Valencia, I.; Knupp, K.G.; et al. Comparative Effectiveness of Levetiracetam vs Phenobarbital for Infantile Epilepsy. JAMA Pediatr. 2018, 172, 352–360. [Google Scholar] [CrossRef] [PubMed]
- Erickson, J.C.; Jabbari, B.; DiFazio, M.P. Basal ganglia injury as a complication of the ketogenic diet. Mov. Disord. 2003, 18, 448–451. [Google Scholar] [CrossRef] [PubMed]
- Desai, A.A.; Thompson, L.M.; Abdelmoity, A.T.; Kayyali, H.; Peter, S.D.S. Management of Symptomatic Cholelithiasis While on Ketogenic Diet: A Case Report. Pediatr. Neurol. 2014, 51, 439–440. [Google Scholar] [CrossRef] [PubMed]
- Kapetanakis, M.; Liuba, P.; Odermarsky, M.; Lundgren, J.; Hallböök, T. Effects of ketogenic diet on vascular function. Eur. J. Paediatr. Neurol. 2014, 18, 489–494. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Role of ketogenic diet in modulation of different diseases.

Figure 2.
Modulation of different diseases by ketogenic diet.


{kind=link}
{kind=link}
Table 1.
The immunomodulatory effects of ketogenic diet on Immune Cells.
| Type of Ketogenic Diet (KD) | Mechanism of Action | Conclusions/Effects | References |
|---|---|---|---|
| Medium-Chain Triglycerides (MCT) | Reduced Leukocyte Count | Anti-Tumor Effect | [79] |
| Eucaloric Ketogenic Diet (EKD) | M1 Recruitment of Neutrophil and Platelets in Thrombo-Inflammation | Prevention of Cytokine Storm in COVID-19. | [80] |
| Very Low Calorie Ketogenic Diet (VLCKD) | Altered Leukocyte Methylation | Obesity Prevention | [81] |
| KD | Reduced WBC & Neutrophil Count | In Epilepsy, baseline immunosuppression does not worsen with KD. | [82] |
| KD | Enhanced activity of Th1 cells | Promoted cellular immune function in a CT26 colon tumor allografts mouse model. | [33] |
| KD | Suppression of Th1 & Th17 | Protection against autoinflammation (Central Nervous System). | [83] |
| KD | Suppression of macrophages & neutrophils | Attenuation of autoinflammation (Muckle-Wells syndrome/ Gout) | [83] |
| KD | Activation of protective γδ T cells and decreases myeloid cell subset | Ketogenesis-induced protection from mCoV-A59-driven inflammatory damage in aging. | [17] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Srivastava, S.; Pawar, V.A.; Tyagi, A.; Sharma, K.P.; Kumar, V.; Shukla, S.K. Immune Modulatory Effects of Ketogenic Diet in Different Disease Conditions. Immuno 2023, 3, 1-15. https://doi.org/10.3390/immuno3010001
AMA Style
Srivastava S, Pawar VA, Tyagi A, Sharma KP, Kumar V, Shukla SK. Immune Modulatory Effects of Ketogenic Diet in Different Disease Conditions. Immuno. 2023; 3(1):1-15. https://doi.org/10.3390/immuno3010001
Chicago/Turabian StyleSrivastava, Shivani, Vishakha Anand Pawar, Anuradha Tyagi, Kanti Prakash Sharma, Vinay Kumar, and Surendra Kumar Shukla. 2023. "Immune Modulatory Effects of Ketogenic Diet in Different Disease Conditions" Immuno 3, no. 1: 1-15. https://doi.org/10.3390/immuno3010001