
Uptake and Advanced Therapy of Butyrate in Inflammatory Bowel Disease
Department of Gastroenterology and Hematology, Hirosaki University Graduate School of Medicine, Hirosaki 036-8216, Japan
*
Author to whom correspondence should be addressed.
Immuno 2022, 2(4), 692-702; https://doi.org/10.3390/immuno2040042
Submission received: 5 October 2022
/
Revised: 14 November 2022
/
Accepted: 24 November 2022
/
Published: 25 November 2022
(This article belongs to the Special Issue GI Tract Immunology and Mucosal Immunity)
Abstract
:The pathogenesis and refractory nature of inflammatory bowel disease (IBD) are related to multiple factors, including genetic factors, environmental factors, and abnormalities in gut microbial diversity, which lead to decreased levels of short-chain fatty acids (SCFAs). Among SCFAs, butyrate plays an important role in mucosal barrier maintenance, serves as an energy source in intestinal epithelial cells (IECs), and exhibits anti-inflammatory effects; therefore, it is a particularly important factor in gut homeostasis. Changes in gut microbiota and butyrate levels affect the outcomes of drug therapy for IBD. Butyrate is mainly absorbed in the large intestine and is transported by monocarboxylate transporter 1 (MCT1) and sodium-coupled monocarboxylate transporter 1 (SMCT1). During gut inflammation, butyrate utilization and uptake are impaired in IECs. Dysbiosis and low abundance of butyrate affect fecal microbiota transplantation and anticancer immunotherapy. Although butyrate administration has been reported as a treatment for IBD, its effects remain controversial. In this review, we discuss butyrate absorption and metabolism in patients with IBD and their relationship with drug therapy.
1. Introduction
Ulcerative colitis (UC) and Crohn’s disease (CD) are considered the major forms of inflammatory bowel disease (IBD) and chronic inflammation in the intestine; however, their etiology is not fully understood. IBD onset is believed to be related to genetic factors, diet, and dysbiosis. IBD has the highest occurrence in developed countries, such as North America and Europe, and has recently been increasing in Asia [1,2]. Japan, Hong Kong, and Korea have an increased incidence of IBD. Regardless of genetic background, the number of patients with IBD has increased. This phenomenon suggests that the rapid increase in the incidence of IBD is parallel to changes in lifestyle, such as diet, breastfeeding, and antibiotic use [3]. Additionally, these parameters are closely related to the gut microbiota. Patients with IBD demonstrate significantly decreased microbial diversity and short-chain fatty acids (SCFAs) compared with healthy controls [4,5,6]. This suggests that environmental factors, such as microbiota and diet, contribute to abnormal immune responses during the onset of IBD.
Recent studies have shown that microbial diversity and fecal abundance of butyrate influence the success of biological therapies and thiopurine treatment [7,8,9,10]. These results suggest that interactions between intestinal dysbiosis and the host response are involved in both the onset of IBD and treatment outcomes. In this review, we describe the multiple effects and advanced therapies of butyrate, as well as the regulation of butyrate uptake into intestinal epithelial cells (IECs).
2. SCFAs and Intestinal Homeostasis in IBD
2.1. The Role of SCFAs in Intestinal Immune Response
SCFAs are metabolites produced by the intestinal microbiota and are mainly composed of acetate, propionate, and butyrate. SCFAs have various physiological effects in humans and play important roles in colon homeostasis [11]. SCFAs have many favorable effects, including their barrier function, serving as an energy source of IEC, activating G-protein coupled receptors (GPCRs), and inhibiting the effects of histone deacetylase (HDAC) on cell differentiation and proliferation [12]. GPCRs include GPR41, GPR43, and GPR109A; anti-inflammatory effects in the intestinal mucosa are due to HDAC inhibition and GPCR activation [13]. GPR41 is preferentially expressed in the adipose tissue, while GPR43 and GPR109A are expressed in IECs and immune cells. SCFAs modulate the immune response [13]. In particular, butyrate strongly induces regulatory T cell (Treg) differentiation by enhancing the production of TGF-β in the colon via acetylation of the Foxp3 promoter in naive T cells [14,15]. Butyrate also promotes IL-22 production from CD4+ T cells and innate lymphoid cells (ILCs); IL-22 plays an important role in maintaining the health of the epithelial barrier [16]. Moreover, butyrate, but not other SCFAs, enhances mucosal barrier function through actin-associated gene expression and AMP-activated protein kinase [17,18]. In colonic cell lines, by activating GPR109A, butyrate suppressed inflammation due to LPS-induced NF-kβ activation [19]. Butyrate suppressed the production of proinflammatory cytokines from neutrophils of patients with IBD and ameliorated inflammation in an animal colitis model by regulating neutrophils [20]. Thus, butyrate plays a major role in suppressing gut inflammation and maintaining epithelial barrier function.
2.2. SCFAs in IBD
Dysbiosis is generally accepted to be involved in IBD onset. Diet plays a role in gut homeostasis and influences gut microbiota composition, intestinal permeability, and mucosal barrier [21]. Westernized diets contain high sugar, high fat, and low fiber levels and high calories; they elevate the risk of IBD onset [22,23]. Butyrate is produced mainly by bacteria in the human colon, particularly Faecalibacterium prausnitzii, Eubacterium rectale, and Roseburia hominis [17,18]. Patients with IBD have diminished microbial diversity, that is, the abundance of F. prausnitzii and R. hominis is reduced [24,25]. Furthermore, IECs metabolism plays an important role in regulating the gut microbiota. In a healthy state, IECs utilize butyrate as an important energy source. Since butyrate metabolism consumes oxygen, surface colonocytes are hypoxic, which promotes the growth of anaerobic bacteria such as Firmicutes in the lumen. Gut inflammation leads to the reduction of Firmicutes and butyrate levels. In the absence of butyrate, IECs obtain energy through fermentation metabolism of glucose; consequently, oxygen concentration in the lumen increases to 3–10%, and it alters the local microbial composition [26]. Its decreases strict anaerobes while allowing the growth of facultative anaerobes such as Escherichia coli pathobionts, which are believed to be involved in IBD [26,27].
3. Butyrate Transport to IECs
Butyrate is a weak acid (pKa = 4.8), and more than 90% exists in an ionized form in the colon lumen; therefore, carrier-mediated transport to IECs is a fundamental pathway [28,29,30,31]. Butyrate exists mainly in the proximal colon [31]. Monocarboxylate transporter 1 (MCT1), encoded by SLC16A1, and sodium-coupled monocarboxylate transporter 1 (SMCT1), encoded by SLC5A8, are expressed in the apical membrane of colonocytes; these are major transporters involved in butyrate absorption [11]. Butyrate uptake mediated by MCT1 is based on Michaelis–Menten kinetics in the 4–10 mM range [32,33]. In contrast, SMCT1 is based on Michaelis–Menten kinetics in the low micromolar range (50–100 μM) [11]. The concentration range of butyrate in the human colon is 10–20 mM [31]. Therefore, SMCT1 is mostly saturated under normal conditions. Butyrate is absorbed by IECs and is metabolized as an energy source. Butyrate undergoes β-oxidation in mitochondria and enters the tricarboxylic acid (TCA) cycle, leading to energy production (Figure 1) [34]. This mechanism provides 70–80% of the IEC energy requirements [28]. Decreased butyrate absorption in IECs causes autophagy via AMPK activation [35]. In contrast, unmetabolized butyrate inhibits histone deacetylase (HDAC) activity and is partially transported to the basolateral side by MCT4 [11,36]. Systemic butyrate concentration in human peripheral blood is 1–10 µM [31,37,38].
4. Regulation and Role of MCT1 in IECs
Luminal butyrate concentration can be modulated by diet, gut inflammation, and dysbiosis. Butyrate itself enhances MCT1 expression via the NF-kβ pathway and mRNA stabilization [39,40]. Abundance of fiber in the diet influences its absorption. In rats and pigs fed with pectin or resistant-starch diets, the expression of MCT1 was increased in the colon [41,42]. Moreover, luminal butyrate activates GPR109A, which is also reported as an SCFA sensor and expressed in the apical membrane of IECs. Activated GPR109A enhances membrane MCT1 expression via decreased cAMP pathway [43]. As a result, Vmax of MCT1-mediated butyrate uptake is increased, which leads to efficient absorption [43]. Previous studies have reported the membrane expression of MCT1 without changes at the transcriptional level. Although precise mechanisms are not known, including the recycling mechanism and protein trafficking systems, calcineurin has been suggested to regulate cAMP degradation and affect cytoskeletal function [33,44,45]. In IBD patients, intestinal MCT1 protein expression is downregulated [46]. This is considered to be due to a decrease in butyrate-producing bacteria and inflammation itself. Interferon γ (IFN-γ) and TNF-α signaling downregulate the expression of MCT1 in HT-29 cells [46]. In pig colonic tissue and Caco-2 cells, TNF-α downregulated MCT1 expression [47]. Thus, MCT1 expression and function are dynamically altered by intestinal butyrate and inflammation. Moreover, in the intestinal mucosa of active UC, the intracellular butyrate availability is reduced due to the downregulation of β-oxidation pathway [48]. Hence, inflammation impair the butyrate uptake and metabolism in IECs, followed by the disruption of gut homeostasis.
In a previous study, the ability of organoid epithelial cells to perform butyrate uptake, oxidization, and metabolism in patients with non-active IBD was comparable to that in healthy individuals; however, in patients with active IBD and upon TNF-α stimulation, this ability was decreased [13,49]. Thus, in the active inflammatory state in IBD, the uptake and utilization of butyrate are decreased, even if abundant butyrate is available from microbiota or supplementation.
5. Gut Microbiota and Systemic Immune Response
The development of new therapeutic agents based on an advanced understanding of IBD pathophysiology has resulted in favorable outcomes and reduced disease-related morbidity. The recently proposed treat-to-target (T2T) strategy has shown that early initiation of advanced therapies, such as anti-TNF-α agents, leads to greater response. As per this concept, personalized treatment according to clinical poor prognostic factors is key. However, prediction of the treatment response to each therapeutic drug based on the mode of action is not yet established. In recent years, the association between the intestinal microbiota and various immune-mediated diseases has attracted attention. Several studies have highlighted the role of gut microbiota and metabolites in the response to treatments, including biologics, fecal microbiota transplantation (FMT), and anti-cancer immunotherapy. Restoration of intestinal dysbiosis reduced diversity, and butyrate abundance by FMT directly targets dysbiosis. Successful treatment with FMT has been associated with the presence of Eubacterium and Roseburia species, SCFA biosynthesis, and secondary bile acids. In contrast, failure of FMT treatment has been associated with Fusobacterium, Sutterella, and Escherichia species [50]. Previous studies have shown a correlation between gut microbiota and anti-cancer immunotherapy, such as cyclophosphamide [51]. Specific species of bacteria induce Th1 cell immune responses against lung and ovarian cancers and predict long-term anticancer responses. The gut microbiota is believed to affect the tumor microenvironment (TME) to regulate anticancer effector T cells and cytokine responses.
In patients with IBD, gut microbes and high butyrate levels predict a better response to biologics and azathioprine (Table 1) [7,8,9,10]. However, the precise pathway by which the gut microbiota affects the response to biological therapies remains obscure. One mechanism hypothesized is that a low level of decreased diversity and butyrate abundance minimizes mucosal barrier disruption, because of which immunomodulatory therapy is expected to easily succeed. Further studies are required to uncover the full mechanistic pathway by which the gut microbiota and its metabolites improve the response to advanced therapies for IBD.
6. Influence of Gut Microbiota on Response to Advanced Therapies
Maier et al. reported that 24% of 1197 drugs against 40 representative gut microbiota strains affect the growth of bacteria, thus modifying the efficacy and side effects of the drugs [52,53]. Butyrate ameliorates gut inflammation through multiple pathways and regulates the mucosal barrier function. Butyrate is considered the principal ATP source for IECs [54]. IEC obtained by biopsy from patients with UC shows impaired butyrate β-oxidation [55,56]. For example, it results from decreased expression of mitochondrial acetoacetyl-CoA thiolase activity [57]. Butyrate enhances mucosal barrier function; however, the barrier is disrupted during active inflammation [58]. The expression of MCT1 in the colon is reportedly decreased in patients with IBD and experimental colitis models [46,47,59]. In patients with active IBD, butyrate utilization and uptake are impaired (Figure 2). The low amount of butyrate and other abnormalities of the microbiota are speculated to contribute to the refractory disease, even if drugs suppress inflammatory cytokines and immune cells. We reported that the efficacy of cyclosporine treatment in an animal colitis model deteriorated with antibiotic pretreatment but recovered with butyrate supplementation [60]. We showed that cyclosporine treatment upregulates the expression of membrane MCT1 in IECs without inducing transcriptional changes. Regarding the efficacy of drugs for IBD, focusing on butyrate absorption and abundance may be necessary. Butyrate exhibits a direct antimicrobial effect, for example, on Acinetobacter baumannii, E. coli, and Staphylococcus pseudintermedius, by inducing bacterial membrane depolarization and cytosolic acidification [61]. Moreover, a previous report showed that butyrate induced antimicrobial peptides, such as LL-37, in IECs in vivo and colon cell lines [62]. Thus, butyrate improves gut microbiota.
Additionally, the gut microbiota is influenced by drug therapy. 5-Aminosalicylic acid (5-ASA) treatment altered the balance between increase in Proteobacteria and decrease in Firmicutes in inflamed mucosa in patients with UC [63]. Furthermore, after anti-TNF-αtherapy for patients with CD, the phyla Proteobacteria and Clostridia increased, and α-diversity was increased in anti-TNF-α therapy responders [64]. The F. prausnitzii/E. coli ratio was associated with the responder and non-responder status; it served as a biomarker of response to anti-TNF-α therapy [64].
7. Advanced Therapy with Butyrate
Treatments focusing on the supply of butyrate and intestinal microflora to the colon have been reported. Several reports have described that butyrate administration improves gut inflammation and relieves symptoms and microbiota changes in patients with IBD [65,66]. However, oral intake of butyrate is unacceptable because of its offensive odor and absorption in the small intestine. There have been several reports on butyrate modification. Tributyrin overcomes this disadvantage [67]. It is an ester composed of butyric acid and glycerol and is used as a food additive. However, till data, no clinical reports indicate that tributyrin improves the condition of patients with IBD [68]. Butyration of starch has been showed to enable effective delivery to the colon [69,70]. Butyrylated starch is resistant to amylolysis in the small intestine and is released in the colon by bacterial esterase [71]. It has shown good efficacy in multiple mouse models of DSS-induced colitis [72]. Mu et al. reported polyvinyl butyrate nanoparticles that were efficiently delivered to intestinal macrophages and were effective against mouse DSS colitis [73]. Clinical trials in patients with IBD are expected.
In patients with mild to moderate UC, oral therapeutics of Firmicutes spores, which were purified from the stool of healthy donors by ethanol treatment, achieved a higher clinical remission rate compared with the placebo [74]. Butyrate did not change significantly after treatment; however, the ratio of butyrate to propionate tended to increase.
Oral treatment with microencapsulated butyrate increased the abundance of SCFA-producing microbiota, and with add-on therapy, maintained remission in patients with IBD [66,75].
However, some reports showed that butyrate treatment not only lacks efficacy in patients with IBD but also worsens colitis (Table 2 and Table 3). Organoid culture sampling from the intestines of patients with UC showed that butyrate worsens barrier function and inflammatory cytokine expression under co-stimulation with TNF-α and IFN-γ [76]. Kaiko et al. reported that butyrate prevents wound repair of intestinal stem cells [77]. Butyrate is potentially toxic at very high concentrations in the colon lumen [78]. Furthermore, reduction in luminal pH is associated with inhibition of the metabolic activity of luminal bacteria, and it can potentially have detrimental effects on other functions of the gut microbiota [68,79]. The luminal concentration of SCFA and the pH profile achieved by these compounds is unknown [68]. Butyrate treatment for IBD may be controversial because of the severity of inflammation in patients participating in clinical studies. Studies have suggested that because of the reduction in butyrate transporter levels, treatment with butyrate fails to improve mucosal damage in patients with active IBD.
8. Conclusions
Therapeutic approaches targeting the gut microbiota may be of great importance for improving treatment outcomes. Butyrate monotherapy is unlikely to cure refractory IBD; however, its plays an adjunctive role in correcting immune abnormalities and epithelial barrier function in the colon, thus enhancing the effects of other drugs. Since the correction of abnormalities in the gut microbiota and appropriate levels of luminal butyrate are necessary for achieving a therapeutic effect, microbiological metabolism and other factors should be considered when conventional therapy has been refractory. The development of a simple marker for easily identifying patients who need gut microbial profile correction is expected. Combining advanced medical therapy with the appropriate restoration of microbial diversity will lead to better outcomes in patients with IBD.
Author Contributions
Conceptualization, S.O. and H.S.; writing—original draft preparation, S.O. and H.S.; writing—review and editing, S.O. and H.S.; visualization, S.O.; supervision, H.S.; project administration, H.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We thank all the staffs of our laboratories for their support of this work.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Molodecky, N.A.; Soon, I.S.; Rabi, D.M.; Ghali, W.A.; Ferris, M.; Chernoff, G.; Benchimol, E.I.; Panaccione, R.; Ghosh, S.; Barkema, H.W.; et al. Increasing Incidence and Prevalence of the Inflammatory Bowel Diseases with Time, Based on Systematic Review. Gastroenterology 2012, 142, 46–54.e42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ng, W.K.; Wong, S.H.; Ng, S.C. Changing Epidemiological Trends of Inflammatory Bowel Disease in Asia. Intest. Res. 2016, 14, 111–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ananthakrishnan, A.N. Epidemiology and Risk Factors for IBD. Nat. Rev. Gastroenterol. Hepatol. 2015, 12, 205–217. [Google Scholar] [CrossRef] [PubMed]
- Glassner, K.L.; Abraham, B.P.; Quigley, E.M.M. The Microbiome and Inflammatory Bowel Disease. J. Allergy Clin. Immunol. 2020, 145, 16–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchesi, J.R.; Holmes, E.; Khan, F.; Kochhar, S.; Scanlan, P.; Shanahan, F.; Wilson, I.D.; Wang, Y. Rapid and Noninvasive Metabonomic Characterization of Inflammatory Bowel Disease. J. Proteome Res. 2007, 6, 546–551. [Google Scholar] [CrossRef] [PubMed]
- Imhann, F.; Vich Vila, A.; Bonder, M.J.; Fu, J.; Gevers, D.; Visschedijk, M.C.; Spekhorst, L.M.; Alberts, R.; Franke, L.; van Dullemen, H.M.; et al. Interplay of Host Genetics and Gut Microbiota Underlying the Onset and Clinical Presentation of Inflammatory Bowel Disease. Gut 2018, 67, 108–119. [Google Scholar] [CrossRef]
- Ananthakrishnan, A.N.; Luo, C.; Yajnik, V.; Khalili, H.; Garber, J.J.; Stevens, B.W.; Cleland, T.; Xavier, R.J. Gut Microbiome Function Predicts Response to Anti-Integrin Biologic Therapy in Inflammatory Bowel Diseases. Cell Host Microbe 2017, 21, 603–610.e3. [Google Scholar] [CrossRef] [Green Version]
- Effenberger, M.; Reider, S.; Waschina, S.; Bronowski, C.; Enrich, B.; Adolph, T.E.; Koch, R.; Moschen, A.R.; Rosenstiel, P.; Aden, K.; et al. Microbial Butyrate Synthesis Indicates Therapeutic Efficacy of Azathioprine in IBD Patients. J. Crohns. Colitis. 2021, 15, 88–98. [Google Scholar] [CrossRef]
- Aden, K.; Rehman, A.; Waschina, S.; Pan, W.-H.; Walker, A.; Lucio, M.; Nunez, A.M.; Bharti, R.; Zimmerman, J.; Bethge, J.; et al. Metabolic Functions of Gut Microbes Associate with Efficacy of Tumor Necrosis Factor Antagonists in Patients with Inflammatory Bowel Diseases. Gastroenterology 2019, 157, 1279–1292.e11. [Google Scholar] [CrossRef] [Green Version]
- Magnusson, M.K.; Strid, H.; Sapnara, M.; Lasson, A.; Bajor, A.; Ung, K.-A.; Öhman, L. Anti-TNF Therapy Response in Patients with Ulcerative Colitis Is Associated with Colonic Antimicrobial Peptide Expression and Microbiota Composition. ECCOJC 2016, 10, 943–952. [Google Scholar] [CrossRef]
- Sivaprakasam, S.; Bhutia, Y.D.; Yang, S.; Ganapathy, V. Short-Chain Fatty Acid Transporters: Role in Colonic Homeostasis. Compr. Physiol. 2017, 8, 299–314. [Google Scholar] [CrossRef]
- Chang, P.V.; Hao, L.; Offermanns, S.; Medzhitov, R. The Microbial Metabolite Butyrate Regulates Intestinal Macrophage Function via Histone Deacetylase Inhibition. Proc. Natl. Acad. Sci. USA 2014, 111, 2247–2252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parada Venegas, D.; De la Fuente, M.K.; Landskron, G.; González, M.J.; Quera, R.; Dijkstra, G.; Harmsen, H.J.M.; Faber, K.N.; Hermoso, M.A. Short Chain Fatty Acids (SCFAs)-Mediated Gut Epithelial and Immune Regulation and Its Relevance for Inflammatory Bowel Diseases. Front. Immunol. 2019, 10, 277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atarashi, K.; Tanoue, T.; Shima, T.; Imaoka, A.; Kuwahara, T.; Momose, Y.; Cheng, G.; Yamasaki, S.; Saito, T.; Ohba, Y.; et al. Induction of Colonic Regulatory T Cells by Indigenous Clostridium Species. Science 2011, 331, 337–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furusawa, Y.; Obata, Y.; Fukuda, S.; Endo, T.A.; Nakato, G.; Takahashi, D.; Nakanishi, Y.; Uetake, C.; Kato, K.; Kato, T.; et al. Commensal Microbe-Derived Butyrate Induces the Differentiation of Colonic Regulatory T Cells. Nature 2013, 504, 446–450. [Google Scholar] [CrossRef]
- Yang, W.; Yu, T.; Huang, X.; Bilotta, A.J.; Xu, L.; Lu, Y.; Sun, J.; Pan, F.; Zhou, J.; Zhang, W.; et al. Intestinal Microbiota-Derived Short-Chain Fatty Acids Regulation of Immune Cell IL-22 Production and Gut Immunity. Nat. Commun. 2020, 11, 4457. [Google Scholar] [CrossRef]
- Wang, R.X.; Lee, J.S.; Campbell, E.L.; Colgan, S.P. Microbiota-Derived Butyrate Dynamically Regulates Intestinal Homeostasis through Regulation of Actin-Associated Protein Synaptopodin. Proc. Natl. Acad. Sci. USA 2020, 117, 11648–11657. [Google Scholar] [CrossRef]
- Peng, L.; Li, Z.-R.; Green, R.S.; Holzman, I.R.; Lin, J. Butyrate Enhances the Intestinal Barrier by Facilitating Tight Junction Assembly via Activation of AMP-Activated Protein Kinase in Caco-2 Cell Monolayers. J. Nutr. 2009, 139, 1619–1625. [Google Scholar] [CrossRef] [Green Version]
- Thangaraju, M.; Cresci, G.A.; Liu, K.; Ananth, S.; Gnanaprakasam, J.P.; Browning, D.D.; Mellinger, J.D.; Smith, S.B.; Digby, G.J.; Lambert, N.A.; et al. GPR109A Is a G-Protein–Coupled Receptor for the Bacterial Fermentation Product Butyrate and Functions as a Tumor Suppressor in Colon. Cancer Res. 2009, 69, 2826–2832. [Google Scholar] [CrossRef] [Green Version]
- Li, G.; Lin, J.; Zhang, C.; Gao, H.; Lu, H.; Gao, X.; Zhu, R.; Li, Z.; Li, M.; Liu, Z. Microbiota Metabolite Butyrate Constrains Neutrophil Functions and Ameliorates Mucosal Inflammation in Inflammatory Bowel Disease. Gut Microbes 2021, 13, 1968257. [Google Scholar] [CrossRef]
- Rizzello, F.; Spisni, E.; Giovanardi, E.; Imbesi, V.; Salice, M.; Alvisi, P.; Valerii, M.C.; Gionchetti, P. Implications of the Westernized Diet in the Onset and Progression of IBD. Nutrients 2019, 11, 1033. [Google Scholar] [CrossRef] [Green Version]
- Hou, J.K.; Abraham, B.; El-Serag, H. Dietary Intake and Risk of Developing Inflammatory Bowel Disease: A Systematic Review of the Literature. Am. J. Gastroenterol. 2011, 106, 563–573. [Google Scholar] [CrossRef] [PubMed]
- Dolan, K.T.; Chang, E.B. Diet, Gut Microbes, and the Pathogenesis of Inflammatory Bowel Diseases. Mol. Nutr. Food Res. 2017, 61, 1600129. [Google Scholar] [CrossRef] [Green Version]
- Kostic, A.D.; Xavier, R.J.; Gevers, D. The Microbiome in Inflammatory Bowel Disease: Current Status and the Future Ahead. Gastroenterology 2014, 146, 1489–1499. [Google Scholar] [CrossRef] [Green Version]
- Louis, P.; Flint, H.J. Formation of Propionate and Butyrate by the Human Colonic Microbiota. Environ. Microbiol. 2017, 19, 29–41. [Google Scholar] [CrossRef] [Green Version]
- Celiberto, L.S.; Graef, F.A.; Healey, G.R.; Bosman, E.S.; Jacobson, K.; Sly, L.M.; Vallance, B.A. Inflammatory Bowel Disease and Immunonutrition: Novel Therapeutic Approaches through Modulation of Diet and the Gut Microbiome. Immunology 2018, 155, 36–52. [Google Scholar] [CrossRef]
- Litvak, Y.; Byndloss, M.X.; Tsolis, R.M.; Bäumler, A.J. Dysbiotic Proteobacteria Expansion: A Microbial Signature of Epithelial Dysfunction. Curr. Opin. Microbiol. 2017, 39, 1–6. [Google Scholar] [CrossRef]
- Gasaly, N.; Hermoso, M.A.; Gotteland, M. Butyrate and the Fine-Tuning of Colonic Homeostasis: Implication for Inflammatory Bowel Diseases. IJMS 2021, 22, 3061. [Google Scholar] [CrossRef]
- Gonçalves, P.; Araújo, J.R.; Pinho, M.J.; Martel, F. Modulation of Butyrate Transport in Caco-2 Cells. Naunyn-Schmied. Arch. Pharm. 2009, 379, 325–336. [Google Scholar] [CrossRef]
- Gonçalves, P.; Martel, F. Regulation of Colonic Epithelial Butyrate Transport: Focus on Colorectal Cancer. Porto. Biomed. J. 2016, 1, 83–91. [Google Scholar] [CrossRef]
- Cummings, J.H.; Pomare, E.W.; Branch, W.J.; Naylor, C.P.; Macfarlane, G.T. Short Chain Fatty Acids in Human Large Intestine, Portal, Hepatic and Venous Blood. Gut 1987, 28, 1221–1227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hadjiagapiou, C.; Schmidt, L.; Dudeja, P.K.; Layden, T.J.; Ramaswamy, K. Mechanism(s) of Butyrate Transport in Caco-2 Cells: Role of Monocarboxylate Transporter 1. Am. J. Physiol. Gastrointest. Liver Physiol. 2000, 279, G775–G780. [Google Scholar] [CrossRef] [PubMed]
- Saksena, S.; Theegala, S.; Bansal, N.; Gill, R.K.; Tyagi, S.; Alrefai, W.A.; Ramaswamy, K.; Dudeja, P.K. Mechanisms Underlying Modulation of Monocarboxylate Transporter 1 (MCT1) by Somatostatin in Human Intestinal Epithelial Cells. Am. J. Physiol. Gastrointest. Liver Physiol. 2009, 297, G878–G885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blachier, F.; de Sá Resende, A.; da Silva Fogaça Leite, G.; Vasques da Costa, A.; Lancha Junior, A.H. Colon Epithelial Cells Luminal Environment and Physiopathological Consequences: Impact of Nutrition and Exercise. Nutrire 2018, 43, 2. [Google Scholar] [CrossRef]
- Donohoe, D.R.; Garge, N.; Zhang, X.; Sun, W.; O’Connell, T.M.; Bunger, M.K.; Bultman, S.J. The Microbiome and Butyrate Regulate Energy Metabolism and Autophagy in the Mammalian Colon. Cell Metab. 2011, 13, 517–526. [Google Scholar] [CrossRef] [Green Version]
- Gill, R.K.; Saksena, S.; Alrefai, W.A.; Sarwar, Z.; Goldstein, J.L.; Carroll, R.E.; Ramaswamy, K.; Dudeja, P.K. Expression and Membrane Localization of MCT Isoforms along the Length of the Human Intestine. Am. J. Physiol. Cell Physiol. 2005, 289, C846–C852. [Google Scholar] [CrossRef] [Green Version]
- Bonomo, R.R.; Cook, T.M.; Gavini, C.K.; White, C.R.; Jones, J.R.; Bovo, E.; Zima, A.V.; Brown, I.A.; Dugas, L.R.; Zakharian, E.; et al. Fecal Transplantation and Butyrate Improve Neuropathic Pain, Modify Immune Cell Profile, and Gene Expression in the PNS of Obese Mice. Proc. Natl. Acad. Sci. USA 2020, 117, 26482–26493. [Google Scholar] [CrossRef]
- Bloemen, J.G.; Venema, K.; van de Poll, M.C.; Olde Damink, S.W.; Buurman, W.A.; Dejong, C.H. Short Chain Fatty Acids Exchange across the Gut and Liver in Humans Measured at Surgery. Clin. Nutr. 2009, 28, 657–661. [Google Scholar] [CrossRef]
- Cuff, M.A.; Lambert, D.W.; Shirazi-beechey, S.P. Substrate-Induced Regulation of the Human Colonic Monocarboxylate Transporter, MCT1. J. Physiol. 2002, 539, 361–371. [Google Scholar] [CrossRef]
- Borthakur, A.; Saksena, S.; Gill, R.K.; Alrefai, W.A.; Ramaswamy, K.; Dudeja, P.K. Regulation of Monocarboxylate Transporter 1 (MCT1) Promoter by Butyrate in Human Intestinal Epithelial Cells: Involvement of NF-κB Pathway. J. Cell Biochem. 2008, 103, 1452–1463. [Google Scholar] [CrossRef]
- Kirat, D.; Kondo, K.; Shimada, R.; Kato, S. Dietary Pectin Up-Regulates Monocaboxylate Transporter 1 in the Rat Gastrointestinal Tract: Pectin Up-Regulates MCT1 in Rat Gastrointestinal Tract. Exp. Physiol. 2009, 94, 422–433. [Google Scholar] [CrossRef]
- Haenen, D.; Zhang, J.; Souza da Silva, C.; Bosch, G.; van der Meer, I.M.; van Arkel, J.; van den Borne, J.J.G.C.; Pérez Gutiérrez, O.; Smidt, H.; Kemp, B.; et al. A Diet High in Resistant Starch Modulates Microbiota Composition, SCFA Concentrations, and Gene Expression in Pig Intestine. J. Nutr. 2013, 143, 274–283. [Google Scholar] [CrossRef] [Green Version]
- Borthakur, A.; Priyamvada, S.; Kumar, A.; Natarajan, A.A.; Gill, R.K.; Alrefai, W.A.; Dudeja, P.K. A Novel Nutrient Sensing Mechanism Underlies Substrate-Induced Regulation of Monocarboxylate Transporter-1. Am. J. Physiol. Gastrointest. Liver Physiol. 2012, 303, G1126–G1133. [Google Scholar] [CrossRef] [Green Version]
- Suk, H.Y.; Zhou, C.; Yang, T.T.C.; Zhu, H.; Yu, R.Y.L.; Olabisi, O.; Yang, X.; Brancho, D.; Kim, J.-Y.; Scherer, P.E.; et al. Ablation of Calcineurin Aβ Reveals Hyperlipidemia and Signaling Cross-Talks with Phosphodiesterases. J. Biol. Chem. 2013, 288, 3477–3488. [Google Scholar] [CrossRef] [Green Version]
- Buyse, M.; Sitaraman, S.V.; Liu, X.; Bado, A.; Merlin, D. Luminal Leptin Enhances CD147/MCT-1-Mediated Uptake of Butyrate in the Human Intestinal Cell Line Caco2-BBE. J. Biol. Chem. 2002, 277, 28182–28190. [Google Scholar] [CrossRef] [Green Version]
- Thibault, R.; De Coppet, P.; Daly, K.; Bourreille, A.; Cuff, M.; Bonnet, C.; Mosnier, J.; Galmiche, J.; Shirazi–Beechey, S.; Segain, J. Down-Regulation of the Monocarboxylate Transporter 1 Is Involved in Butyrate Deficiency during Intestinal Inflammation. Gastroenterology 2007, 133, 1916–1927. [Google Scholar] [CrossRef]
- Villodre Tudela, C.; Boudry, C.; Stumpff, F.; Aschenbach, J.R.; Vahjen, W.; Zentek, J.; Pieper, R. Down-Regulation of Monocarboxylate Transporter 1 (MCT1) Gene Expression in the Colon of Piglets Is Linked to Bacterial Protein Fermentation and pro-Inflammatory Cytokine-Mediated Signalling. Br. J. Nutr. 2015, 113, 610–617. [Google Scholar] [CrossRef] [Green Version]
- De Preter, V.; Arijs, I.; Windey, K.; Vanhove, W.; Vermeire, S.; Schuit, F.; Rutgeerts, P.; Verbeke, K. Impaired Butyrate Oxidation in Ulcerative Colitis Is Due to Decreased Butyrate Uptake and a Defect in the Oxidation Pathway. Inflamm. Bowel Dis. 2012, 18, 1127–1136. [Google Scholar] [CrossRef]
- Ferrer-Picón, E.; Dotti, I.; Corraliza, A.M.; Mayorgas, A.; Esteller, M.; Perales, J.C.; Ricart, E.; Masamunt, M.C.; Carrasco, A.; Tristán, E.; et al. Intestinal Inflammation Modulates the Epithelial Response to Butyrate in Patients with Inflammatory Bowel Disease. Inflamm. Bowel Dis. 2020, 26, 43–55. [Google Scholar] [CrossRef] [Green Version]
- Paramsothy, S.; Nielsen, S.; Kamm, M.A.; Deshpande, N.P.; Faith, J.J.; Clemente, J.C.; Paramsothy, R.; Walsh, A.J.; van den Bogaerde, J.; Samuel, D.; et al. Specific Bacteria and Metabolites Associated with Response to Fecal Microbiota Transplantation in Patients with Ulcerative Colitis. Gastroenterology 2019, 156, 1440–1454.e2. [Google Scholar] [CrossRef]
- Daillère, R.; Vétizou, M.; Waldschmitt, N.; Yamazaki, T.; Isnard, C.; Poirier-Colame, V.; Duong, C.P.M.; Flament, C.; Lepage, P.; Roberti, M.P.; et al. Enterococcus Hirae and Barnesiella Intestinihominis Facilitate Cyclophosphamide-Induced Therapeutic Immunomodulatory Effects. Immunity 2016, 45, 931–943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Chu, C.-Q. Gut Microbiota–Medication Interaction in Rheumatic Diseases. Front. Immunol. 2021, 12, 796865. [Google Scholar] [CrossRef] [PubMed]
- Maier, L.; Pruteanu, M.; Kuhn, M.; Zeller, G.; Telzerow, A.; Anderson, E.E.; Brochado, A.R.; Fernandez, K.C.; Dose, H.; Mori, H.; et al. Extensive Impact of Non-Antibiotic Drugs on Human Gut Bacteria. Nature 2018, 555, 623–628. [Google Scholar] [CrossRef] [PubMed]
- Roediger, W.E. Utilization of Nutrients by Isolated Epithelial Cells of the Rat Colon. Gastroenterology 1982, 83, 424–429. [Google Scholar] [CrossRef]
- Thibault, R.; Blachier, F.; Darcy-Vrillon, B.; de Coppet, P.; Bourreille, A.; Segain, J.-P. Butyrate Utilization by the Colonic Mucosa in Inflammatory Bowel Diseases: A Transport Deficiency. Inflamm. Bowel Dis. 2010, 16, 684–695. [Google Scholar] [CrossRef]
- De Preter, V.; Geboes, K.P.; Bulteel, V.; Vandermeulen, G.; Suenaert, P.; Rutgeerts, P.; Verbeke, K. Kinetics of Butyrate Metabolism in the Normal Colon and in Ulcerative Colitis: The Effects of Substrate Concentration and Carnitine on the β-Oxidation Pathway: Kinetics of Butyrate Metabolism in Ulcerative Colitis. Aliment. Pharmacol. Ther. 2011, 34, 526–532. [Google Scholar] [CrossRef]
- Santhanam, S.; Venkatraman, A.; Ramakrishna, B.S. Impairment of Mitochondrial Acetoacetyl CoA Thiolase Activity in the Colonic Mucosa of Patients with Ulcerative Colitis. Gut 2007, 56, 1543–1549. [Google Scholar] [CrossRef] [Green Version]
- Michielan, A.; D’Incà, R. Intestinal Permeability in Inflammatory Bowel Disease: Pathogenesis, Clinical Evaluation, and Therapy of Leaky Gut. Mediat. Inflamm. 2015, 2015, 628157. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Chen, L.; Zhou, R.; Wang, X.; Song, L.; Huang, S.; Wang, G.; Xia, B. Increased Proportions of Bifidobacterium and the Lactobacillus Group and Loss of Butyrate-Producing Bacteria in Inflammatory Bowel Disease. J. Clin. Microbiol. 2014, 52, 398–406. [Google Scholar] [CrossRef] [Green Version]
- Ota, S.; Sakuraba, H.; Hiraga, H.; Yoshida, S.; Satake, M.; Akemoto, Y.; Tanaka, N.; Watanabe, R.; Takato, M.; Murai, Y.; et al. Cyclosporine Protects from Intestinal Epithelial Injury by Modulating Butyrate Uptake via Upregulation of Membrane Monocarboxylate Transporter 1 Levels. Biochem. Biophys. Rep. 2020, 24, 100811. [Google Scholar] [CrossRef]
- Kennedy, G.M.; Min, M.Y.; Fitzgerald, J.F.; Nguyen, M.T.; Schultz, S.L.; Crum, M.T.; Starke, J.A.; Butkus, M.A.; Bowman, D.D.; Labare, M.P. Inactivation of the Bacterial Pathogens Staphylococcus Pseudintermedius and Acinetobacter Baumannii by Butanoic Acid. J. Appl. Microbiol. 2019, 126, 752–763. [Google Scholar] [CrossRef]
- Schauber, J. Expression of the Cathelicidin LL-37 Is Modulated by Short Chain Fatty Acids in Colonocytes: Relevance of Signalling Pathways. Gut 2003, 52, 735–741. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Chen, N.; Wu, Z.; Song, Y.; Zhang, Y.; Wu, N.; Zhang, F.; Ren, X.; Liu, Y. 5-Aminosalicylic Acid Alters the Gut Bacterial Microbiota in Patients with Ulcerative Colitis. Front. Microbiol. 2018, 9, 1274. [Google Scholar] [CrossRef] [PubMed]
- Sanchis-Artero, L.; Martínez-Blanch, J.F.; Manresa-Vera, S.; Cortés-Castell, E.; Valls-Gandia, M.; Iborra, M.; Paredes-Arquiola, J.M.; Boscá-Watts, M.; Huguet, J.M.; Gil-Borrás, R.; et al. Evaluation of Changes in Intestinal Microbiota in Crohn’s Disease Patients after Anti-TNF Alpha Treatment. Sci. Rep. 2021, 11, 10016. [Google Scholar] [CrossRef]
- Sossai, P. Butyric Acid: What Is the Future for This Old Substance? Swiss Med. Wkly 2012, 42, w13596. [Google Scholar] [CrossRef]
- Facchin, S.; Vitulo, N.; Calgaro, M.; Buda, A.; Romualdi, C.; Pohl, D.; Perini, B.; Lorenzon, G.; Marinelli, C.; D’Incà, R.; et al. Microbiota Changes Induced by Microencapsulated Sodium Butyrate in Patients with Inflammatory Bowel Disease. Neurogastroenterol. Motil. 2020, 32, e13914. [Google Scholar] [CrossRef]
- Edelman, M.J.; Bauer, K.; Khanwani, S.; Tait, N.; Trepel, J.; Karp, J.; Nemieboka, N.; Chung, E.-J.; Van Echo, D. Clinical and Pharmacologic Study of Tributyrin: An Oral Butyrate Prodrug. Cancer Chemother. Pharm. 2003, 51, 439–444. [Google Scholar] [CrossRef]
- Gill, P.A.; van Zelm, M.C.; Muir, J.G.; Gibson, P.R. Review Article: Short Chain Fatty Acids as Potential Therapeutic Agents in Human Gastrointestinal and Inflammatory Disorders. Aliment. Pharm. Ther. 2018, 48, 15–34. [Google Scholar] [CrossRef] [Green Version]
- Clarke, J.M.; Topping, D.L.; Christophersen, C.T.; Bird, A.R.; Lange, K.; Saunders, I.; Cobiac, L. Butyrate Esterified to Starch Is Released in the Human Gastrointestinal Tract. Am. J. Clin. Nutr. 2011, 94, 1276–1283. [Google Scholar] [CrossRef] [Green Version]
- Clarke, J.M.; Bird, A.R.; Topping, D.L.; Cobiac, L. Excretion of Starch and Esterified Short-Chain Fatty Acids by Ileostomy Subjects after the Ingestion of Acylated Starches. Am. J. Clin. Nutr. 2007, 86, 1146–1151. [Google Scholar] [CrossRef]
- Nielsen, T.; Canibe, N.; Larsen, F. Butyrylation of Maize and Potato Starches and Characterization of the Products by Nuclear Magnetic Resonance and In Vitro Fermentation. Foods 2018, 7, 79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Cheng, L.; Li, Z.; Li, C.; Hong, Y.; Gu, Z. Butyrylated Starch Protects Mice from DSS-Induced Colitis: Combined Effects of Butyrate Release and Prebiotic Supply. Food Funct. 2021, 12, 11290–11302. [Google Scholar] [CrossRef] [PubMed]
- Mu, Y.; Kinashi, Y.; Li, J.; Yoshikawa, T.; Kishimura, A.; Tanaka, M.; Matsui, T.; Mori, T.; Hase, K.; Katayama, Y. Polyvinyl Butyrate Nanoparticles as Butyrate Donors for Colitis Treatment. ACS Appl. Bio Mater. 2021, 4, 2335–2341. [Google Scholar] [CrossRef]
- Henn, M.R.; O’Brien, E.J.; Diao, L.; Feagan, B.G.; Sandborn, W.J.; Huttenhower, C.; Wortman, J.R.; McGovern, B.H.; Wang-Weigand, S.; Lichter, D.I.; et al. A Phase 1b Safety Study of SER-287, a Spore-Based Microbiome Therapeutic, for Active Mild to Moderate Ulcerative Colitis. Gastroenterology 2021, 160, 115–127.e30. [Google Scholar] [CrossRef]
- Vernero, M.; De Blasio, F.; Ribaldone, D.G.; Bugianesi, E.; Pellicano, R.; Saracco, G.M.; Astegiano, M.; Caviglia, G.P. The Usefulness of Microencapsulated Sodium Butyrate Add-On Therapy in Maintaining Remission in Patients with Ulcerative Colitis: A Prospective Observational Study. JCM 2020, 9, 3941. [Google Scholar] [CrossRef]
- Vancamelbeke, M.; Laeremans, T.; Vanhove, W.; Arnauts, K.; Ramalho, A.S.; Farré, R.; Cleynen, I.; Ferrante, M.; Vermeire, S. Butyrate Does Not Protect Against Inflammation-Induced Loss of Epithelial Barrier Function and Cytokine Production in Primary Cell Monolayers from Patients with Ulcerative Colitis. J. Crohns Colitis 2019, 13, 1351–1361. [Google Scholar] [CrossRef] [Green Version]
- Kaiko, G.E.; Ryu, S.H.; Koues, O.I.; Collins, P.L.; Solnica-Krezel, L.; Pearce, E.J.; Pearce, E.L.; Oltz, E.M.; Stappenbeck, T.S. The Colonic Crypt Protects Stem Cells from Microbiota-Derived Metabolites. Cell 2016, 165, 1708–1720. [Google Scholar] [CrossRef] [Green Version]
- Peng, L.; He, Z.; Chen, W.; Holzman, I.R.; Lin, J. Effects of Butyrate on Intestinal Barrier Function in a Caco-2 Cell Monolayer Model of Intestinal Barrier. Pediatr. Res. 2007, 61, 37–41. [Google Scholar] [CrossRef] [Green Version]
- Perman, J.A.; Modler, S.; Olson, A.C. Role of PH in Production of Hydrogen from Carbohydrates by Colonic Bacterial Flora. Studies In Vivo and In Vitro. J. Clin. Investig. 1981, 67, 643–650. [Google Scholar] [CrossRef]
- Breuer, R.I.; Buto, S.K.; Christ, M.L.; Bean, J.; Vernia, P.; Paoluzi, P.; Di Paolo, M.C.; Caprilli, R. Rectal Irrigation with Short-Chain Fatty Acids for Distal Ulcerative Colitis: Preliminary Report. Dig. Dis. Sci. 1991, 36, 185–187. [Google Scholar] [CrossRef]
- Scheppach, W.; Sommer, H.; Kirchner, T.; Paganelli, G.-M.; Bartram, P.; Christl, S.; Richter, F.; Dusel, G.; Kasper, H. Effect of Butyrate Enemas on the Colonic Mucosa in Distal Ulcerative Colitis. Gastroenterology 1992, 103, 51–56. [Google Scholar] [CrossRef]
- Scheppach, W.; German-Austrian Scfa Study Group. Treatment of Distal Ulcerative Colitis with Short-Chain Fatty Acid Enemas a Placebo-Controlled Trial. Dig. Dis. Sci. 1996, 41, 2254–2259. [Google Scholar] [CrossRef] [PubMed]
- Vernia, P.; Marcheggiano, A.; Caprilli, R.; Frieri, G.; Corrao, G.; Valpiani, D.; Paolo, M.C.D.; Paoluzi, P.; Torsoli, A. Short-Chain Fatty Acid Topical Treatment in Distal Ulcerative Colitis. Aliment. Pharmacol. Ther. 2007, 9, 309–313. [Google Scholar] [CrossRef]
- Steinhart, A.H.; Hiruki, T.; Brzezinski, A.; Baker, J.P. Treatment of Left-Sided Ulcerative Colitis with Butyrate Enemas: A Controlled Trial. Aliment. Pharmacol. Ther. 1996, 10, 729–736. [Google Scholar] [CrossRef]
- Breuer, R.I.; Soergel, K.H.; Lashner, B.A.; Christ, M.L.; Hanauer, S.B.; Vanagunas, A.; Harig, J.M.; Keshavarzian, A.; Robinson, M.; Sellin, J.H.; et al. Short Chain Fatty Acid Rectal Irrigation for Left-Sided Ulcerative Colitis: A Randomised, Placebo Controlled Trial. Gut 1997, 40, 485–491. [Google Scholar] [CrossRef]
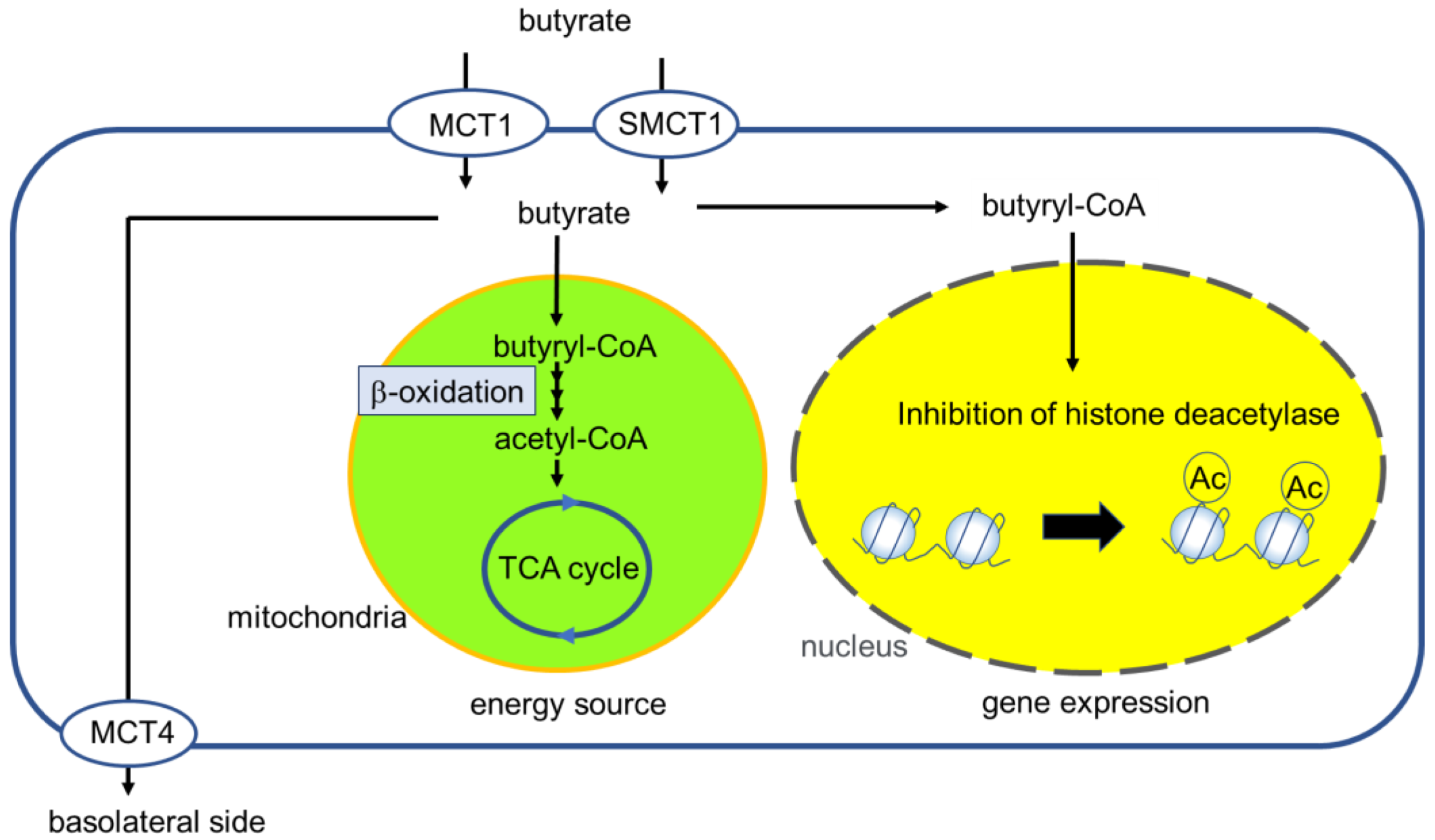
Figure 1.
Overview of butyrate uptake, dynamics, and physiological effects in intestinal epithelial cells (IECs). Butyrate fermented by microbiota is transported to IEC by monocarboxylate transporter 1 (MCT1) and sodium-coupled monocarboxylate transporter 1 (SMCT1). By β-oxidization in the mitochondria, butyrate is metabolized to acetyl-CoA. Unmetabolized butyrate inhibits histone deacetylase and is transported into the lamina propria.
Figure 1.
Overview of butyrate uptake, dynamics, and physiological effects in intestinal epithelial cells (IECs). Butyrate fermented by microbiota is transported to IEC by monocarboxylate transporter 1 (MCT1) and sodium-coupled monocarboxylate transporter 1 (SMCT1). By β-oxidization in the mitochondria, butyrate is metabolized to acetyl-CoA. Unmetabolized butyrate inhibits histone deacetylase and is transported into the lamina propria.

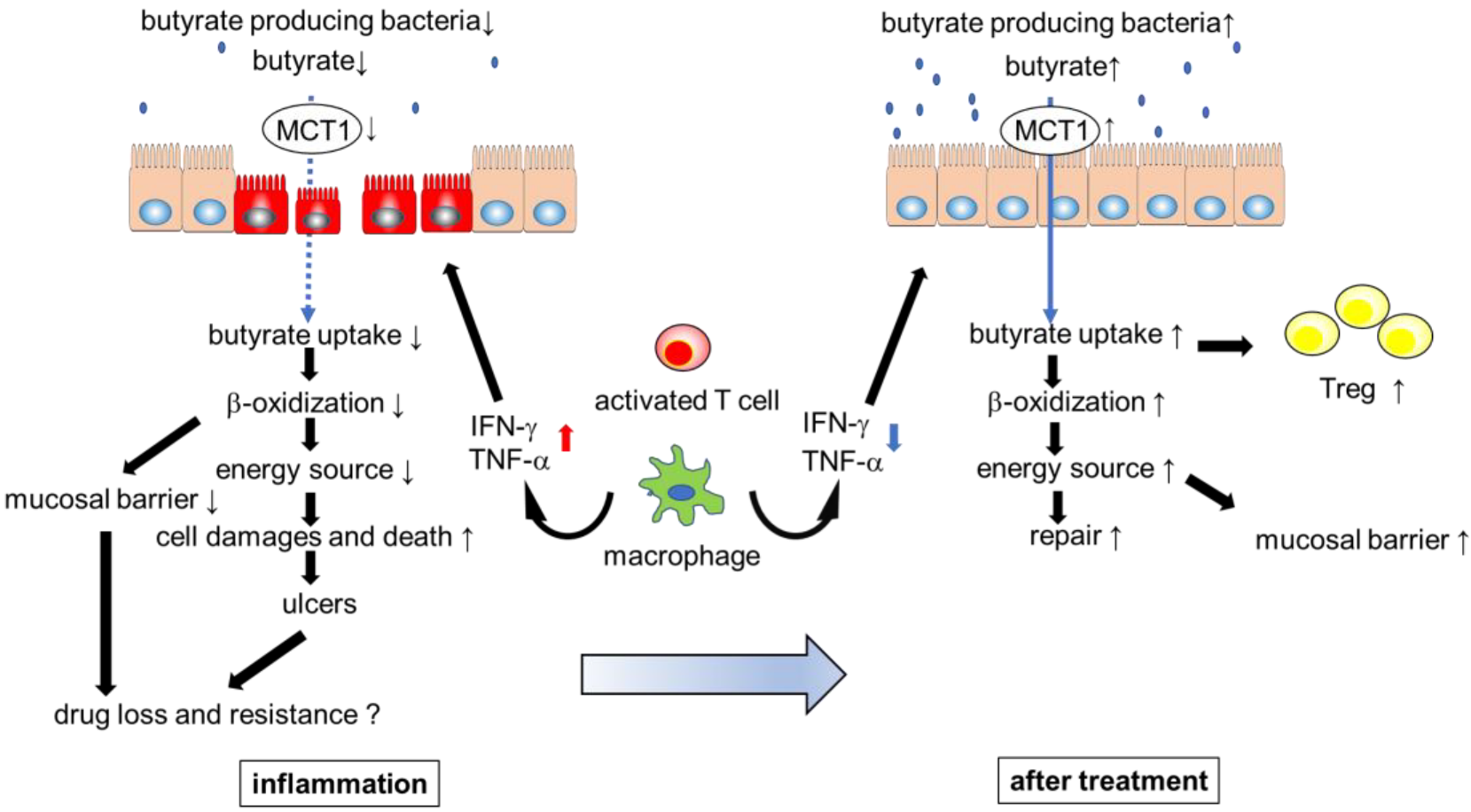
Figure 2.
Inflammatory cytokines impair butyrate uptake, barrier function, and metabolism in intestinal epithelial cells (IECs). Inflammation and low abundance of butyrate contribute to the refractory nature of inflammatory bowel disease. Appropriate treatment ameliorates the inflammation that leads to dysbiosis and decreased butyrate uptake. As a result, increased butyrate levels in the colon contribute to gut homeostasis and drug efficacy.
Figure 2.
Inflammatory cytokines impair butyrate uptake, barrier function, and metabolism in intestinal epithelial cells (IECs). Inflammation and low abundance of butyrate contribute to the refractory nature of inflammatory bowel disease. Appropriate treatment ameliorates the inflammation that leads to dysbiosis and decreased butyrate uptake. As a result, increased butyrate levels in the colon contribute to gut homeostasis and drug efficacy.


{kind=link}
{kind=link}
Table 1.
Effects of butyrate and butyrate-producing bacteria on therapeutic efficacy of drugs.
| Drug | Patients | Outcome | Ref. |
|---|---|---|---|
| anti-TNF-α | 56 UC (50 IFX, 6 ADA) | Lower dysbiosis indexes and higher abundance of Faecalibacterium prausnitzii predict responders | [10] |
| vedolizumab | 43 UC, 42 CD | Butyrate producer promote response of vedolizmab treatment | [7] |
| anti-TNF-α | 18 CD, 17 UC | In low response patients, lower exchanged among bacterial communities of butyrate, ethanol or acetaldehyde (involved in butyrate synthesis) | [9] |
| azathioprine | 43 CD, 22 UC | Butyrate production of patients in remission is at baseline higher compared with patients without remission | [8] |
Table 2.
Clinical trial results for topical butyrate treatment.
| Disease | Sample Size | Treatment | Dose, Duration | Outcome | Ref. |
|---|---|---|---|---|---|
| distal UC | 12 | SCFAs (80:30:40) | 100 mL, twice daily, 6 weeks | 9 patients improved disease activity index and mucosal histrogical scores | [80] |
| 10 | 100 mM butyrate vs. placebo | 100 mL, twice daily, 2 weeks | Improving endoscopic and histrogical scores | [81] | |
| 47 | 130 mM SCFAs (46:23:31) or 100 mM butyrate alone vs. saline | 60 mL, twice daily, 8 weeks | No differences in disease activity, clinical response, endoscopic, histological scores | [82] | |
| 40 | 150 mM SCFAs (53:20:27) vs. saline | 100 mL, twice daily, 6 weeks | Improving intestinal bleeding, urgency, the patient self-evaluation scores | [83] | |
| 38 | 80 mM butyrate vs. saline | 60 mL, once nightly, 6 weeks | No difference in clinical disease activity index scores | [84] | |
| 103 | 150 mM SCFAs (53:20:27) vs. saline | 100 mL, twice daily, 6 weeks | No difference in clinical and histological activity scores | [85] |
Ratio = acetate:propionate:butyrate.
Table 3.
Research showing negative effects of butyrate on barrier function and cytoprotection.
| Model | Result | Mechanism | Ref. |
|---|---|---|---|
| DSS induced colitis mice with antibiotics pretreatment | Not ameliorate colitis | Decreased IL-6 production by butyrate treatment inhibit intestinal tissue repair and cytoprotection | [12] |
| Caco-2 cells | Low concentration; intestinal barrier ↑ | Excessive butyrate induces epithelial cell apoptosis | [78] |
| Excessive concentration; intestinal barrier ↓ | |||
| Primary intestinal epithelial monolayer cultures from UC patients + TNF-α and IFN-γ | Intestinal barrier ↓, IL-8 mRNA ↑ many inflammatory protein ↑ | unknown (There is a possibility that butyrate suppresses the expansion of proliferating cells under inflammatory mediators) | [76] |
| Mouse and human colonic and small intestinal crypts | Suppressing colonic epithelial stem/progenitor cell proliferation, delayed wound repair | By HDAC inhibition, promoter activity for the negative cell-cycle regulator Foxo3 was increased | [77] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ota, S.; Sakuraba, H. Uptake and Advanced Therapy of Butyrate in Inflammatory Bowel Disease. Immuno 2022, 2, 692-702. https://doi.org/10.3390/immuno2040042
AMA Style
Ota S, Sakuraba H. Uptake and Advanced Therapy of Butyrate in Inflammatory Bowel Disease. Immuno. 2022; 2(4):692-702. https://doi.org/10.3390/immuno2040042
Chicago/Turabian StyleOta, Shinji, and Hirotake Sakuraba. 2022. "Uptake and Advanced Therapy of Butyrate in Inflammatory Bowel Disease" Immuno 2, no. 4: 692-702. https://doi.org/10.3390/immuno2040042