
The Influence of Eurasian Beaver (Castor fiber L.) Activity on the Transformation and Functioning of Riparian Phytocoenoses in the Southern Boreal Zone (European Russia)


 and
and 
Abstract
:1. Introduction
2. Materials and Methods
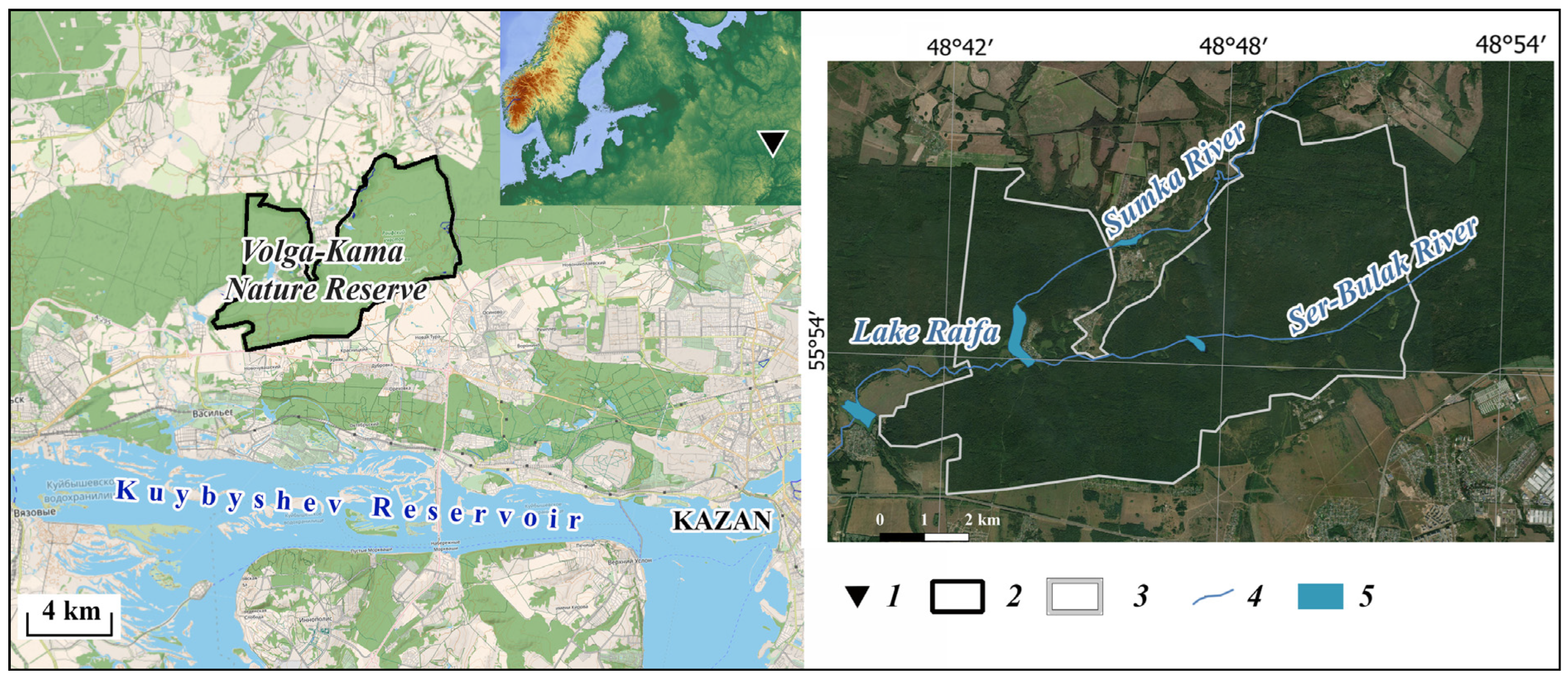
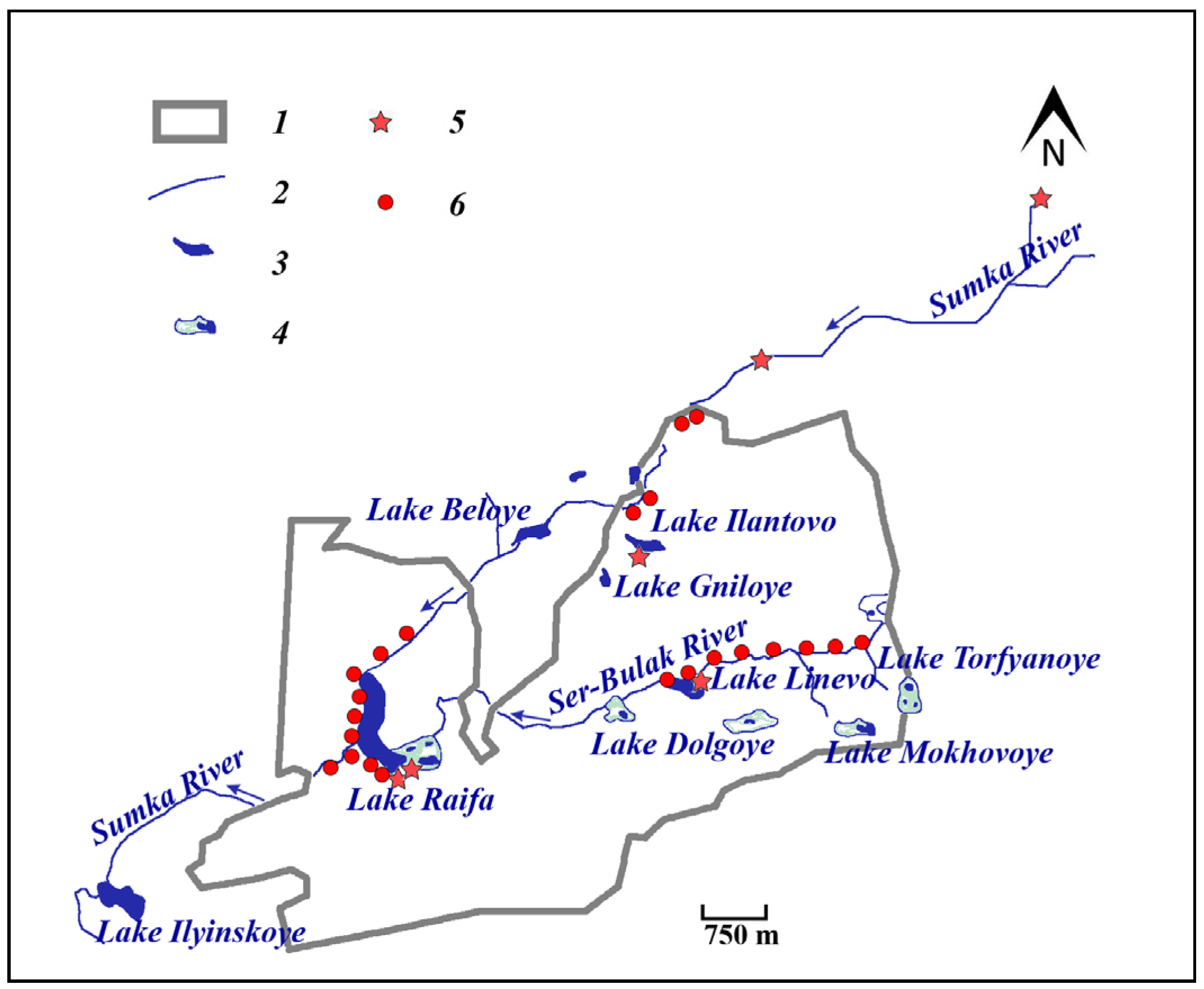
2.1. Study Area
2.2. Reintroduction of Castor fiber L.
2.3. Materials
2.4. Methods
3. Results and Discussion
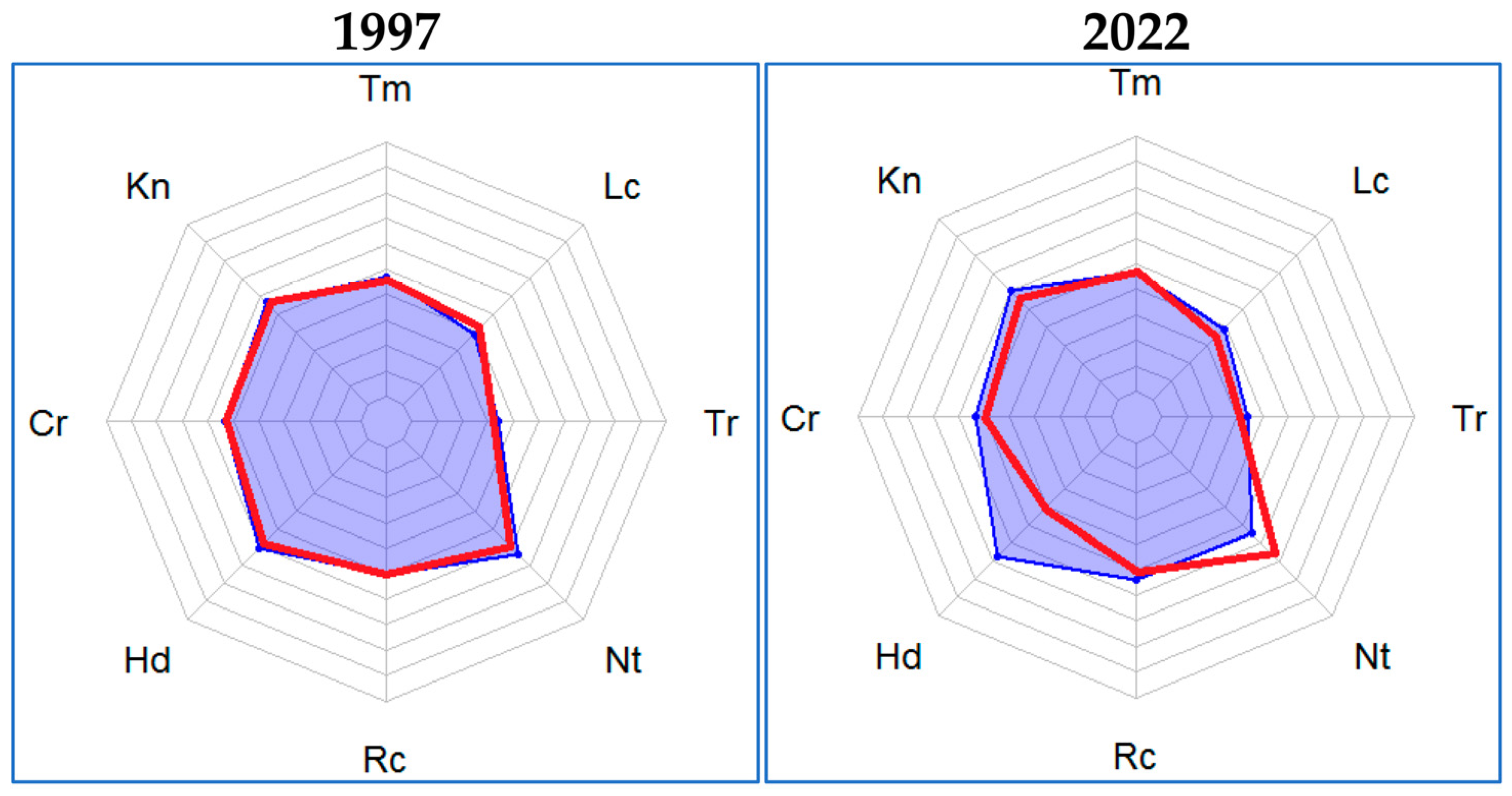
- The air temperature regime (Tm) refers to the type of boreal-nemoral zones with an annual value of total solar radiation of 40 kcal/cm2;
- Continental climate (Kn), semi-arid, according to the ombroclimatic factor;
- The cryoclimatic factor (Cr) was characterized by moderate winters (average air temperatures of the coldest month were from −8 to −16 °C);
- Conditions of humid forest-meadow zones according to the moisture factor (Hd);
- Poor soils in terms of the salt regime (Tr);
- Nitrogen-poor soils under conditions of nitrogen richness (Nt);
- Slightly acidic soils (pH = 5.5–6.5) according to the soil acidity regime (Rc) (for light forest conditions–Lc, for a zone of relatively stable moisture–Bk [25]).
4. Limitations
5. Conclusions
- The activity of C. fiber does not affect the functioning of riparian phytocoenoses associated with such more global factors as climate continentality, air temperature regime, ombroclimatism, and cryoclimatism. Statistically significant changes in microclimatic factors of the environment under the influence of the activity of C. fiber in the Raifa forest sector of the Volga-Kama State Nature Biosphere Reserve were revealed. The influence of the beaver reintroduction factor on the change in the regime of moistening and illumination of habitats, the richness of soils with nitrogen, and the acidity and salt regime of soils was also revealed.
- Under the conditions of fodder and construction activities of the beaver, an increase in the proportion of groups of aquatic, wetland, and boreal plant species, as well as an expansive nature of the change in their activity, were noted. A decrease in the activity of distribution of ruderal species under the conditions of beaver activity was noted.
- Changes in factors such as humidity and illumination of the habitat are most likely directly related to the activity and lifestyle of the beaver. This is due to the direct removal of trees from the forest stand, which increases the illumination of riparian zones.
- Beaver-induced flooding and underflooding result in an increase in moisture of riparian soils and grounds in quantities sufficient to cause the replacement of some ecological–coenotic groups by others that are more adapted to changing conditions.
- From the obtained results, it follows that in the areas affected by C. fiber, the effect of adverse climatic phenomena, such as drought, is smoothed out, and the most optimal conditions for the conservation and development of boreal plant species are provided.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Halley, D.J.; Saveljev, A.P.; Rosell, F. Population and distribution of beavers Castor fiber and Castor canadensis in Eurasia. Mammal Rev. 2021, 51, 1–24. [Google Scholar] [CrossRef]
- Dezhkin, V.V.; D’yakov, Y.V.; Safonov, V.G. Beaver; Agroprom Publisher: Moscow, Russia, 1986. (In Russian) [Google Scholar]
- Balodis, M.M. Biological Bases of Beaver Farming (on the Example of Latvia). Ph.D. Thesis, A.N. Severtsov Institute of Ecology and Evolution of the Russian Academy of Sciences, Moscow, Russia, 1989. (In Russian). [Google Scholar]
- Abaturov, B.D. Mammals as a Component of the Ecosystems; Nauka Publisher: Moscow, Russia, 1984. (In Russian) [Google Scholar]
- Bratton, S.P. The effect of the European wild boar, Sus scrofa, on gray beech forest in the Great Smokey Mountains. Ecology 1975, 56, 1356–1366. [Google Scholar] [CrossRef]
- McNaughton, S.J. Ecology of a grazing ecosystem: The Serengheti. Ecol. Monogr. 1985, 55, 259–294. [Google Scholar] [CrossRef]
- Huntley, N.J. Influence of refuging consumers (pikas: Ochotona princeps) on subalpine meadow vegetation. Ecology 1987, 68, 274–283. [Google Scholar] [CrossRef]
- Johnston, C.A.; Naiman, R.J. Boundary dynamics at the aquatic-terrestrial interface: The influence of beaver and geomorphology. Landsc. Ecol. 1987, 1, 47–57. [Google Scholar] [CrossRef]
- Gorshkov, D.Y.; Gorshkov, Y.A.; Kochetkov, N.V. The features of the feeding behavior of the beavers in Volga-Kama National Nature Zapovednik. In Proceedings of the First Euro-American Beaver Congress, 24–28 August 1999; Busher, P.E., Gorshkov, Y., Eds.; Kluwer Academic/Plenum Publishers: New York, NY, USA, 1999; pp. 14–16. [Google Scholar]
- Zavyalov, H.A.; Krylov, A.V.; Bobrov, A.A.; Ivanov, V.K.; Dgebuadze, Y.Y. Effects of River Beaver on the Ecosystems of Small Rivers; Nauka Publisher: Moscow, Russia, 2005. (In Russian) [Google Scholar]
- Dezhkin, V. Beaver in the modern world: Necessity of the population management on the national and the international level. In Proceedings of the First Euro-American Beaver Congress, 24–28 August 1999; Busher, P.E., Gorshkov, Y., Eds.; Kluwer Academic/Plenum Publishers: New York, NY, USA, 1999; pp. 20–26. [Google Scholar]
- Popov, V.A. Mammals of the Volga-Kama Region; Kazan Branch of the Academy of Sciences of the USSR Publisher: Kazan, Russia, 1960. (In Russian) [Google Scholar]
- Grigoriev, N.D. The current state of beaver colonies in the Volga-Kama region, growth prospects and using. Proc. Voronezhsky State Reserve 1969, 16, 86–99. (In Russian) [Google Scholar]
- Dgebuadze, Y.Y. Ecology of invasions and population contacts of animals: General approaches. In Invasive Species in the European Seas of Russia; Matishev, G.G., Ed.; Kola Scientific Center of the Russian Academy of Sciences Publisher: Apatity, Russia, 2000; pp. 35–50. (In Russian) [Google Scholar]
- Great Volzhsko-Kamsky Biosphere Reserve, According to UNESCO. Available online: https://en.unesco.org/biosphere/eu-na/great-volzhsko-kamsky (accessed on 31 March 2023).
- Porfiriev, V.S. Vegetation of Raifa. Proc. Volga-Kama Nat. Reserve 1968, 1, 17–23. (In Russian) [Google Scholar]
- Taisin, A.S. Raifa Forest in the Boreal Forests of Eurasia; Kazan University Publisher: Kazan, Russia, 2008. (In Russian) [Google Scholar]
- Legeyda, I.S. Environment-Forming Activity of Beavers and Protection of Coastal Biogeocenoses of Ukraine. Ph.D. Thesis, Institute of Ecology and Evolution of the Russian Academy of Sciences, Moscow, Russia, 1992. (In Russian). [Google Scholar]
- Lisetskii, F.N.; Poletaev, A.O.; Buryak, Z.A. Geoinformation support for studies of the boundaries of flood zones in urban areas. J. Phys. Conf. Ser. 2022, 2388, 012134. [Google Scholar] [CrossRef]
- Taisin, A.S.; Dedkov, A.P. Pliocene valleys and Quaternary terraces of the Raifa. Proc. Volga-Kama Nat. Reserve 2005, 6, 115–127. (In Russian) [Google Scholar]
- Gorshkov, Y.A.; Gorshkov, D.Y.; Easter-Pilcher, A.L.; Pilcher, B.K. First results of beaver (Castor fiber) reintroduction in Volga-Kama National Nature Zapovednik (Russia). Folia Zool. 2002, 51, 67–74. [Google Scholar]
- Easter-Pilcher, A.L. Forage Utilization, Habitat Selection and Population Indices of Beaver in Northwest Montana. Master’s Thesis, University of Montana, Missoula, MT, USA, 1987. [Google Scholar]
- Drude, O. Handbuch der Pflanzengeographie; J. Engelhorn: Stuttgart, Germany, 1890; 582p. (In German) [Google Scholar]
- Prokhorov, V.E.; Rogova, T.V.; Kozhevnikova, M.V. Vegetation Database of Tatarstan. Pytocoenologia 2017, 47, 309–313. [Google Scholar] [CrossRef]
- Tsyganov, D.N. Phytoindication of Ecological Regimes in the Subzone of Coniferous-Deciduous Forests; Nauka Publisher: Moscow, Russia, 1983. (In Russian) [Google Scholar]
- Ellenberg, H. Zeigerwerte der Gefässpflanzen Mitteleuropas. Scr. Geobot. 1974, 9, 1–166. (In German) [Google Scholar]
- Landolt, E. Okologische Zeigerwerts zur Sweizer Flora. In Veröffentlichungen des Geobotanischen Institutes der ETH; Stiftung Rübel: Zürich, Switzerland, 1977; Volume 64, 208p. (In German) [Google Scholar]
- Ramensky, L.G.; Tsatsenkin, I.A.; Chizhikov, O.N.; Antipin, N.A. Ecological Assessment of Fodder Lands by Vegetation Cover; Sel’khozhiz: Moscow, Russia, 1956; 472p. (In Russian) [Google Scholar]
- Egorova, N.Y. Influence of Ecological Factors on the Population-Ontogenetic Parameters of Vaccinium vitis-idaea L. in Forest Ecosystems of the European Northeast of Russia. Contemp. Probl. Ecol. 2020, 13, 656–662. [Google Scholar] [CrossRef]
- Hellegers, M.; Ozinga, W.A.; Hinsberg, A.; Huijbregts, M.A.J.; Hennekens, S.M.; Schaminée, J.H.J.; Dengler, J.; Schipper, A.M. Evaluating the ecological realism of plant species distribution models with ecological indicator values. Ecography 2020, 43, 161–170. [Google Scholar] [CrossRef]
- Fyodorov, F.; Yakimova, A. Changes in Ecosystems of the Middle Taiga due to the Impact of Beaver Activities, Karelia, Russia. Balt. For. 2012, 18, 278–287. [Google Scholar]
- Danilov, P.I.; Fyodorov, F.V. The history and legacy of reintroduction of beaver in the European North of Russia. Russ. J. Theriol. 2016, 15, 43–48. [Google Scholar] [CrossRef]
- Law, A.; Jones, K.C.; Willby, N.J. Medium vs. short- term effects of herbivory by Eurasian beaver on aquatic vegetation. Aquat. Bot. 2014, 116, 27–34. [Google Scholar] [CrossRef]
- Elmeros, M.; Madsen, A.B.; Berthelsen, J.P. Monitoring of reintroduced beavers (Castor fiber) in Denmark. Lutra 2003, 46, 153–162. [Google Scholar]
- Donkor, N.T.; Fryxell, J.M. Lowland boreal forests characterization in Algonquin Provincial Park relative to beaver (Castor canadensis) foraging and edaphic factors. Plant Ecol. 2000, 148, 1–12. [Google Scholar] [CrossRef]
- Porokhov, A.A. Reacclimatization and Biocenotic Role of River Beavers Castor fiber L. in the Territory of the Priilmenskaya Lowland. Ph.D. Thesis, Russian Research Institute of Game Management and Fur Farming, Kirov, Russia, 1998. (In Russian). [Google Scholar]
- Pollock, M.M.; Beechie, T.; Wheaton, J.; Jordan, C.; Bouwes, N.; Weber, N.; Volk, C. Using beaver dams to restore incised stream ecosystems. BioScience 2014, 64, 279–290. [Google Scholar] [CrossRef]
- Sjoberg, G.; Ball, J.P. (Eds.) Restoring the European Beaver: 50 Years of Experience; Pensoft Publisher: Sofia, Bulgaria, 2011. [Google Scholar]
- McMaster, R.T.; McMaster, N.D. Composition, structure, and dynamics of vegetation in fifteen beaver-impacted wetlands in Western Massachusetts. Rhodora 2001, 103, 293–320. [Google Scholar]
- Müller-Schwarze, D.; Sun, L. The Beaver. Natural History of a Wetlands Engineer; Cornell University Press: New York, NY, USA, 2003. [Google Scholar]
- Rosell, F.; Bozser, O.; Collen, P.; Parker, H. Ecological impact of beavers Castor fiber and Castor canadensis and their ability to modify ecosystems. Mammal Rev. 2005, 35, 248–276. [Google Scholar] [CrossRef]
- Simonavičiūtė, L.; Ulevičius, A. Structure of phytocenoses in beaver meadows in Lithuania. Ekologija 2007, 53, 34–44. [Google Scholar]
- Bonner, J.; Anderson, J.; Rentch, J.; Grafton, W. Vegetative composition and community structure associated with beaver ponds in Canaan valley, West Virginia, USA. Wetl. Ecol. Manag. 2009, 17, 543–554. [Google Scholar] [CrossRef]
- Odum, E.P. Fundamentals of Ecology, 3rd ed.; W.B. Saunders Publisher: Philadelphia, PA, USA, 1971. [Google Scholar]
- Aleinikov, A.A. Castor fiber as an ecosystem engineer in «Bryanskiy Les» reserve and its protective zone. Bull. Tver State Univ. Ser. Biol. Ecol. 2010, 18, 60–68. (In Russian) [Google Scholar]
- Zavyalov, N.A. Dynamics of the Number and Environment-Forming Activity of the Beaver in the Darwin Reserve. Ph.D. Thesis, Institute of Forestry of Russian Academy of Sciences, Moscow, Russia, 1999. (In Russian). [Google Scholar]
- Albov, S.A.; Andreeva, M.V.; Bashinskiy, I.V.; Golubkov, V.V.; Katsman, E.A.; Goriaynova, Z.I.; Krylov, A.V.; Onipchenko, V.G.; Prokin, A.A.; Khlyap, L.A. European Beaver (Castor fiber L.) as a Key Species of a Small River Ecosystem (Prioksko-Terrasnyi Nature Biosphere Reserve); Association of Scientific Publications Publisher: Moscow, Russia, 2012. (In Russian) [Google Scholar]
- Oliger, T.I. Environment-forming role of the beaver in the Nizhnesvirsky Reserve. In Proceedings of the Preservation and Study of Geo- and Biodiversity in the Protected Areas of the European North of Russia, Izhevsk, Russia, 2–5 September 2014; pp. 166–170. (In Russian). [Google Scholar]
- Ustabaeva, E.V. Regional Ecological Features of the River Beaver Population with an Assessment of Its Impact on Steppe Bio-Coenoses of Orenburg Oblast. Ph.D. Thesis, Russian State Agrarian University named after Timiryazev, Moscow, Russia, 2013. (In Russian). [Google Scholar]




 control test plots,
control test plots,  experimental test plots.
control test plots, experimental test plots.
experimental test plots.
control test plots, experimental test plots.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Group | Ecological–Coenotic Group, % | ||||
|---|---|---|---|---|---|---|
| Forest | Meadow | Wetland | Steppe | Ruderal | ||
| 1997 | Control | 51.2 | 37.7 | 6.7 | 0 | 4.4 |
| Experimental | 46.0 | 27.0 | 10.2 | 3.3 | 13.5 | |
| 2010 | Control | 47.7 | 30.6 | 12.7 | 1.6 | 7.4 |
| Experimental | 44.5 | 19.4 | 25.7 | 1.7 | 8.7 | |
| 2022 | Control | 63.2 | 21.0 | 0 | 0 | 15.8 |
| Experimental | 46.3 | 23.1 | 28.2 | 0 | 2.4 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nazarov, N.G.; Prokhorov, V.E.; Sharifullin, A.G.; Gusarov, A.V.; Lisetskii, F.N. The Influence of Eurasian Beaver (Castor fiber L.) Activity on the Transformation and Functioning of Riparian Phytocoenoses in the Southern Boreal Zone (European Russia). Earth 2023, 4, 384-397. https://doi.org/10.3390/earth4020021
Nazarov NG, Prokhorov VE, Sharifullin AG, Gusarov AV, Lisetskii FN. The Influence of Eurasian Beaver (Castor fiber L.) Activity on the Transformation and Functioning of Riparian Phytocoenoses in the Southern Boreal Zone (European Russia). Earth. 2023; 4(2):384-397. https://doi.org/10.3390/earth4020021
Chicago/Turabian StyleNazarov, Nail G., Vadim E. Prokhorov, Aidar G. Sharifullin, Artyom V. Gusarov, and Fedor N. Lisetskii. 2023. "The Influence of Eurasian Beaver (Castor fiber L.) Activity on the Transformation and Functioning of Riparian Phytocoenoses in the Southern Boreal Zone (European Russia)" Earth 4, no. 2: 384-397. https://doi.org/10.3390/earth4020021