

Effects of Ammonia on Juvenile Sunray Surf Clam (Mactra chinensis Philippi) in Laboratory Tests


1
Tianjin Fisheries Research Institute, Hexi District, Tianjin 300221, China
2
College of Fisheries and Life Science, Dalian Ocean University, Ganjingzi District, Dalian 116023, China
3
InnoTech Alberta, Hwy 16A & 75 Street, P.O. Box 4000, Vegreville, AB T9C 1T4, Canada
*
Authors to whom correspondence should be addressed.
†
This Paper is a Part of the Master Thesis of Yuanyuan Dai, Presented at Dalian Ocean University (China).
Pollutants 2023, 3(2), 232-242; https://doi.org/10.3390/pollutants3020017
Submission received: 29 December 2022
/
Revised: 16 February 2023
/
Accepted: 10 April 2023
/
Published: 14 April 2023
(This article belongs to the Special Issue Marine Pollutants - Volume 2)
Abstract
:The current study aimed to determine the acute and sub-chronic toxicity of ammonia to juvenile surf clams (Mactra chinensis Philippi). Acute toxicity tests were conducted with seven concentrations of ammonium chloride using a 96 h static-renewal approach. Sub-chronic ammonia exposure tests (20 d exposures) were conducted with 6 concentrations at 20 °C. The 96 h median lethal concentration (96 h LC50) was 11.1 (10.0; 12.0) mg/L total ammonia nitrogen (TAN) and 0.56 (0.50; 0.60) mg/L unionized ammonia (NH3). The relative growth rate was significantly reduced at concentrations higher than 1.6 mg/L TAN (0.075 mg/L NH3) in the 20 d tests. The estimated maximum acceptable toxicant concentration (MATC) based on the reduced growth of juvenile M. chinensis was between 0.8 and1.6 mg/L TAN (0.038–0.075 mg/L NH3). Histopathological changes were evaluated in the surviving clams after 20 days of exposure. Exposure to 14.1 mg/L TAN (0.661 mg/L NH3) resulted in changes in the mantle, foot and digestive diverticulum. We also examined the antioxidant enzyme activities of superoxide dismutase (SOD) and catalase (CAT) in 10 d and 20 d at 6 different levels (plus a control) of ammonia from 0.8 mg/L to 14.1 mg/L TAN. Ammonia exposure at 0.8 mg/L TAN (0.038 mg/L NH3) significantly affected SOD and CAT activities. The level of enzymic activity decreased with the increasing concentration of TAN. The results improved our understanding of oxidative damage under ammonia exposure and provided data for the aquaculture of sunray surf clams.
1. Introduction
Global mollusc production from aquaculture has recently increased to 17.7 million tonnes (USD 29.8 billion) [1]. The sunray surf clam (also called hen clam, Mactra chinensis Philippi) is an essential commercial bivalve mollusc living in the intertidal zone of the Asia-Pacific region [2]. This species is vital to producing seafood in China, Korea and Japan [3]. However, its supply often cannot fulfill the market demand, and its farming is limited because of the high mortality rate due to pollution [4].
Ammonia is a constituent of concern due to its high likelihood of causing toxicity to marine molluscs [5]. Elevated ammonia levels have been found to compromise the survival, growth and reproduction of various aquatic organisms [6]. High concentrations of ammonia can alter various biological processes of marine bivalves, such as affecting the metabolic pathways involved in oxidative stress [7]. However, marine water quality guidelines for ammonia are rarely established to protect seawater organisms, mainly due to a lack of data on the toxicity of ammonia to marine organisms [8].
Recently, the toxic effects of ammonia have received increasing attention because of regional excessive nitrogen discharge [9]. Two forms of ammonia exist in seawater: ionized ammonia (NH4+) and unionized ammonia (NH3) [10]. Generally, the sum of NH4+ and NH3 is expressed as the total ammonia nitrogen (TAN) [11]. The proportion of each form mainly depends on water temperature, ionic strength and pH, while NH3 presents a more toxic impact [12]. A recent review of ammonia toxicity to shrimp and crabs concluded that the mechanisms of ammonia toxicity are crucial for the aquaculture of crustaceans [13]. Therefore, it is essential to discuss the mechanism of ammonia detoxification and provide practical guidance for marine aquaculture [14].
The toxic effects of ammonia on marine bivalves vary among taxa. For example, the surf clam species Spisula solidissima is considered one of the more ammonia-sensitive marine species, with a 48 h lethal concentration (LC50) at 10.6 mg/L TAN and 0.53 mg/L NH3 at the larvae life stage [15]. Histological observations revealed that the gill tissue of Ruditapes philippinarum was loose with large vacuoles scattered under the exposure of 0.1 mg/L NH3 [16]. However, ammonia-induced toxic effects are often tested using standard toxicology model organisms, such as freshwater mussel fatmucket (Lampsilis siliquoidea) [17]. Despite their contribution to the regional economy, very little ecotoxicological information could be found for M. chinensis under ammonia stress.
Our objectives in this study are to determine the lethal and sublethal effects of ammonia on juvenile M. chinensis in laboratory tests. The results provide data for water quality control in aquaculture and assess the protectiveness of the existing threshold guidance. In addition, we aim to examine the antioxidant enzyme activities of superoxide dismutase (SOD) and catalase (CAT) in order to provide information on the detoxification mechanism of marine bivalves under environmental ammonia exposure.
2. Materials and Methods
2.1. Test Organisms
Parental Mactra chinensis Philippi was collected from the Jianshan natural scenic area of Zhuanghe in Dalian, China. Juvenile M. chinensis propagated at Zhuanghe marine shellfish farm (Dalian, China). Healthy juvenile clams of uniform size (averaging 1.00 ± 0.10 cm in shell length (SL) and 0.75 ± 0.04 cm in shell height) were selected for toxicity tests. Juvenile SL and height were measured using an electronic vernier caliper for 21 randomly chosen individuals.
2.2. Test Water and Water Chemistry
The control water was prepared with sand-filtered seawater from Heishijiao (Dalian, China). Several routine water chemistry parameters of the control water were periodically collected and characterized, including temperature (19.8 ± 0.5 °C), pH (8.22 ± 0.04), salinity (29.6 ± 0.4 psu), dissolved oxygen (DO, 7.0 ± 0.8 mg/L), chemical oxygen demand (COD, 1.15 ± 0.09 mg/L), ammonia nitrogen (NH+4-N, 23.5 ± 4.6 µg/L) and nitrite nitrogen (NO2−, 15 ± 0.8 µg/L), as per the specification of the National Standards of the People’s Republic of China for marine monitoring (GB 17378.1-2007).
Salinity was measured with a portable salinometer (WYYⅡ, Chengdu Hao Chuang Photoelectric Instrument, Chengdu, China). DO was measured by the iodometric method, and COD was tested based on the basic potassium permanganate method. Ammonia and nitrite concentrations (hypobromite oxidation method) and pH were monitored using a UV-visible spectrophotometer (V-1800, Shanghai Mapade Instruments, Shanghai, China) and pH meter (pHs-3C, Shanghai Precision Science Instrument, Shanghai, China).
2.3. Acute Exposure (96 h)
The exposure bioassays were conducted as 96 h acute tests without feeding or aeration. During the acute ammonia nitrogen exposure experiment, 100% of filtered seawater was replaced every 24 h, and an ammonia stock solution was added to the seawater relative to the designated ammonia level (10, 15, 20, 50, 70, 100 and 200 mg/L TAN plus a control).
Ammonium chloride (NH4Cl, analytical reagent, 20 mg/mL) was purchased from Shenyang Chemical Reagent Co. (Shenyang, China), and the stock solution was then diluted to the desired ammonia concentration. The incubation was performed in 2 L glass beakers immersed in 20 L water baths (polyethylene container) with temperature controlled at 19.8 ± 0.5 °C. The 3 replicates of 20 individuals in each beaker were tested and observed daily for mortality. At the start and end of each 24 h interval, pH and ammonia were measured in all concentrations in the control samples.
2.4. Sub-Chronic Exposure (20 d)
A sub-chronic 20 d ammonia test was conducted using aeration with juvenile clams in beakers. As conducted during acute exposure, three replicate beakers per concentration were placed in water baths for sub-chronic exposure. The water temperature was maintained at 19.5 ± 0.5 °C. The clams (20 juveniles) in each replicate beaker were fed with a 20 mL algae mixture 4 times per day, which was previously used to culture these juveniles. The algae mixture (5–8 × 103/mL) mainly contained Prymnesium parvum and Chlorella vulgaris. The exposure solutions were renewed daily.
At the end of the 10 d and 20 d exposure, the clams in each replicate beaker were examined under a microscope (VANOX-S AH-2, Olympus (Beijing) Sales & Service, Beijing, China) to check the reaction and mortality during the acute test. The sub-chronic toxicity experimental concentration of TAN was designed as 0 (control), 1, 2, 3, 5, 9 and 15 mg/L, and mean concentrations were measured as 0.56 ± 0.07, 0.85 ± 0.09, 1.57 ± 0.10, 2.72 ± 0.20, 4.36 ± 0.33, 8.43 ± 0.43 and 14.12 ± 0.50 mg/L (n = 15 for each concentration). The survival and SL of 10 juveniles selected without any particular order were determined on test days 10 and 20.
2.5. Histological Procedures and Assessment
Histological analysis of the live clams at 14.1 mg/L TAN after the 20 d exposure was conducted using histologic sections and the microscopy method [18]. Briefly, the tissue samples were embedded in paraffin wax. The digestive diverticulum, the mantle and foot tissues were fixed in Bouin’s solution. The wax was cut into 5–6 μm thick sections using a microtome (Leica RM2135, Leica Instruments, Nussloch, Germany) and stained with hematoxylin and eosin. The sections were observed using a microscope (VANOX-S AH2, Olympus (Beijing) Sales & Service, Beijing, China) at ×10 and ×40 magnifications, and photographs were obtained using a digital microscope camera (DP71, Olympus (Beijing) Sales & Service, Beijing, China).
2.6. Antioxidant Enzyme Activities
The superoxide dismutase (SOD) and catalase activity (CAT) were determined from whole-body (soft tissue) homogenates. The samples (soft tissue of survived clams) were homogenized in 2 mL of ice-cool 1:9 physiological saline solution [19]. The homogenate was centrifuged at 5000 rpm for 15 min at 4 °C to eliminate cellular debris and cartilage fragments, and the supernatant was used for antioxidant enzyme measurements. The Coomassie Brilliant Blue Total Protein Assay kit (Nanjing Jiancheng Bioengineering Institute, Nanjing, China) was used to determine the total protein contents in tissue samples [20]. Enzyme activities were measured with diagnostic reagent kits purchased from Nanjing Jian Cheng Bioengineering Institute (Nanjing, China). Each enzymatic assay was performed in triplicate. The results were expressed as enzymatic units per mg proteins [21]: an SOD unit was defined as the enzyme amount necessary to inhibit 50% of the reaction rate per mg protein in 1 mL reaction solution; a CAT unit was defined as the enzyme amount that transforms 1 µmol of H2O2 per second per mg protein.
2.7. Data Analysis
Data analysis was conducted using the statistical program SPSS Version 13.0. Data are presented as means ± standard deviation (SD). Differences between treatments were tested by one-way analysis of variance (ANOVA) with the least significant difference (LSD) method for multiple comparisons and by using Duncan’s multiple range test. The level of statistical significance was set at p < 0.05. A freely available internet platform, MOSAIC [22], was used to estimate LC50 with a 95% credible interval (CI). The relative growth rate (RGR) was calculated as follows:
where L0 and L1 are the shell lengths measured before and after exposure.
The concentrations of NH3 were calculated as follows [23]:
where C(TAN) and C(NH3) are in mg/L; pH is the negative log10 of the hydrogen ion's activity; and pK is the acid dissociation constant for water (Equation (4)):
where T is the water temperature in °C; S is salinity in the practical salinity unit (psu). Although applying a safety factor on toxicity data to estimate the “safe level” of environmental ammonia is still under debate [24], the safe concentration (SC) of TAN and NH3 for M. chinensis is estimated by multiplying an application factor 0.1 times the 96 h LC50 [25].
C(NH3) = C(TAN) × fNH3
fNH3 = 1/(10^(pK − pH))
pK = 9.245 + 0.002949 × S + 0.0324 × (25 − T)
3. Results
3.1. Acute Toxicity Test
The mortality of M. chinensis increased along with the increasing concentration of ammonia. The LC50 values for M. chinensis are illustrated in Table 1. The LC50-96 h and SC of TAN for juvenile M. chinensis were 11.1 (10.0; 12.0) and 1.11 mg/L, respectively. The 96 h LC50 and SC of NH3 for juvenile M. chinensis were 0.56 (0.50; 0.60) and 0.056 mg/L, respectively.
3.2. Sub-Chronic Toxicity Test
The mean survival rate was 100% in all treatments with concentrations lower than 8.4 and 14.1 mg/L TAN. The survival rates of the clam were observed at 99.3 ± 0.9% and 97.7 ± 1.5% at 8.4 and 14.1 mg/L TAN, respectively.
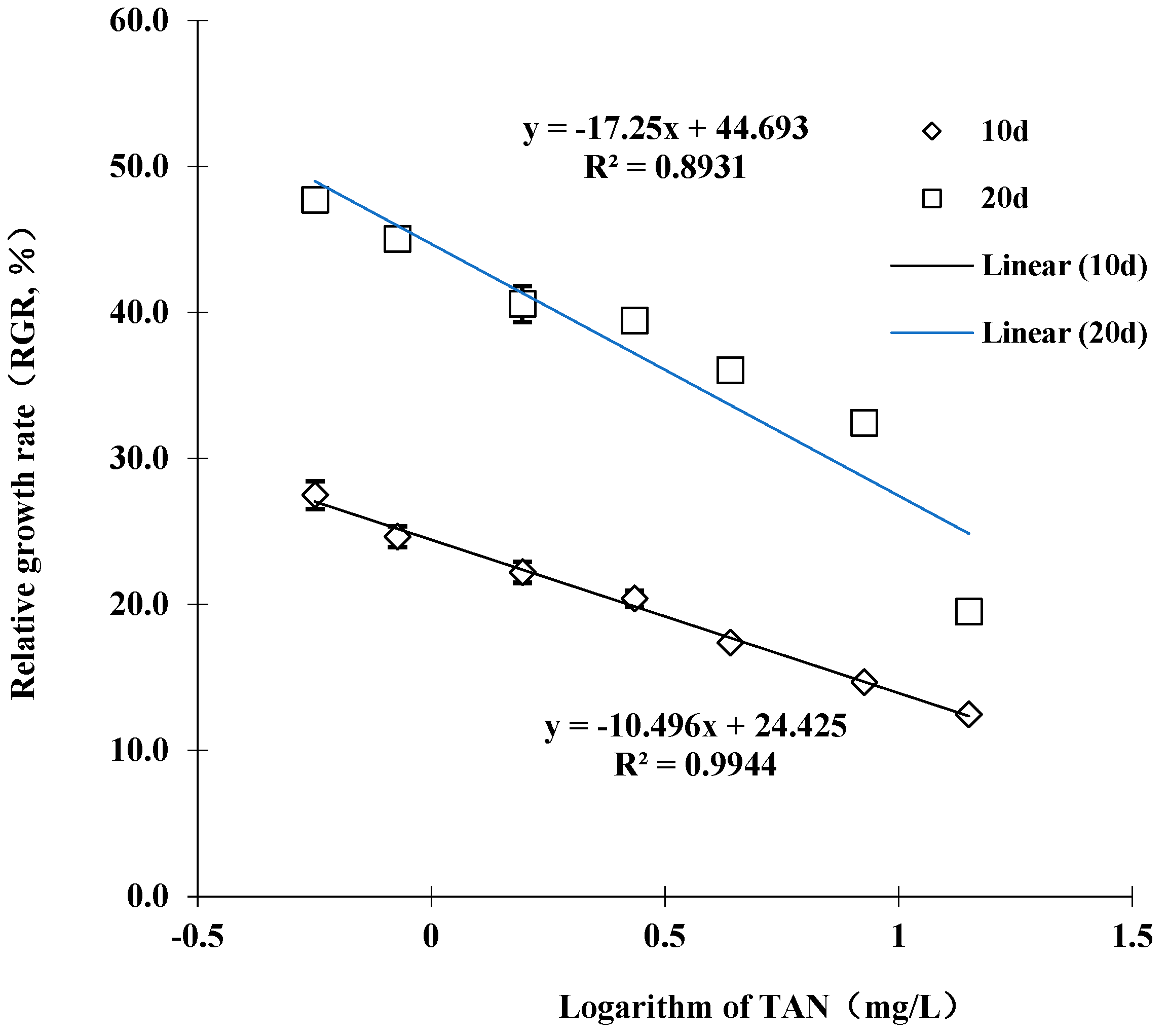
The RGR decreased as the TAN concentration increased (Figure 1). A linear relationship was obtained between the RGR of juvenile M. chinensis and the logarithm of TAN concentration (R2 = 0.99 for 10 d and 0.89 for 20 d, respectively). In the control group, the juvenile had grown by 0.48 ± 0.02 cm of SL on day 20 of exposure. However, the mean increment of SL at 14.1 mg/L TAN (0.661 mg/L NH3) was only 0.20 ± 0.01 cm, which was significantly lower compared to the control (p < 0.05). On day 20 of the experiment, significant differences in SL growth were found among juveniles exposed to a TAN of 1.6 mg/L (0.41 ± 0.02 cm), 2.7 mg/L (0.39 ± 0.01 cm), 4.4 mg/L (0.36 ± 0.01 cm), 8.4 mg/L (0.32 ± 0.01 cm) and the control (p < 0.05). The estimated maximum acceptable toxicant concentration (MATC) for TAN was between 0.8 and 1.6 mg/L TAN (0.038–0.075 mg/L NH3), which was in agreement with the SC obtained from acute toxicity tests (1.11 mg/L TAN and 0.056 mg/L NH3).
3.3. Histological Observations
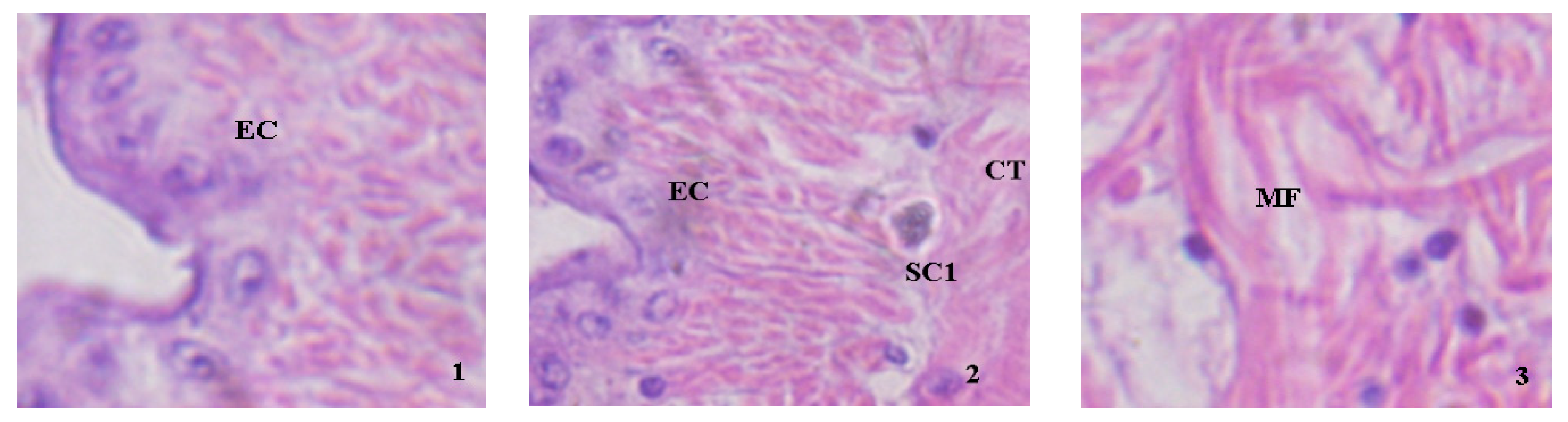
Figure 2 shows the histological observations of juvenile clams exposed at 14.1 mg/L TAN (0.661 mg/L NH3) for 20 days. The normal structure of the mantle is shown in Figure 2(1), and the mantle cavity is identified as the epithelial cell (EC: a tight and neat layer of the epithelium). However, several impairments were observed at 14.1 mg/L TAN (0.661 mg/L NH3) exposure, such as the large granular basophilic cells produced (Figure 2(2)), vesicles-like connective tissue and disordered muscle fiber (Figure 2(3)).
The normal phasic activity of the digestive diverticulum (gland) was observed in the control clams (Figure 2(4)). The gland exposed to ammonia showed vesicles-like acinar (Figure 2(5)), produced abnormal objects (Figure 2(6)), a ruptured acinar, collapsibility, dissolved necrosis (Figure 2(7)) and increased basophilic cells (Figure 2(8)).
The juvenile control clams had an extensive muscular foot that showed connective tissue and muscle fibers (Figure 2(9)). Under the exposure of 14.1 mg/L TAN (0.661 mg/L NH3), histopathological damages in the foot were observed, including epithelial cell break or fall off (Figure 2(10)), muscle fiber disorder (Figure 2(11)) and mucus cell hyperplasia (Figure 2(12)).
3.4. Enzyme Activities under Sub-Chronic Ammonia Stress
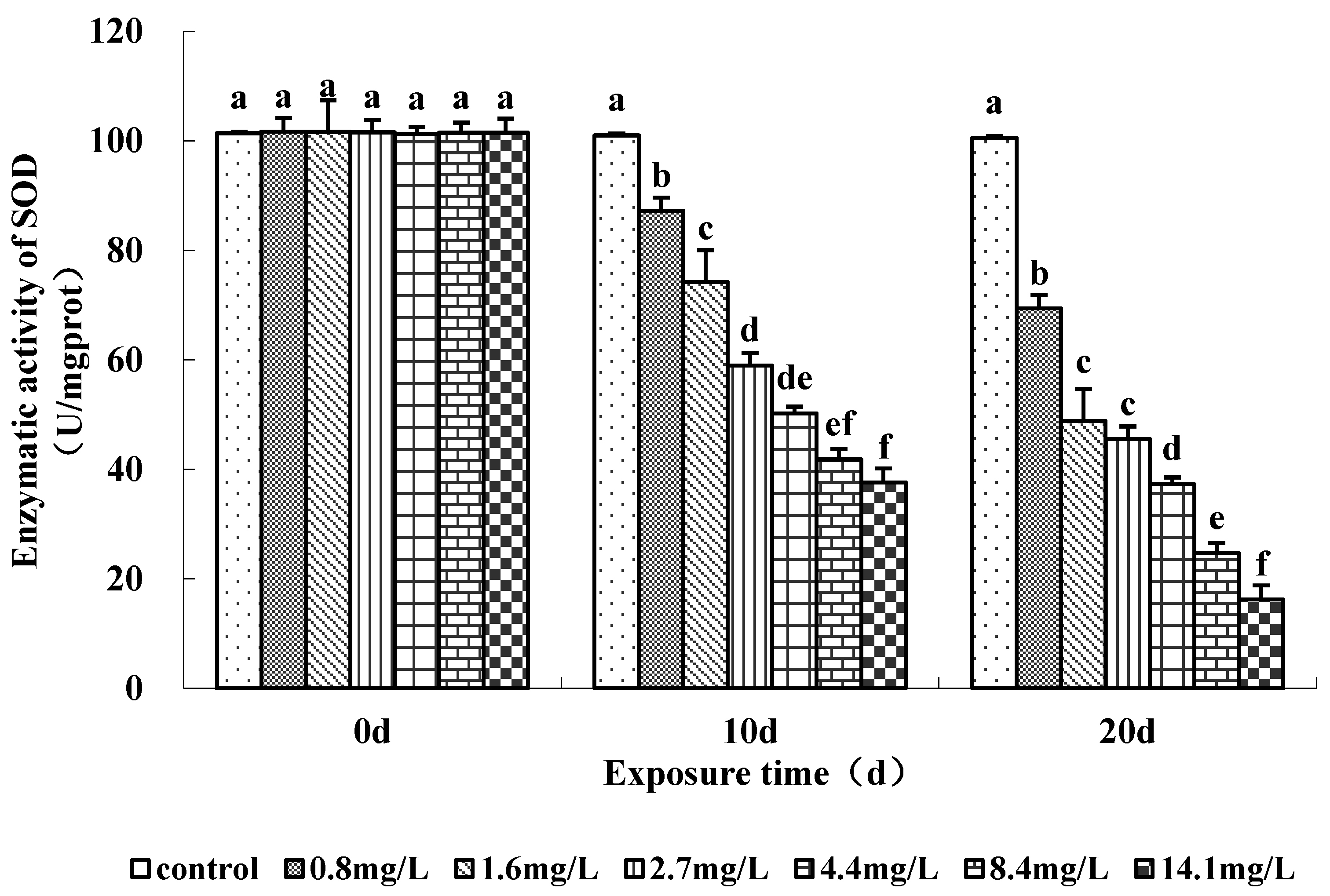
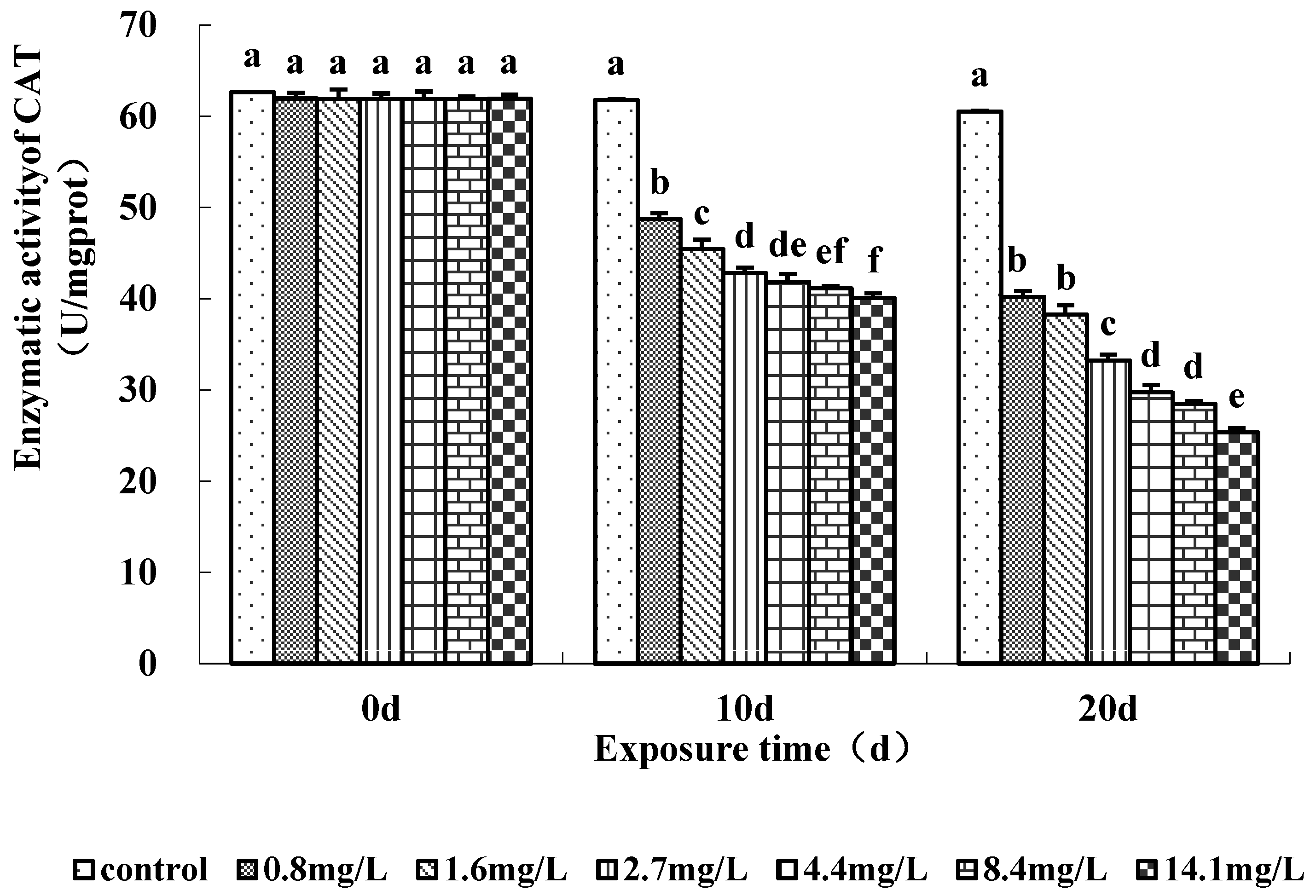
The activities of antioxidant enzymes (SOD and CAT) measured in the soft tissue of the clams were significantly affected by ammonia exposure (Figure 3 and Figure 4). SOD activity tended to decrease with the increase in ammonia concentration on days 10 and 20 (Figure 3). SOD activity was significantly lower in clams exposed to ≥0.8 mg/L TAN (0.038 mg/L NH3) on days 10 and 20. Clams exposed to the highest ammonia level (14.1 mg/L) showed a mean activity of 16% compared to the control at day 20. CAT activity also decreased in ammonia-exposed clams during the experiment (Figure 4). CAT activity was significantly lower in the ≥0.8 mg/L TAN (0.038 mg/L NH3) concentration groups than the control group during days 10 and 20. There were no increases in SOD and CAT activity observed.
4. Discussions and Conclusions
The sensitivity of juvenile M. chinensis to ammonia is comparable to that of other bivalve species. Table 2 summarizes the selected studies using ammonia toxicological values for bivalve species. The 48 h LC50 concentration of larvae surf clam Spisula solidissima was reported as 10.6 mg/L TAN (0.53 mg/L NH3) under similar water quality (pH = 8.14 ± 0.06, T = 19.7 ± 0.4 °C and salinity = 31 ± 0.5‰) [15], which is close to the current study (48 h LC50 = 16.5 mg/L TAN, 0.79 mg/L NH3). Recently, another species of economically important marine clam Cyclina sinensis was found at 96 h LC50 at 80.7 mg/L TAN (4.01 mg/L NH3) (pH = 8.0 ± 0.3, T = 24 °C, salinity = 21 ppt) [26], which is an order of magnitude higher compared to the current study (96 h LC50 = 11.1 mg/L TAN, 0.56 mg/L NH3). As for SC, Batley and Simpson [27] proposed a guideline value of 0.46 mg/L TAN for ammonia in estuarine and marine waters using species sensitivity distributions. In the literature, the guideline value was derived using toxicity data at a pH value from 7 to 8.3 and temperatures from 6 to 28 °C, resulting in a varied fraction of NH3 of TAN [27]. It should be cautioned that NH3 is more toxic compared to NH4+ [28], and guideline values should always be converted to mg/L NH3.
Grice and Bell [29] observed that approximately 0.6–0.85 mg/L of ammonium treatment resulted in significantly a slower growth of juvenile giant clam (Tridacna maxima) in terms of SL during the 25 days. Although pH, temperature and salinity information were not clearly disclosed in the literature [29], it is similar to that observed in the current study, in which case the growth of juvenile clams was significantly reduced at 1.6 mg/L TAN. In the present study, variations in sensitivity to the mortality of juveniles were observed between acute and sub-chronic test results. The deviation, i.e., the survival endpoint that was less sensitive during the 20 d assay, could be due to the testing organisms from different batches. The reference toxicant and reference toxicity tests should be developed to establish an acceptable deviation range in future tests.
In a recent study, Ma et al. [31] reported that the MATC on the growth of the ‘Zebra 2’ strain of the manila clam (Ruditapes philippinarum) juveniles ranges from 58.05 to 95.69 mg/L TAN (3.01–4.97 mg/L NH3, pH = 8.18, T = 27.65 °C, salinity = 27.5). This range is almost two orders of magnitude compared to the current study (0.8–1.6 mg/L TAN, 0.038–0.075 mg/L NH3). However, the exposure period was only 96 h [31], which is shorter than the current study (20 days). Yuan et al. [30] reported an MATC of 0.86–1.80 mg/L TAN (0.054–0.113 mg/L NH3) based on the 24 h hatching rate of bay scallops (Argopecten irradians) at a water temperature of 21.5–22.5 °C, pH of 8.20–8.30 and a salinity of 27–28. The reported MATC range is comparable to the current study.
Although the literature indicated the addition of ammonium as a nutrient for clams [18], growth stimulation was not observed at any ammonium concentrations in the current study. The significant reduction in growth caused by 1.6 mg/L TAN (0.075 mg/L NH3) should be considered in an aquacultural setting or a natural ecosystem.
Clams usually exhibit an extreme retraction of their mantle edge before death, and foot protrusion is also one of the stress manifestations [32]. Lasee [33] conducted histological assessments of juvenile Lampsilis ventricose and observed severe effects with respect to the digestive glands and mantle by 30 and 50 µg/L Cd. In addition, the most evident histopathological effects were vacuolization, necrosis and tissue separations [33], which agree with our current observations. Recently, Liu et al. [18] performed a histological analysis using marine clams (Meretrix meretrix), and epithelial cell exfoliation and necrosis were also observed in the digestive glands exposed to functionalized polystyrene nanoplastics. Necrosis has been considered an uncontrolled form of cell death [34], and colliquative necrosis may be used as a biomarker of exposure to ammonia.
Compared to SOD, CAT is considered a more sensitive biomarker of oxidative stress, although under certain circumstances. [35]. However, our data demonstrated a similar decrease in CAT and SOD activity under ammonium exposure. The decrease in SOD and CAT activity at 0.8 mg/L TAN (0.038 mg/L NH3) indicating the level of ammonium is sufficient to impair the oxidative potential of juveniles.
Liu and Chen [36] found that SOD activity in the hemocytes of white shrimps (Litopenaeus vannamei) decreased significantly at 21.60 mg/L ammonia nitrogen (0.696 mg/L NH3) after 7 days. However, increased CAT activity in the gill of Asian clam (Corbicula fluminea) was found in under 25 mg/L of TAN exposure for 48 h, although the SOD and CAT activity in the digestive gland was suppressed significantly (pH = 8.2, T = 20 ± 0.5 °C) [37]. Recently, Zhao et al. [38] reported the impairment of SOD and CAT activities in the hepatopancreas and gills of triangle sail mussels (Hyriopsis cumingii) exposed to 6.43 mg/L NH3 for 96 h. Although whole-body tissue was examined in the current study, the ammonia level is considered more environmentally relevant.
The current results demonstrated that the relatively sensitive endpoint for ammonia toxicity in this study was antioxidant enzyme activities, compared to survival and growth inhibition. Therefore, the recommended water quality criteria to protect juvenile clams from ammonia would be 0.8 mg/L TAN (0.038 mg/L NH3). In Australia and New Zealand, the NH3 guideline for the protection of aquaculture species in saltwater production is < 0.1 mg/L [39], which seems insufficient for the protection of M. chinensis. Han et al. [40] suggested that NH3 should be controlled below 0.020 mg/L during the breeding of Manila clams (Ruditapes philippinarum).
While ammonia is not a common guideline indicator for marine water quality, it is generally traced in freshwater and fisheries. For example, Canada set a criterion for protecting freshwater aquatic life at <0.019 mg/L NH3 [41]. Similarly, the Chinese water quality standard for fisheries was developed and set at ≤0.02 mg/L NH3 [42]. The recommended water quality criteria derived from the current study were comparable to both guideline values, indicating that adopting freshwater/fishery guidelines for the marine environment could still be promising.
Our work revealed that the sensitivity of M. chinensis to ammonia is comparable to other species of surf clams and economically crucial marine clams. Sub-chronic ammonia exposure for 10 d and 20 d caused a decrease in CAT and SOD activity ≥0.8 mg/L TAN (0.038 mg/L NH3), indicating that ammonia exposure could induce oxidative stress by decreasing antioxidant enzyme activity. A significant effect on growth was observed using 1.6 mg/L of TAN (0.075 mg/L NH3). The LC50 of TAN (NH3) for juvenile M. chinensis after 24, 48 and 96 h were 90.4 (4.14), 16.5 (0.79) and 11.1 (0.56) mg/L TAN, respectively. Various histological effects were observed at 14.1 mg/L TAN (0.661 mg/L NH3) for 20 d exposure. The results indicated that decreased antioxidant enzyme activity could be a sensitive indicator of ammonia stress.
Overall, under the current experimental condition in seawater (pH = 8.22 ± 0.04, T = 19.5 ± 0.5 °C and salinity = 29.6 ± 0.4 psu), the recommended criterion for the protection of M. chinensis from ammonia would be 0.8 mg/L TAN (0.038 mg/L NH3). As the mixtures of ammonium associated with other pollutants may be more toxic than the individual concentration, the regional freshwater/fishery guidelines of ammonia should be advised for the marine environment. The current study provided ecotoxicological insights into the mechanisms underlying ammonia toxicity effects in clams, and it also provided relevant information for clam development and healthy aquaculture. Additional studies are needed to test other life stages of clams for a more extended period.
Author Contributions
Conceptualization, Y.D. (Yuanyuan Dai) and F.Y.; methodology, Y.D. (Yuanyuan Dai); software, Y.D. (Yubo Dong); validation, J.J., Y.D. (Yuanyuan Dai) and Z.C.; formal analysis, Y.D. (Yuanyuan Dai); investigation, Z.C.; resources, F.Y.; data curation, Y.D. (Yubo Dong); writing—original draft preparation, Y.D. (Yuanyuan Dai); writing—review and editing, Z.C. and F.Y.; visualization, J.J.; supervision, F.Y.; project administration, Y.D. (Yuanyuan Dai); funding acquisition, F.Y. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National High-tech Research and Development Program of China (863 Program), grant 2006AA10A410; Key Research and Development Program of Liaoning Province of China (990387); and the Foundation of Liaoning Province Education Administration of China (2007T104).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available upon request from the corresponding author.
Acknowledgments
The authors thank Xiwu Yan at Dalian Ocean University for administrative support.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- FAO. The State of World Fisheries and Aquaculture 2022. Towards Blue Transformation; FAO: Rome, Italy, 2022; Available online: https://www.fao.org/documents/card/en/c/cc0461en (accessed on 16 January 2023).
- Kim, S.H.; Chung, E.; Lee, K. Oocyte Degeneration Associated with Follicle Cells in Female Mactra chinensis (Bivalvia: Mactridae). Dev. Reprod. 2014, 18, 321–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Res, M.; Reunov, A.; Lutaenko, K.; Vekhova, E.; Zhang, J.; Zakharov, E.; Sharina, S.; Alexandrova, Y.; Reunova, Y.; Akhmadieva, A.; et al. In the Asia-Pacific region, the COI DNA test revealed the divergence of the bivalve mollusc Mactra chinensis into three species; can these species be distinguished using shell coloration and sperm structure ? Helgol. Mar. Res. 2021, 75, 7. [Google Scholar] [CrossRef]
- Zhang, X.; Li, Z.; Huo, Z.; Yan, X.; Yang, F.; Liu, H.; Zhang, X. Effect of substrate component on the growth and survival of juvenile sunray surf clam (Mactra chinensis Philippi). J. Ocean Univ. China 2016, 15, 676–680. [Google Scholar] [CrossRef]
- Cong, M.; Li, Y.; Xu, H.; Lv, J.; Wu, H.; Zhao, Y. Ammonia nitrogen exposure caused structural damages to gill mitochondria of clam Ruditapes philippinarum. Ecotoxicol. Environ. Saf. 2021, 222, 112528. [Google Scholar] [CrossRef] [PubMed]
- Camargo, J.A.; Alonso, Á. Ecological and toxicological effects of inorganic nitrogen pollution in aquatic ecosystems: A global assessment. Environ. Int. 2006, 32, 831–849. [Google Scholar] [CrossRef]
- Lu, J.; Yao, T.; Shi, S.; Ye, L. Effects of acute ammonia nitrogen exposure on metabolic and immunological responses in the Hong Kong oyster Crassostrea hongkongensis. Ecotoxicol. Environ. Saf. 2022, 237, 113518. [Google Scholar] [CrossRef]
- Boardman, G.D.; Starbuck, S.M.; Hudgins, D.B.; Li, X.; Kuhn, D.D. Toxicity of ammonia to three marine fish and three marine invertebrates. Environ. Toxicol. 2004, 19, 134–142. [Google Scholar] [CrossRef]
- Liu, M.; Li, Y.; Wang, H.; Wang, H.; Qiao, R.; Jeppesen, E. Ecosystem complexity explains the scale-dependence of ammonia toxicity on macroinvertebrates. Water Res. 2022, 226, 119266. [Google Scholar] [CrossRef]
- Cong, M.; Li, Y.; Xu, H.; Lv, J.; Wu, H.; Zhao, Y.; Wang, N.; Ingersoll, C.G.; Hardesty, D.K.; Ivey, C.D.; et al. Effects of acute ammonia exposure on antioxidant and detoxification metabolism in clam Cyclina sinensis. Environ. Toxicol. Chem. 2021, 26, 2569–2575. [Google Scholar]
- Kallqvist, T.; Svenson, A. Assessment of ammonia toxicity in tests with the microalga, Nephroselmis pyriformis, Chlorophyta. Water Res. 2003, 37, 477–484. [Google Scholar] [CrossRef]
- Chen, J.; Lei, S.-C. Toxicity of Ammonia and Nitrite to Penueus monodon Juveniles. J. World Aquac. Soc. 1990, 21, 300–306. [Google Scholar] [CrossRef]
- Lin, W.; Luo, H.; Wu, J.; Hung, T.C.; Cao, B.; Liu, X.; Yang, J.; Yang, P. A Review of the Emerging Risks of Acute Ammonia Nitrogen Toxicity to Aquatic Decapod Crustaceans. Water 2023, 15, 27. [Google Scholar] [CrossRef]
- Lv, L.; Ren, J.; Zhang, H.; Sun, C.; Dong, Y.; Lin, Z. Transcriptomic Analysis of Gill and Hepatopancreas in Razor Clam (Sinonovacula constricta) Exposed to Acute Ammonia. Front. Mar. Sci. 2022, 9, 832494. [Google Scholar] [CrossRef]
- Ferretti, J.A.; Calesso, D.F. Toxicity of ammonia to surf clam (Spisula solidissima) larvae in saltwater and sediment elutriates. Mar. Environ. Res. 2011, 71, 189–194. [Google Scholar] [CrossRef] [Green Version]
- Cong, M.; Wu, H.; Yang, H.; Zhao, J.; Lv, J. Gill damage and neurotoxicity of ammonia nitrogen on the clam Ruditapes philippinarum. Ecotoxicology 2017, 26, 459–469. [Google Scholar] [CrossRef]
- USEPA. Aquatic Life Ambient Water Quality Criteria for Ammonia—Freshwater 2013; USEPA: Washington, DC, USA, 2013; Volume EPA-822-R-18-002. Available online: https://www.epa.gov/sites/default/files/2015-08/documents/aquatic-life-ambient-water-quality-criteria-for-ammonia-freshwater-2013.pdf (accessed on 16 January 2023).
- Liu, L.; Zheng, H.; Luan, L.; Luo, X.; Wang, X.; Lu, H.; Li, Y.; Wen, L.; Li, F.; Zhao, J. Functionalized polystyrene nanoplastic-induced energy homeostasis imbalance and the immunomodulation dysfunction of marine clams (Meretrix meretrix) at environmentally relevant concentrations. Environ. Sci. Nano 2021, 8, 2030–2048. [Google Scholar] [CrossRef]
- Chen, C.; Ma, C.; Dai, Y.; Wang, X.; Bai, D.; Shang, X.; Jia, L. Effect of dietary nano-Se on oxidation resistance of juvenile tongue sole (Cynogiossus semilaevis). Feed Res. 2019, 1, 25–30. (In Chinese) [Google Scholar] [CrossRef]
- Maoxiao, P.; Xiaojun, L.; Donghong, N.; Bo, Y.; Tianyi, L.; Zhiguo, D. Survival, growth and physiology of marine bivalve (Sinonovacula constricta) in long-term low-salt culture. Sci. Rep. 2019, 9, 2819. [Google Scholar] [CrossRef] [Green Version]
- Sabatini, S.E.; Brena, B.M.; Luquet, C.M.; San Julián, M.; Pirez, M.; de Molina, M.d.C.R. Microcystin accumulation and antioxidant responses in the freshwater clam Diplodon chilensis patagonicus upon subchronic exposure to toxic Microcystis aeruginosa. Ecotoxicol. Environ. Saf. 2011, 74, 1188–1194. [Google Scholar] [CrossRef]
- Charles, S.; Veber, P.; Delignette-Muller, M.L. MOSAIC: A web-interface for statistical analyses in ecotoxicology. Environ. Sci. Pollut. Res. 2018, 25, 11295–11302. [Google Scholar] [CrossRef]
- Lei, Y. Aquaculture Water Environmental Chemistry; China Agricultural Press: Beijing, China, 2004; ISBN 7109085813. [Google Scholar]
- Zhang, L.; Xu, E.G.; Li, Y.; Liu, H.; Vidal-Dorsch, D.E.; Giesy, J.P. Ecological risks posed by ammonia nitrogen (AN) and un-ionized ammonia (NH3) in seven major river systems of China. Chemosphere 2018, 202, 136–144. [Google Scholar] [CrossRef]
- Mummert, A.K.; Neves, R.J.; Newcomb, T.J.; Cherry, D.S. Sensitivity of juvenile freshwater mussels (Lampsilis fasciola, Villosa iris) to total and un-ionized ammonia. Environ. Toxicol. Chem. 2003, 22, 2545–2553. [Google Scholar] [CrossRef]
- Ge, H.; Liu, J.; Ni, Q.; Wang, F.; Dong, Z. Effects of acute ammonia exposure and post-exposure recovery on nonspecific immunity in Clam Cyclina sinensis. Isr. J. Aquac. 2022, 74, 1–10. [Google Scholar] [CrossRef]
- Batley, G.E.; Simpson, S.L. Development of guidelines for ammonia in estuarine and marine water systems. Mar. Pollut. Bull. 2009, 58, 1472–1476. [Google Scholar] [CrossRef] [PubMed]
- Hao, M.; Zuo, Q.; Zhang, W.; Feng, Y.; Wang, L.; Yu, L.; Zhang, X.; Li, J.; Huang, Z. Toxicological Assessment of Ammonia Exposure on Carassius auratus red var. Living in Landscape Waters. Bull. Environ. Contam. Toxicol. 2019, 103, 814–821. [Google Scholar] [CrossRef] [PubMed]
- Grice, A.M.; Bell, J.D. Application of ammonium to enhance the growth of giant clams (Tridacna maxima) in the land-based nursery: Effects of size class, stocking density and nutrient concentration. Aquaculture 1999, 170, 17–28. [Google Scholar] [CrossRef]
- Yuan, H.; Yang, C.; Yang, F.; Yan, X. Impact of total ammonia nitrogen on survival and growth of larval and juvenile bay scallop Argopecten irradians. J. Dalian Ocean Univ. 2017, 32, 268–274. (In Chinese) [Google Scholar] [CrossRef]
- Ma, Q.; Jin, Y.; Jahan, K.; Jami, M.E.H.; Xu, Q.; Huo, Z.; Yang, F.; Yan, X. Effects of ammonia nitrogen on early growth and survival of the ‘Zebra 2’ strain of the manila clam (Ruditapes philippinarum). Aquac. Res. 2021, 52, 4767–4775. [Google Scholar] [CrossRef]
- Soon, T.K.; Ransangan, J. Extrinsic Factors and Marine Bivalve Mass Mortalities: An Overview. J. Shellfish Res. 2019, 38, 223–232. [Google Scholar] [CrossRef]
- Lasee, B.A. Histological and Ultrastructural Studies of Larval and Juvenile Lampsilis (Bivalvia) from the Upper Mississippi River; Iowa State University: Ames, IA, USA, 1991. [Google Scholar]
- Carella, F.; Feist, S.W.; Bignell, J.P.; De Vico, G. Comparative pathology in bivalves: Aetiological agents and disease processes. J. Invertebr. Pathol. 2015, 131, 107–120. [Google Scholar] [CrossRef]
- Vlahogianni, T.; Dassenakis, M.; Scoullos, M.J.; Valavanidis, A. Integrated use of biomarkers (superoxide dismutase, catalase and lipid peroxidation) in mussels Mytilus galloprovincialis for assessing heavy metals’ pollution in coastal areas from the Saronikos Gulf of Greece. Mar. Pollut. Bull. 2007, 54, 1361–1371. [Google Scholar] [CrossRef]
- Liu, C.H.; Chen, J.C. Effect of ammonia on the immune response of white shrimp Litopenaeus vannamei and its susceptibility to Vibrio alginolyticus. Fish Shellfish Immunol. 2004, 16, 321–334. [Google Scholar] [CrossRef]
- Zhang, T.; Yan, Z.; Zheng, X.; Wang, S.; Fan, J.; Liu, Z. Effects of acute ammonia toxicity on oxidative stress, DNA damage and apoptosis in digestive gland and gill of Asian clam (Corbicula fluminea). Fish Shellfish Immunol. 2020, 99, 514–525. [Google Scholar] [CrossRef]
- Zhao, Q.; Feng, K.; Zhang, L.; Bai, Y.; Yao, W. Effects of acute ammonia stress on antioxidant responses, histopathology and ammonia detoxification metabolism in triangle sail mussels (Hyriopsis cumingii). Water 2021, 13, 425. [Google Scholar] [CrossRef]
- Australian and New Zealand Environment and Conservation Council. Australian and New Zealand Guidelines for Fresh and Marine Water Quality Volume 1; Australian and New Zealand Environment and Conservation Council: Canberra, Australia, 2000. Available online: https://www.waterquality.gov.au/sites/default/files/documents/anzecc-armcanz-2000-guidelines-vol1.pdf (accessed on 29 December 2022).
- Han, T.; Yang, F.; Jia, J.; Li, X.; Yan, X. Effects of total ammonia nitrogen content on early growth and development of Manila clam Ruditapes philippinarum. J. Dalian Ocean Univ. 2018, 33, 210–216. (In Chinese) [Google Scholar] [CrossRef]
- Canadian Council of Ministers of the Environment. Canadian Water Quality Guidelines for the Protection of Aquatic Life: Ammonia; Canadian Council of Ministers of the Environment: Winnipeg, Canada, 2010; Available online: https://ccme.ca/en/res/ammonia-en-canadian-water-quality-guidelines-for-the-protection-of-aquatic-life.pdf (accessed on 3 December 2022).
- GB11607-89; Water Quality Standard for Fisheries. Ministry of Ecology and Environment of the People’s Republic of China: Beijing, China, 1990. Available online: https://www.chinesestandard.net/PDF/English.aspx/GB11607-1989 (accessed on 30 November 2022).
Figure 1.
Relative growth rate based on the shell length (SL) of juvenile M. chinensis exposed at different ammonia concentrations for 10 and 20 days. Error bars represent standard deviation (n = 30).
Figure 1.
Relative growth rate based on the shell length (SL) of juvenile M. chinensis exposed at different ammonia concentrations for 10 and 20 days. Error bars represent standard deviation (n = 30).

Figure 2.
The photomicrographs of histological sections of juvenile clams after 20 days of exposure. (1) Control mantle without ammonia exposure (X 400); (2,3) impaired mantle exposed with 14.1 mg/L TAN (X 400); (4) normal structure of digestive diverticulum (X 100); (5–8) impaired digestive diverticulum (X 400); (9) juvenile foot in the control group (X 200); (10–12) impaired foot exposed at 14.1 mg/L TAN (400). EC: Epithelial cell; SC1: large granular basophilic cells; CT: connective tissue; MF: muscle fiber.
Figure 2.
The photomicrographs of histological sections of juvenile clams after 20 days of exposure. (1) Control mantle without ammonia exposure (X 400); (2,3) impaired mantle exposed with 14.1 mg/L TAN (X 400); (4) normal structure of digestive diverticulum (X 100); (5–8) impaired digestive diverticulum (X 400); (9) juvenile foot in the control group (X 200); (10–12) impaired foot exposed at 14.1 mg/L TAN (400). EC: Epithelial cell; SC1: large granular basophilic cells; CT: connective tissue; MF: muscle fiber.


Figure 3.
Changes in superoxide dismutase (SOD) activity (U/mg proteins) in the soft tissues of clams. Ammonia concentrations were measured at 0.6 (control), 0.8, 1.6, 2.7, 4.4, 8.4 and 14.1 mg/L TAN. Error bars represent the standard deviation (n = 3). Treatments that do not share a common letter significantly differ with respect to exposure time (p < 0.05).
Figure 3.
Changes in superoxide dismutase (SOD) activity (U/mg proteins) in the soft tissues of clams. Ammonia concentrations were measured at 0.6 (control), 0.8, 1.6, 2.7, 4.4, 8.4 and 14.1 mg/L TAN. Error bars represent the standard deviation (n = 3). Treatments that do not share a common letter significantly differ with respect to exposure time (p < 0.05).

Figure 4.
Catalase activity (U/mg proteins) in the soft tissues of M. chinensis Philippi for 20 days. Ammonia concentrations were measured at 0.6 (control), 0.8, 1.6, 2.7, 4.4, 8.4 and 14.1 mg/L TAN. Error bars represent standard deviation (n = 3). Treatments that do not share a common letter indicate a significant difference from each other with respect to exposure time (p < 0.05).
Figure 4.
Catalase activity (U/mg proteins) in the soft tissues of M. chinensis Philippi for 20 days. Ammonia concentrations were measured at 0.6 (control), 0.8, 1.6, 2.7, 4.4, 8.4 and 14.1 mg/L TAN. Error bars represent standard deviation (n = 3). Treatments that do not share a common letter indicate a significant difference from each other with respect to exposure time (p < 0.05).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
The median lethal concentration (LC50) with 95% credible interval (CI) for juvenile M. chinensis.
Table 1.
The median lethal concentration (LC50) with 95% credible interval (CI) for juvenile M. chinensis.
| Time (h) | LC50-TAN (mg/L) | LC50-NH3 (mg/L) |
|---|---|---|
| 24 | 90.4 (78.0; 106.0) | 4.14 (3.57; 4.85) |
| 48 | 16.5 (15.1; 18.3) | 0.79 (0.72; 0.88) |
| 96 | 11.1 (10.0; 12.0) | 0.56 (0.50; 0.60) |
Table 2.
Selected studies using bivalve species in the ecotoxicological assessments of ammonia.
| Species | Stage | Effect | Response | Duration (h) | pH | T (°C) | Salinity | Toxicological Values (TAN/NH3, mg/L) |
|---|---|---|---|---|---|---|---|---|
| Spisula Solidissima [15] | Larvae | Mortality | LC50 | 48 | 8.14 ± 0.06 | 19.7 ± 0.4 | 31 ± 0.5‰ | 10.6/0.53 |
| Growth | EC50 | 2.35/0.12 | ||||||
| Cyclina Sinensis [26] | 3.38 ± 0.21 g | Mortality | LC50 | 72 | 8.0 ± 0.3 | 24 | 21 ppt | 105.0/5.22 |
| 96 | 80.7/4.01 | |||||||
| Ruditapes philippinarum [29] | Larvae | Growth | MATC | 96 | 8.1 | 23.05 | 27 | 2.98–5.85/0.09–0.18 |
| Juvenile | 8.18 | 27.65 | 27.5 | 58.05–95.69/3.01–4.97 | ||||
| Argopecten Irradians [30] | Larvae | Growth | EC50 | 144 | 7.95–8.10 | 23–25 | 27–28 | 2.023/0.089 |
| Embryo | Hatching | MATC | 24 | 8.20–8.30 | 21.5–22.5 | 0.86–1.80/0.054–0.113 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Dai, Y.; Dong, Y.; Yang, F.; Chen, Z.; Jia, J. Effects of Ammonia on Juvenile Sunray Surf Clam (Mactra chinensis Philippi) in Laboratory Tests. Pollutants 2023, 3, 232-242. https://doi.org/10.3390/pollutants3020017
AMA Style
Dai Y, Dong Y, Yang F, Chen Z, Jia J. Effects of Ammonia on Juvenile Sunray Surf Clam (Mactra chinensis Philippi) in Laboratory Tests. Pollutants. 2023; 3(2):232-242. https://doi.org/10.3390/pollutants3020017
Chicago/Turabian StyleDai, Yuanyuan, Yubo Dong, Feng Yang, Zhongzhi Chen, and Jia Jia. 2023. "Effects of Ammonia on Juvenile Sunray Surf Clam (Mactra chinensis Philippi) in Laboratory Tests" Pollutants 3, no. 2: 232-242. https://doi.org/10.3390/pollutants3020017