
Taxonomic Diversity of the Microbial Biofilms Collected along the Thermal Streams on Kunashir Island

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Description of the Sample Collection Sites
2.2. DNA Extraction, Purification, Amplification, and Sequencing
2.3. Data Analysis
3. Results
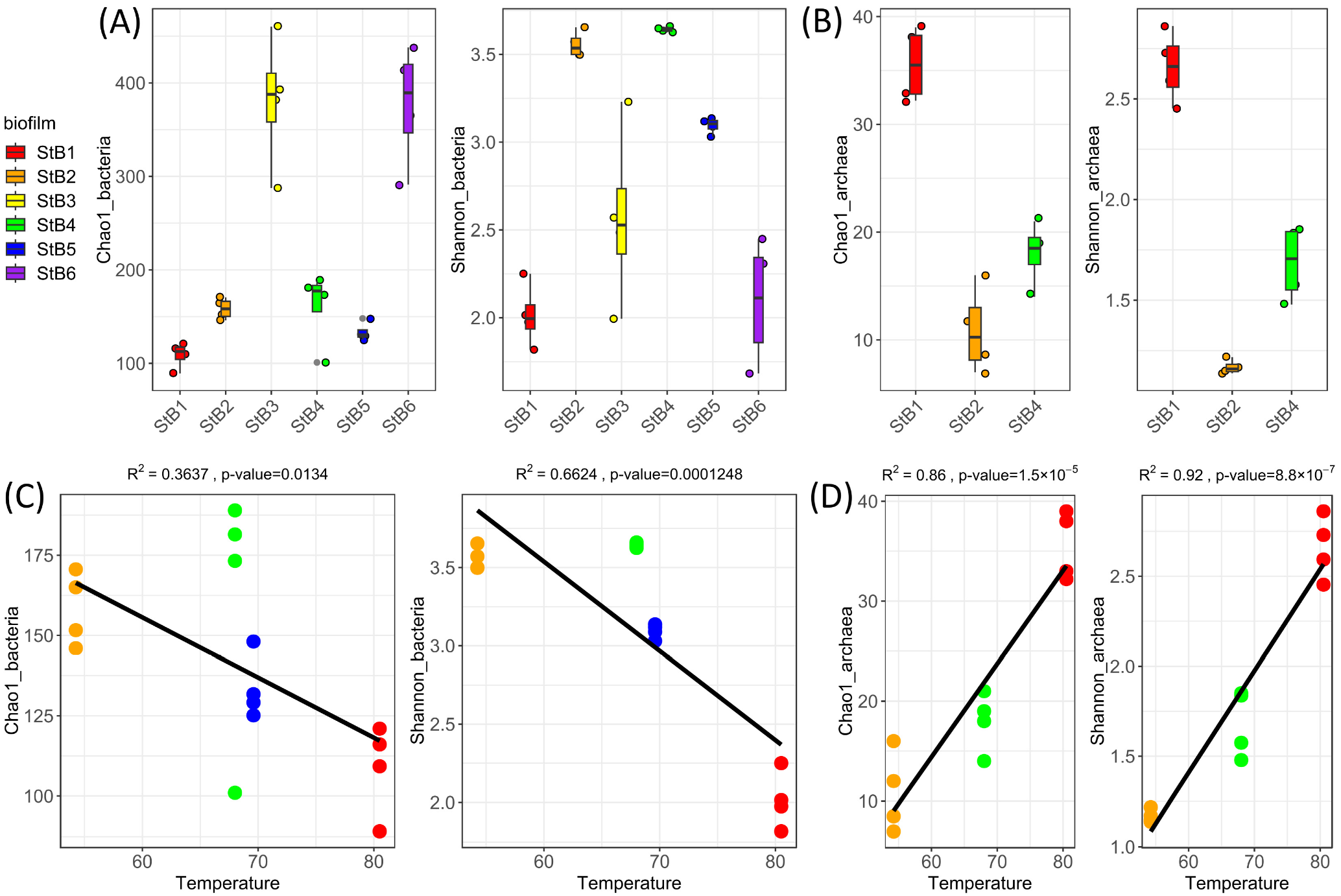
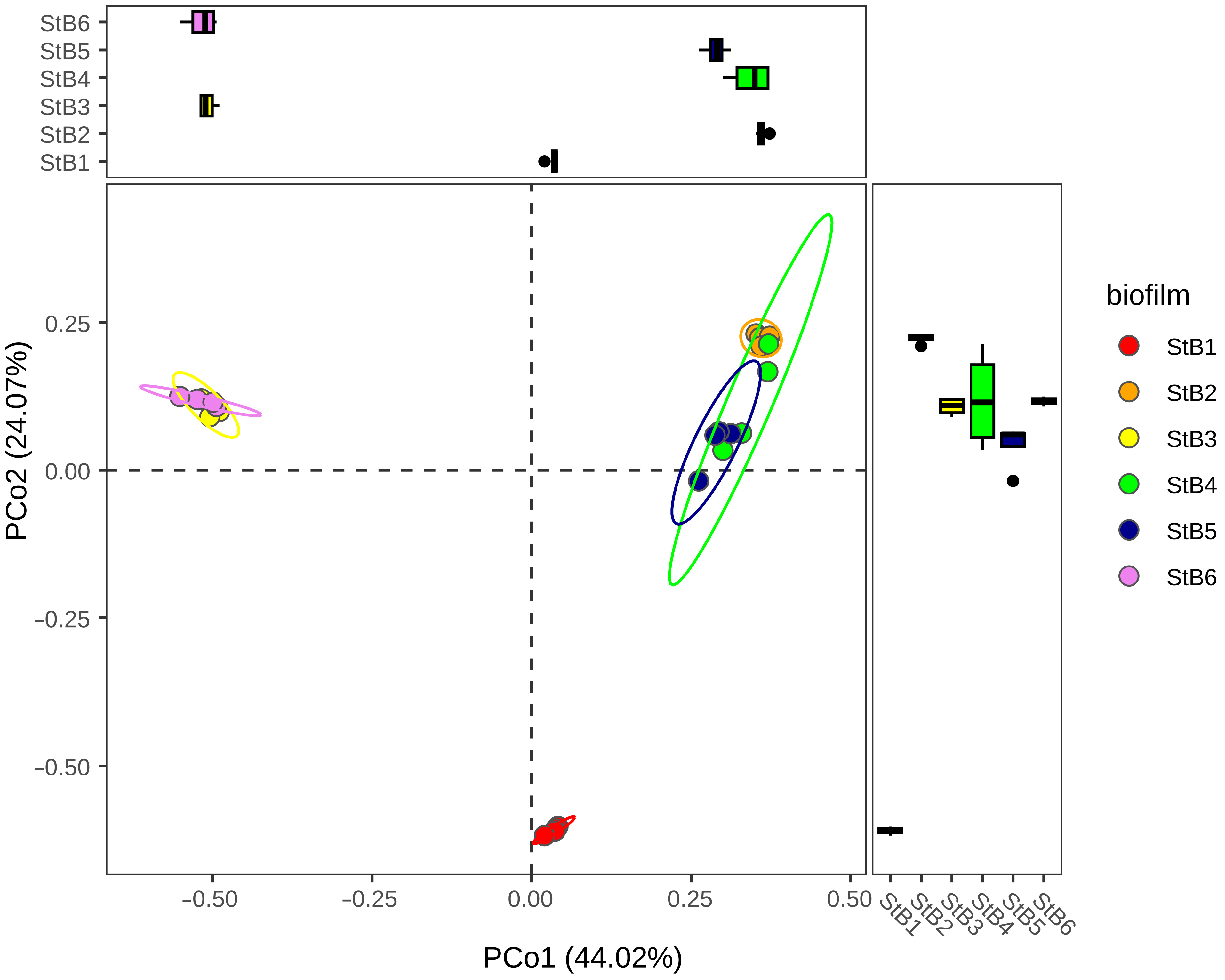
3.1. Estimation of Richness and Diversity of the Biofilm-Forming Microbial Communities
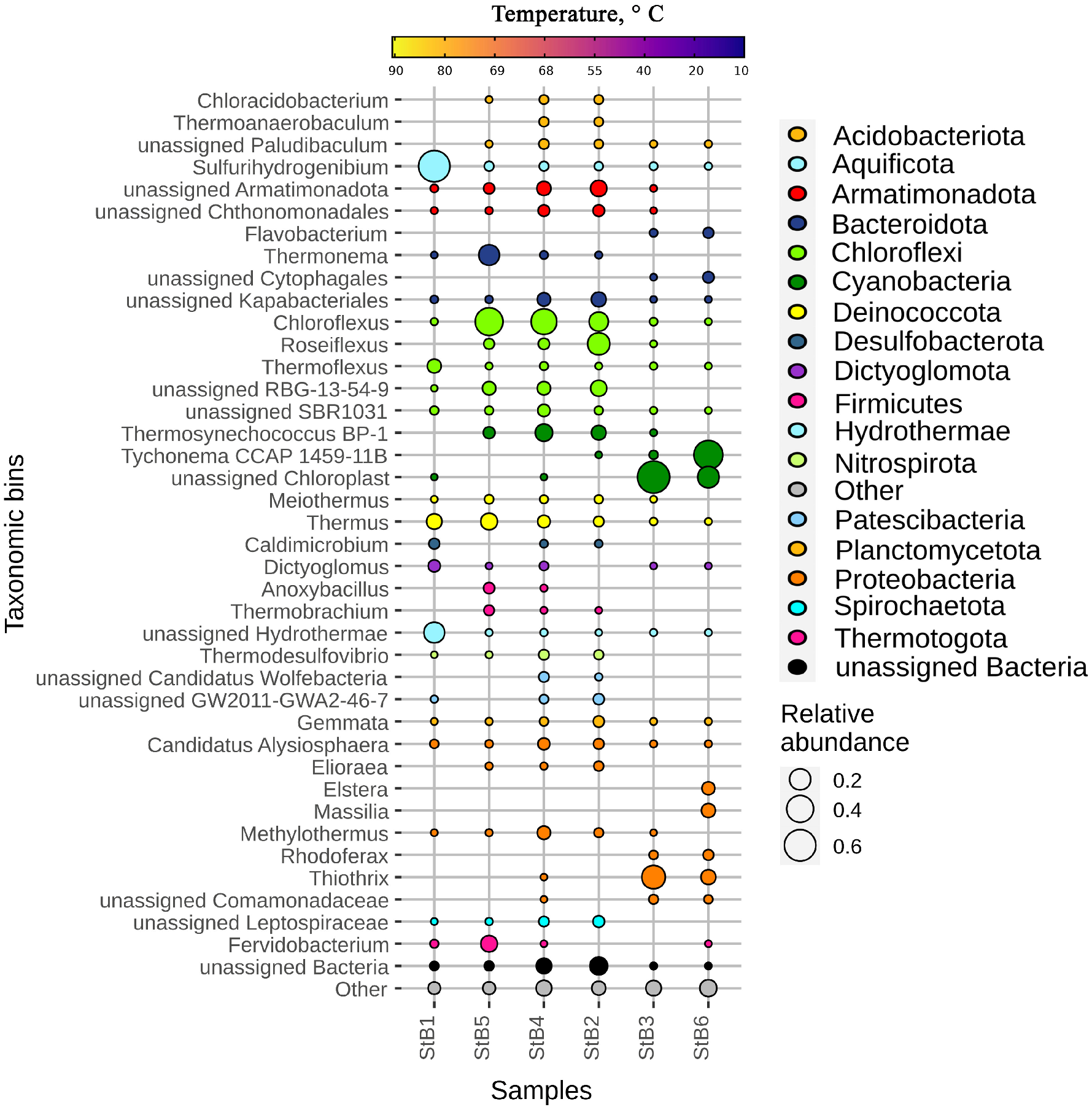
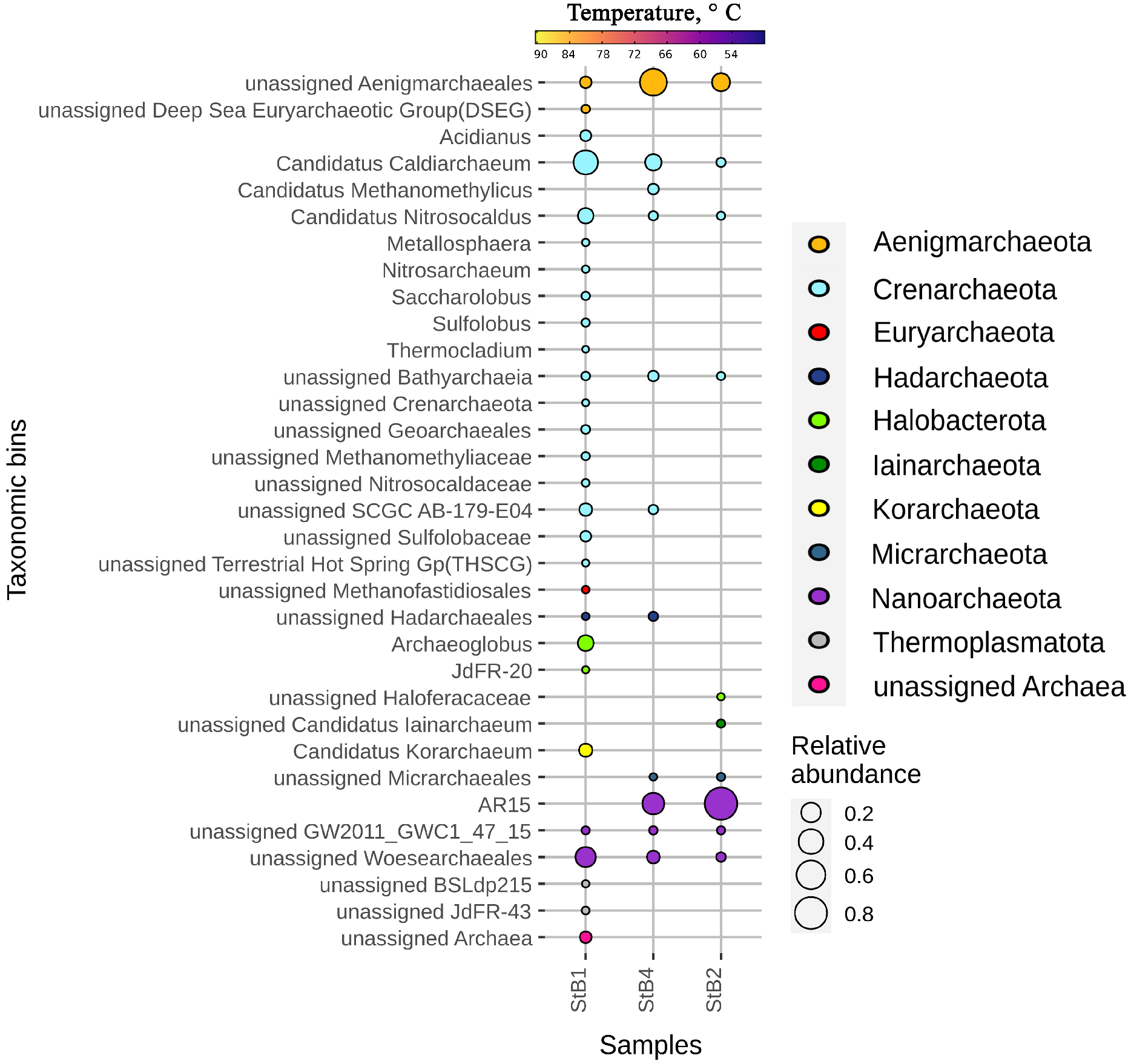
3.2. The Taxonomic Diversity of Bacteria and Archaea in the Biofilm-Forming Communities
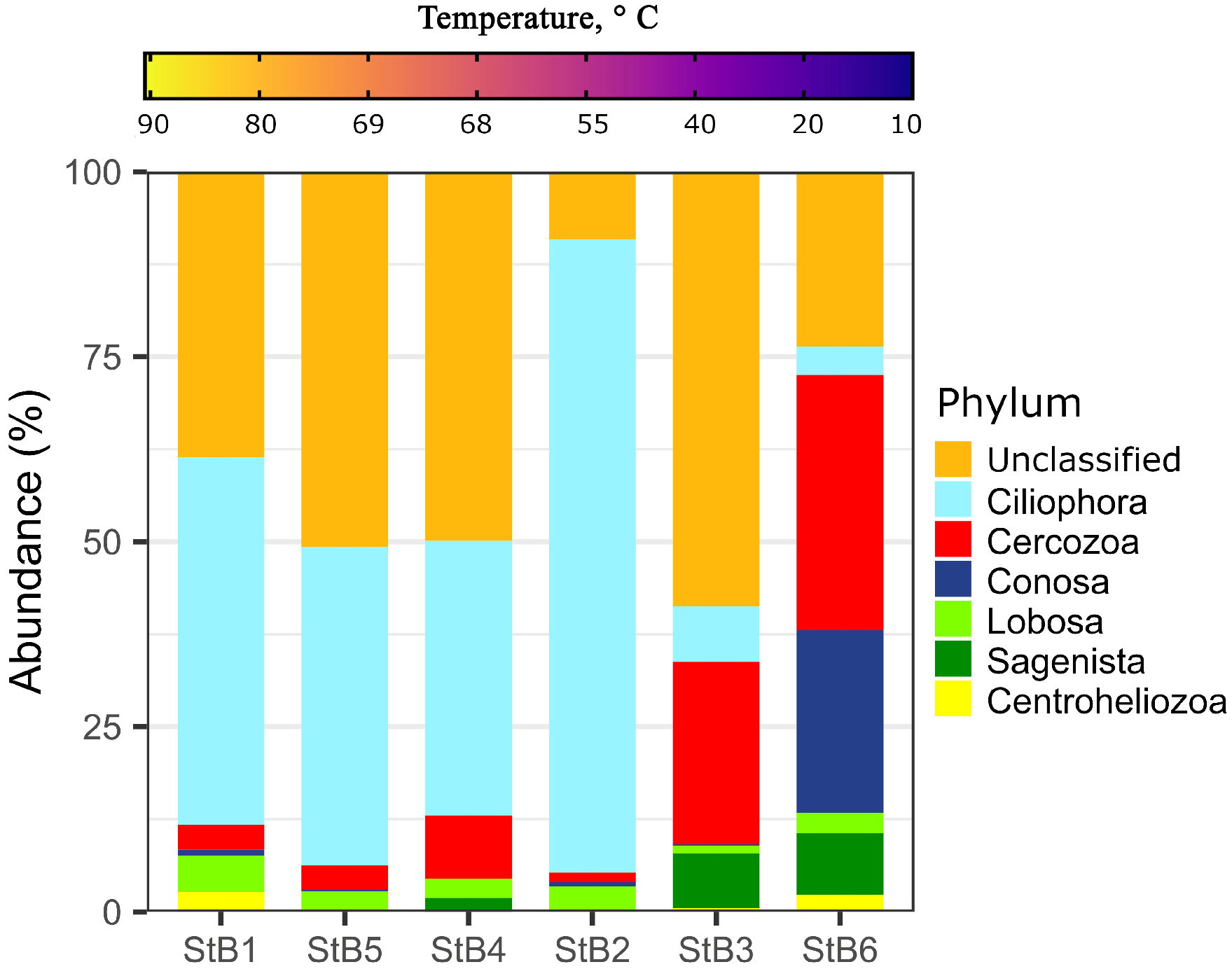
3.3. Taxonomic Diversity of Protists in the Biofilm-Forming Microbial Communities
4. Discussion
4.1. Prokaryotic Community Changes Correlate with the Temperature Gradient
4.2. The Evidence of Presence of Ciliates in the Hot Springs
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zharkov, R.V. Active volcanoes and thermal springs of Kunashir Island (Russia). IOP Conf. Ser. Earth Environ. Sci. 2020, 459, 022039. [Google Scholar] [CrossRef]
- Kalacheva, E.G.; Taran, Y.A. Processes responsible for variations in the isotopic composition (ΔD and Δ18O) of thermal waters of the Kuril Islands arc. Volcanolog. Seismol. 2019, 4, 3–17. [Google Scholar] [CrossRef]
- Markhinin, E.K.; Stratula, D.S. Hydrothermal Systems of Kuril Islands; Nauka: Moscow, Russia, 1977; p. 212. (In Russian) [Google Scholar]
- Straub, C.T.; Counts, J.A.; Nguyen, D.M.N.; Wu, C.H.; Zeldes, B.M.; Crosby, J.R.; Conway, J.M.; Otten, J.K.; Lipscomb, G.L.; Schut, G.J.; et al. Biotechnology of extremely thermophilic archaea. FEMS Microbiol. Rev. 2018, 42, 543–578. [Google Scholar] [CrossRef] [PubMed]
- Sani, R.; Rathinam, N. Extremophilic Microbial Processing of Lignocellulosic Feedstocks to Biofuels, Value-Added Products, and Usable Power; Springer: Berlin/Heidelberg, Germany, 2018; Chapter 8; ISBN 978-3-319-74457-5. [Google Scholar]
- Khaire, K.C.; Patel, S.; Gavande, P.V.; Moholkar, V.A.S.; Goyal, A. Extremophilic biofilms: Exploring the prospects. In Introduction to Biofilm Engineering; ACS Symposium Series; American Chemical Society: Washington, DC, USA, 2019; Volume 1323, pp. 141–157. ISBN 978-0-8412-3473-4. [Google Scholar]
- Castenholz, R.W. Thermophilic blue-green algae and the thermal environment. Bacteriol. Rev. 1969, 33, 476–504. [Google Scholar] [CrossRef] [PubMed]
- Brock, T.D. Thermophilic Microorganisms and Life at High Temperatures; Springer Series in Microbiology; Springer: New York, NY, USA, 1978; ISBN 978-1-4612-6286-2. [Google Scholar]
- Kublanov, I.V.; Perevalova, A.A.; Slobodkina, G.B.; Lebedinsky, A.V.; Bidzhieva, S.K.; Kolganova, T.V.; Kaliberda, E.N.; Rumsh, L.D.; Haertlé, T.; Bonch-Osmolovskaya, E.A. Biodiversity of thermophilic prokaryotes with hydrolytic activities in hot springs of Uzon Caldera, Kamchatka (Russia). Appl. Environ. Microbiol. 2009, 75, 286–291. [Google Scholar] [CrossRef] [PubMed]
- DeCastro, M.-E.; Rodríguez-Belmonte, E.; González-Siso, M.-I. Metagenomics of thermophiles with a focus on discovery of novel thermozymes. Front. Microbiol. 2016, 7, 1521. [Google Scholar] [CrossRef]
- López-López, O.; Cerdán, M.E.; González-Siso, M.-I. Hot spring metagenomics. Life 2013, 3, 308–320. [Google Scholar] [CrossRef]
- Oliverio, A.M.; Power, J.F.; Washburne, A.; Cary, S.C.; Stott, M.B.; Fierer, N. The ecology and diversity of microbial eukaryotes in geothermal springs. ISME J. 2018, 12, 1918–1928. [Google Scholar] [CrossRef]
- Nakamura, Y.; Kaneko, T.; Sato, S.; Ikeuchi, M.; Katoh, H.; Sasamoto, S.; Watanabe, A.; Iriguchi, M.; Kawashima, K.; Ki-mura, T.; et al. Complete genome structure of the thermophilic cyanobacterium Thermosynechococcus elongatus BP-1. DNA Res. 2002, 9, 123–130. [Google Scholar] [CrossRef]
- Slobodkina, G.; Allioux, M.; Merkel, A.; Cambon-Bonavita, M.A.; Alain, K.; Jebbar, M.; Slobodkin, A. Physiological and genomic characterization of a hyperthermophilic archaeon Archaeoglobus neptunius sp. nov. isolated from a deep-sea hydrothermal vent warrants the reclassification of the genus Archaeoglobus. Front. Microbiol. 2021, 16, 679245. [Google Scholar] [CrossRef]
- Hu, X. Ciliates in extreme environments. J. Eukaryot. Microbiol. 2014, 61, 410–418. [Google Scholar] [CrossRef]
- Hickey, D.A.; Singer, G.A. Genomic and proteomic adaptations to growth at high temperature. Genome Biol. 2004, 5, 117. [Google Scholar] [CrossRef]
- Kizilova, A.; Sukhacheva, M.; Pimenov, N.; Yurkov, A.; Kravchenko, I. Methane oxidation activity and diversity of aerobic methanotrophs in pH-neutral and semi-neutral thermal springs of the Kunashir Island, Russian Far East. Extremophiles 2014, 18, 207–218. [Google Scholar] [CrossRef]
- Rozanov, A.S.; Korzhuk, A.V.; Shlyakhtun, V.N.; Shipova, A.A.; Peltek, S.E. Metagenomic data on prokaryotic diversity of Kunashir Island geothermal spring. Data Brief 2020, 32, 106244. [Google Scholar] [CrossRef] [PubMed]
- Rozanov, A.S.; Korzhuk, A.V.; Shekhovtsov, S.V.; Vasiliev, G.V.; Peltek, S.E. Microorganisms of two thermal pools on Kunashir Island, Russia. Biology 2021, 10, 924. [Google Scholar] [CrossRef] [PubMed]
- Kalacheva, E.G.; Taran, Y.A.; Kotenko, T.A.; Inguaggiato, S.; Voloshina, E.V. The hydrothermal system of Mendeleev Volcano, Kunashir Island, Kuril Islands: The geochemistry and the transport of magmatic components. J. Volcanolog. Seismol. 2017, 11, 335–352. [Google Scholar] [CrossRef]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef] [PubMed]
- Takai, K.; Horikoshi, K. Rapid detection and quantification of members of the archaeal community by quantitative PCR using fluorogenic probes. Appl. Environ. Microbiol. 2000, 66, 5066–5072. [Google Scholar] [CrossRef]
- Stoeck, T.; Bass, D.; Nebel, M.; Christen, R.; Jones, M.D.M.; Breiner, H.-W.; Richards, T.A. Multiple marker parallel tag environmental DNA sequencing reveals a highly complex eukaryotic community in marine anoxic water. Mol. Ecol. 2010, 19 (Suppl. 1), 21–31. [Google Scholar] [CrossRef]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef]
- Callahan, B.; McMurdie, P.; Holmes, S. Exact sequence variants should replace operational taxonomic units in marker-gene data analysis. ISME J. 2017, 11, 2639–2643. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- McMurdie, P.J.; Holmes, S. Phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef]
- Wickham, H.; Chang, W.; Henry, L.; Pedersen, T.; Takahashi, K.; Wilke, C.; Woo, K.; Yutani, H.; Dunnington, D. RStudio. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Xu, S.; Yu, G. Microbiota Process: A Comprehensive R Package for Managing and Analyzing Microbiome and Other Ecological Data within the Tidy Framework. R Package Version 1.8.1. Available online: https://github.com/YuLab-SMU/MicrobiotaProcess (accessed on 29 December 2022).
- Lahti, L.; Shetty, S. Tools for Microbiome Analysis in R. R PackageVersion 1.19.1. Available online: http://microbiome.github.com/microbiome (accessed on 29 December 2022).
- Huber, R.; Stetter, K.O. Fervidobacterium. In Bergey’s Manual of Systematics of Archaea and Bacteria; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2015; pp. 1–5. ISBN 978-1-118-96060-8. [Google Scholar]
- Suda, S.; Watanabe, M.M.; Otsuka, S.; Mahakahant, A.; Yongmanitchai, W.; Nopartnaraporn, N.; Liu, Y.; Day, J.G. Taxonomic revision of water-bloom-forming species of oscillatorioid cyanobacteria. Int. J. Syst. Evol. Microbiol. 2002, 52 Pt 5, 1577–1595. [Google Scholar] [CrossRef] [PubMed]
- Kato, S.; Itoh, T.; Yamagishi, A. Archaeal diversity in a terrestrial acidic spring field revealed by a novel PCR primer targeting archaeal 16S rRNA genes. FEMS Microbiol. Lett. 2014, 319, 34–43. [Google Scholar] [CrossRef] [PubMed]
- Baumgartner, M.; Stetter, K.O.; Foissner, W. Morphological, small subunit rRNA, and physiological characterization of Trimyema minutum (Kahl, 1931), an anaerobic ciliate from submarine hydrothermal vents growing from 28 °C to 52 °C. J. Eukaryot. Microbiol. 2002, 49, 227–238. [Google Scholar] [CrossRef] [PubMed]
- Sharp, C.E.; Brady, A.L.; Sharp, G.H.; Grasby, S.E.; Stott, M.B.; Dunfield, P.F. Humboldt’s Spa: Microbial diversity Is controlled by temperature in geothermal environments. ISME J. 2014, 8, 1166–1174. [Google Scholar] [CrossRef]
- Chiriac, C.M.; Szekeres, E.; Rudi, K.; Baricz, A.; Hegedus, A.; Dragoş, N.; Coman, C. Differences in temperature and water chemistry shape distinct diversity patterns in thermophilic microbial communities. Appl. Environ. Microbiol. 2017, 83, e01363-17. [Google Scholar] [CrossRef]
- Chan, C.S.; Chan, K.-G.; Ee, R.; Hong, K.-W.; Urbieta, M.S.; Donati, E.R.; Shamsir, M.S.; Goh, K.M. Effects of physiochemical factors on prokaryotic biodiversity in Malaysian circumneutral hot springs. Front. Microbiol. 2017, 8, 1252. [Google Scholar] [CrossRef] [Green Version]
- Power, J.F.; Carere, C.R.; Lee, C.K.; Wakerley, G.L.J.; Evans, D.W.; Button, M.; White, D.; Climo, M.D.; Hinze, A.M.; Morgan, X.C.; et al. Microbial biogeography of 925 geothermal springs in New Zealand. Nat. Commun. 2018, 9, 2876. [Google Scholar] [CrossRef]
- Massello, F.L.; Chan, C.S.; Chan, K.-G.; Goh, K.M.; Donati, E.; Urbieta, M.S. Meta-analysis of microbial communities in hot springs: Recurrent taxa and complex shaping factors beyond pH and temperature. Microorganisms 2020, 8, 906. [Google Scholar] [CrossRef]
- Podar, P.T.; Yang, Z.; Björnsdóttir, S.H.; Podar, M. Comparative Analysis of microbial diversity across temperature gradients in hot springs from Yellowstone and Iceland. Front. Microbiol. 2020, 11, 1625. [Google Scholar] [CrossRef]
- Escuder-Rodríguez, J.-J.; DeCastro, M.-E.; Saavedra-Bouza, A.; Becerra, M.; González-Siso, M.-I. Insights on microbial communities inhabiting non-volcanic hot springs. Int. J. Mol. Sci. 2022, 23, 12241. [Google Scholar] [CrossRef] [PubMed]
- Kubo, K.; Knittel, K.; Amann, R.; Fukui, M.; Matsuura, K. Sulfur-metabolizing bacterial populations in microbial mats of the Nakabusa hot spring, Japan. Syst. Appl. Microbiol. 2011, 34, 293–302. [Google Scholar] [CrossRef]
- Inskeep, W.P.; Rusch, D.B.; Jay, Z.J.; Herrgard, M.J.; Kozubal, M.A.; Richardson, T.H.; Macur, R.E.; Hamamura, N.; Jennings, R.D.; Fouke, B.W. Metagenomes from high-temperature chemotrophic systems reveal geochemical controls on microbial community structure and function. PLoS ONE 2010, 5, e9773. [Google Scholar] [CrossRef] [PubMed]
- Merkel, A.; Pimenov, N.; Rusanov, I.; Slobodkin, A.; Slobodkina, G.; Tarnovetckii, I.; Frolov, E.; Dubin, A.; Perevalova, A.; Bonch-Osmolovskaya, E. Microbial diversity and autotrophic activity in Kamchatka hot springs. Extremophiles 2017, 21, 307–317. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, T.; Fukui, M. Molecular characterization of community structures and sulfur metabolism within microbial streamers in Japanese hot springs. Appl. Environ. Microbiol. 2003, 69, 7044–7057. [Google Scholar] [CrossRef]
- Hetzer, A.; Morgan, H.; Mcdonald, I.; Daughney, C. Microbial life in champagne pool, a geothermal spring in Waiotapu, New Zealand. Extremophiles 2007, 11, 605–614. [Google Scholar] [CrossRef]
- Sahm, K.; John, P.; Nacke, H.; Wemheuer, B.; Grote, R.; Daniel, R.; Antranikian, G. High abundance of heterotrophic prokaryotes in hydrothermal springs of the Azores as revealed by a network of 16S rRNA gene-based methods. Extremophiles 2013, 17, 649–662. [Google Scholar] [CrossRef]
- Dong, Y.; Sanford, R.A.; Inskeep, W.P.; Srivastava, V.; Bulone, V.; Fields, C.J.; Yau, P.M.; Sivaguru, M.; Ahrén, D.; Fouke, K.W.; et al. Physiology, metabolism, and fossilization of hot-spring filamentous microbial mats. Astrobiology 2019, 19, 1442–1458. [Google Scholar] [CrossRef] [PubMed]
- Bowen De León, K.; Gerlach, R.; Peyton, B.M.; Fields, M.W. Archaeal and bacterial communities in three alkaline hot springs in Heart Lake geyser basin, Yellowstone National Park. Front. Microbiol. 2013, 4, 330. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Hou, W.; Dong, H.; Jiang, H.; Huang, L.; Wu, G.; Zhang, C.; Song, Z.; Zhang, Y.; Ren, H.; et al. Control of temperature on microbial community structure in hot springs of the Tibetan Plateau. PLoS ONE 2013, 8, e62901. [Google Scholar] [CrossRef]
- Thiel, V.; Wood, J.M.; Olsen, M.T.; Tank, M.; Klatt, C.G.; Ward, D.M.; Bryant, D.A. The dark side of the Mushroom Spring microbial Mat: Life in the Shadow of Chlorophototrophs. I. Microbial diversity based on 16S rRNA gene amplicons and metagenomic sequencing. Front. Microbiol. 2016, 17, 919. [Google Scholar] [CrossRef] [PubMed]
- Rozanov, A.S.; Bryanskaya, A.V.; Ivanisenko, T.V.; Malup, T.K.; Peltek, S.E. Biodiversity of the microbial mat of the Garga hot spring. BMC Evol. Biol. 2017, 17, 254. [Google Scholar] [CrossRef] [PubMed]
- Shu, W.-S.; Huang, L.-N. Microbial diversity in extreme environments. Nat. Rev. Microbiol. 2022, 20, 219–235. [Google Scholar] [CrossRef]
- Klatt, C.G.; Inskeep, W.P.; Herrgard, M.J.; Jay, Z.J.; Rusch, D.B.; Tringe, S.G.; Niki Parenteau, M.; Ward, D.M.; Boomer, S.M.; Bryant, D.A.; et al. Community structure and function of high-temperature chlorophototrophic microbial mats inhabiting diverse geothermal environments. Front. Microbiol. 2013, 4, 106. [Google Scholar] [CrossRef]
- Darima, B.; Buryukhaev, S.; Dambaev, V.; Tsyrenova, D.; Lavrentieva, E. Taxonomical and functional diversity of microbial communities in two hot springs of the Baikal rift zone. IOP Conf. Ser. Earth Environ. Sci. 2021, 908, 012001. [Google Scholar] [CrossRef]
- Boomer, S.M.; Noll, K.L.; Geesey, G.G.; Dutton, B.E. Formation of multilayered photosynthetic biofilms in an alkaline thermal spring in Yellowstone National Park, Wyoming. Appl. Environ. Microbiol. 2009, 75, 2464–2475. [Google Scholar] [CrossRef]
- Nübel, U.; Bateson, M.; Vandieken, V.; Wieland, A.; Kühl, M.; Ward, D. Microscopic examination of distribution and phenotypic properties of phylogenetically diverse Chloroflexaceae-related bacteria in hot spring microbial matst. Appl. Environ. Microbiol. 2002, 68, 4593–4603. [Google Scholar] [CrossRef] [Green Version]
- Bennett, A.C.; Murugapiran, S.K.; Hamilton, T.L. Temperature impacts community structure and function of phototrophic Chloroflexi and Cyanobacteria in two alkaline hot springs in Yellowstone National Park. Environ. Microbiol. Rep. 2020, 12, 503–513. [Google Scholar] [CrossRef] [PubMed]
- Tank, M.; Thiel, V.; Ward, D.; Bryant, D. A Panoply of Phototrophs: An Overview of the Thermophilic Chlorophototrophs of the Microbial Mats of Alkaline Siliceous Hot Springs in Yellowstone National Park, WY, USA; Springer International Publishing: Berlin/Heidelberg, Germany, 2017; pp. 87–137. [Google Scholar] [CrossRef]
- Hamilton, T.; Havig, J. Meet me in the middle: Median temperatures impact Cyanobacteria and photoautotrophy in eruptive Yellowstone hot springs. mSystems 2022, 7, e01450-21. [Google Scholar] [CrossRef] [PubMed]
- Mammola, S.; Meierhofer, M.B.; Borges, P.A.V.; Colado, R.; Culver, D.C.; Deharveng, L.; Delić, T.; Di Lorenzo, T.; Dražina, T.; Ferreira, R.L.; et al. Towards evidence-based conservation of subterranean ecosystems. Biol. Rev. Camb. Philos. Soc. 2022, 97, 1476–1510. [Google Scholar] [CrossRef] [PubMed]
- Yamaoka, T.; Satoh, K.; Katoh, S. Photosynthetic activities of a thermophilic blue-green alga. Plant Cell Physiol. 1978, 19, 943–954. [Google Scholar] [CrossRef]
- Katoh, H.; Itoh, S.; Shen, J.R.; Ikeuchi, M. Functional analysis of psbV and a novel c-type cytochrome gene psbV2 of the thermophilic cyanobacterium Thermosynechococcus elongatus strain BP-1. Plant Cell Physiol. 2001, 42, 599–607. [Google Scholar] [CrossRef]
- Honda, D.; Yokota, A.; Sugiyama, J. Detection of seven major evolutionary lineages in cyanobacteria based on the 16S rRNA gene sequence analysis with new sequences of five marine Synechococcus strains. J. Mol. Evol. 1999, 48, 723–739. [Google Scholar] [CrossRef]
- Klatt, C.G.; Liu, Z.; Ludwig, M.; Kühl, M.; Jensen, S.I.; Bryant, D.A.; Ward, D.M. Temporal metatranscriptomic patterning in phototrophic Chloroflexi inhabiting a microbial mat in a geothermal spring. ISME J. 2013, 7, 1775–1789. [Google Scholar] [CrossRef]
- Brigmon, R.L.; Furlong, M.; Whitman, W.B. Identification of Thiothrix unzii in two distinct ecosystems. Lett. Appl. Microbiol. 2003, 36, 88–91. [Google Scholar] [CrossRef]
- Bauer, F.; Stix, M.; Bartha-Dima, B.; Geist, J.; Raeder, U. Spatio-temporal monitoring of benthic anatoxin-a-producing Tychonema sp. in the River Lech, Germany. Toxins 2022, 14, 357. [Google Scholar] [CrossRef]
- Alcamán-Arias, M.; Pedrós-Alió, C.; Tamames, J.; Fernández, C.; Pérez-Pantoja, D.; Vásquez, M.; Díez, B. Diurnal changes in active carbon and nitrogen pathways along the temperature gradient in porcelana hot spring microbial mat. Front. Microbiol. 2018, 9, 2353. [Google Scholar] [CrossRef]
- Lehr, C.R.; Frank, S.D.; Norris, T.B.; D’Imperio, S.; Kalinin, A.V.; Toplin, J.A.; Castenholz, R.W.; McDermott, T.R. Cyanidia (cyanidiales) population diversity and dynamics in an acid-sulfate-chloride spring in Yellowstone National Park. J. Phycol. 2007, 43, 3–14. [Google Scholar] [CrossRef]
- Rampelotto, P.H. Extremophiles and extreme environments. Life 2013, 3, 482–485. [Google Scholar] [CrossRef]
- Groussin, M.; Gouy, M. Adaptation to environmental temperature is a major determinant of molecular evolutionary rates in Archaea. Mol. Biol. Evol. 2011, 28, 2661–2674. [Google Scholar] [CrossRef] [PubMed]
- Yim, L.C.; Hongmei, J.; Aitchison, J.C.; Pointing, S.B. Highly diverse community structure in a remote central Tibetan geothermal spring does not display monotonic variation to thermal stress. FEMS Microbiol. Ecol. 2006, 57, 80–91. [Google Scholar] [CrossRef] [PubMed]
- Zhao-Qi, S.; Chen, J.-Q.; Jiang, H.; Zhou, E.-M.; Tang, S.-K.; Zhi, X.-Y.; Zhang, L.-X.; Zhang, L.; Li, W.-J. Diversity of Crenarchaeota in terrestrial hot springs in Tengchong, China. Extremophiles 2010, 14, 287–296. [Google Scholar] [CrossRef]
- Nunoura, T.; Takaki, Y.; Kakuta, J.; Nishi, S.; Sugahara, J.; Kazama, H.; Chee, G.J.; Hattori, M.; Kanai, A.; Atomi, H.; et al. Insights into the evolution of Archaea and eukaryotic protein modifier systems revealed by the genome of a novel archaeal group. Nucleic Acids Res. 2011, 39, 3204–3223. [Google Scholar] [CrossRef]
- Kimura, H.; Mori, K.; Tashiro, T.; Kato, K.; Yamanaka, T.; Ishibashi, J.I.; Hanada, S. Culture-independent estimation of optimal and maximum growth temperatures of archaea in subsurface habitats based on the G+C content in 16S rRNA gene sequences. Geomicrobiol. J. 2010, 27, 114–122. [Google Scholar] [CrossRef]
- Stefanova, K.; Tomova, I.; Tomova, A.; Radchenkova, N.; Atanassov, I.; Kambourova, M. Archaeal and bacterial diversity in two hot springs from geothermal regions in Bulgaria as demostrated by 16S rRNA and GH-57 genes. Int. Microbiol. 2015, 18, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Takami, H.; Arai, W.; Takemoto, K.; Uchiyama, I.; Taniguchi, T. Functional classification of uncultured “Candidatus Caldiarchaeum subterraneum” using the maple system. PLoS ONE 2015, 21, e0132994. [Google Scholar] [CrossRef] [Green Version]
- Rinke, C.; Chuvochina, M.; Mussig, A.J.; Chaumeil, P.A.; Davín, A.A.; Waite, D.W.; Whitman, W.B.; Parks, D.H.; Hugenholtz, P. A standardized archaeal taxonomy for the Genome Taxonomy Database. Nat. Microbiol. 2021, 6, 946–959. [Google Scholar] [CrossRef] [PubMed]
- Stetter, K.O. Archaeoglobus fulgidus gen. Nov., sp. nov.: A new taxon of extremely thermophilic Archaebacteria. Syst. Appl. Microbiol. 1988, 10, 172–173. [Google Scholar] [CrossRef]
- Burggraf, S.; Jannasch, H.W.; Nicolaus, B.; Stetter, K.O. Archaeoglobus profundus sp. nov., represents a new species within the sulfur-reducing Archaebacteria. Syst. Appl. Microbiol. 1990, 13, 24–28. [Google Scholar] [CrossRef]
- Tansey, M.R.; Brock, T.D. The upper temperature limit for eukaryotic organisms. Proc. Natl. Acad. Sci. USA 1972, 69, 2426–2428. [Google Scholar] [CrossRef] [PubMed]
- Brown, P.B.; Wolfe, G.V. Protist genetic diversity in the acidic hydrothermal environments of Lassen Volcanic National Park, USA. J. Eukaryot. Microbiol. 2006, 53, 420–431. [Google Scholar] [CrossRef] [PubMed]
- Baumgartner, M.; Yapi, A.; Gröbner-Ferreira, R.; Stetter, K.O. Cultivation and properties of Echinamoeba thermarum n. sp., an extremely thermophilic amoeba thriving in hot springs. Extremophiles 2003, 7, 267–274. [Google Scholar] [CrossRef]
- Kahan, D. Cyclidium citrullus Cohn, a ciliate from the hot springs of Tiberias (Israel). J. Protozool. 1972, 19, 593–597. [Google Scholar] [CrossRef]
- Qu, Z.; Groben, R.; Marteinsson, V.; Agatha, S.; Filker, S.; Stoeck, T. Redescription of Dexiotricha colpidiopsis (Kahl, 1926) Jankowski, 1964 (Ciliophora, Oligohymenophorea) from a hot spring in Iceland with identification key for Dexiotricha species. Acta Protozool. 2019, 57, 95–106. [Google Scholar] [CrossRef]
- Zhu, R.; Qu, Z.; Zhang, Q.; Filker, S.; Stoeck, T.; Li, F.; Hu, X. A New record of Oxytricha granulifera granulifera Foissner and Adam, 1983 (Protozoa, Ciliophora, Oxytrichidae) from a hot spring in Iceland, with notes on its abnormal form during cultivation. Front. Marin. Sci. 2021, 8, 621349. [Google Scholar] [CrossRef]
- Roy, C.; Rameez, M.J.; Haldar, P.K.; Peketi, A.; Mondal, N.; Bakshi, U.; Mapder, T.; Pyne, P.; Fernandes, S.; Bhattacharya, S.; et al. Microbiome and ecology of a hot spring-microbialite system on the Trans-Himalayan Plateau. Sci. Rep. 2020, 10, 5917. [Google Scholar] [CrossRef] [Green Version]
- Takeshita, K.; Yamada, T.; Kawahara, Y.; Narihiro, T.; Ito, M.; Kamagata, Y.; Shinzato, N. Tripartite symbiosis of an anaerobic scuticociliate with two hydrogenosome-associated endosymbionts, a Holospora-related alphaproteobacterium and a methanogenic archaeon. Appl. Environ. Microbiol. 2019, 85, e00854-19. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Name | Latitude, North | Longitude, East | Temperature, °C | Location | Morphology |
|---|---|---|---|---|---|
| StB1 | 44.007009 | 145.683265 | 79.5–81.5 | 15–30 cm away from the hot spring 1 | White biofilm with thick filamentous structure |
| StB2 | 44.006997 | 145.683311 | 50.0–58.5 | Pool of the hot spring 2 | Layered structure of dark green, brown and orange brick color |
| StB3 | 44.007046 | 145.683308 | 18.0/46.0 | Main stream (thermocline zone) | Thin brownish filamentous structure |
| StB4 | 44.007092 | 145.683204 | 67.0–69.0 | 4 m downstream StB5 | Layered structure, similar to StB2 biofilm |
| StB5 | 44.007025 | 145.683300 | 69.0–70.2 | Mixing point of two hot springs | Layered structure, almost transparent and resembled a rigid and dense jelly |
| StB6 | 44.007186 | 145.683210 | 17.0 | Main stream bed | Amorphous, brown-black, and covered with gas bubbles |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Malygina, A.; Balkin, A.; Polyakova, E.; Stefanov, S.; Potekhin, A.; Gogoleva, N. Taxonomic Diversity of the Microbial Biofilms Collected along the Thermal Streams on Kunashir Island. Ecologies 2023, 4, 106-123. https://doi.org/10.3390/ecologies4010009
Malygina A, Balkin A, Polyakova E, Stefanov S, Potekhin A, Gogoleva N. Taxonomic Diversity of the Microbial Biofilms Collected along the Thermal Streams on Kunashir Island. Ecologies. 2023; 4(1):106-123. https://doi.org/10.3390/ecologies4010009
Chicago/Turabian StyleMalygina, Alexandra, Alexander Balkin, Elena Polyakova, Sergey Stefanov, Alexey Potekhin, and Natalia Gogoleva. 2023. "Taxonomic Diversity of the Microbial Biofilms Collected along the Thermal Streams on Kunashir Island" Ecologies 4, no. 1: 106-123. https://doi.org/10.3390/ecologies4010009