
Effects of Deadwood on Macroinvertebrate Assemblages in Three Sand-Type Lowland Streams

1
Department of Water and Waste Management, University of Applied Sciences Magdeburg-Stendal, Breitscheidstr. 2, D-39114 Magdeburg, Germany
2
School of Public Health, San Diego State University, San Diego, CA 92182, USA
*
Author to whom correspondence should be addressed.
Ecologies 2023, 4(1), 88-105; https://doi.org/10.3390/ecologies4010008
Submission received: 3 December 2022
/
Revised: 26 January 2023
/
Accepted: 30 January 2023
/
Published: 9 February 2023
(This article belongs to the Special Issue The Ecology of Rivers, Floodplains and Oxbow Lakes)
Abstract
:The objective of the manuscript was to study the effects of dead wood on macroinvertebrates (MI) assemblages occupying different habitat patches. Three streams were studied, with control and wooded sections in each stream from which MI were sampled for all substrate types (wood, fine detritus, gravel, sand). This contribution deals with three German streams Ihle, Nieplitz, and Verlorenwasser, which belong to the stream type 14 (sand-dominated lowland stream) and are located in the landscape unit Fläming. This type of stream, where large wood could be most influential, has been relatively understudied. This study focused on quantifying substrate distribution and macroinvertebrate taxonomy and performing statistical analysis of MI densities with regard to substrate type. The area-specific invertebrate assemblages were differentiated for MI density, dry matter density and taxonomic richness of invertebrates. In addition to macroinvertebrates, a special focus was on aquatic and semi-aquatic insects. There were significant differences between the control and wooden sections. The macroinvertebrate assemblages (individual density, dry matter density and species richness) of the detritus of the wood sections only in the Verlorenwasser was significantly and positively affected (two or three times) by the deadwood. Detritus in the wood sections of the Nieplitz and Verlorenwasser contained more sensitive species like EPT than detritus from the control sections. On the other hand, no significant difference in MI populations were determined for the sand of the wood sections compared to the sand of the control sections. In all water bodies, the wood substrate promoted all investigated indicators of macrozoobenthos as compared to sand of the wood sections. Detritus significantly promoted macrozoobenthos colonization in the wood sections as compared to sand of the Nieplitz and the Verlorenwasser. The MI density and dry matter density in the Ihle was also significantly promoted by detritus, although not significantly with regard species richness. Two significant correlations were also identified. The more sand that was present in the Ihle and the Verlorenwasser, the more the macroinvertebrate assemblages were inhibited. Additionally, the larger the area of detritus in the Ihle and the area of wood in the Nieplitz, the more MI populations were encouraged. The influence of the deadwood position on MI was also investigated, but there was no significant difference. Incline bridges, flow guides and parallel wood position types were settled in similar quality and quantity.
1. Introduction
Insects, such as dragonflies, mayflies, and caddisflies, go through an aquatic stage before settling on the land. According to the “Krefeld study”, a decrease in the total biomass of flying insects by more than 75% was observed over a period of 27 years in several German protected areas [1]. It is also conceivable that the dramatic loss of insect species and biomass worldwide has been underestimated [2].
The impoverishment of the landscape structure through the dredging and straightening of water bodies, intensive maintenance measures (such as the removal of deadwood structures), eutrophication, and the entry of pesticides are major stress factors [2,3]. In a lot of cases, macroinvertebrates (MI) including aquatic stages of insects, no longer find suitable habitat conditions (such as deadwood) due to changed abiotic conditions. A decline in insects, the most species-rich animal class in terms of species numbers and population sizes, directly impacts the ecosystem’s status and the populations of animal and plant species [2]. For example, the individual density and biomass of MI are important for the structure and development of fish populations [4,5].
This research aimed to determine how deadwood in its various basic types affects MI distribution in streams.
Under the European Water Framework Directive (WFD), all water bodies must be developed to a good ecological status (GES) by 2027. However, the metrics for defining GES only consider species composition and abundance and not population density and biomass. Accordingly, the following hypotheses were tested using statistical analyses of three metrics, population density, dry matter density and species richness of MI and insect assemblages.
- Deadwood (DW) serves as a food source for MI, increases habitat and species diversity, and individual density [6]. Compared to sand, wood and detritus promote area-specific assemblages by MI in general and by aquatic insects specifically.
- The MI assemblages of detritus are higher in wood sections compared to the control sections. In contrast wood has no effect on MI assemblages of the sand of wood sections, so the sand of wood sections is not significantly different from that of control sections.
- The percentage of substrates per unit area directly influences the individual density and the dry matter of a water body section. Each section of wood and detritus promotes the total MI assemblages in a stream (including all substrates), while sand inhibits species richness and density of individuals.
- Wood promotes MI assemblages in a water segment (including detritus, sand and gravel) in contrast to a section of water without wood (control section).
This study derives remediation measures based on the hypotheses examined. For this reason, our work examines whether MI assemblages are dependent on wood structure type (incline bridges, flow guides and parallel wood position).
2. Material and Methods
2.1. Project Area
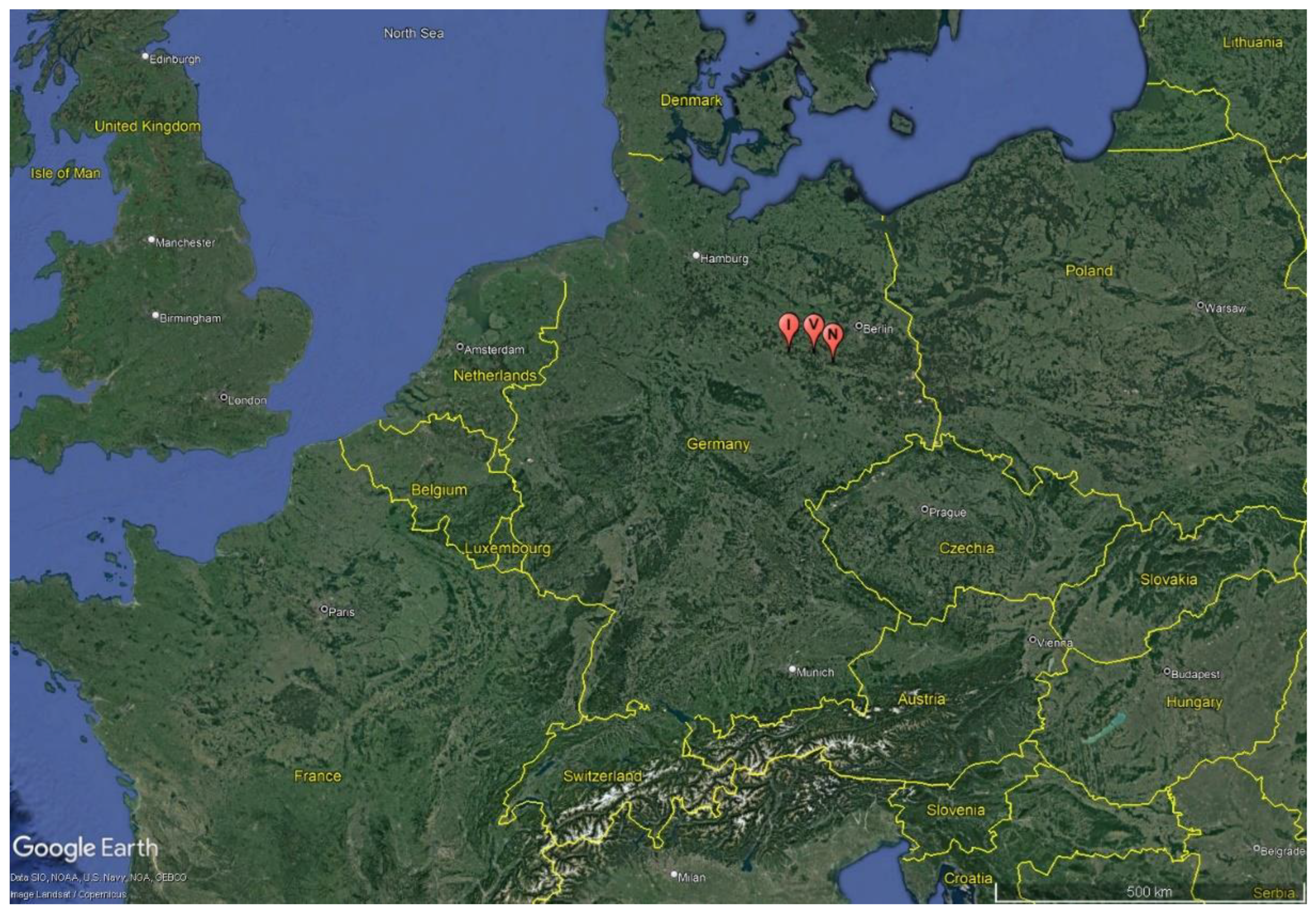
Three streams, the Ihle (I), Nieplitz (N) and Verlorenwasser (V) were studied. These are all located in the North German lowlands, the Fläming (Figure 1). The Fläming is a mountain range shaped by the Ice Age with landscape forms such as headwaters, deep meadow valleys and mixed forests. The “Hoher Fläming natural park” is a dense network of watercourses with intact upper reaches. In the surrounding area of the Nieplitz and Verlorenwasser streams, mixed deciduous and coniferous forests and small grasslands dominate, and wet grassland and deciduous forest dominate around the Ihle. The banks of these streams are settled by alder trees (Alnus glutinosa) which shade large parts of the water body. Arable land or human settlements are rare in these areas so urban development and agriculture have minimum influence in these catchment areas [4].
All three water bodies are sand- and gravel-dominated lowland streams. The test stretches are permanently flowing waters of the Rithral and belong to LAWA stream type 14 “sand-type lowland streams” [9,10]. The watercourses have a near-natural, predominantly meandering course and a near-natural cross profile. The catchment area is at least 10 km2. The saprobity (organic load) is ‘good’ and ‘better’ which rule out chemical stress as an additional and over-laying stressor. Species inventory and recolonization potential is as “intact” as possible [4].
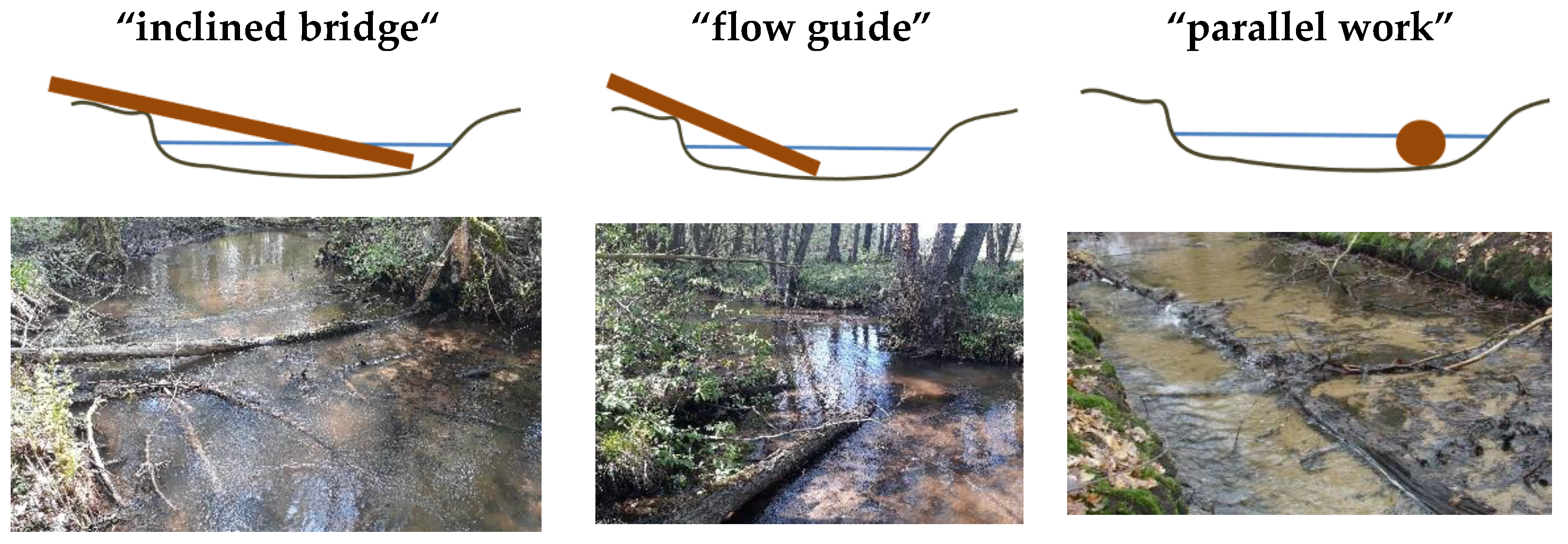
Wood structures caused by self-dynamics and natural input are present in all three streams. However, the Ihle additionally contains some wood that was brought in as part of restoration measures. These wooden structures were designed in such a way (e.g., felling of a whole tree) that they are similar to the natural wooden structures in the other two streams. The proportion of wood in the total bottom surface is at least seven percent. The control sections were between the wood sections, have similar boundary conditions and contained no or only little wood. The wood sections have inclined bridges, flow guides and parallel wood position types (Figure 2).
2.2. Methods
2.2.1. Sampling and Taxonomies
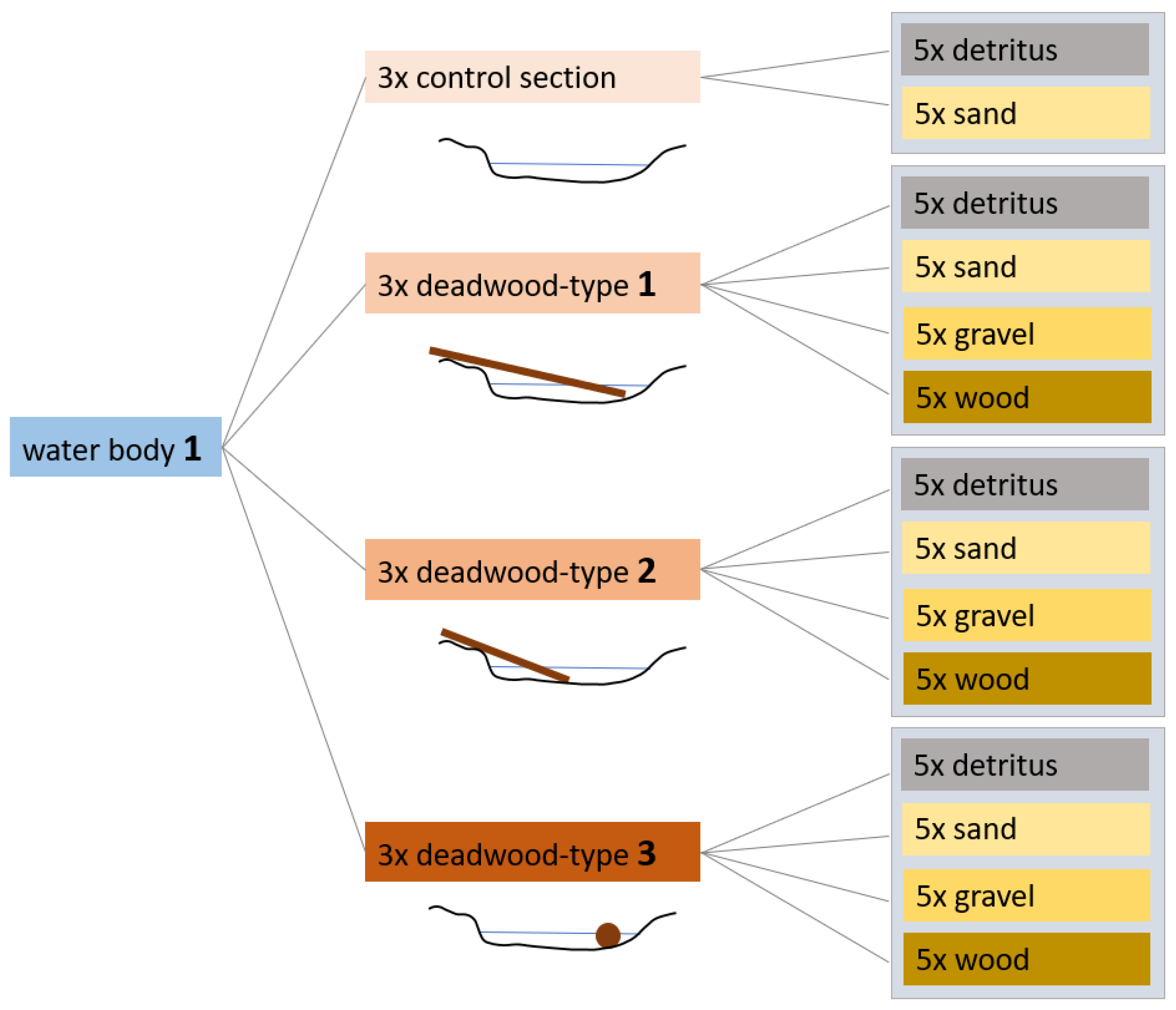
The three streams were sampled in April 2019, with three control (CS) and nine wooded section (WS) per stream. In WS, the three wood structure types were separated with three replicates each. During each sampling, the associated substrates, including the MI, in the sections were sampled: wood, detritus, sand, and gravel. Five representative sub-samples were taken for each substrate in a section and combined into a composite sample (Figure 3). A total of 27 substrate samples were analyzed from the 3 different wood sections in Nieplitz and Verlorenwasser. Additionally, because gravel is present in the stream Ihle, a total of 33 samples were analyzed in these three wood sections. The 3 replicates in the CS per stream each consisted of three detritus and three sand substrate samples. The study was conducted according to the guidelines of the Declaration of Helsinki, and approved by the Institutional Review Board of the Institute for Water Management and Ecotechnology by date of 1 September 2022 (Protocol Code: 09-01-2022-001).
For a detailed description of the MI settlement, it is necessary to include all available substrates and their areas. The considered substrates’ surface proportions were measured on site in the sub-areas of the streams, and the types of wood structures were as documented. The section’s length and average width were measured for a percentage estimation of the individual substrates’ share of area in a given section.
MI organisms were sampled with a surber (mesh size 500 µm, net depth 70 cm). A sub-sample covered an area of 25 cm × 25 cm (the projected frame dimensions of the landing net; 0.0625 m2). Sand and gravel were sampled down to a depth of 3–5 cm, detritus down to the underlying mineral layer (depending on the layer 1–10 cm) and wood (including attached branches and foliage) at the surface. The samples were immediately fixed with 70% alcohol.
Organisms from the sampled substrate were wholly sorted out with tweezers and counted. In the case of taxonomic groups with a high number of individuals, such as Chironomidae Gen. sp. or Pisidium sp., these were sorted out in a sub-sample and all other taxa were extrapolated to the overall sample (by area reference).
The determination of the organisms was carried out with the binocular “Nikon SMZ 800” and by using determination literature according to [13,14,15,16,17,18]. In addition to macroinvertebrates, a special focus was on the aquatic and semi-aquatic insects. Except for most Diptera, the organisms were identified down to species level.
In the further evaluation, the aquatic insects and the EPT (Ephemeroptera, Plecoptera, Trichoptera) taxa were emphasized among the MI, since it has been shown that these groups are particularly suitable as bioindicators due to their high sensitivity [19].
2.2.2. Measurement of Dry Matter
For dry matter determination, individual samples were grouped into processing units according to their habitat affiliation, e.g., the detritus samples from the Ihle wood sections were combined for each taxon. The caddis of caddisfly larvae (Trichoptera) were removed before drying. The exception was Sericostoma personatum, because most of these caddises could not be removed. The dry mass of S. personatum larvae and total dry matter (larvae and caddis) were determined in a subsample of the Nieplitz River (WS, D). The larvae had a percentage of 6.53% of the total dry mass. Dry masses of non-removable larvae were approximated by multiplying with the factor of 0.0653 for the species S. personatum.
The samples were then dried in an oven for four hours at 105 °C and cooled in a desiccator. Dry matter was determined by weighing the dried individuals to four decimal places (in g). A mean individual dry weight per taxon was calculated from the dry matter of the pooled samples and multiplied by the number of individuals at a sampling site, including the individuals that were separated due to high abundances (Chironomidae, Pisidium spp.).
2.2.3. Statistics
Non-parametric tests were carried out with the “IBM SPSS Statistics 14.0” program to estimate the differences in the colonization of two individual samples, and boxplots were created.
The substrates were combined as averages in the calculations. For example, substrates from the wood sections (n = 9) were combined as average values and compared with the average values of substrates from the control sections (n = 3).
To determine if the differences in macroinvertebrate composition among the three streams and substrate types were significant, we used the ANOSIM function from the R-Studios Vegan package. The function compares the mean dissimilarity between groups to the mean dissimilarity within groups. A Bray–Curtis distance and 9999 permutations were used as settings for the ANOSIM function. The calculation yields an R value and its significance. An R value close to 0 indicates homogeneous species composition between groups while an R value close to 1 indicates true dissimilarity [20].
Two-tailed nonparametric correlation tests were performed to examine the relationship between two dependent variables. The Spearman test was used for this. The test is based on a rank correlation (ranking of the data; di = rank difference). The Spearman correlation coefficient rsp is calculated using the following formula 1 (n = number of samples):
Spearman’s correlation coefficient rsp can assume values between −1 (high negative correlation) and +1 (high positive correlation). The probability of error of two samples were tested using the Wilcoxon-Mann–Whitney test (Mann–Whitney U).
3. Results
3.1. Stream Bottom
The dominant bottom substrate in all streams is sand, except in the CS of the Ihle where detritus predominates. In Verlorenwasser, the bottom substrate in the CS (n = 3) consists of around 84% sand and is significantly higher than in the WS (n = 9) (60% on average). In the Nieplitz, the percentage of sand in the WS was 65%, which is lower than in the CS (75%). Wood covered 7% of the wood sections in both water bodies (Verlorenwasser Min 3; Max 13; Nieplitz Min 2; Max 15). The detritus percentage was higher in both streams’ WS than in their CS (Table 1).
In Ihle, the degree of sand coverage in the WS was higher than in the CS. That is different from the Verlorenwasser and the Nieplitz. The Ihle is the only sampled stream containing gravel substrates. These occupy 6% (Min 0, Max 14) of the coverage in the WS, with gravel found in six of the nine WS. Wood covers 9% (Min 5; Max 15) and detritus 37% of the wood runs. In the CS of the Ihle, the proportion of detritus, at 58%, was higher than the proportion of sand, at 42% (Table 1).
3.2. MI Colonization in the Streams
For the three streams with a sampled bottom area of 6.56 m2, 111 taxa with 68,671 individuals having a dry matter of 162.59 g (excluding freshwater molluscs) were identified to species level. These belong to 54 families and 14 orders (see Supplementary Materials).
The comparison of the three streams shows that MI populations were very different. The Ihle was populated with 89 taxa with gravel (sampled area of 2.44 m2), respectively, 85 taxa without gravel (sampled area of 2.06 m2). As many as 65 taxa were identified in Nieplitz and the fewest taxa found for the Verlorenwasser (57 taxa). Likewise, most insects (58), and EPT (Ephemeroptera, Plecoptera and Trichoptera; 36) were found in the Ihle (without G). The fewest insects (38) and EPT (23) were collected in the Verlorenwasser (Table 2).
Sand-dominated lowland streams host typical species of sand. These include, for example, the mayfly (Ephemeroptera) Ephemera danica and the stonefly Isoptena serricornis, which colonized the sampled waters. Deadwood and detritus are colonized by caddisflies (Trichoptera) such as Sericostoma personatum, Notidobia ciliaris, Lepidostoma basale, Lype sp., Potamophylax sp., Hydropsyche saxonica, and Micropterna sequax.
Species of all 14 orders occurred in the Ihle. In the Nieplitz, no individuals of the order Heteroptera such as Aphelocheirus aestivalis and Sigara sp. were found, as well as no Turbellaria such as Dugesia gonocephala were found in the Verlorenwasser. One neozoon occurred in each of the water bodies. The snail Potamopyrgus antipodarum, originated from New Zealand, was found in both the Ihle and the Nieplitz. The clam Corbicula fluminea was found in the Verlorenwasser.
The average density of individuals in the Ihle was 6215 individuals per m2. The highest density was in Nieplitz (20,492 individuals per m2), and the lowest density was in the Verlorenwasser (5457 individuals per m2) (Table 2). The average dry matter concentration in the Nieplitz was 56.86 g per m2, higher than both the Ihle (11.05 g per m2) and the Verlorenwasser (8.90 g per m2) (Table 2).
3.3. Species Composition of MI in Dependence on Substrates
3.3.1. Wood and Control Sections
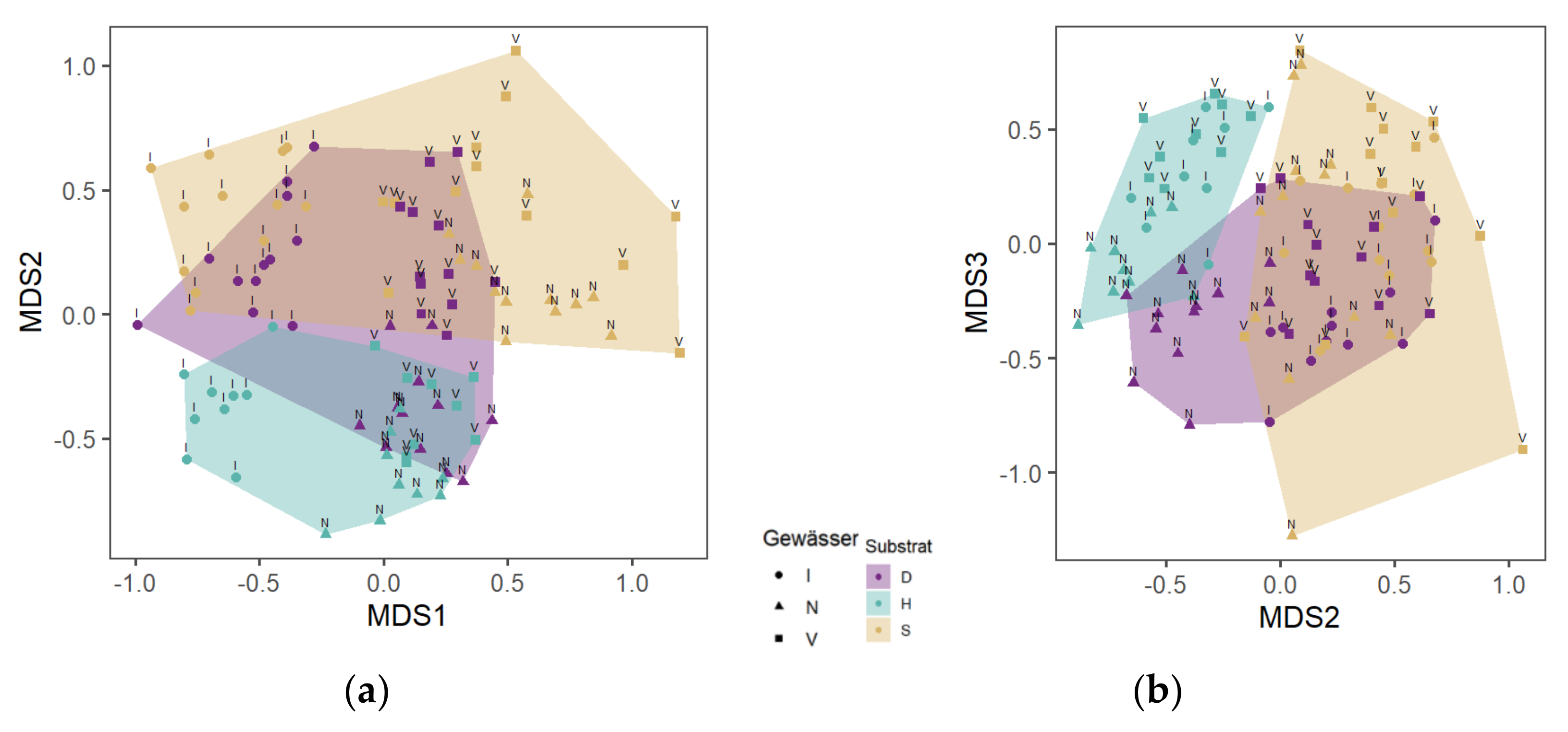
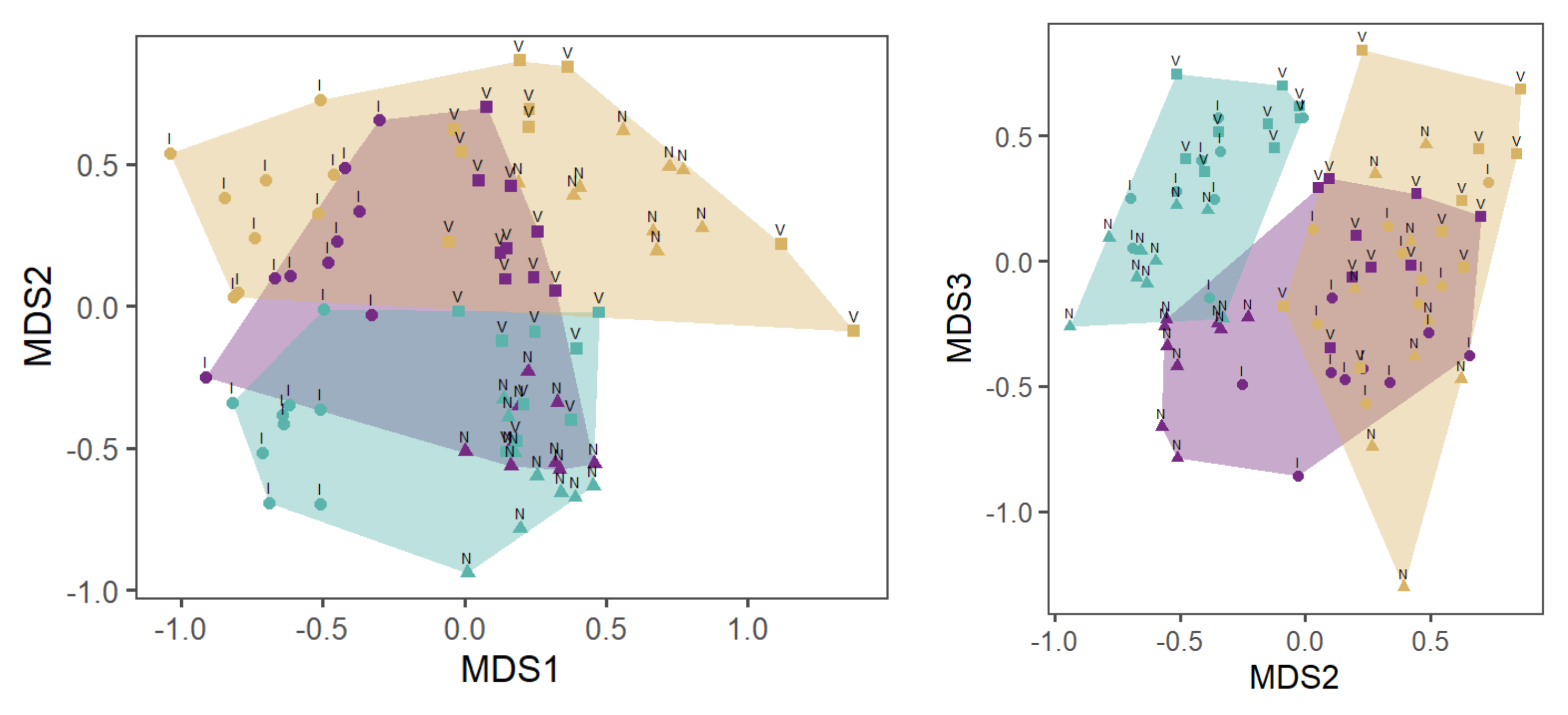
Species composition on wood in the WS and CS compared to that in sand has the highest dissimilarity between all substrates (without G) (Figure 4). The R value of the species composition on wood compared to that in sand determined with ANOSIM was 0.35 (ANOSIM; p < 0.001). Twenty-five taxa on the wood could not be found in the sand. In contrast, there were seven taxa in the sand that were not present on the wood.
The R-value of detritus compared to sand was 0.30, indicating an adequate dissimilarity with some overlap between both substrates (ANOSIM; p < 0.001) Figure 4). Twenty taxa in the detritus were not found in the sand. In contrast, nine taxa that did not populate detritus were collected in the sand.
Detritus and wood were most similar with large overlaps (ANOSIM; R = 0.13 p < 0.01) (see Figure 4). Twenty-one taxa were found in the wood and not in the detritus. On the contrary, 14 species were collected from detritus and not from wood.
3.3.2. Wood Sections
Species composition on wood compared to sand of WS had the highest dissimilarity among all the substrates (without G) (Figure 5). A comparison between wood and sand provided an R value of 0.41 (ANOSIM; p < 0.001). Twenty-nine taxa were found on and in wood that were not found in sand. Conversely, only five taxa were found in sand.
The R value of detritus compared to sand was 0.31, indicating some dissimilarity but with some overlap between both substrates (ANOSIM; p < 0.001). Twenty-five taxa were found in the detritus but were not present in the sand. In contrast, ten taxa that did not populate the detritus were found in the sand.
The R-value of detritus versus wood is nearly identical between the WS and the CS. Species composition in detritus compared to wood was different, but there were large overlaps (ANOSIM; R = 0.13; p < 0.01). Twenty-two taxa were found on wood and not in detritus. Fourteen species did not colonize wood but were collected from the detritus.
3.4. Colonization of MI and Insects in Dependence on Substrates
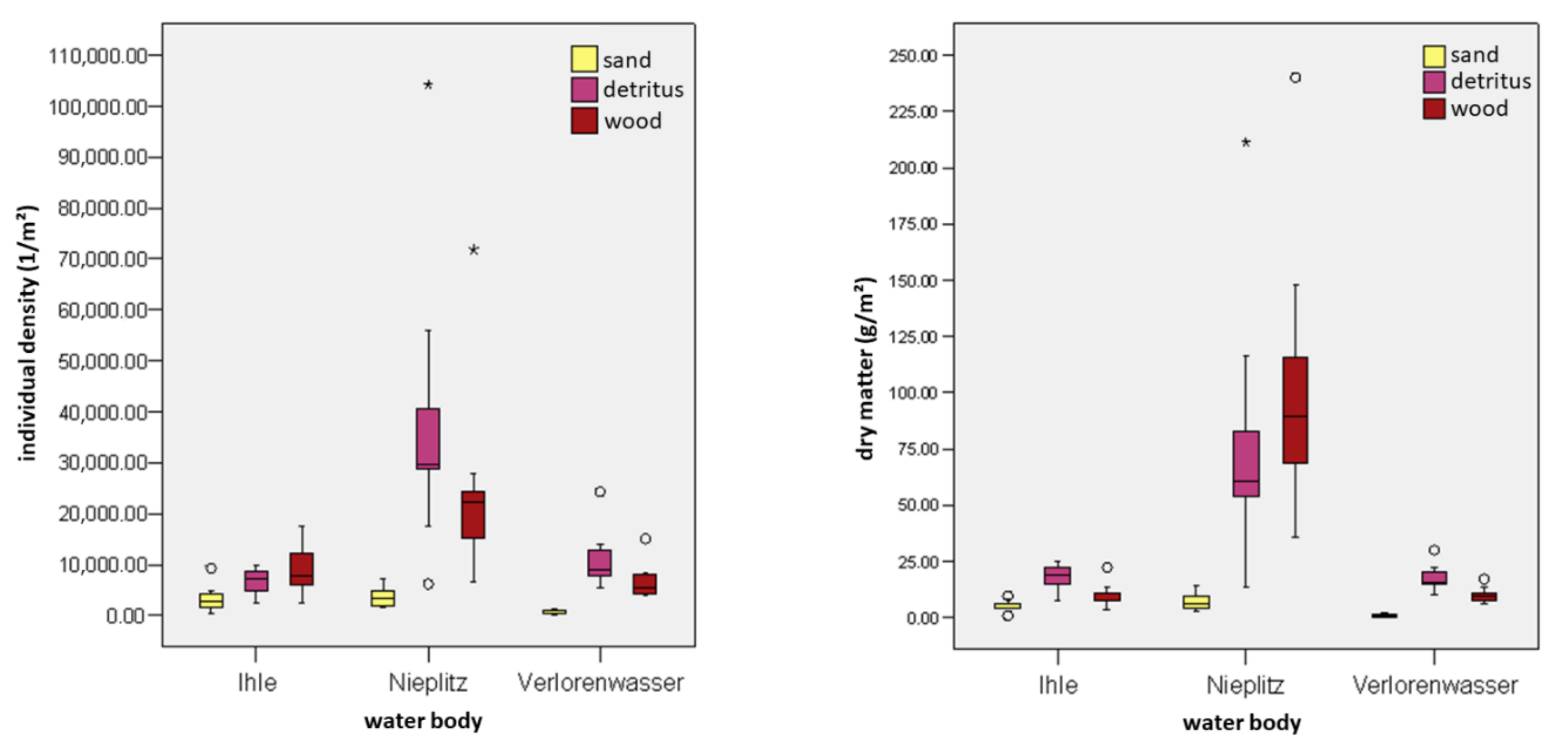
It was checked whether the abundance of MI and insects on wood was higher than in sand. Compared to sand, wood promotes the area-specific settlement of MI in the WS of all water bodies. Figure 6 shows boxplots of individual density and dry matter in the threee streams Ihle, Nieplitz and Verlorenwasser. Figure 7 shows the species richness of substrates (except gravel). The number of individuals of MI in all streams was higher in wood than in sand. In the Ihle, the density of individuals per m2 on the wood was 2.7 times (p < 0.01) than in the sand. For the Nieplitz and the Verlorenwasser, the MI densities were 6.9 times (p < 0.001) and 11.3 times (p < 0.001) higher, respectively, than in the sand. In all streams, the wood contained an average of 13,650 individuals per m2 (min 6741 [ØV]; max 24,942 [ØN]) compared to the sand with an average of 2538 individuals per m2 (min 599 [ØV]; max 3636 [ØN]). Dry matter density of MI that populated wood was twice that for sand in the Ihle (p < 0.01), 14.8 times higher than that in sand for the Nieplitz (p < 0.001), and 10.9 times (p < 0.001) higher than in sand for the Verlorenwasser.
For (semi)aquatic insects, all streams (one exception was the dry matter of the Ihle) showed significantly higher densities on the wood as compared to that of the sand. Additionally, density/m2 of insect species on wood was twice as high than in sand.
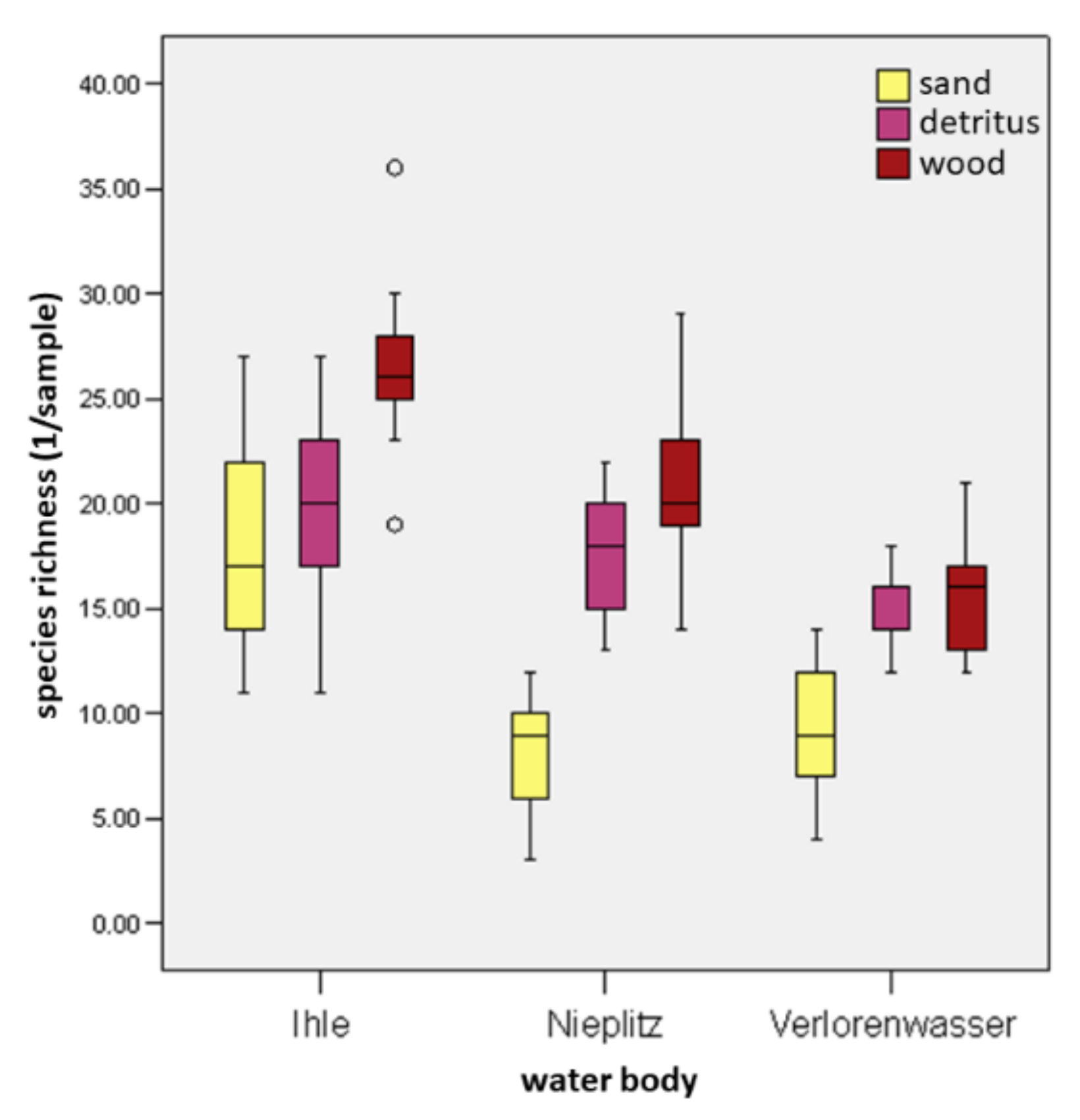
Detritus significantly promoted MI abundance in WS of all streams as compared to sand, with the exception of species abundance in Ihle stream. For this stream, species richness of detritus in wood as compared to sand did not show a significant difference (Figure 6 and Figure 7). In detritus of the WS, the number of individuals were 1.9-fold (p < 0.05) more in Ihle, 10.6-fold (p < 0.001) more in Nieplitz and 18.1-fold (p < 0.001) more in Verlorenwasser than in the sand. For all streams, detritus was colonized by an average of 18,574 individuals per m2 (min 6741 [ØI]; max 24,942 [ØN]), but for sand only by an average of 2538 individuals per m2 (min 599 [ØV]; max 3636 [ØN]).
MI densities in the detritus, as compared to in the sand, were 3.2-fold higher for the Ihle, 11.2-fold higher for the Nieplitz, and 19.4-fold higher (p < 0.001) for the Verlorenwasser. For all streams, dry matter of MI in detritus averaged 38.14 g per m2 (min 17.38 [ØI]; max 78.94 [ØN]) and of sand averaged 4.46 g per m2 (min 0.93 [ØV]; max 7.04 [ØN]). For insects, in all streams (except for individual density and species richness of Ihle) MI densities of detritus were significantly higher, as compared to that of the sand.
3.5. Relationship between the MI Population and the Area Percentage of Substrates
The Spearman rank correlation test was used in order to determine whether MI individual density and dry matter density were significantly affected by the substrate type. Table 3 shows the correlation coefficients (rsp) of the substrates sand, wood and detritus of the WS and the CS (n = 12) as a function of the individual density and the dry matter density.
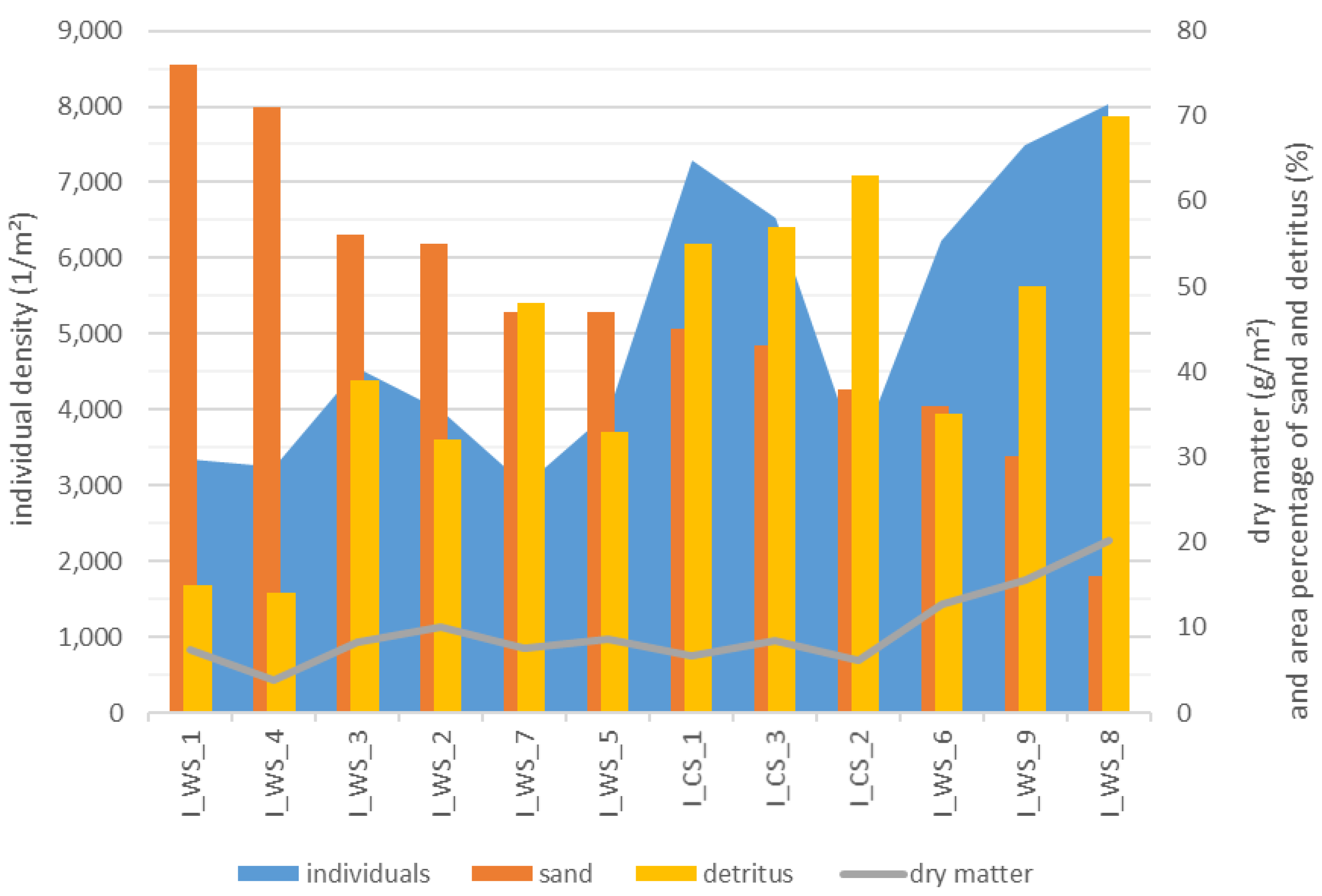
In the Ihle, the higher the fraction of sand, the lower the density of individuals and the dry matter density of MI. In the Nieplitz, the density of MI and dry matter had a strong correlation (with values of 0.73 resp. 0.72) with wood.
In addition, the density of MI individuals in Ihle correlates significantly with the proportion of detritus (Table 3 and Figure 8).
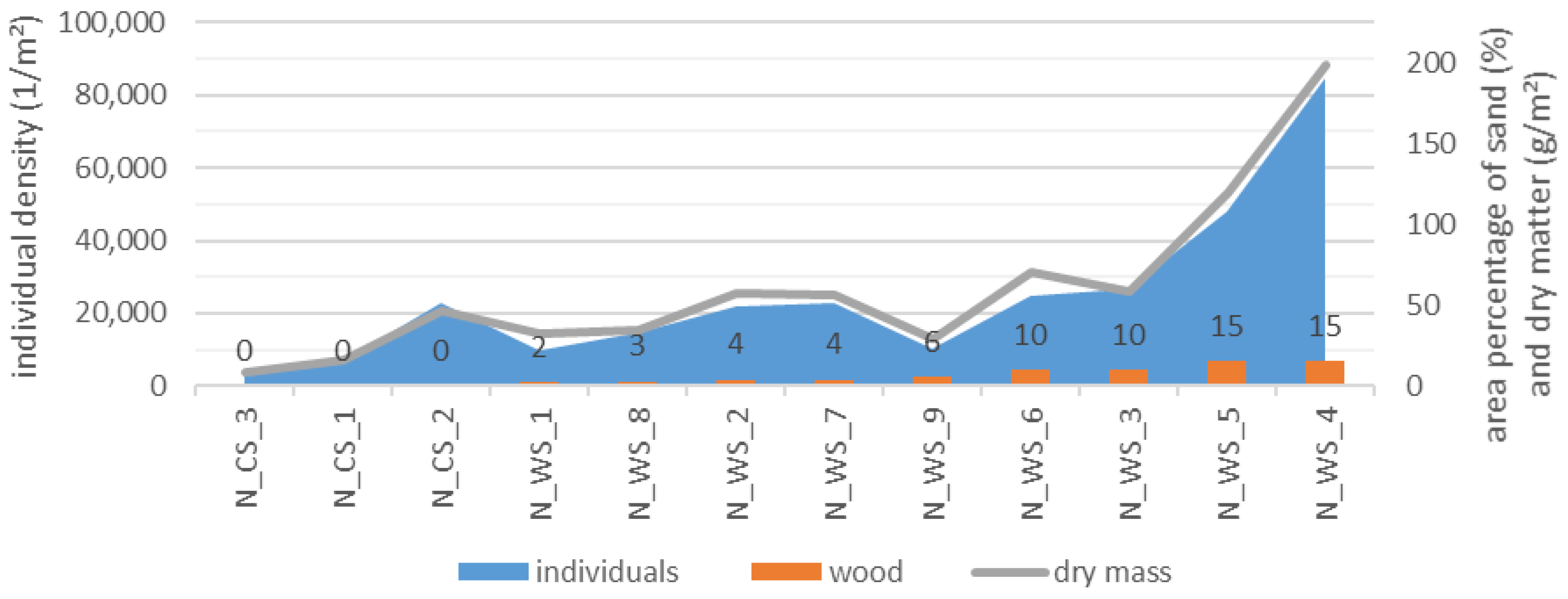
In Nieplitz, individual density and dry matter per m2 increased with increasing proportion of wood (Figure 9). Both relationships show a high correlation. The correlation coefficient for the MI individual density as a function of wood was 0.73 and that of the dry matter 0.72 (p < 0.05).
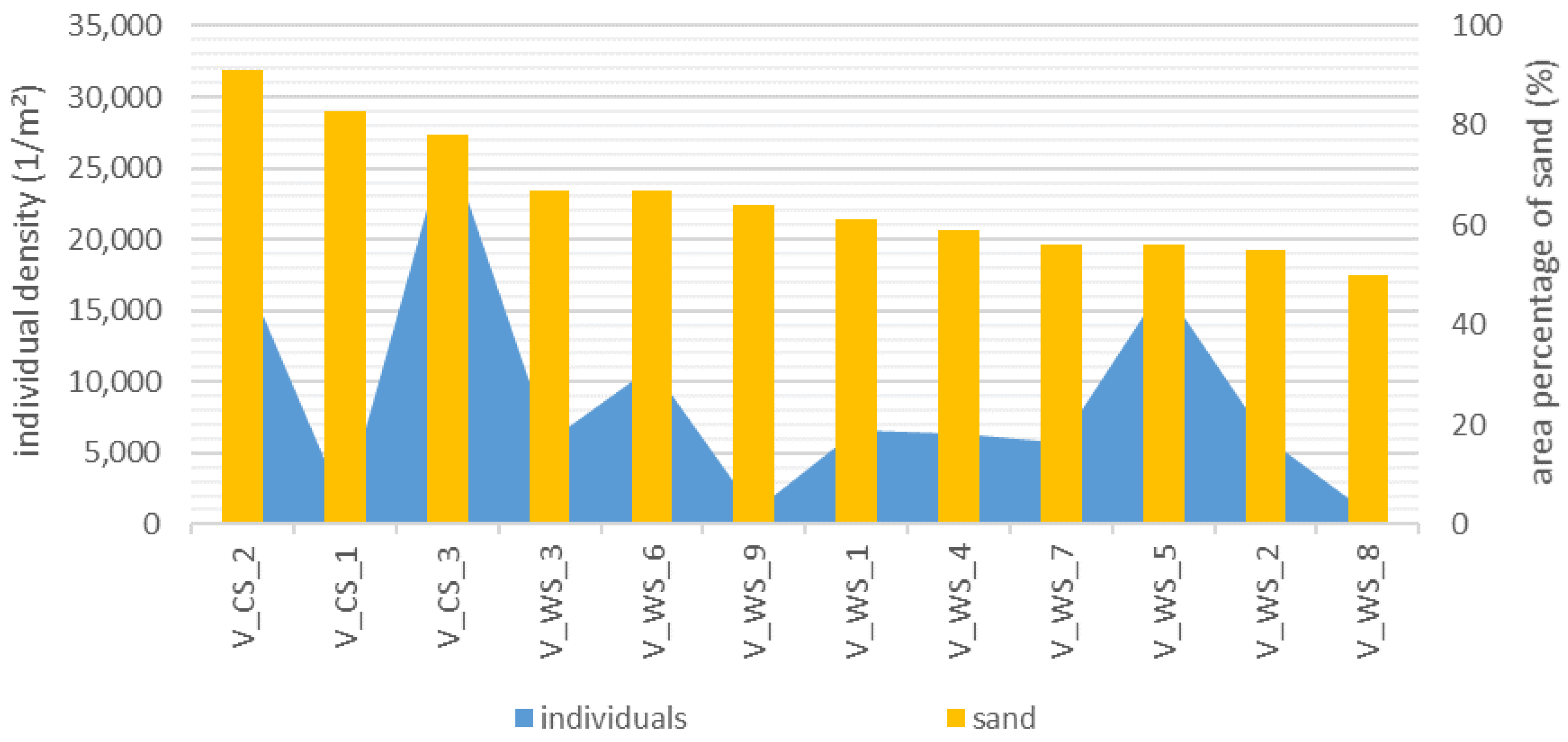
In the Verlorenwasser, the density of individuals per m2 correlated with the proportion of sand with a coefficient of −0.58 (p = 0.05) (Figure 10). Spearman testing of whether the fraction of wood correlated with the proportion of detritus showed no significant correlation (rsp = 0.28; p = 0.16).
3.6. Wood Structure Types
The colonization parameters (individual density, species richness, dry matter) of MI and insects did not differ depending on the type of deadwood (incline bridge, flow guide, parallel work). Furthermore, according to ANOSIM any differences in species composition depending on the three wood structure types were not significant (R < −0.019; p < 0.99).
4. Discussion
4.1. Influence of Deadwood and Restoration on the MI-Fauna
It is not a new research finding that deadwood affects geomorphic processes and functions in rivers and streams. As part of a meta-study evaluating 137 original papers, Polednikova and Galia [21] were able to show that increased channel heterogeneity and the creation or development of specific habitats are by far the most important environmental services provided by naturally or anthropogenically introduced deadwood. Thus, placement of wood and woody debris in streams is one of the most widespread and common techniques to improve riverine fish and MI habitat [22]. Concerning fish, Whiteway et al. [23] found for Canadian streams that woody instream structures lead to localized increases in fish abundance (number and density) for salmonid fishes. Positive effects on fish by deadwood were statistically verified in the framework of a meta-study by Roni et al. [22]. Of the 122 studies that they reviewed evaluating wood placement, 81 concerned fish and, among them, 68 reported a positive response in fish abundance, biomass, species diversity or richness, or survival. However, studies concerning aquatic MI are less frequent and mainly concern mountainous streams [24]. Moreover, up to the present, very little is known about the influence of deadwood in lowland sand-bed streams [25], although some of this research gap has been filled by some recent research on the success of lowland stream and river restoration projects [26,27].
One of the streams investigated in this study (Ihle) was subjected to extensive restoration measures in 2002 and 2003 [28] and contain large amounts of gravel substrate in the river bottom. Due to anthropogenic and natural input of deadwood and coarse detritus the diversity of substrate and current is very high. Under such conditions, the Ihle has shown the highest species inventory among the sampled streams, and MI-based ecological assessment indicates a good ecological status with tendency to very good [26]. High diversity of substrate and flow velocity create valuable habitats—mixtures of gravel and deadwood—for sensitive species from the groups of Trichoptera and Plecoptera. These conditions are found in the Ihle but are less evident in the other two streams of this study. Thus, in addition to the Ihle showing the highest species inventory of MI, insects and EPT, most of the species that were rare in the other two water bodies were found in the Ihle (see Supplementary Materials). Meanwhile, the species composition of Verlorenwasser and Nieplitz shows a high similarity. The similarity of Ihle to these two other streams studied is rather lower due to the geographical distance and the effects of restoration measures. On the other hand, hydraulic engineering structures could impair biodiversity of the water bodies, particularly in the headwaters of Verlorenwasser, since lacking continuity can inhibit the potential for recolonization and species exchange. Although the headwaters of the Verlorewasser do not contain any hydraulic structures, the sections below do. Additionally, the Verlorenwasser is impaired in the lower reaches by several hydraulic structures with nine falls [29]. This circumstance can be an additional reason why the fewest MI, insects (38) and EPT (23) were detected in Verlorenwasser.
4.2. Effect of Substrates on MI Populations
Formation of the waterbed depends on hydromorphological factors such as depth and width variance, gradient and roughness. Depending on the flow velocities, sediments are washed out or deposited. As a result of flow, oxygen is introduced into the water according to the turbulence and the fauna reacts to all these factors by the occurrence or absence of species with high or low demands for current, substrate type or oxygen saturation [30].
In sandy lowland rivers where hydromorphological structures are rare, deadwood is a hotspot for many species of macroinvertebrates [31,32,33,34,35,36]. Benke et al. [31] recorded 59% of all species only on wood in the sandy lowland stream Satilla River in Georgia. Wood occupied a maximum of only 6% on the riverbed. Sand, on the other hand, dominated with an area of 85% and had only 36% of the taxa. The results of our study show a tendency similar to Benke et al. [31]. In the examined WS in our streams, wood occupied an area of 7% but 71% of the species (total from n = 9, i.e., approx. 0.56 m2) were recorded there. The sand of WS covered an average of 57% and was colonized by only 47% of the species (n = 9). On average, 36% of all species were found exclusively on wood and 11% only in sand. A study by Speth et al. [32] also shows that more species were found in wood of a sandy stream in the north German lowlands (Osterau, Schleswig-Holstein) than in sand. However, this previous study did not differentiate MI with regard to the substrate type or have the taxonomic resolution of the present study.
We found that MI colonized wood in all in all the sampled streams with an average of 21 taxa, more than twice than in sand. However, absolute individual and taxa numbers as well as dry matter of the macroinvertebrates should be differentiated, since there are species that are very tolerant to stressors, such as a high number of white flies (Chironomidae Gen. sp.) and black flies (Simulium sp.). Thus, special consideration was given among MI to insects and, more specifically, to sensitive taxa such as EPT species. We were able to show that not only species richness of MI was promoted, but also that of insects and sensitive species. Species richness (number of taxa) of MI, insects and EPT were approximately twice as high on the wood than in the sand.
For example, the riffle beetles Elmis sp. as a sensitive species only were found on wood in all three bodies of water, but never on the sand. Likewise, the alderflies Sialis fuliginosa (only found in Ihle) and Sialis lutaria (only found in Nieplitz and Verlorenwasser) were only found on wood and not in sand. Alderflies (Sialis spp.) prefer shaded stream banks with structures hanging over the stream, such as bridge deadwood structures, to lay their eggs near the water surface. After hatching, the larvae fall into the water and begin the aquatic stadium [32,33]. Caddisflies (Trichoptera) such as Lype reducta and L. phaeopa, in contrast, choose the soft tissue of submerged wood to lay their eggs.
The wood also offers a large food supply for xylophagous species such as L. phaeopa, L. reducta or Lepidostoma basale [33]. For example, L. phaeopa was only found on the wood and not in the sand of Nieplitz and Verlorenwasser. In addition, some species, including Diptera such as Tipulidae, burrow into the surface of the wood to protect their pupae more effectively against runoff or predation [32]. However, the wood also provides food for non-xylophagous taxa. Grazers and sediment eaters, which have also been detected on the wood substrates of all the streams, feed on the biofilm on the wood. This film consists of fungi, bacteria and algae. In all streams, wood caused a significantly higher dry matter density of MI than sand on the wood sections, which can be explained by the higher food supply.
Apparently, however, some of the mayflies (Ephemeroptera), stoneflies (Plecoptera) or caddisflies (Trichoptera) species are especially adapted to life on the current-exposed sandy areas of the sandy lowland streams. Sand offers, for example, to sand quiver-building caddisflies such as Sericostoma personatum and Notidobia ciliaris, building material or to the stonefly Isoptena serricornis larvae a habitat to bury. For example, the species Isoptena serricornis [35], which is characteristic in sandy lowland streams, only occurred in the sand of the Verlorenwasser. The larvae of these stoneflies are typical residents of near-natural, largely unaffected water bodies, and prefer sand as a colonization substrate [35]. However, they are only found sporadically in the sand due to water quality stressors [26,35]. In addition, more individuals of the mayfly Ephemera danica were collected in sand of all water bodies as compared to on wood. E. danica digs U-shaped burrows in the sandy substrate and feeds by actively filtering mineral substrate and detritus [36]. The higher dry matter per individual on wood than in sand suggests a higher food supply despite the lower number of individuals.
Ho et al. [24] summarize the mechanisms through which wood structures diversify MI-communities. These are food resources retained by wood, accidental migration from wood surfaces, variation in flow velocities, diversification of stream bed substrates and reduced drift loss during high-flow events. In our former study concerning the Ihle [26], we were able to show that these mechanisms were enhanced in the process of natural succession by structural diversification of deadwood structures themselves.
Effects of deadwood can also be enhanced by interaction with other organic substrates [4,12]. Detritus is formed from organic material such as fallen leaves. Organic material such as rootstocks or leaves can be retained to a greater extent by wooden structures [25].
Putri et al. [37] found that with a higher proportion of organic substrate, abundance of MI is promoted. This was confirmed in our study. We showed that the presence of wood leads to significant differences in colonization of detritus compared to sand in the wood sections of all streams. Individual density and dry matter of MI, insects (with the exception of the individual density of Ihle) and EPT of wood sections were higher in detritus than in sand. In control sections, there were no significant differences between colonization of detritus compared to sand. Therefore, for individuals in sand, it does not seem to make any difference whether wood was present nearby, probably due to their adaptation to current velocities.
In addition, 20 taxa that were not found in sand (incongruent species) were present in the detritus of all streams (see Supplementary Materials). These included, for example, the caddisfly species Lepidostoma basale (only found in Ihle), that is typical for sandy lowland streams. For the occurrence of L. basale, deadwood accumulations and detritus input as food sources are important prerequisites [33].
4.3. Substrates in WS and CS
It was not possible to determine clearly whether the sections with wood were more conducive to MI populations in the detritus than in the wood-free sections. This hypothesis was confirmed for only one stream. Wood in WS in Verlorenwasser had a positive effect on individual densities of MI and insects in the subsequent substrate detritus (twice to three times more individuals). WS of Verlorenwasser also promoted taxa richness (twice as high) and the dry matter density of the insects (twice as much) of the detritus compared to the CS. Detritus from WS of Verlorenwasser tended to contain species that are more characteristic for the water body type and/or the presence of ecologically valuable structures (such as Elmis sp.) than the detritus from the CS. It was remarkable, however, that 40% fewer individuals of MI and insects were found in the detritus of WS in the Ihle, but no significant results were found with regard to dry matter density and species richness of EPT. In Nieplitz, no statistically significant effect was found for MI of detritus from WS as compared to CS, possibly due to the diversity of interacting influences on MI. Nevertheless, the detritus of Nieplitz WS was more conducive to MI populations, insects and EPT (although not significantly so).
Deadwood had no significant influence on the populations of all other substrates. For the sand individuals, there was no significant difference between WS and CS, possibly due to adjustment to flow velocities. However, significantly more individuals of the species Halesus sp. and Sericostoma personatum live in WS because on unstable sandy areas, caddisfly species can only move with heavy quivers [32].
4.4. Comparison of Stream Sections in WS and CS
Wood is an important habitat for benthic MI in running waters. The studies by Noack et al. [38] and Ho et al. [24] demonstrated that the occurrence of deadwood creates an additional mesohabitat. A higher diversity of meso- and microhabitats leads to the colonization by a type-specific MI community with specific needs and stages of development.
Pilotto et al. [34] demonstrated that hydromorphological site variables were higher on sections with wood than on wood-poor sites (speed: 85% higher, turbulence: 89%, grain size: 126%, sorting index: 67% higher). Due to this varying flow behavior in a section of water with wood, fast and slow-flowing areas are formed. The substrate formation can be predicted on the basis of the deadwood structure types. Gravel is often washed out in the fast-flowing areas, while detritus (organic material) is deposited in the slower-moving areas. In Verlorenwasser and Nieplitz, the proportion of detritus in the WS was therefore higher than in the CS. However, significant differences in MI populations of the respective sections of water were only shown for the Verlorenwasser.
In this study, we also observed whether there was a difference in MI populations as related to different surface areas of wood, detritus or sand, as MI habitat depends on the proportion of certain substrates. Accordingly, MI populations in Nieplitz were promoted by wood and in the Ihle by detritus, and tended to be inhibited by sand in the Verlorenwasser and the Ihle. However, due to the higher area percentage of detritus and the lower percentage of sand in Ihle CS, the maximum values of individual density and dry matter density was higher than in the WS.
4.5. Wood Texture Types
The null hypothesis which states that there are no clear differences in colonization by MI, insects and sensitive taxa such as EPT depending on the wood structure type, has been confirmed [39]. The MI populations (density of individuals, density of dry matter and species richness) were not different between incline bridge, flow guide and parallel work wood types. All three structure types contributed to the formation of a rich substrate distribution and served as a microhabitat regardless of location. So far, there are no comparative results in the literature, since this issue has not yet been investigated.
5. Conclusions and Recommendations for Improvement Measures
From our study, measures can be derived which can lead to an improvement in the framework conditions for MI in the sandy watercourses of the North German lowlands.
Priority considerations and actions should be with:
- Make surrounding land use more MI friendly: Riparian strips should be identified and secured so that deadwood can be brought in a natural way. This input can promote detritus formation and MI colonization, particularly in sections that have little or no deadwood. Care should be taken to ensure that the formation of colonization-promoting substrates such as detritus and wood is favored but sand is not completely covered.
- Create diverse water body structures: Settlement by MI is dependent on the available surface area of wood and less on volume. Thus, when present in sufficient quantities, branches (0.05 to 0.1 m in diameter) and coarse organic material, such as large logs, have the potential as additional microhabitats.
- Considering the surface of deadwood: More individuals would be found under the bark of older deadwood because they are protected from drifting. When putting in deadwood, care should be taken to ensure that deadwood with bark is among.
- Use of deadwood also in Heavily Modified Water Bodies: The effect of introduced deadwood in unnatural streams compared to near-natural streams with natural deadwood is similar, the species richness did not differ significantly from each other.
- Further development of monitoring: In order to identify problems and to check effectiveness, monitoring and further research are of great importance for the promotion of colonization by MI, insects and sensitive species by means of deadwood.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/ecologies4010008/s1, Table S1: Taxa list and numbers of individuals per body of water.
Author Contributions
Conceptualization, J.M.Z.; methodology, J.M.Z. and V.L.; software, J.M.Z.; validation, J.M.Z., V.L. and R.M.G.; formal analysis, J.M.Z.; investigation, J.M.Z. and V.L.; resources, V.L.; data curation, J.M.Z.; writing—original draft preparation, V.L. and J.M.Z.; writing—review and editing, V.L., R.M.G. and J.M.Z.; visualization, J.M.Z.; supervision, R.M.G.; project administration, V.L.; funding acquisition, J.M.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
The animal study protocol was approved by the Institutional Review Board of the Institute for Water Management and Ecotechnology by date of 1st of September 2022 (Protocol Code: 09-01-2022-001).
Data Availability Statement
Data are available in the Supplementary Materials.
Acknowledgments
We thank Michael Seidel for his conceptual work and the support with the investigations in the field. Katja Westphal deserves our appreciation for the valuable help with the statistical investigations. Special thanks also to the editor and two anonymous reviewers for their constructive comments on the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hallmann, C.A.; Sorg, M.; Jongejans, E.; Siepel, H.; Hofland, N.; Schwan, H.; Stenmans, W.; Müller, A.; Sumser, H.; Hörren, T.; et al. More than 75 percent decline over 27 years in total flying insect biomass in protected areas. PLoS ONE 2017, 12, e0185809. [Google Scholar] [CrossRef] [PubMed]
- Hornberg, C.; Niekisch, M.; Calliess, C.; Kemfert, C.; Lucht, W.; Messari-Becker, L.; Rotter, V.S. For a Surface-Effective Insect Protection. Opinion; Scientific Advisory Board for Biodiversity and Genetic Resources at the Federal Ministry of Food and Agriculture, Ed.; German Advisory Council on the Environment (SRU): Berlin, Germany, 2016; Available online: https://www.umweltrat.de/SharedDocs/Downloads/DE/04_Stellungnamen/2016_2020/2018_10_AS_Insektenschutz.pdf?__blob=publicationFile&v=19 (accessed on 4 August 2020).
- Liess, M.; Liebmann, L.; Vormeier, P.; Weisner, O.; Altenburger, R.; Borchardt, D.; Brack, W.; Chatzinotas, A.; Escher, B.; Foit, K.; et al. Pesticides are the dominant stressors for vulnerable insects in lowland streams. Water Res. 2021, 201, 117262. [Google Scholar] [CrossRef] [PubMed]
- Seidel, M. Naturnaher Einsatz von Holz zur Entwicklung von Fließgewässern im Norddeutschen Tiefland. Schriftenreihe des Instituts für Wasserwirtschaft und Ökotechnologie; Ettmer, B., Lüderitz, V., Eds.; Shaker Verlag GmbH: Aachen, Germany, 2018. [Google Scholar] [CrossRef]
- UBA. Umweltbundesamt (2019): Gewässertyp des Jahres 2018—Sandiger Tieflandbach. 2019. Available online: https://www.umweltbundesamt.de/themen/wasser/gewaessertyp-des-jahres/gewaessertyp-des-jahres-2018-sandiger-tieflandbach#kurzbeschreibung (accessed on 10 November 2020).
- Röthele, I.; Mertens, M.; Küry, D. Erhebung Biomasse Makrozoobenthos 2014 Violenbach, Orisbach, Ergolz, Frenke, Hintere Frenke, Vordere Frenke. 2014. Available online: https://www.baselland.ch/politik-und-behorden/direktionen/volkswirtschafts-und-gesundheitsdirektion/jagd-und-fischerei/fischereiwesen/fischfauna-untersuchungsberichte/downloads/erhebung-biomasse-makrozoobenthos_2014.pdf/@@download/file/erhebung-biomasse-makrozoobenthos_2014.pdf (accessed on 10 May 2020).
- Neuhaus, V.; Mende, M. Engineered Large Wood Structures in Stream Restoration Projects in Switzerland: Practice-Based Experiences. Water 2021, 13, 2520. [Google Scholar] [CrossRef]
- Google. n.d. Locations of Ihle, Nieplitz and Verlorenwasser. Data SIO, NOAA, U.S. Navy, NGA, GEBCA. Image Landsat/Copernicus. Available online: https://earth.google.com/web/search/ihle/@49.42246021,8.12620422,-57.14636488a,1828973.53736222d,35y,-0.32186261h,23.21376508t,0r/data=CigiJgokCZXaZMslE0VAEYEcRHJFEEVAGQC0GiHTVQRAIauPEd9hMQRA (accessed on 26 December 2022).
- UBA. Umweltbundesamt. Fließgewässertyp des Jahres. GIS-Karte. 2016. Available online: https://gis.uba.de/maps/resources/apps/gdj/index.html?lang=de (accessed on 24 November 2020).
- Pottgiesser, T.; Sommerhäuser, M. Aktualisierung der Steckbriefe der Bundesdeutschen Fließgewässertypen (Teil A) und Ergänzung der Steckbriefe der Deutschen Fließgewässertypen um Typspezifische Referenzbedingungen und Bewertungsverfahren aller Qualitätselemente (Teil B). Essen. 2008. Available online: http://gewaesser-bewertung.de/files/steckbriefe_fliessgewaessertypen_april2008.pdf (accessed on 30 April 2020).
- Gerhard, M.; Reich, M. Totholz in Fließgewässern: Empfehlungen zur Gewässerentwicklung; Gemeinnützige Fortbildungsgesellschaft für Wasserwirtschaft und Landschaftsentwicklung (GFG) mbH: Mainz, Germany; WBW Fortbildungsgesellschaft für Gewässerentwicklung mbH: Karlsruhe, Germany, 2001; pp. 12–19. Available online: https://wbw-fortbildung.de/sites/default/files/2021-10/Totholz%20in%20Flie%C3%9Fgew%C3%A4ssern%20-%20Empfehlung%20zur%20Gew%C3%A4sserentwicklung.pdf (accessed on 4 August 2020).
- Seidel, M.; Brunke, M. Impulsgebende Maßnahmen in Tieflandbächen Schleswig-Holsteins—Wirkung und mögliche Einbauvarianten für eine naturnähere Gestaltung. Wasser Wirtsch. 2015, 12, 55–60. [Google Scholar] [CrossRef]
- Bellmann, H. Der Kosmos Libellenführer: Die Arten Mitteleuropas Sicher Bestimmen; Franckh-Kosmos Verlags-GmbH &Co. KG: Stuttgart, Germany, 2007. [Google Scholar]
- Eiseler, B. Bildbestimmungshilfen für die Eintagsfliegenlarven der deutschen Mittelgebirge und des Tieflandes. In Lauterbornia. Internationale Zeitschrift für Faunistik und Floristik der Binnengewässer Europas; Heft 53; Mauch, E., Ed.; Kessler Verlagsdruckerei: Dinkelscherben, Germany, 2005. [Google Scholar]
- Eiseler, B. Bestimmungshilfen—Makrozoobenthos (1). Arbeitsblatt 14. In Landesamt für Natur, Umwelt und Verbraucherschutz Nordrhein-Westfalen (Ed.): Taxonomie für die Praxis; LANUV: Recklinghausen, Germany, 2010. [Google Scholar]
- Eiseler, B.; Hess, M. Bestimmungshilfen—Makrozoobenthos (2). Arbeitsblatt 20. In Landesamt für Natur, Umwelt und Verbraucherschutz Nordrhein-Westfalen (Ed.): Taxonomie für die Praxis; LANUV: Recklinghausen, Germany, 2013. [Google Scholar]
- Glöer, P. Süßwassermollusken. Ein Bestimmungsschlüssel für die Bundesrepublik Deutschland (14. Auflage); Deutscher Jugendbund für Naturbeobachtung, Ed.; SENSER-DRUCK: Augsburg, Germany, 2015. [Google Scholar]
- Waringer, J.; Graf, W. Atlas der Österreichischen Köcherfliegenlarven unter Einschluß der Angrenzenden Gebiete; Facultas Universitätsverlag: Vienna, Austria, 1997. [Google Scholar]
- Jiang, X.; Song, Z.; Xiong, J.; Xie, Z. Can excluding non-insect taxa from stream macroinvertebrate surveys enhance the sensitivity of taxonomic distinctness indices to human disturbance? Ecol. Indic. 2014, 41, 175–182. [Google Scholar] [CrossRef]
- Chapman, M.G.; Underwood, A.J. Ecological patterns in multivariate assemblages: Information and interpretation of negative values in ANOSIM tests. Mar. Ecol. Prog. Ser. 1999, 180, 257–265. [Google Scholar]
- Poledniková, Z.; Galia, T. Ecosystem Services of Large Wood: Mapping the Research Gap. Water 2021, 13, 2594. [Google Scholar] [CrossRef]
- Roni, P.; Beechie, T.; Pess, G.; Hanson, K. Wood placement in river restoration: Fact, fiction, and future direction. Can. J. Fish. Aquat. Sci. 2015, 72, 466–478. [Google Scholar] [CrossRef]
- Whiteway, S.L.; Biron, P.M.; Zimmermann, A.; Venter, O.; Grant, J.W. Do in-stream restoration structures enhance salmonid abundance? A meta-analysis. Can. J. Fish. Aquat. Sci. 2010, 67, 831–841. [Google Scholar] [CrossRef]
- Lo, H.W.; Smith, M.; Klaar, M.; Woulds, C. Potential secondary effects of in-stream wood structures installed for natural flood management: A conceptual model. WIREs Water 2021, 8, e1546. [Google Scholar] [CrossRef]
- Mutz, M. Influences of Woody Debris on Flow Patterns and Channel Morphology in a Low Energy, Sand-Bed Stream Reach. Int. Rev. Hydrobiol. 2000, 1, 107–121. [Google Scholar] [CrossRef]
- Seidel, M.; Li, F.; Langheinrich, U.; Gersberg, R.M.; Lüderitz, V. Self-dynamics as a driver for restoration success in a lowland stream reach. Limnologica 2021, 88, 125873. [Google Scholar] [CrossRef]
- Seidel, M.; Li, F.; Winking, C.; Sommerhäuser, M.; Lüderitz, V. Should We Sample More than Required by the European Water Framework Directive? Case study: Emscher Catchment. CLEAN—Soil Air Water 2022, 50, 2000391. [Google Scholar] [CrossRef]
- Lüderitz, V.; Jüpner, R.; Müller, S.; Feld, C.K. Renaturalization of streams and rivers—The special importance of integrated ecological methods in measurement of success. An example from Saxony-Anhalt (Germany). Limnologica 2004, 34, 249–263. [Google Scholar] [CrossRef]
- Unselt, C.; Haack, S.; Dreger, F.; Gebauer, D.; Brunk, I. Pflege- und Entwicklungsplan Naturpark Hoher Fläming. Landesamt Brandenburg—Landesanstalt für Großschutzgebiete (Ed.). Available online: https://www.hoher-flaeming-naturpark.de/fileadmin/user_upload/PDF/Hoher-Flaeming/PEP_Hauptstudie_Naturpark_Hoher_Flaeming_2006_bearbeitet.pdf (accessed on 25 November 2020).
- Dahm, V.; Kupilas, B.; Rolauffs, P.; Hering, D. Strategien zur Optimierung von Fließgewässer-Renaturierungsmaßnahmen und ihrer Erfolgskontrolle. Umweltbundesamt (Ed.). Texte 43/2014. Dessau-Roßlau. Available online: https://www.umweltbundesamt.de/sites/default/files/medien/378/publikationen/texte_43_2014_strategien_zur_optimierung_von_fliessgewaesser-renaturierung_0.pdf (accessed on 24 October 2020).
- Benke, A.C.; Van Arsdall, T.C.; Gillespie, D.M.; Parrish, F.K. Invertebrate Productivity in a Subtropical Blackwater River: The Importance of Habitat and Life History. Ecol. Monogr. 1984, 54, 25–63. [Google Scholar] [CrossRef]
- Speth, S.; Böttger, K. Die sustratspezifische Verteilung der Ephemeroptera, Plecoptera und Trichoptera (Insecta) in einem sandigen Bach des norddeutschen Tieflnads (Osterau, Schleswig-Holstein). Limnologica 1993, 23, 369–380. [Google Scholar]
- Hoffmann, A.; Hering, D. Wood-Associated Macroinvertebrate Fauna in Central European Streams. Int. Rev. Hydrobiol. 2000, 85, 25–48. [Google Scholar] [CrossRef]
- Pilotto, F.; Harvey, G.L.; Wharton, G.; Pusch, M.T. Simple large wood structures promote hydromorphological heterogeneity and benthic macroinvertebrate diversity in low-gradient rivers. Aquat. Sci. 2016, 78, 755–766. [Google Scholar] [CrossRef]
- Boiché, A.; Mondy, C.; Jacquemin, G.; Thiebaut, G. Découverte d’Isoptena serricornis (Pictet, 1841) en France [Plecoptera, Chloroperlidae]/First record of Isoptena serricornis (Pictet, 1841) in France [Plecoptera, Chloroperlidae]. Ephemera 2009, 10, 1–6. [Google Scholar]
- Brandt, J. Ephemera danica im Alstereinzugsgebiet. Institut für Hygiene und Umwelt—Hamburger Landesinstitut für Lebensmittelsicherheit Gesundheitsschutz und Umweltuntersuchungen. 2011. Available online: https://www.hamburg.de/contentblob/2996626/cd3f7d6818384ae51eca59f1170a7873/data/ephemera-danica.pdf (accessed on 2 November 2020).
- Putri, A.M.S.; Suryanti, S.; Widyorini, N. Hubungan Tekstur Sedimen Dengan Kandungan Bahan Organik Dan Kelimpahan Makrozoobenthos Di Muara Sungai Banjir Kanal Timur Semarang The Relation of Sediment Texture to Organic Matter and Macrozoobenthos Abundance in the Estuarine of Banjir Kanal Timur River. Saintek Perikan. Indones. J. Fish. Sci. Technol. 2017, 12, 75–80. [Google Scholar] [CrossRef]
- Noack, M.; Podraza, P.; Lechner, A. Untersuchungen von Mikrohabitaten und deren Bedeutung für das Makrozoobenthos in renaturierten Gewässerabschnitten von Sorpe und Röhr. In Korrespondenz Wasserwirtschaft; Osnabrück University: Osnabrück, Austria, 2017. [Google Scholar]
- Lester, R.E.; Wright, W.; Jones-Lennon, M.; Rayment, P. Large versus small wood in streams: The effect of wood dimension on macroinvertebrate communities. Fundam. Appl. Limnol. 2009, 174, 339–351. [Google Scholar] [CrossRef]
Figure 1.
Location of sampled streams in Germany, published from google earth [8].
Figure 1.
Location of sampled streams in Germany, published from google earth [8].

Figure 2.
Schematic sketch and photos of three frequent deadwood structures [11,12], modified, adapted with permission from photographer, Michael Seidel.

Figure 3.
Design of the experiment for a total of three sampled streams. Different wood structure types and the control sections were sampled in three replicates. Different substrates detritus, sand, gravel and wood were each taken as a mixed sample from five individual samples [12], modified.
Figure 3.
Design of the experiment for a total of three sampled streams. Different wood structure types and the control sections were sampled in three replicates. Different substrates detritus, sand, gravel and wood were each taken as a mixed sample from five individual samples [12], modified.

Figure 4.
Non-metric multidimensional scaling (NMDS) of MI samples from the WS (a) and the CS (b) of streams I—Ihle, N—Nieplitz and V—Verlorenwasser and the substrates D—detritus, H—wood and S—sand. Stress—0.17 (©Katja Westphal).
Figure 4.
Non-metric multidimensional scaling (NMDS) of MI samples from the WS (a) and the CS (b) of streams I—Ihle, N—Nieplitz and V—Verlorenwasser and the substrates D—detritus, H—wood and S—sand. Stress—0.17 (©Katja Westphal).

Figure 5.
Nonmetric multidimensional scaling (NMDS) of MI samples from WS with I = Ihle, N = Nieplitz and V = Verlorenwasser as well as D = detritus, H = wood and S = sand. stress = 0.16 (©Katja Westphal).
Figure 5.
Nonmetric multidimensional scaling (NMDS) of MI samples from WS with I = Ihle, N = Nieplitz and V = Verlorenwasser as well as D = detritus, H = wood and S = sand. stress = 0.16 (©Katja Westphal).

Figure 6.
Density of individuals per m2 (left) and dry matter in g per m2 (right) of MI for the substrates sand, detritus and wood of WS, divided among the water bodies Ihle, Nieplitz, and Verlorenwasser. The box plots show the minimum (lower antenna), the two quartiles, the median, and the maximum (upper antenna) of the distributions. The circles marks are suspected to be outliers, and the star marks the extreme values of the distributions.
Figure 6.
Density of individuals per m2 (left) and dry matter in g per m2 (right) of MI for the substrates sand, detritus and wood of WS, divided among the water bodies Ihle, Nieplitz, and Verlorenwasser. The box plots show the minimum (lower antenna), the two quartiles, the median, and the maximum (upper antenna) of the distributions. The circles marks are suspected to be outliers, and the star marks the extreme values of the distributions.

Figure 7.
Species richness of the macroinvertebrates per sample for the substrates sand, detritus and wood of the WS, related to the streams Ihle (except gravel), Nieplitz, and Verlorenwasser. The box plots show the minimum (lower antenna), the two quartiles, the median, and the maximum (upper antenna) of the distributions. The circles marks are suspected to be outliers.
Figure 7.
Species richness of the macroinvertebrates per sample for the substrates sand, detritus and wood of the WS, related to the streams Ihle (except gravel), Nieplitz, and Verlorenwasser. The box plots show the minimum (lower antenna), the two quartiles, the median, and the maximum (upper antenna) of the distributions. The circles marks are suspected to be outliers.

Figure 8.
Individual density and dry matter in g per m2 depend on the area percentage of sand and detritus (%) of WS and CS in Ihle (I) stream sorted according to the proportion of sand.
Figure 8.
Individual density and dry matter in g per m2 depend on the area percentage of sand and detritus (%) of WS and CS in Ihle (I) stream sorted according to the proportion of sand.

Figure 9.
Individual density per sample and dry matter in g per m2 as a function of the area percentage of wood (%) in WS and CS in Nieplitz (N) sorted according to the proportion of detritus.
Figure 9.
Individual density per sample and dry matter in g per m2 as a function of the area percentage of wood (%) in WS and CS in Nieplitz (N) sorted according to the proportion of detritus.

Figure 10.
Individual density per m2 as a function of the area percentage of sand (%) in WS and CS of Verlorenwasser (V) sorted according to the proportion of sand.
Figure 10.
Individual density per m2 as a function of the area percentage of sand (%) in WS and CS of Verlorenwasser (V) sorted according to the proportion of sand.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Area percentage (min.–max.) of the substrates in % of the wood sections (WS, n = 9) and the control sections (CS, n = 3) of the streams Ihle, Nieplitz and Verlorenwasser. Gravel was not present in 3 of 9 sections of the Ihle River, for this reason the values (n = 9 and n = 6) are presented in the table.
Table 1.
Area percentage (min.–max.) of the substrates in % of the wood sections (WS, n = 9) and the control sections (CS, n = 3) of the streams Ihle, Nieplitz and Verlorenwasser. Gravel was not present in 3 of 9 sections of the Ihle River, for this reason the values (n = 9 and n = 6) are presented in the table.
| Substrate | Ihle | Nieplitz | Verlorenwasser |
|---|---|---|---|
| detritus (WS) | 37 (15–70) | 28 (26–43) | 34 (26–43) |
| sand (WS) | 48 (16–76) | 65 (16–76) | 60 (50–67) |
| wood (WS) | 9 (5–15) | 7 (2–15) | 7 (3–13) |
| gravel (WS) | 6 (0–14); with n = 6: 8 (6–14) | 0 | 0 |
| detritus (CS) | 58 (55–63) | 25 (23–29) | 16 (9–23) |
| sand (CS) | 42 (38–45) | 75 (71–76) | 84 (78–91) |
Table 2.
MI-colonization of streams Ihle, Nieplitz and Verlorenwasser.
| Ihle | Nieplitz | Verlorenwasser | |
|---|---|---|---|
| Total number of taxa (species richness) | 85 (without G, 2.06 m2) 89 (with G, 2.44 m2) | 65 (2.06 m2) | 57 (2.06 m2) |
| number of families | 46 (without G) 50 (with G) | 37 | 35 |
| number of orders | 14 (without and with G) | 13 | 13 |
| taxa of insects | 58 (without G) 61 (with G) | 47 | 38 |
| taxa of EPT | 36 (without G) 38 (with G) | 33 | 23 |
| individual density per m2 | 6215 (with G) | 20,492 | 5457 |
| dry matter in g/m2 | 11.05 (with G) | 56.86 | 8.90 |
Table 3.
Results of the correlation of the area percentages of the substrates with the density of individuals and the dry matter density of the MI (n = 12; ns = not significant).
Table 3.
Results of the correlation of the area percentages of the substrates with the density of individuals and the dry matter density of the MI (n = 12; ns = not significant).
| Individuals | Dry Matter | ||
|---|---|---|---|
| Ihle | sand | −0.71 (p < 0.05) | −0.59 (p < 0.05) |
| detritus | 0.59 (p < 0.05) | ns | |
| Nieplitz | wood | 0.73 (p < 0.05) | 0.72 (p < 0.05) |
| Verlorenwasser | sand | −0.58 (p = 0.05) | ns |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zigann, J.M.; Gersberg, R.M.; Lüderitz, V. Effects of Deadwood on Macroinvertebrate Assemblages in Three Sand-Type Lowland Streams. Ecologies 2023, 4, 88-105. https://doi.org/10.3390/ecologies4010008
AMA Style
Zigann JM, Gersberg RM, Lüderitz V. Effects of Deadwood on Macroinvertebrate Assemblages in Three Sand-Type Lowland Streams. Ecologies. 2023; 4(1):88-105. https://doi.org/10.3390/ecologies4010008
Chicago/Turabian StyleZigann, Julia Marie, Richard M. Gersberg, and Volker Lüderitz. 2023. "Effects of Deadwood on Macroinvertebrate Assemblages in Three Sand-Type Lowland Streams" Ecologies 4, no. 1: 88-105. https://doi.org/10.3390/ecologies4010008