
Wild Felid Diversity, Space Use and Activity Patterns in the Eastern Himalaya, India


1
Wildlife Institute of India, Chandrabani, Dehradun 248001, Uttarakhand, India
2
Centre of Research for Development, University of Kashmir, Hazratbal 190006, J&K, India
3
Department of Conservation Biology, Durgapur Govt. College, Durgapur 713214, WB, India
4
Namchi Government College, Kamrang, Namchi 737126, Sikkim, India
*
Author to whom correspondence should be addressed.
Ecologies 2023, 4(1), 41-54; https://doi.org/10.3390/ecologies4010005
Submission received: 28 November 2022
/
Revised: 10 January 2023
/
Accepted: 19 January 2023
/
Published: 25 January 2023
(This article belongs to the Special Issue Feature Papers of Ecologies 2022)
Abstract
:Species with similar resource requirements cannot coexist for long. To facilitate co-existence, carnivores adapt different strategies to partition the available resources in space and time. In high-altitude ecosystems with limited resources the operation becomes complex. We assessed species diversity and spatio-temporal resource partitioning among felid species inhabiting the intricate habitats of Khangchendzonga Biosphere Reserve (KBR), Eastern Himalaya. We used systematic camera trap surveys along with abundance and overlap indices to generate information on their space use and activity patterns. We recorded six species of felids belonging to five genera, including four species of global conservation importance. The differential use of space by felids was observed with respect to the habitat types determined by sharp altitudinal gradients with varied levels of spatial and altitudinal overlap. Maximum overlap was observed among Asiatic golden cat Catopuma temminckii, jungle cat Felis chaus and clouded leopard Neofelis nebulosa. The felids had distinct activity profiles from one other, particularly between golden cat and leopard cat Prionailurus bengalensis. Pianka’s index also provided weak evidence for temporal overlap among species. The overall results indicate that habitat heterogeneity facilitates primary separation at the spatial scale, although species are also segregated temporally. Golden cat was the most adaptive felid in the area, depicting a peculiar pattern of spatio-temporal segregation with other species. We suspect either intense competition or dietary segregation among some species, and propose a further investigation of their diet and the activity patterns of their prey. The study presents an early template of carnivore community organization in resource-scarce high-altitude environments, thus, offering huge ecological and conservation significance.
1. Introduction
Understanding the organization of ecological communities has long been a topic of interest among ecologists [1]. Resource availability is a key factor for shaping the community structure and its functioning, since it is affected by assemblages of species interacting in space and time [2,3,4]. As all ecosystems have limited resources, species coexist in a range of settings and are, therefore, subjected to various ecological processes at different scales [5,6,7]. In such an arrangement, even carnivores at the top of the food chain are forced to share their niches, which often results in an overlap in resource use, leading to coexistence or competition [8,9]. To facilitate co-existence and avoid competition, carnivores often adapt different strategies in order to partition the limited available resources [10,11,12,13]. These interactions become complex in high-altitude landscapes where resources are even scarcer [14].
According to the competitive exclusion principle, species with similar resource needs cannot coexist for very long [15]. Partitioning by the selection of prey species based on body sizes [16,17], foraging habits [18], activity patterns [16,19] and use of space [18,20] has been used for a long time to describe how carnivores coexist in natural systems [8,9]. The differential use of resources by carnivores therefore facilitates co-existence and influences community structure [21]. A way towards understanding community organizations can, therefore, be through a measure of resource use overlap between several species in a community guild at various ecological scales [10,13,21].
Owing to their high relevance towards understanding the functioning of the ecosystem at higher trophic levels, generating information on carnivore diversity and their interactions at spatial and temporal scale seems crucial [22,23]. Given the conservation importance of this group [24,25], less attention has been given to the non-fragmented habitats, particularly those in high-altitude landscapes such as the Himalayas. The primary reason for this has been the inaccessibility and ruggedness of the Himalayan terrain that makes field research and monitoring challenging [26]. Such intricacy of terrain and species diversity makes it more intriguing to understand the ways by which carnivores tend to cope in such resource-scarce environments. Since felids represent the most versatile and obligate flesh-eating carnivores at the highest trophic level [27] and receive top conservation priority among mammals [24,28,29], we made a serious attempt towards assessing their: (i) diversity and space use with respect to different habitat types and elevation gradients, (ii) temporal activity patterns, and (iii) the spatio-temporal overlap among species in the Khangchendzonga Biosphere Reserve (henceforth KBR) landscape. Our aim was to understand how felid species in the area segregate in space and time to partition the available resources in such resource-scarce environments. Given the intricacy of the terrain, the challenging field conditions and the elusive nature of the study species, a study like this has not still been performed at such altitudes in the Himalayas and, therefore, offers huge ecological and conservation advantages.
2. Material and Methods
2.1. Study Area
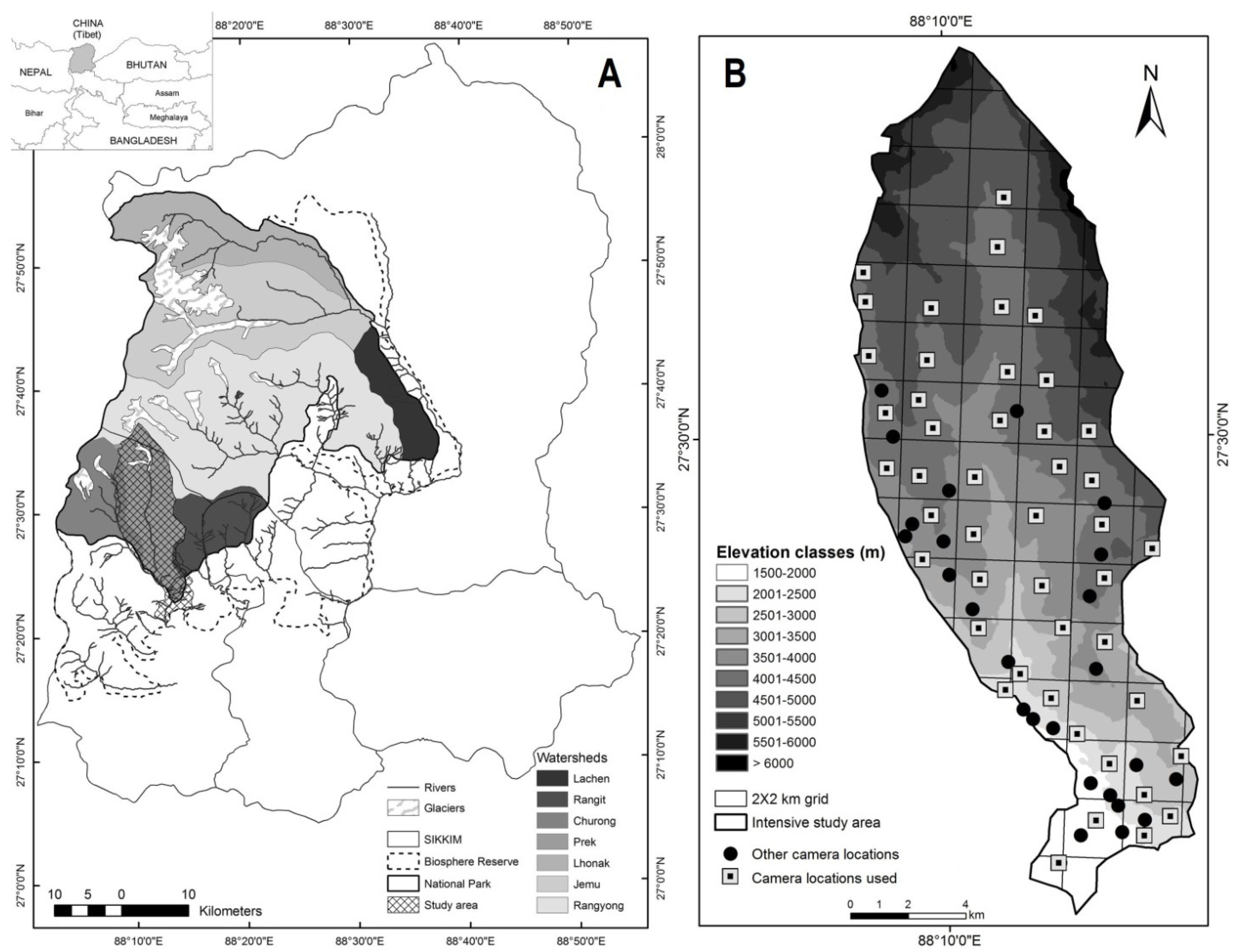
The study area represents the westernmost part of the Eastern Himalayan global biodiversity hotspot [30] and is home to the third highest peak in the world (Mt. Kanchenjunga [8586 m]). Positioned at the junction of two biogeographic realms, viz., the Palaearctic and the Oriental, the area has high potential for such ecological assessment, as it can sustain a variety of both ancient and recently evolved species of carnivores. The KBR landscape (2620 km2) includes a sharp variation in elevation from 1220 m to 8586 m, which is accompanied by an array of habitat types [31]. The KBR comprises seven river catchments, viz., Lhonak, Jemu, Lachen, Rangyong, Rangit, Prek and Churong (Figure 1A). Among these, we selected the Prek catchment (182 km2) for intensive sampling due to its topographic analogy with the entire KBR [32]. The Prek catchment has an altitudinal range of 2200 m to 6691 m, covering major habitat types such as mixed temperate (17%), sub-alpine (36%), alpine pastures (5%), rock and snow cover (41%), and water bodies (1%), respectively. We carried out our study over a period of over three years from 2010 to 2012. All field activities were carried out in the expedition mode due to the terrain’s topography and remoteness, and they involved trekking, camping and sampling different parts of the catchment.
2.2. Field Sampling and Camera Trapping
Reconnaissance surveys employing sign surveys, trail monitoring and ridge walking were conducted throughout the Prek catchment to identify appropriate locations for the deployment of camera traps [32]. The study area was divided into grids measuring 2 km × 2 km within a Geographic Information System (GIS), using ARC GIS 9.3 for better visualization of the habitat types and elevation categories in the area, as well as to monitor the camera deployment design. We deployed 27 infrared-triggered cameras at 71 different locations of likely animal-use based on the information generated during reconnaissance surveys. Cameras were fixed to tree trunks or rocks that were 3–5 m away from the expected animal activity and 15–30 cm above the ground. The cameras were programmed for 24-h monitoring across all seasons and checked at 15–20 days intervals. Information on the GPS coordinates, elevation and habitat type were noted at each camera location. To ensure efficient sampling design, at least one camera was deployed in each accessible grid by maintaining a certain inter-trap distance (minimum 500 m). Due to the failure of the cameras at few locations, data only from the cameras that were continuously operational for at least a period of one month were considered. Keeping the above criteria, 46 camera locations were finally considered for data use covering major habitat types in the area (Figure 1B).
2.3. Data Analysis
The spatial and elevational distribution of species was assessed with respect to their detection in cameras and interpreted in terms of their frequencies of occurrence. Space use by felids was represented in terms of their relative abundance index in each habitat type deduced from camera detections and interpreted as photo-capture rates [33]. We calculated photo capture rate (CR) as the number of independent photographs (with minimum 1 h duration) of a species captured at a given camera location, divided by the number of trap days for which the camera was operational at that location, and expressed per 100 trap days [34,35,36]. A Mann–Whitney U test was performed to test for variations in the CRs of species between different habitats. The cameras’ occurrence records of species were also analysed to calculate the spatial and altitudinal range overlap among pairs of felid species at different camera locations. Two species’ spatial overlap was assessed using the Jaccard similarity index (S), which ranges from 0 to 1 [37]. The index value 0 (no overlap) indicates detections at completely distinct camera locations and value 1 (complete overlap) indicates detections at exactly the same locations for both species. The altitudinal range overlap between two species was evaluated by calculating the fraction of overlapping range out of the species’ entire altitudinal range in the area.
The activity pattern of felids was evaluated with reference to the date and time stamps on the independent camera trap images of species, under the assumption that the quantity of photographs taken was correlated with the activity levels of the species [36,38]. The mean activity period of each species was calculated by converting all independent time records of each species for each hour into a percentage of photographic captures during that hour in a 24-h period, using the program Oriana 4.0 [39]. To determine if a species was nocturnal or diurnal, the activity level percentage was used. Nocturnal activity was considered from 1800 to 0500 h (the approximate times of sunset and sunrise in the study area). The Rayleigh test was used to assess the consistency of each felid species’ activity pattern. Watson’s U2 test was also used to examine the variations in activity patterns between felid species and within each felid species (for pairwise and single species). Further analysis was conducted on the percentage of photographic captures of species for each hour to assess the temporal overlap between their activity patterns (pairwise) through Pianka’s niche overlap index [40]. This was implemented using a simulation-based software program, TimeOverlap version 1.0 [41], which employs a randomization algorithm (ROSARIO) by performing random alterations of entire distributions within the time frame of analyses. For each randomization, the program calculates the amount of overlap of each group of randomized distributions for each iteration and determines its significance by a comparison of the randomized overlap values to the amount of empirical overlap. The analysis was performed involving 10,000 iterations at a 0.05 level of statistical significance.
3. Results
We obtained the sampling data of 6910 camera days from 46 camera locations during the study period, including 1862 in temperate (1200–3000 m), 3087 in sub-alpine (3000–4000 m), and 1961 in alpine (above 4000 m) habitats. We could confirm the occurrence of six species of felids in the study area belonging to five different genera, including all three species of leopard, viz., snow leopard Panthera uncia, clouded leopard Neofelis nebulosa and leopard Panthera pardus; the Asiatic golden cat Catopuma temminckii; leopard cat Prionailurus bengalensis; and jungle cat Felis chaus (Figure 2). Felid species were distributed throughout the catchment with varied frequency of occurrence in different habitat types. Overall, we documented five species from the temperate, three from the sub-alpine and one from the alpine habitats. Due to the elusory nature of the species, direct encounters were rare, viz., snow leopard (n = 1), leopard cat (n = 2) and jungle cat (n = 1).
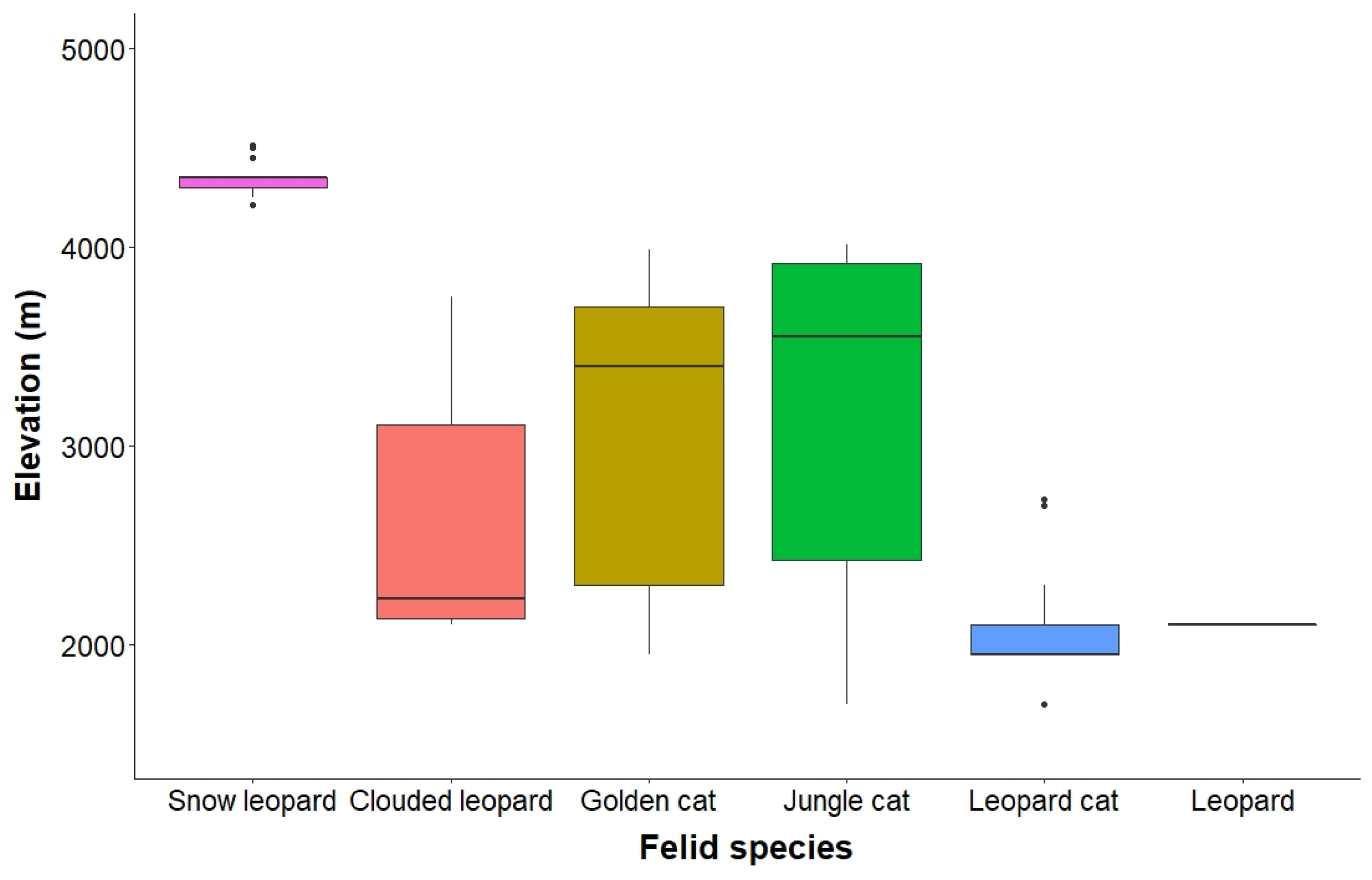
The occurrence of snow leopard was detected at six camera locations confined only to the alpine and rocky areas above 4000 m. Clouded leopard on the other hand was detected only at three camera locations positioned in sub-alpine and temperate habitats ranging from 2100 m to 3750 m (Figure 3). The occurrence of leopard cat was detected at 10 camera locations. Interestingly, the leopard was detected only once during the study. However, both these species were restricted only to the temperate habitat below 3000 m. Both golden cat and jungle cat were generally detected in temperate and sub-alpine habitats at 10 and 5 camera locations, though their highest elevation records were detected at 3990 m and 4010 m (in the Krummholz zone), respectively.
The relative space use by felids (expressed as capture rates) with relation to various habitat types are presented in Table 1. Although the overall trend in the relative space use index of felids was depicted as leopard cat > snow leopard > golden cat > jungle cat > clouded leopard > leopard, their capture rates did not vary significantly among habitat types. Furthermore, felid species showed varied levels of spatial and altitudinal overlap, as depicted by the Jaccard similarity index (S) and altitudinal range overlap values. The maximum overlap was, however, observed among golden cat–jungle cat–clouded leopard, while the minimum overlap was observed between snow leopard and other species (Table 2).
Among the six species of felids, temporal activity profiles could only be deduced for snow leopard, golden cat and leopard cat. A very low number of detections in the cameras limited the statistical evaluation of the activity patterns of clouded leopard (n = 6), jungle cat (n = 6) and leopard (n = 1). However, based on the percent frequency of records in the cameras, clouded leopard and jungle cat showed 67 and 83% of nocturnal activity, respectively. Furthermore, the single detection of leopard also occurred during night hours.
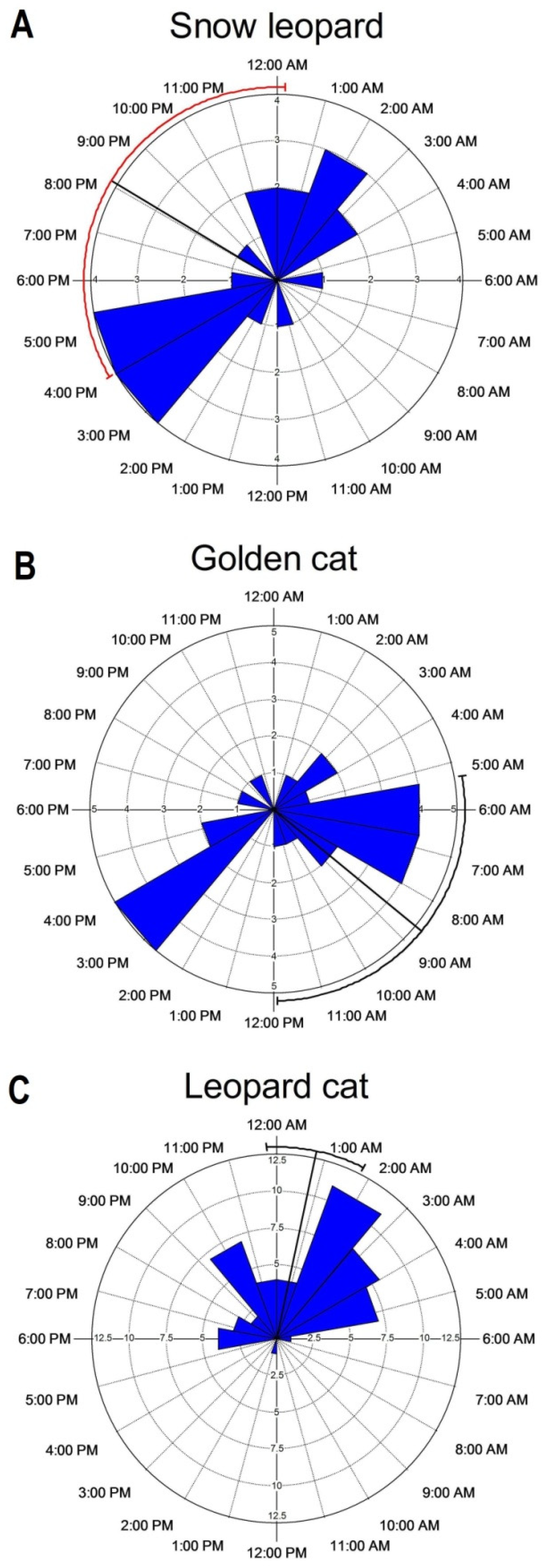
The activity profile of snow leopard was bimodal in general. Despite being more active later in the day, around sunset and at midnight (Figure 4A), its overall activity pattern was mostly constant (Rayleigh Z = 1.622, p = 0.199), with no significant difference between activities at different times of the day (Watson’s U2 = 0.186, p > 0.05). The circular statistics (below) also indicate the mean time activity of the snow leopard at 20:09 ± 02:04 h (Table 3). The golden cat displayed an irregular pattern of activity and was more active during crepuscular hours (Figure 4B). Although the activity profile was virtually evenly distributed throughout the day (Z = 2.436, p = 0.087), the intensity of activity varied significantly during the day (U2 = 0.215, p < 0.05). On the contrary, typical nocturnal behaviour was displayed by the leopard cat (Figure 4C), with mean time activity at 00:51 ± 00:30 h, and significant non-uniform and distinct pattern during different times of the day (Z = 21.381, p < 0.001; U2 = 1.187, p < 0.005). The activity profiles of felids were different from one another, as indicated by the pairwise Watson’s U2 test, and the most significant difference was observed between golden cat and leopard cat (U2 = 0.865, p < 0.001; Table 3). On the other hand, Pianka’s index also showed weak support for temporal overlap in the activity patterns of the felids.
4. Discussion
Landscapes provide the ecological template for the life history and behavioural processes of species through their ecological complexity and flexibility [42], which ultimately determine the spacing patterns and habitat use of animals. Our study provides the first photographic documentation of six felid species from KBR, comprising four species with significant global conservation importance (three Vulnerable and one Near-Threatened) listed in the IUCN Red List of Threatened Species. Belonging to five genera, these represent elements from both the Palaearctic (snow leopard, leopard) and Oriental (clouded leopard, Asiatic golden cat, jungle cat, leopard cat) origins [43]. The overall distribution patterns of felids with respect to elevation and habitat types validate the ecological significance of the area, both in terms of its habitat diversity and biogeography. However, the decrease in the felid diversity with an increase in elevation (from temperate to alpine) demonstrates the typical Eastern Himalayan trend in species distributions, indicating more diversity in the middle elevation range, i.e., 2000–3500 m [32,44].
4.1. Space Use
The distribution of carnivores in general is essentially determined by the spatial arrangement of their prey [13,22]. However, in the Himalayas, habitat and elevation also have a significant impact on how carnivores utilise their surroundings. In the Prek catchment of the KBR, felids were dispersed according to several habitats dictated by severe elevation gradients with a considerable amount of overlap among their ranges. Although, some species (snow leopard, leopard, and leopard cat) were restricted to a specific habitat or elevation class, most of them (Asiatic golden cat, clouded leopard, and jungle cat) were wide ranging.
The confined distribution of snow leopard to rocky and alpine habitat corresponds to the species altitudinal range (3000–5400 m) reported in previous studies [45]. Its occurrence strictly above 4000 m (alpine and rocky areas) is attributed to the higher treeline, which is a characteristic of Eastern Himalaya. However, frequent detections of snow leopard at other specific camera locations could be related to their positioning around sites that are well connected with the sub-alpine and Krummholz areas of the Rangit catchment, and the alpine and rocky habitats of the Somiti–Goechala and Dzongri–Rathong areas. Such habitat connectedness would allow the snow leopard to access a variety of prey sources from various areas (such as Himalayan tahr and musk deer from sub-alpine and Krummholz, and blue sheep from alpine). Furthermore, continuity of the region with the Kanchenjunga Conservation Area of Nepal also facilitates the cross-border movement of snow leopards, thus, enhancing their detection in the sampled area.
Clouded leopard, on the other hand, mainly inhabits dense evergreen forests [46]. Its occurrence in the upper temperate and sub-alpine habitat validates its tendency for similar areas due to its arboreal habit [46,47]. The current study marks a new altitudinal record for clouded leopard at 3750 m, which stands much higher compared to the species’ earlier record (up to 2500 m) in the Himalayan foothills [47], thus, indicating a wider altitudinal range for the species in the Eastern Himalayas. This could again be due to the higher treeline in the Eastern Himalaya that supports forest dwelling species such as clouded leopard to explore higher elevation, and help them minimize competition with other carnivores. Furthermore, the occurrence of Asiatic golden cat at 3990 m in the sub-alpine and Krummholz habitat also represents its adaptability to montane forests (dominated by Abies and Rhododendron) and treeline with dwarf rhododendrons. The species is otherwise known to primarily inhabit tropical and sub-tropical evergreen to mixed and dry deciduous forests at lower elevations [47,48].
The restricted distribution of leopard cat to the sub-tropical and temperate habitats below 2750 m observed in our study does not seem comparable to the overall wide altitudinal range of the species extending up to 4474 m [49,50,51]. However, such a peculiar space use pattern of leopard cat could indicate a strategy to avoid competition with the other carnivores, by occupying lower areas close to human habitation in the buffer zone with access to domestic prey, as also depicted through its narrow spatial overlap with other felids in the area.
The occurrence of jungle cat at 4010 m also marks an interesting altitudinal record contrary to its general preference for lowland habitats [47,52]. Compared to other felids, the low detection of leopard outside its versatile habitat range reaching 5200 m in Himalaya [47] could either indicate a restricted range distribution to lower sub-tropical areas (below the lowest elevation limit of the study area), or a strategy to avoid low prey (medium to large sized prey) density areas of the region. Another potential reason for its rare detection could be the recent persecution by local people in retaliation to their livestock losses that almost eradicated leopards from the lower areas of the Prek catchment in KBR (Usha Lachungpa pers comm). In reference to this, leopard occurrence is generally reported from the low-elevation sub-tropical areas of the Khangchendzonga landscape below the lowest elevation of our study area.
4.2. Activity Pattern
Although species activity patterns are a part of the fundamental knowledge of natural history, they are helpful in understanding the mechanisms that shape ecological communities. Activity patterns help us to know whether potentially competing species overlap or avoid each other temporally, and how larger species influence activities of their smaller cohorts in the same habitat. In the present study, snow leopard was found to be mostly active at twilight and during the night, lying nearly in coherence with the observations in Southern Mongolia [53]. The species is typically thought to be primarily nocturnal [54]; however a few studies have also shown its pattern of activity during the crepuscular hours [55,56]. Such expanding activity pattern could be related to the scarcity of prey base in the area that compels the snow leopard to spend more time active in foraging. Moreover, snow leopards are known to benefit from the cover of darkness, as the semi-darkness of twilight would appear optimal for stalking the prey [57]. The strict nocturnal activity pattern of the leopard cat was, however, in coherence with previous findings [38,58,59], but it also contradicted a few other studies reporting crepuscular [60], arrhythmic [61] and diurnal activity [62] patterns. This could be explained in a way that the daily activity of small felids is associated with the activity pattern of their principal prey, i.e., murids, in the case of leopard cat [59,63], which was also confirmed through our previous study in the area [64].
Asiatic golden cats, on the other hand, are noted for their nocturnal behaviour [65]. However, in reference to the present records (arrhythmic and crepuscular activity) as well as based on other studies (indicating daytime and crepuscular activity peaks), the species may not be considered exclusively nocturnal, as previously thought [46,66]. Other factors, such as prey distribution and activity, inter- and intra-guild structure, habitat complexity and topography, and competition could also play a significant role in shaping the species activity profile in the area.
In the case of other species (jungle cat, clouded leopard and leopard), due to the scarce number of records in the cameras, no conclusive inference could be drawn in regard to their activity profiles. However, their dominant nocturnal activity patterns were both in coherence (for clouded leopard) as well as contradiction (for jungle cat and leopard) with the species’ general behavioural ecology [67]. More field-based information is, therefore, required to further elucidate the activity profiles of these species in KBR, which is yet to be performed.
4.3. Spatio-Temporal Overlap and Segregation
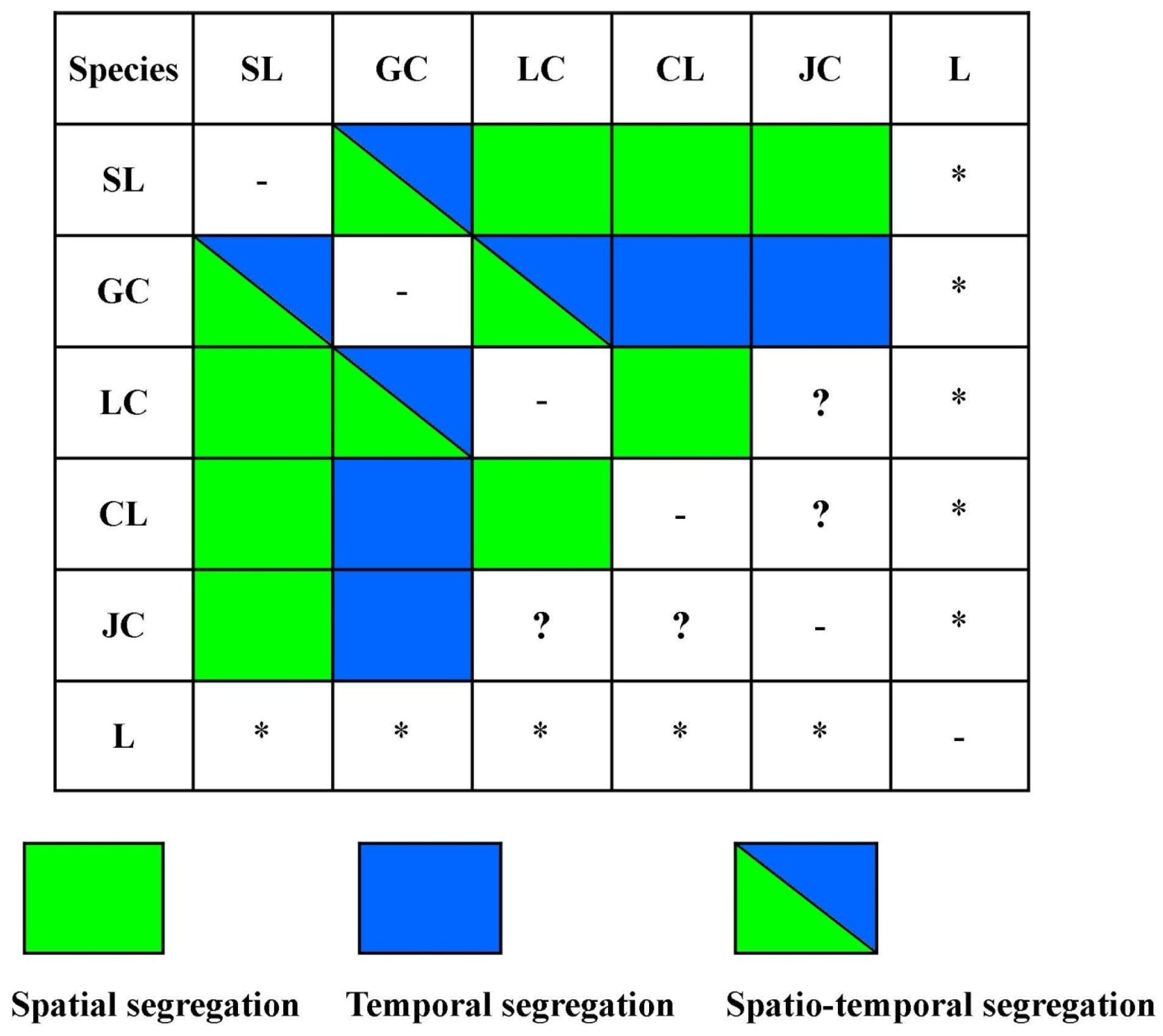
Niche theory states that when two or more species co-exist, their usage of the environment, diet, activity and effect on the community structure does not entirely overlap [10,68,69]. In order to facilitate co-existence, minimize competition and cope with limited resources, species segregate at different scales to partition the use of available resources [70,71]. Differential resource selection therefore operates as a fundamental process enabling species to coexist. The overall findings of the present study suggest that the felid assemblage inhabiting this high-altitude ecosystem of Khangchendzonga has a tendency to partition the available resources at distinct scales. As the region is shaped into vast altitudinal gradients and a wide variety of habitats, it is further influenced by diverse weather conditions. The evolutionary adaptation of carnivores to such conditions may, therefore, favour a differential pattern of space use in such diverse landscapes [67,72] and, thus, allow primary segregation at the spatial scale, as observed. This was shown by snow leopard (confined to >4000 m) showing spatial (altitudinal) segregation with other felids, and also between clouded leopard and leopard cat (Figure 5). This elucidates niche complementarily between snow leopard and other felids involving low spatial overlap and high temporal overlap between species [13,68,69]. Strong spatial overlap among golden cat–jungle cat–clouded leopard associated with the arrhythmic and crepuscular activity of golden cat indicates resource sharing by golden cat only at the spatial scale during different time periods of the day, thus, depicting temporal segregation. The similar (nocturnal) activity pattern of clouded leopard and jungle cat, however, indicates both spatial and temporal overlap among the two species, thereby suggesting segregation at some different (e.g., dietary) scale or competition among the species. Based on their spatio-temporal patterns, a similar inference could be drawn while explaining the mode of resource partitioning between leopard cat and jungle cat, considering diet as an important axes of niche space. Due to very limited information available on the leopard, no attempt was made to derive any inference on its resource use pattern in the area. Furthermore, we observed that golden cat occupied the middle elevation range of the study area and showed spatio-temporal segregation with lowland (e.g., leopard cat) and highland (e.g., snow leopard) species. However, with respect to other middle elevation species (i.e., clouded leopard and jungle cat), golden cat depicted temporal segregation. Such a peculiar pattern of spatio-temporal segregation signifies the versatility of golden cat in the region, further corroborating its adaptive behaviour, as already exhibited through its phenotypic polymorphism reported from the study area [73].
Owing to the similarities among felid species in terms of their body size and basic ecology, intense spatial and temporal overlaps may result in competition both at interspecific and intraspecific levels. This may compel species to move to new areas with an increased risk of human interaction, thus, leading to conflict. Such an antagonistic relationship between carnivores could even become intense among species of a similar body size, e.g., leopard cat and jungle in the present context, thereby disrupting the functioning of the ecosystem.
5. Conclusions
Through this study, we attempted to assess the spatio-temporal aspects of the ecology of these little-known elusive species of wild felids. Give the small dataset, our inference on resource partitioning remains preliminary and requires further studies to validate the patterns observed. Considering the range and overlap of body sizes between species, we expect dietary segregation to play a major role in facilitating co-existence among the felids of KBR. However, this explanation needs to be tested through further investigation on the diet and activity patterns of species and their primary prey. This can aid in obtaining a better understanding of the mechanisms by which carnivore communities structure themselves in such intricate and fragile environments. The ecological data generated in this study from an exhaustive sampling effort are of key significance, as they can set the tone for more comprehensive studies, including a detailed assessment of the carnivore prey base in the region, extending the preliminary information generated in this study (Supplementary Table S1). The study underlines the immense scope for further research in the area, validates the importance of the region as a high-altitude biodiversity hotspot, and warrants its effective conservation and management.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/ecologies4010005/s1, Table S1: Photo-capture rate (±standard error) of potential prey species of carnivores in different habitats of the Prek catchment of Khangchendzonga Biosphere Reserve.
Author Contributions
Conceptualization, T.B. (Tawqir Bashir) and S.S.; Methodology, T.B. (Tawqir Bashir); Software, T.B. (Tawqir Bashir); Validation, T.B. (Tawqir Bashir) and S.S.; Formal Analysis, T.B. (Tawqir Bashir); Investigation, T.B. (Tawqir Bashir), T.B. (Tapajit Bhattacharya) and K.P.; Resources, T.B. (Tawqir Bashir) and S.S.; Data Curation, T.B. (Tawqir Bashir); Writing—Original Draft Preparation, T.B. (Tawqir Bashir); Writing—Review and Editing, T.B. (Tawqir Bashir), T.B. (Tapajit Bhattacharya) and S.S.; Visualization, T.B. (Tawqir Bashir); Supervision, S.S.; Project Administration, T.B. (Tawqir Bashir) and S.S.; Funding Acquisition, S.S. All authors have read and agreed to the published version of the manuscript.
Funding
The funding support for conducting this study including field surveys, procurement of equipments and the infrastructural facilities were provided by the Wildlife Institute of India from its Grant-in-Aid (WII/GRT/208).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author. The data are not publicly available due to institutional data privacy policy.
Acknowledgments
We thank the Wildlife Institute of India, Dehradun, for providing the funding support required for conducting this study. We are grateful to the Department of Forests, Environment and Wildlife Management, Government of Sikkim, for providing research permits for our field surveys. The first author appreciates the support from the DST-INSPIRE Faculty award grant that supported the data analysis and writing of the draft manuscript. We also thank the three anonymous reviewers whose comments significantly helped in improving the quality of the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Colorado-Zuluaga, G.J. How ecological communities are structured: A review on ecological assembly rules. Revista EIA 2015, 12, 27–53. [Google Scholar]
- Lockwood, J.; Powell, R.D.; Nott, M.P.; Pimm, S.L. Assembling ecological communities in time and space. Oikos 2017, 80, 549–553. [Google Scholar] [CrossRef]
- Fox, B.J.; Fox, M.D. Factors determining mammal species richness on habitat islands and isolates: Habitat diversity, disturbance, species interactions and guild assembly rules. Glob. Ecol. Biogeogr. 2000, 9, 19–37. [Google Scholar] [CrossRef]
- Zupo, V.; Alexander, T.J.; Edgar, G.J. Relating trophic resources to community structure: A predictive index of food availability. R. Soc. Open Sci. 2017, 4, 160515. [Google Scholar] [CrossRef] [Green Version]
- Kelt, D.A.; Taper, M.L.; Meserve, P.L. Assessing the impact of competition on the assembly of communities: A case study using small mammals. Ecology 1995, 76, 1283–1296. [Google Scholar] [CrossRef]
- Campbell, L.A. Distribution and Habitat Associations of Mammalian Carnivores in the Central and Southern Sierra Nevada. Ph.D. Thesis, University of California, Davis, CA, USA, 2004. [Google Scholar]
- Tadesse, S.A. Community structure and trophic level interactions in the terrestrial ecosystems: A review. Int. J. Avian. Wildl. Biol. 2017, 2, 185–192. [Google Scholar] [CrossRef] [Green Version]
- Linnell, J.D.C.; Strand, O. Interference interactions, co-existence and conservation of mammalian carnivores. Divers. Distrib. 2000, 6, 169–176. [Google Scholar]
- De Satgé, J.; Teichman, K.; Cristescu, B. Competition and coexistence in a small carnivore guild. Oecologia 2017, 184, 873–884. [Google Scholar] [CrossRef]
- Schoener, T.W. Resource partitioning in ecological communities. Science 1974, 185, 27–39. [Google Scholar]
- Sunquist, M.E.; Sunquist, F.C. Ecological constraints on predation by large felids. In Carnivore Behavior, Ecology and Evolution; Gittleman, J.L., Ed.; Cornell University Press: Ithaca, NY, USA, 1989; pp. 283–301. [Google Scholar]
- Di Bitetti, M.S.; De Angelo, C.D.; Di Blanco, Y.E.; Paviolo, A. Niche partitioning and species coexistence in a Neotropical felid assemblage. Acta. Oecol. 2010, 36, 403–412. [Google Scholar]
- Sunarto, S.; Kelly, M.J.; Parakkasi, K.; Hutajulu, M.B. Cat coexistence in central Sumatra: Ecological characteristics, spatial and temporal overlap, and implications for management. J. Zool. 2015, 296, 104–115. [Google Scholar]
- Davis, M.L.; Kelly, M.J.; Staufer, D.F. Carnivore co-existence and habitat use in the Mountain Pine Ridge Forest Reserve, Belize. Anim. Conserv. 2011, 14, 56–65. [Google Scholar] [CrossRef]
- Kneitel, J. Gause’s Competitive Exclusion Principle. In Encyclopedia of Ecology; Jørgensen, S.E., Fath, B.D., Eds.; Academic Press: Cambridge, MA, USA, 2008; pp. 1731–1734. [Google Scholar]
- Karanth, K.U.; Sunquist, M.E. Behavioral correlates of predation by tiger (Panthera tigris), leopard (Panthera pardus) and dhole (Cuon alpinus) in Nagarahole, India. J. Zool. 2000, 250, 255–265. [Google Scholar] [CrossRef]
- Gittleman, J.L. Carnivore body size; ecological and taxonomical correlates. Oecologia 1985, 67, 540–554. [Google Scholar] [PubMed]
- Palomares, F.; Ferreras, P.; Fedriani, J.M.; Delibes, M. Spatial relationships between Iberian lynx and other carnivores in an area of south-western Spain. J. Appl. Ecol. 1996, 33, 5–13. [Google Scholar] [CrossRef]
- Fedriani, J.M.; Palomares, F.; Delibes, M. Niche relationships among three sympatric Mediterranean carnivores. Oecologia 1999, 121, 138–148. [Google Scholar] [CrossRef] [Green Version]
- Durant, S.M. Competition refuges and co-existence: An example from Serengeti carnivores. J. Anim. Ecol. 1998, 67, 370–386. [Google Scholar] [CrossRef] [Green Version]
- Simberlof, D.; Dayan, T. The guild concept and the structure of ecological communities. Annu. Rev. Ecol. Syst. 1991, 22, 115–143. [Google Scholar] [CrossRef]
- Karanth, K.U.; Srivathsa, A.; Vasudev, D.; Puri, M.; Parameshwaran, R.; Kumar, N.S. Spatio-temporal interactions facilitate large carnivore sympatry across a resource gradient. Proc. Biol. Sci. 2017, 284, 20161860. [Google Scholar] [CrossRef] [Green Version]
- Hoeks, S.; Huijbregts, M.A.J.; Busana, M.; Harfoot, M.B.J.; Svenning, J.-C.; Santini, L. Mechanistic insights into the role of large carnivores for ecosystem structure and functioning. Ecography 2020, 43, 1752–1763. [Google Scholar] [CrossRef]
- Ceballos, G.; Erlich, P.R.; Soberon, J.; Salazar, I.; Fay, J.P. Global mammal conservation: What must we manage? Science 2005, 309, 603–607. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Sepúlveda, J.; Martín, C.A. Conservation status of the world’s carnivorous mammals (order Carnivora). Mamm. Biol. 2022. [Google Scholar] [CrossRef]
- Kuemmerle, T.; Bluhm, H.; Ghoddousi, A.; Arakelyan, M.; Askerov, E.; Bleyhl, B.; Ghasabian, M.; Gavashelishvili, A.; Heidelberg, A.; Malkhasyan, A.; et al. Identifying priority areas for restoring mountain ungulates in the Caucasus ecoregion. Conserv. Sci. Pract. 2020, 2, e276. [Google Scholar]
- Macdonald, D.W.; Loveridge, A.J. (Eds.) Biology and Conservation of Wild Felids; Oxford University Press: Oxford, UK, 2010; p. 762. [Google Scholar]
- Brodie, J.F. Is research effort allocated efficiently for conservation? Felidae as a global case study. Biodivers. Conserv. 2009, 18, 2927–2939. [Google Scholar] [CrossRef]
- Macdonald, E.A.; Burnham, D.; Hinks, A.E.; Dickman, A.J.; Malhi, Y.; Macdonald, D.W. Conservation inequality and the charismatic cat: Felis felicis. Glob. Ecol. Conserv. 2015, 3, 851–866. [Google Scholar] [CrossRef] [Green Version]
- Myers, N.; Mittermier, R.A.; Mittermier, C.G.; da Fonseca, G.A.B.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Tambe, S. Ecology and Management of the Alpine Landscape in the Khangchendzonga National Park, Sikkim Himalaya. Ph.D. Thesis, FRI University, Dehradun, India, 2007; p. 232. [Google Scholar]
- Sathyakumar, S.; Bashir, T.; Bhattacharya, T.; Poudyal, K. Assessing mammal distribution and abundance in intricate Eastern Himalayan habitats of Khangchendzonga, Sikkim, India. Mammalia 2011, 75, 257–268. [Google Scholar] [CrossRef]
- Carbone, C.; Christie, S.; Conforti, K.; Coulson, T.; Franklin, N.; Ginsberg, J.R.; Griffiths, M.; Holden, J.; Kawanishi, K.; Kinnaird, M.F.; et al. The use of photographic rates to estimate densities of tigers and other cryptic mammals. Anim. Conserv. 2001, 4, 75–79. [Google Scholar] [CrossRef] [Green Version]
- Kawanishi, K.; Sahak, A.M.; Sunquist, M. Preliminary analysis on abundance of large mammals at Sungai Relau, Taman Negara. J. Wildli. Parks. 1999, 17, 62–82. [Google Scholar]
- O’Brien, T.G.; Kinnaird, M.F.; Wibisono, H.T. Crouching tigers, hidden prey: Sumatran tiger and prey populations in a tropical forest landscape. Anim. Conserv. 2003, 6, 131–139. [Google Scholar] [CrossRef] [Green Version]
- Bowkett, A.E.; Rovero, F.; Marshall, A.R. The use of camera-trap data to model habitat use by antelope species in the Udzungwa Mountain forests, Tanzania. Afr. J. Ecol. 2007, 46, 479–487. [Google Scholar] [CrossRef]
- Magguran, A.E. Measuring Biological Diversity; Blackwell Publishing: Oxford, UK, 2004; p. 215. [Google Scholar]
- Lynam, A.J.; Jenks, K.E.; Tantipisanuh, N.; Chutipong, W.; Ngoprasert, D.; Steinmetz, R.; Sukmasuang, R.; Grassman, L.I., Jr.; Cutter, P.; Kitamura, S. Terrestrial activity patterns of wild cats from camera-trapping. Raffles Bull. Zool. 2013, 61, 407–415. [Google Scholar]
- Kovach, W.L. Oriana—Circular Statistics for Windows, ver. 4.; Kovach Computing Services: Pentraeth, Wale, 2011. [Google Scholar]
- Pianka, E.R. The structure of lizard communities. Annu. Rev. Ecol. Evol. Syst. 1973, 4, 53–74. [Google Scholar] [CrossRef] [Green Version]
- Castro-Arellano, I.; Lacher, T.E., Jr.; Willig, M.R.; Rangel, T. Assessment of assemblage-wide temporal niche segregation using null models. Methods Ecol. Evol. 2010, 1, 311–318. [Google Scholar]
- Davies, A.B.; Tambling, C.J.; Marneweck, D.G.; Ranc, N.; Druce, D.J.; Cromsigt, J.P.G.M.; le Roux, E.; Asner, G.P. Spatial heterogeneity facilitates carnivore coexistence. Ecology 2021, 102, e03319. [Google Scholar] [PubMed]
- Chattopadhayay, S.; Saha, S.S.; Ghosh, M.K.; Agrawal, V.C. Mammalia. In Fauna of Sikkim; State Fauna Series, 9 (Part1); Zoological Survey of India: Kolkata, India, 2006; pp. 33–76. [Google Scholar]
- Mishra, C.; Madhusudanand, M.D.; Datta, A. Mammals of the high altitudes of western Arunachal Pradesh, Eastern Himalaya: An assessment of threats and conservation needs. Oryx 2006, 40, 29–35. [Google Scholar] [CrossRef] [Green Version]
- McCarthy, T.; Mallon, D.; Jackson, R.; Zahler, P.; McCarthy, K. Panthera Uncia; The IUCN Red List of Threatened Species; IUCN: Gland, Switzerland, 2017; p. e.T22732A50664030. [Google Scholar] [CrossRef]
- Grassman, L.I.; Tewes, M.E.; Silvy, N.J.; Kreetiyutanont, K. Ecology of three sympatric felids in a mixed evergreen forest in north-central Thailand. J. Mammal 2005, 86, 29–38. [Google Scholar] [CrossRef]
- Nowell, K.; Jackson, P. Wild Cats: Status Survey and Conservation Action Plan; IUCN/SSC Cat Specialist Group, IUCN: Gland, Switzerland, 1996; p. 382. [Google Scholar]
- Choudhury, A. Sighting of Asiatic golden cat in the grasslands of Assam’s Manas National Park. Cat News 2007, 47, 29. [Google Scholar]
- Grassman, L.I.; Tewes, M.E.; Silvy, N.J.; Kreetiyutanont, K. Spatial organization and diet of the leopard cat (Prionailurus bengalensis) in north-central Thailand. J. Zool. 2005, 266, 45–54. [Google Scholar] [CrossRef]
- Rajaratnam, R.; Sunquist, M.; Rajaratnam, L.; Ambu, L. Diet and habitat selection of the leopard cat (Prionailurus bengalensis borneoensis) in an agricultural landscape in Sabah, Malaysian Borneo. J. Trop. Ecol. 2007, 23, 209–217. [Google Scholar] [CrossRef]
- Thapa, K.; Pradhan, N.; Barker, J.; Dahal, M.; Bhandari, A.R.; Gurung, G.S.; Rai, D.P.; Thapa, G.J.; Shrestha, S.; Singh, G.R. High elevation record of a leopard cat in the Kanchenjunga Conservation Area, Nepal. Cat News 2013, 58, 26–27. [Google Scholar]
- Duckworth, J.W.; Poole, C.M.; Tizard, R.J.; Walston, J.L.; Timmins, R.J. The Jungle Cat Felis chaus in Indochina: A threatened population of a widespread and adaptable species. Biodivers. Conserv. 2005, 14, 1263–1280. [Google Scholar] [CrossRef]
- McCarthy, T.M.; Fuller, T.K.; Munkhtsog, B. Movements and activities of snow leopard in South-western Mongolia. Biol. Conserv. 2005, 124, 527–537. [Google Scholar] [CrossRef]
- Mallon, D. The snow leopard in Ladakh. In International Pedigree Book of Snow Leopards; Helsinki Zoo: Helsinki, Finland, 1984; Volume 4, pp. 23–37. [Google Scholar]
- Schaller, G.B.; Tserendeleg, J.; Amarsanaa, G. Observations on snow leopards in Mongolia. Proc. Int. Snow Leopard Symp. 1994, 7, 33–42. [Google Scholar]
- Jackson, R.M. Home Range, Movements and Habitat Use of Snow Leopard (Uncia uncia) in Nepal. Ph.D. Thesis, University of London, London, UK, 1996; p. 233. [Google Scholar]
- Johansson, O. Unveiling the Ghost of the Mountain; Snow Leopard Ecology and Behaviour. Ph.D. Thesis, Faculty of Forest Science, Department of Ecology, Swedish University of Agricultural Science, Uppsala, Sweden, 2017; p. 51. [Google Scholar]
- Cheyne, S.M.; Macdonald, D.W. Wild felid diversity and activity patterns in Sabangau peat-swamp forest, Indonesian Borneo. Oryx 2011, 45, 119–124. [Google Scholar]
- Rostro-García, S.; Kamler, J.F.; Minge, C.; Caragiulo, A.; Crouthers, R.; Groenenberg, M.; Gray, T.N.E.; In, V.; Pin, C.; Sovanna, P.; et al. Small cats in big trouble? Diet, activity, and habitat use of jungle cats and leopard cats in threatened dry deciduous forests, Cambodia. Ecol. Evol. 2021, 11, 4205–4217. [Google Scholar] [CrossRef]
- Grassman, L.I.; Haines, A.M.; Janečka, J.E.; Tewes, M.E. Activity periods of photo-captured mammals in north central Thailand. Mammalia 2006, 70, 306–309. [Google Scholar] [CrossRef]
- Austin, S.C.; Tewes, M.E.; Grassman, L.I., Jr.; Silvy, N.J. Ecology and conservation of leopard cat Prionailurus bengalensis and clouded leopard Neofelis Nebulosa in Khao Yai National Park, Thailand. Acta Zool. Sin. 2007, 53, 1–14. [Google Scholar]
- Saxena, A.; Rajvanshi, A. Diurnal activity of leopard cat in Rajaji National Park, India. Cat News 2014, 61, 21. [Google Scholar]
- Schmidt, K.; Nakanishi, N.; Izawa, M.; Okamura, M.; Watanabe, S.; Tanaka, S.; Doi, T. The reproductive tactics and activity patterns of solitary carnivores: The Iriomote cat. J. Ethol. 2009, 27, 165–174. [Google Scholar] [CrossRef]
- Bashir, T.; Bhattacharya, T.; Poudyal, K.; Sathyakumar, S.; Qureshi, Q. Integrating aspects of ecology and predictive modeling: Implications for the conservation of the leopard cat (Prionailurus bengalensis) in the Eastern Himalaya. Acta Theriol. 2014, 59, 35–47. [Google Scholar] [CrossRef]
- Kawanishi, K.; Sunquist, M.E. Food habits and activity patterns of the Asiatic golden cat (Catopuma temminckii) and dhole (Cuon alpinus) in a primary rainforest of Peninsular Malaysia. Mamm. Stud. 2008, 33, 173–177. [Google Scholar] [CrossRef]
- Azlan, J.M.; Sharma, D.S.K. The diversity and activity patterns of wild felids in a secondary forest in Peninsular Malaysia. Oryx 2006, 40, 36–41. [Google Scholar] [CrossRef] [Green Version]
- Gittleman, J.L. Carnivore Behavior, Ecology and Evolution; Cornell University Press: Ithaca, NY, USA; London, UK, 1996; Volume 2, p. 664. [Google Scholar]
- Cornell, H. Niche Overlap. In Encyclopedia of Theoretical Ecology; Hastings, A., Gross, L.J., Eds.; University of California: Berkeley, CA, USA, 2011; pp. 489–498. [Google Scholar]
- Mittelbach, G.G.; McGill, B.J. Species coexistence and niche theory. In Community Ecology, 2nd ed.; Oxford Academic: Oxford, UK, 2019; pp. 141–157. [Google Scholar]
- Pringle, R.M. Ecology: A revolution in resource partitioning. Curr. Biol. 2021, 31, R1474–R1476. [Google Scholar] [CrossRef]
- Curras, M.R.; Donadio, E.; Middleton, A.D.; Pauli, J.N. Carnivore Niche Partitioning in a Human Landscape. Am. Nat. 2022, 199, 496–509. [Google Scholar] [CrossRef]
- Sunquist, M.; Sunquist, F. Wild Cats of the World; University of Chicago Press: Chicago, IL, USA, 2000; p. 452. [Google Scholar]
- Bashir, T.; Bhattacharya, T.; Poudyal, K.; Sathyakumar, S. Notable observations on the melanistic Asiatic Golden cat (Pardofelis temminckii) of Sikkim, India. NeBIO 2011, 2, 1–4. [Google Scholar]
Figure 1.
Location of Khangchendzonga Biosphere Reserve in Sikkim: (A) the inset’s shaded region indicates the positioning of Sikkim in north-east India, showing the National Park and the seven watersheds, including the Prek catchment (study area); (B) digital elevation model (DEM) of the Prek catchment showing locations of cameras deployed in the area.
Figure 1.
Location of Khangchendzonga Biosphere Reserve in Sikkim: (A) the inset’s shaded region indicates the positioning of Sikkim in north-east India, showing the National Park and the seven watersheds, including the Prek catchment (study area); (B) digital elevation model (DEM) of the Prek catchment showing locations of cameras deployed in the area.

Figure 2.
Camera trap photos of the felid species recorded in the study area; (A) snow leopard; (B) clouded leopard; (C) leopard; (D) Asiatic golden cat; (E) leopard cat; and (F) jungle cat.
Figure 2.
Camera trap photos of the felid species recorded in the study area; (A) snow leopard; (B) clouded leopard; (C) leopard; (D) Asiatic golden cat; (E) leopard cat; and (F) jungle cat.

Figure 3.
Boxplot showing the altitudinal distribution of six felid species recorded in the KBR.

Figure 4.
Temporal activity patterns of felids in the Prek catchment of KBR.

Figure 5.
Block chart depicting segregation and resource partitioning among the felid species at different temporal and spatial scales. (?) suggests segregation at some different scale, (-) shows no relationship within the same species and (*) indicates insufficient data to generate meaningful inference on resource portioning between species.
Figure 5.
Block chart depicting segregation and resource partitioning among the felid species at different temporal and spatial scales. (?) suggests segregation at some different scale, (-) shows no relationship within the same species and (*) indicates insufficient data to generate meaningful inference on resource portioning between species.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Photo-capture rate (±standard error) of felids in different habitats of the Prek catchment with Mann–Whitney U statistics (p denotes statistical significance).
Table 1.
Photo-capture rate (±standard error) of felids in different habitats of the Prek catchment with Mann–Whitney U statistics (p denotes statistical significance).
| Species | N | Temperate | Sub-Alpine | Alpine | Overall CR | MW-U χ2 | p |
|---|---|---|---|---|---|---|---|
| Snow leopard | 24 | - | - | 0.85 (0.35) | 0.85 (0.35) | - | - |
| Clouded leopard | 6 | 0.15 (0.15) | 0.04 (0.03) | - | 0.09 (0.06) | 114.5 | 0.73 |
| Leopard | 1 | 0.04 (0.04) | - | - | 0.04 (0.04) | - | - |
| Asiatic golden cat | 25 | 0.40 (0.24) | 0.43 (0.13) | - | 0.41 (0.13) | 95.5 | 0.28 |
| Jungle cat | 6 | 0.13 (0.09) | 0.11 (0.07) | - | 0.10 (0.05) | 116 | 0.85 |
| Leopard cat | 62 | 2.86 (0.87) | - | - | 2.86 (0.87) | - | - |
Table 2.
Spatial overlap (Jaccard similarity index ‘S’) and altitudinal range overlap between pairs of felid species.
Table 2.
Spatial overlap (Jaccard similarity index ‘S’) and altitudinal range overlap between pairs of felid species.
| Species 1 | Species 2 | S-Index | % Altitudinal Range Overlap | |
|---|---|---|---|---|
| Species 1 | Species 2 | |||
| Snow leopard | All others | 0 | - | - |
| Clouded leopard | Leopard | 0.333 | 7.40 | 4.00 |
| Clouded leopard | Leopard cat | 0.083 | 25.92 | 35.00 |
| Clouded leopard | Golden cat | 0.681 | 100.00 | 71.43 |
| Clouded leopard | Jungle cat | 0.733 | 100.00 | 59.73 |
| Leopard | Leopard cat | 0.100 | 30.00 | 75.00 |
| Leopard | Golden cat | 0 | 16.00 | 21.16 |
| Leopard | Jungle cat | 0 | 30.00 | 33.18 |
| Leopard cat | Golden cat | 0.111 | 65.00 | 34.39 |
| Leopard cat | Jungle cat | 0.538 | 100.00 | 44.25 |
| Golden cat | Jungle cat | 0.812 | 100.00 | 83.63 |
Table 3.
Circular statistics of temporal activity pattern, pairwise difference in activity patterns (Watson’s U2 test statistics) and temporal overlap (Pianka’s overlap index) among felid species (p is the level of significance in the test statistics).
Table 3.
Circular statistics of temporal activity pattern, pairwise difference in activity patterns (Watson’s U2 test statistics) and temporal overlap (Pianka’s overlap index) among felid species (p is the level of significance in the test statistics).
| Species | Mean Vector | S.E | 95% CI | Circular Variance |
|---|---|---|---|---|
| Snow leopard | 20:06 | 02:04 | 16:00–00:09 | 0.729 |
| Golden cat | 08:38 | 01:41 | 05:19–11:56 | 0.688 |
| Leopard cat | 00:51 | 00:30 | 23:48–01:47 | 0.359 |
| Pairwise | Watson’s U2 | p | Pianka’s index | p |
| Snow leopard–Golden cat | 0.292 | <0.01 | 0.346 | 0.618 |
| Snow leopard–Leopard cat | 0.226 | <0.05 | 0.422 | 0.352 |
| Golden cat–Leopard cat | 0.865 | <0.001 | 0.298 | 0.724 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Bashir, T.; Bhattacharya, T.; Poudyal, K.; Sathyakumar, S. Wild Felid Diversity, Space Use and Activity Patterns in the Eastern Himalaya, India. Ecologies 2023, 4, 41-54. https://doi.org/10.3390/ecologies4010005
AMA Style
Bashir T, Bhattacharya T, Poudyal K, Sathyakumar S. Wild Felid Diversity, Space Use and Activity Patterns in the Eastern Himalaya, India. Ecologies. 2023; 4(1):41-54. https://doi.org/10.3390/ecologies4010005
Chicago/Turabian StyleBashir, Tawqir, Tapajit Bhattacharya, Kamal Poudyal, and Sambandam Sathyakumar. 2023. "Wild Felid Diversity, Space Use and Activity Patterns in the Eastern Himalaya, India" Ecologies 4, no. 1: 41-54. https://doi.org/10.3390/ecologies4010005