
The Necrobiome of Deadwood: The Life after Death


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. The Necrobiome of Deadwood
2.1. Fungi
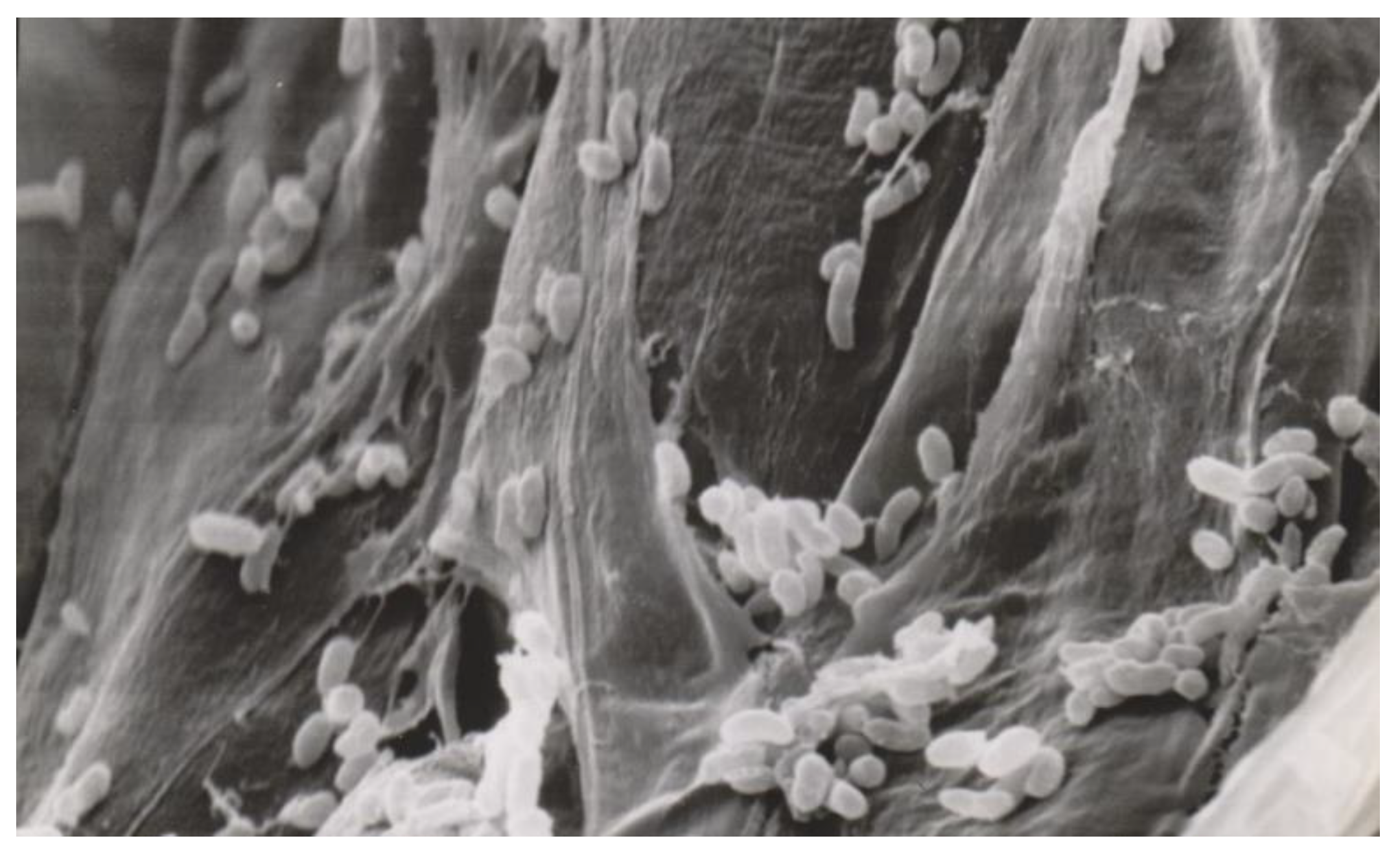
2.2. Bacteria
2.3. Archaea
2.4. Ciliate
3. Factors Affecting Decomposition
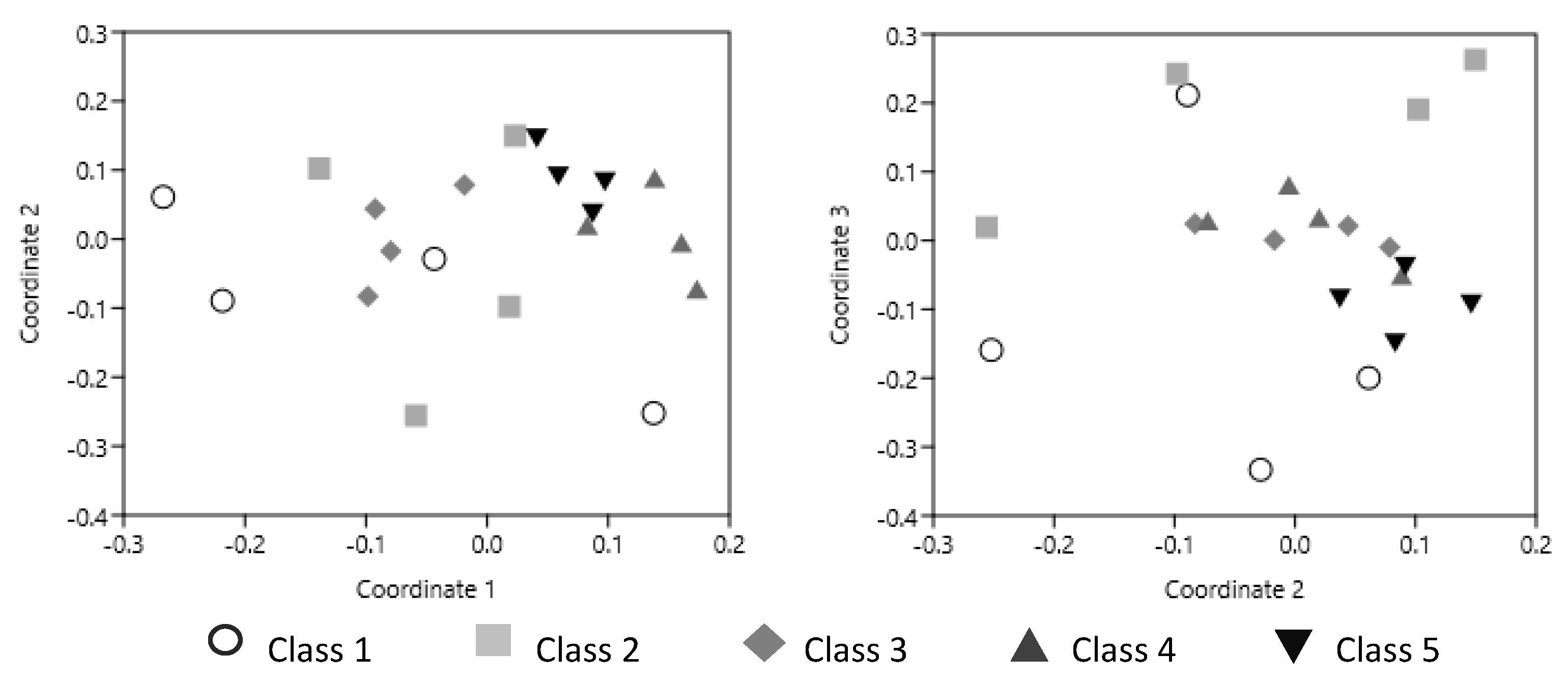
4. Interspecific Interactions within the Necrobiome
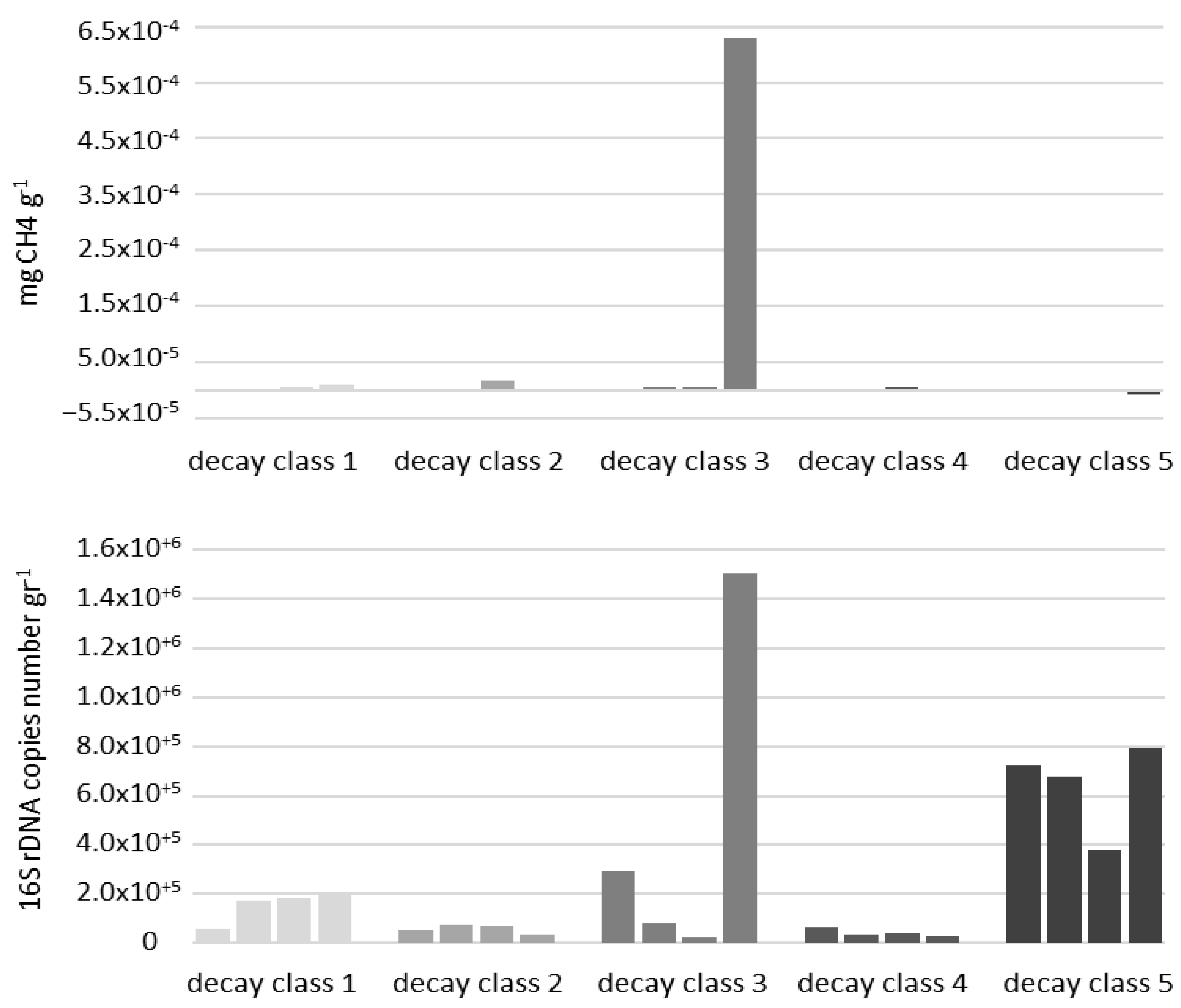
5. Role of the Necrobiome in Climate Change
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fichtner, A.; Härdtle, W. Forest Ecosystems: A Functional and Biodiversity Perspective. In Perspectives for Biodiversity and Ecosystems, Environmental Challenges and Solutions, 1st ed.; Hobohm, C., Ed.; Springer: Cham, Switzerland, 2021; Volume 16, pp. 383–405. [Google Scholar]
- Lassauce, A.; Paillet, Y.; Jactel, H.; Bouget, C. Deadwood as a surrogate for forest biodiversity: Meta-analysis of correlations between deadwood volume and species richness of saproxylic organisms. Ecol. Indic. 2011, 11, 1027–1039. [Google Scholar] [CrossRef]
- Penman, J.; Gytarsky, M.; Hiraishi, T.; Kruger, D.; Pipatti, R.; Buendia, L.; Miwa, K.; Ngara, T.; Tanabe, K.; Wagner, F. (Eds.) IPCC–Good Practice Guidance for Land Use, Land-Use Change and Forestry; Intergovernmental Panel on Climate Change (IPCC), IPCC/IGES: Hayama, Japan, 2003. [Google Scholar]
- FAO 2004. Global Forest Resources Assessment Update 2005: Terms and definitions (Working Papers 83/E); Forest Resources Assessment Programme: Rome, Italy, 2004. [Google Scholar]
- Hagemann, U.; Moroni, M.T.; Makeschin, F. Deadwood abundance in Labrador high-boreal black spruce forests. Can. J. For. Res. 2009, 39, 131–142. [Google Scholar] [CrossRef]
- La Fauci, A.; Bagnato, S.; Gugliotta, O.I.; Mercurio, R. Osservazioni preliminari sulla necromassa in popolamenti di pino laricio nel Parco Nazionale dell’Aspromonte. Forest@ 2006, 3, 54–62. [Google Scholar] [CrossRef] [Green Version]
- Camia, A.; Barbosa, P.; Amatulli, G.; San-Miguel-Ayanz, J. Fire danger rating in the European Forest Fire Information System (EFFIS): Current developments. For. Ecol. Manag. 2006, 234, S20. [Google Scholar] [CrossRef]
- Harmon, M.E.; Franklin, J.F.; Swanson, F.J.; Sollins, P.; Gregory, S.V.; Lattin, J.D.; Anderson, N.H.; Cline, S.P.; Aumen, N.G.; Sedell, J.R.; et al. Ecology of coarse woody debris in temperate ecosystems. Adv. Ecol. Res. 1986, 15, 133–302. [Google Scholar]
- Deng, Y.; Bai, Y.; Cao, R.; Jiang, Y.; Wang, Z.; Li, F.; Gong, H.; Yang, W. Key drivers of soil arthropod community shift across a subalpine forest series vary greatly with litter and topsoil layers. Eur. J. Soil Biol. 2022, 111, 103421. [Google Scholar] [CrossRef]
- Herrero, C.; Krankina, O.; Monleon, V.J.; Bravo, F. Amount and distribution of coarse woody debris in pine ecosystem of north-western Spain, Russia and the United States. IForest 2014, 7, 53–60. [Google Scholar] [CrossRef] [Green Version]
- Krankina, O.N.; Harmon, M.E. The impact of intensive forest management on carbon stores in forest ecosystems. World Res. Rev. 1994, 6, 161–177. [Google Scholar]
- Bragg, D.C.; Kershner, J.L. Coarse woody debris in riparian zones. J. For. 1999, 4, 30–35. [Google Scholar]
- Ravindranath, N.H.; Ostwald, M. Carbon Inventory Methods: Handbook for Greenhouse Gas Inventory, Carbon Mitigation and Roundwood Production Projects; Springer: Berlin/Heidelberg, Germany, 2008. [Google Scholar]
- Piaszczyk, W.; Lasota, J.; Błońska, E. Effect of organic matter released from deadwood at different decomposition stages on physical properties of forest soil. Forests 2019, 11, 24. [Google Scholar] [CrossRef] [Green Version]
- Dunker, R.; Bullock, D.; Bollero, G.; Armstrong, K. A system to evaluate prime farmland proclamation success based on spatial soil properties. In Proceedings of the American Society for Surface Mining and Reclamation, Tupelo, MS, USA, 8–15 June 2012; pp. 103–132. [Google Scholar]
- Fridman, J.; Walheim, M. Amount, structure, and dynamics of dead wood on managed forestland in Sweden. For. Ecol. Manag. 2000, 131, 23–36. [Google Scholar] [CrossRef]
- Paletto, A.; Ferretti, F.; De Meo, I.; Cantiani, P.; Focacci, M. Ecological and environmental role of deadwood in managed and unmanaged forests. In Sustainable Forest Management–Current Research; García, J.M., Diez Casero, J.J., Eds.; InTech: Reijeka, Croatia, 2012; pp. 219–238. [Google Scholar]
- Foissner, W.; Berger, H.; Xu, K.; Zechmeister-Boltenstern, S. A huge, undescribed soil ciliate (Protozoa: Ciliophora) diversity in natural forest stands of Central Europe. Biodivers. Conserv. 2005, 14, 617–701. [Google Scholar] [CrossRef]
- Jia, T.; Liang, X.; Guo, T.; Chai, B. Impact of nutrients on protozoa community diversity and structure in litter of two natural grass species in a copper tailings dam, China. Microorganisms 2021, 9, 2250. [Google Scholar] [CrossRef] [PubMed]
- Wolynski, A. Close-to-Nature Forestry in the Trentino/North Italy; Forst und Holz: Berlin, Germany, 2001. [Google Scholar]
- Benbow, M.E.; Lewis, A.J.; Tomberlin, J.K.; Pechal, J.L. Seasonal necrophagous insect community assembly during vertebrate carrion decomposition. J. Med. Entomol. 2013, 50, 440–450. [Google Scholar] [CrossRef] [PubMed]
- Benbow, M.E.; Barton, P.S.; Ulyshen, M.D.; Beasley, J.C.; DeVault, T.L.; Strickland, M.S.; Tomberlin, J.K.; Jordan, H.R.; Pechal, J.L. Necrobiome framework for bridging decomposition ecology of autotrophically and heterotrophically derived organic matter. Ecol. Monogr. 2019, 89, e01331. [Google Scholar] [CrossRef] [Green Version]
- Mali, T.; Mäki, M.; Hellen, H.; Heinonsalo, J.; Bäck, J.; Lundell, T. Decomposition of spruce wood and release of volatile organic compounds depend on decay type, fungal interactions and enzyme production patterns. FEMS Microb. Ecol. 2019, 95, fiz135. [Google Scholar] [CrossRef]
- Parisi, F.; Pioli, S.; Lombardi, F.; Fravolini, G.; Marchetti, M.; Tognetti, R. Linking deadwood traits with saproxylic invertebrates and fungi in European forests-a review. iForest 2018, 11, 423. [Google Scholar] [CrossRef] [Green Version]
- Domonell, A.; Brabender, M.; Nitsche, F.; Bonkowski, M.; Arndt, H. Community structure of cultivable protists in different grassland and forest soils of Thuringia. Pedobiologia 2013, 56, 1–7. [Google Scholar] [CrossRef]
- Ho, A.; Angel, R.; Veraart, A.J.; Daebeler, A.; Jia, Z.; Kim, S.Y.; Kerckhof, F.M.; Boon, N.; Bodelier, P.L. Biotic interactions in microbial communities as modulators of biogeochemical processes: Methanotrophy as a model system. Front. Microbiol. 2016, 7, 1285. [Google Scholar] [CrossRef] [Green Version]
- Lladó, S.; López-Mondéjar, R.; Baldrian, P. Forest soil bacteria: Diversity, involvement in ecosystem processes, and response to global change. Microbiol. Mol. Biol. Rev. 2017, 81, e00063-16. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.R.; Oberle, B.; Olivas, W.; Young, D.F.; Zanne, A.E. Wood construction more strongly shapes deadwood microbial communities than spatial location over 5 years of decay. Environ. Microbiol. 2020, 22, 4702–4717. [Google Scholar] [CrossRef] [PubMed]
- Petrillo, M.; Cherubini, P.; Sartori, G.; Abiven, S.; Ascher, J.; Bertoldi, D.; Camin, F.; Barbero, A.; Larcher, R.; Egli, M. Decomposition of Norway spruce and European larch coarse woody debris (CWD) in relation to different elevation and exposure in an Alpine setting. iForest-Biogeosci. For. 2015, 9, 154. [Google Scholar] [CrossRef] [Green Version]
- Zhou, L.; Dai, L.M.; Gu, H.Y.; Zhong, L. Review on the decomposition and influence factors of coarse woody debris in forest ecosystem. J. For. Res. 2007, 18, 48–54. [Google Scholar] [CrossRef]
- Rajala, T.; Tuomivirta, T.; Pennanen, T.; Mäkipää, R. Habitat models of wood-inhabiting fungi along a decay gradient of Norway spruce logs. Fungal Ecol. 2015, 18, 48–55. [Google Scholar] [CrossRef] [Green Version]
- Magnússon, R.Í.; Tietema, A.; Cornelissen, J.H.C.; Hefting, M.M.; Kalbitz, K. Tamm Review: Sequestration of carbon from coarse woody debris in forest soils. For. Ecol. Manag. 2016, 377, 1–15. [Google Scholar] [CrossRef]
- Purahong, W.; Wubet, T.; Lentendu, G.; Hoppe, B.; Jariyavidyanont, K.; Arnstadt, T.; Baber, K.; Otto, P.; Kellner, H.; Hofrichter, M.; et al. Determinants of deadwood-inhabiting fungal communities in temperate forests: Molecular evidence from a large scale deadwood decomposition experiment. Front. Microbiol. 2018, 9, 2120. [Google Scholar] [CrossRef]
- Pastorelli, R.; Agnelli, A.E.; De Meo, I.; Graziani, A.; Paletto, A.; Lagomarsino, A. Analysis of microbial diversity and greenhouse gas production of decaying pine logs. Forests 2017, 8, 224. [Google Scholar] [CrossRef] [Green Version]
- Pastorelli, R.; Paletto, A.; Agnelli, A.E.; Lagomarsino, A.; De Meo, I. Microbial communities associated with decomposing deadwood of downy birch in a natural forest in Khibiny Mountains (Kola Peninsula, Russian Federation). For. Ecol. Manag. 2020, 455, 117643. [Google Scholar] [CrossRef]
- Pastorelli, R.; Paletto, A.; Agnelli, A.E.; Lagomarsino, A.; De Meo, I. Microbial diversity and ecosystem functioning in deadwood of black pine of a temperate forest. Forests 2021, 12, 1418. [Google Scholar] [CrossRef]
- Strukelj, M.; Brais, S.; Quideau, S.A.; Angers, V.A.; Kebli, H.; Drapeau, P.; Oh, S.W. Chemical transformations in downed logs and snags of mixed boreal species during decomposition. Can. J. For. Res. 2013, 43, 785–798. [Google Scholar] [CrossRef]
- Moll, J.; Kellner, H.; Leonhardt, S.; Stengel, E.; Dahl, A.; Bässler, C.; Buscot, F.; Hofrichter, M.; Hoppe, B. Bacteria inhabiting deadwood of 13 tree species are heterogeneously distributed between sapwood and heartwood. Environ. Microb. 2018, 20, 3744–3756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rock, J.; Badeck, F.W.; Harmon, M.E. Estimating decomposition rate constants for European tree species from literature sources. Eur. J. For. Res. 2008, 127, 301–313. [Google Scholar] [CrossRef]
- Wei, Y.; Dai, Y. Ecological function of wood-inhabiting fungi in forest ecosystem. J. Appl. Ecol. 2004, 15, 1935–1938. [Google Scholar]
- Longa, C.M.O.; Francioli, D.; Gómez-Brandón, M.; Ascher-Jenull, J.; Bardelli, T.; Pietramellara, G.; Insam, H. Culturable fungi associated with wood decay of Picea abies in subalpine forest soils: A field-mesocosm case study. iForest 2018, 11, 781. [Google Scholar] [CrossRef]
- Gómez-Brandón, M.; Probst, M.; Siles, J.A.; Peintner, U.; Bardelli, T.; Egli, M.; Insam, H.; Ascher-Jenull, J. Fungal communities and their association with nitrogen-fixing bacteria affect early decomposition of Norway spruce deadwood. Sci. Rep. 2020, 10, 8025. [Google Scholar] [CrossRef]
- Fukasawa, Y.; Osono, T.; Takeda, H. Dynamics of physicochemical properties and occurrence of fungal fruit bodies during decomposition of coarse woody debris of Fagus crenata. J. For. Res. 2009, 14, 20–29. [Google Scholar] [CrossRef]
- Voříšková, J.; Baldrian, P. Fungal community on decomposing leaf litter undergoes rapid successional changes. ISME J. 2013, 7, 477–486. [Google Scholar] [CrossRef]
- Fukami, T.; Dickie, I.A.; Paula Wilkie, J.; Paulus, B.C.; Park, D.; Roberts, A.; Buchanan, P.K.; Allen, R.B. Assembly history dictates ecosystem functioning: Evidence from wood decomposer communities. Ecol. Lett. 2010, 13, 675–684. [Google Scholar] [CrossRef]
- Yang, S.; Limpens, J.; Sterck, F.J.; Sass-Klaassen, U.; Cornelissen, J.H.; Hefting, M.; van Logtestijn, R.S.P.; Goudzwaard, L.; Dam, N.; Dam, M.; et al. Dead wood diversity promotes fungal diversity. Oikos 2021, 130, 2202–2216. [Google Scholar] [CrossRef]
- Paliwal, R.; Giri, K.; Rai, J.P.N. Microbial ligninolysis: Avenue for natural ecosystem management. In Biotechnology: Concepts, Methodologies, Tools, and Applications; IGI Global: Hershey, PA, USA, 2019; pp. 1399–1423. [Google Scholar]
- Berg, B.; McClaugherty, C. Decomposer organisms. In Plant Litter; Springer: Cham, Switzerland, 2020; pp. 45–65. [Google Scholar]
- Savory, J.G. Breakdown of timber by ascomycetes and fungi imperfecti. Ann. Appl. Biol. 1954, 41, 336–347. [Google Scholar] [CrossRef]
- Daniel, G.; Nilsson, T. Developments in the study of soft rot and bacterial decay. In Forest Products Biotechnology, 1st ed.; Bruce, A., Palfreyman, J.W., Eds.; Taylor and Francis: London, UK, 1997; pp. 47–72. [Google Scholar]
- Ward, G.; Hadar, Y.; Dosoretz, C.G. The biodegradation of lignocellulose by white rot fungi. Mycol. Ser. 2004, 21, 393–408. [Google Scholar]
- Hatakka, A.; Hammel, K.E. Fungal biodegradation of lignocelluloses. In Industrial applications. The Mycota, 1st ed.; Hofrichter, M., Ed.; Springer: Berlin/Heidelberg, Germany, 2011; Volume 10, pp. 319–340. [Google Scholar]
- Allen, R.B.; Buchanan, P.K.; Clinton, P.W.; Cone, A.J. Composition and diversity of fungi on decaying logs in a New Zealand temperate beech (Nothofagus) forest. Can. J. For. Res. 2000, 30, 10251033. [Google Scholar] [CrossRef]
- Kielak, A.M.; Scheublin, T.R.; Mendes, L.W.; Van Veen, J.A.; Kuramae, E.E. Bacterial community succession in pine-wood decomposition. Front. Microbiol. 2016, 7, 231. [Google Scholar] [CrossRef] [Green Version]
- Schwarze, F.W.M.R.; Engels, J.; Mattheck, C. Fungal Strategies of Wood Decay in Trees; Springer: Berlin/Hedelberg, Germany; New York, NY, USA,, 2000. [Google Scholar]
- Blanchette, R.A. A review of microbial deterioration found in archaeological wood from different environments. Int. Biodeterior. Biodegrad. 2000, 46, 189–204. [Google Scholar] [CrossRef]
- Fukasawa, Y.; Osono, T.; Takeda, H. Wood decomposing abilities of diverse lignicolous fungi on nondecayed and decayed beech wood. Mycologia 2011, 3, 474–482. [Google Scholar] [CrossRef]
- Blaser, S.; Prati, D.; Senn-Irlet, B.; Fischer, M. Effects of forest management on the diversity of deadwood-inhabiting fungi in Central European forests. For. Ecol. Manag. 2013, 304, 42–48. [Google Scholar] [CrossRef]
- Parfitt, D.; Hunt, J.; Dockrell, D.; Rogers, H.J.; Boddy, L. Do all trees carry the seeds of their own destruction? PCR reveals numerous wood decay fungi latently present in sapwood of a wide range of angiosperm trees. Fungal Ecol. 2010, 3, 338–346. [Google Scholar] [CrossRef]
- Prewitt, L.; Kang, Y.; Kakumanu, M.L.; Williams, M. Fungal and bacterial community succession differs for three wood types during decay in a forest soil. Microb. Ecol. 2014, 68, 212–221. [Google Scholar] [CrossRef] [PubMed]
- Kubartová, A.; Ottosson, E.; Dahlberg, A.; Stenlid, J. Patterns of fungal communities among and within decaying logs, revealed by 454 sequencing. Mol. Ecol. 2012, 21, 4514–4532. [Google Scholar]
- Ovaskainen, O.; Schigel, D.; Ali-Kovero, H.; Auvinen, P.; Paulin, L.; Nordén, B.; Nordén, J. Combining high-throughput sequencing with fruit body surveys reveals contrasting life-history strategies in fungi. ISME J. 2013, 7, 1696–1709. [Google Scholar] [CrossRef]
- Hoppe, B.; Purahong, W.; Wubet, T.; Kahl, T.; Bauhus, J.; Arnstadt, T.; Hofrichter, M.; Buscot, F.; Krüger, D. Linking molecular deadwood-inhabiting fungal diversity and community dynamics to ecosystem functions and processes in Central European forests. Fungal Divers. 2016, 77, 367–379. [Google Scholar] [CrossRef]
- Purahong, W.; Pietsch, K.A.; Lentendu, G.; Schöps, R.; Bruelheide, H.; Wirth, C.; Buscot, F.; Wubet, T. Characterization of unexplored deadwood mycobiome in highly diverse subtropical forests using culture-independent molecular technique. Front. Microbiol. 2017, 8, 574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tláskal, V.; Brabcová, V.; Větrovský, T.; Jomura, M.; López-Mondéjar, R.; Oliveira Monteiro, L.M.; Saraiva, J.P.; Human, Z.R.; Cajthaml, T.; da Rocha, U.N.; et al. Complementary roles of wood-inhabiting fungi and bacteria facilitate deadwood decomposition. mSystems 2021, 6, e01078-20. [Google Scholar] [CrossRef]
- Probst, M.; Gómez-Brandón, M.; Bardelli, T.; Egli, M.; Insam, H.; Ascher-Jenull, J. Bacterial communities of decaying Norway spruce follow distinct slope exposure and time-dependent trajectories. Environ. Microbiol. 2018, 20, 3657–3670. [Google Scholar] [CrossRef]
- Díaz-García, L.; Bugg, T.D.; Jiménez, D.J. Exploring the lignin catabolism potential of soil-derived lignocellulolytic microbial consortia by a gene-centric metagenomic approach. Microb. Ecol. 2020, 80, 885–896. [Google Scholar] [CrossRef]
- Greaves, H. The bacterial factor in wood decay. Wood Sci. Technol. 1971, 5, 6–16. [Google Scholar] [CrossRef]
- Clausen, C.A. Bacterial associations with decaying wood: A review. Int. Biodeterior. Biodegrad. 1996, 37, 101–107. [Google Scholar] [CrossRef]
- Singh, A.P.; Kim, Y.S.; Singh, T. Chapter 9-Bacterial Degradation of Wood. In Secondary Xylem Biology-Origins, Functions, and Applications; Kim, Y.S., Funada, R., Singh, A.P., Eds.; Academic Press: Cambridge, MA, USA, 2016; pp. 169–190. [Google Scholar]
- Hagge, J.; Bässler, C.; Gruppe, A.; Hoppe, B.; Kellner, H.; Krah, F.S.; Müller, J.; Seibold, S.; Stengel, E.; Thorn, S. Bark coverage shifts assembly processes of microbial decomposer communities in deadwood. Proc. Royal Soc. B 2019, 286, 20191744. [Google Scholar] [CrossRef] [Green Version]
- De Boer, W.; Folman, L.B.; Summerbell, R.C.; Boddy, L. Living in a fungal world: Impact of fungi on soil bacterial niche development. FEMS Microbiol. Rev. 2005, 29, 795–811. [Google Scholar] [CrossRef] [Green Version]
- Folman, L.B.; Klein Gunnewiek, P.J.; Boddy, L.; De Boer, W. Impact of white-rot fungi on numbers and community composition of bacteria colonizing beech wood from forest soil. FEMS Microbiol. Ecol. 2008, 63, 181–191. [Google Scholar] [CrossRef] [Green Version]
- Sun, H.; Terhonen, E.; Kasanen, R.; Asiegbu, F.O. Diversity and community structure of primary wood-inhabiting bacteria in boreal forest. Geomicrobiol. J. 2014, 31, 315–324. [Google Scholar] [CrossRef]
- Větrovský, T.; Steffen, K.T.; Baldrian, P. Potential of cometabolic transformation of polysaccharides and lignin in lignocellulose by soil Actinobacteria. PLoS ONE 2014, 9, e89108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lynd, L.R.; Weimer, P.J.; van Zyl, W.H.; Pretorius, I.S. Microbial cellulose utilization: Fundamentals and biotechnology, Microbiol. Mol. Biol. Rev. 2020, 66, 506–577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.B.; Yang, M.X.; Tu, R. Unexpectedly high bacterial diversity in decaying wood of a conifer as revealed by a molecular method. Int. Biodeterior. Biodegrad. 2008, 62, 471–474. [Google Scholar] [CrossRef]
- Janusz, G.; Pawlik, A.; Sulej, J.; Świderska-Burek, U.; Jarosz-Wilkołazka, A.; Paszczyński, A. Lignin degradation: Microorganisms, enzymes involved, genomes analysis and evolution. FEMS Microbiol. Rev. 2017, 41, 941–962. [Google Scholar] [CrossRef] [Green Version]
- Kleinsteuber, S.; Müller, F.D.; Chatzinotas, A.; Wendt-Potthoff, K.; Harms, H. Diversity and in situ quantification of Acidobacteria subdivision 1 in an acidic mining lake. FEMS Microbiol. Ecol. 2008, 63, 107–117. [Google Scholar] [CrossRef]
- Radajewski, S.; Webster, G.; Reay, D.S.; Morris, S.A.; Ineson, P.; Nedwell, D.B.; Prosser, J.I.; Murrell, J.C. Identification of active methylotroph populations in an acidic forest soil by stable isotope probing. Microbiology 2002, 148, 2331–2342. [Google Scholar] [CrossRef] [Green Version]
- Vorob’ev, A.V.; de Boer, W.; Folman, L.B.; Bodelier, P.L.; Doronina, N.V.; Suzina, N.E.; Trotsenko, Y.A.; Dedysh, S.N. Methylovirgula ligni gen. nov., sp. nov., an obligately acidophilic, facultatively methylotrophic bacterium with a highly divergent mxaF gene. Int. J. System Evol. Microbiol. 2009, 59, 2538–2545. [Google Scholar] [CrossRef] [Green Version]
- Mäkipää, R.; Leppänen, S.M.; Munoz, S.S.; Smolander, A.; Tiirola, M.; Tuomivirta, T.; Fritze, H. Methanotrophs are core members of the diazotroph community in decaying Norway spruce logs. Soil Biol. Biochem. 2018, 120, 230–232. [Google Scholar] [CrossRef]
- Dedysh, S.N.; Khmelenina, V.N.; Suzina, N.E.; Trotsenko, Y.A.; Semrau, J.D.; Liesack, W.; Tiedje, J.M. Methylocapsa acidiphila gen. nov., sp. nov., a novel methane-oxidizing and dinitrogen-fixing acidophilic bacterium from Sphagnum bog. Int. J. Syst. Evol. Microbiol. 2002, 52, 251–261. [Google Scholar] [CrossRef] [Green Version]
- Hoppe, B.; Kahl, T.; Karasch, P.; Wubet, T.; Bauhus, J.; Buscot, F.; Krüger, D. Network analysis reveals ecological links between N-fixing bacteria and wood-decaying fungi. PLoS ONE 2014, 9, E88141. [Google Scholar] [CrossRef] [Green Version]
- Zumft, W.G. Cell biology and molecular basis of denitrification. Microbiol. Mol. Biol. Rev. 1997, 61, 533–616. [Google Scholar] [PubMed]
- Shaw, L.J.; Nicol, G.W.; Smith, Z.; Fear, J.; Prosser, J.I.; Baggs, E.M. Nitrosospira spp. can produce nitrous oxide via a nitrifier denitrification pathway. Environ. Microbiol. 2006, 8, 214–222. [Google Scholar] [CrossRef] [PubMed]
- Wainø, M.; Ingvorsen, K. Production of β-xylanase and β-xylosidase by the extremely halophilic archaeon Halorhabdus utahensis. Extremophiles 2003, 7, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Rinta-Kanto, J.M.; Sinkko, H.; Rajala, T.; Al-Soud, W.A.; Sørensen, S.J.; Tamminen, M.V.; Timonen, S. Natural decay process affects the abundance and community structure of Bacteria and Archaea in Picea abies logs. FEMS Microbiol. Ecol. 2016, 92, 087. [Google Scholar] [CrossRef] [Green Version]
- Topp, E.; Pattey, E. Soils as sources and sinks for atmospheric methane. Can. J. Soil Sci. 1997, 77, 167–177. [Google Scholar] [CrossRef]
- Zeikus, J.G.; Henning, D.L. Methanobacterium arbophilicum sp. nov. An obligate anaerobe isolated from wetwood if living trees. Anton Leeuw. 1975, 41, 543–552. [Google Scholar] [CrossRef]
- Covey, K.R.; de Mesquita, C.P.; Oberle, B.; Maynard, D.S.; Bettigole, C.; Crowther, T.W.; Duguid, M.C.; Steven, B.; Zanne, A.E.; Lapin, M.; et al. Greenhouse trace gases in deadwood. Biogeochemistry 2016, 130, 215–226. [Google Scholar] [CrossRef]
- Mukhin, V.A.; Voronin, P.Y. A new source of methane in boreal forests. Appl. Biochem. Microbiol. 2008, 44, 297–299. [Google Scholar] [CrossRef]
- Kudo, T. Termite-microbe symbiotic system and its efficient degradation of lignocellulose. Biosci. Biotechnol. Biochem. 2009, 73, 2561–2567. [Google Scholar] [CrossRef]
- Hongoh, Y.; Ohkuma, M. Termite gut flagellates and their methanogenic and eubacterial symbionts. In Endosymbiotic Methanogenic Archaea; Hackstein, J.H.P., Ed.; Springer: Heidelberg, Germany, 2010; Volume 19, pp. 55–79. [Google Scholar]
- Foissner, W. An updated compilation of world soil ciliates (Protozoa, Ciliophora), with ecological notes, new records, and descriptions of new species. Eur. J. Protistol. 1998, 34, 195–235. [Google Scholar] [CrossRef]
- Adl, M.S.; Gupta, V.S. Protists in soil ecology and forest nutrient cycling. CA J. For. Res. 2006, 36, 1805–1817. [Google Scholar] [CrossRef]
- Rivera, F.; Lugo, A.; Ramirez, E.; Bonilla, P.; Calderon, A.; Rodriguez, S.; Ortiz, R.; Gallegos, E.; Labastida, A.; Chavez, M.P. Seasonal distribution of air-borne protozoa in Mexico City and its suburbs. Water Air Soil Poll. 1992, 61, 17–36. [Google Scholar] [CrossRef]
- Pastorelli, R.; Cucu, M.A.; Lagomarsino, A.; Paletto, A.; De Meo, I. Analysis of ciliate community diversity in decaying Pinus nigra logs. Forests 2022, 13, 642. [Google Scholar] [CrossRef]
- Chesson, P. Mechanisms of maintenance of species diversity. Annu. Rev. Ecol. Syst. 2000, 31, 343–366. [Google Scholar] [CrossRef] [Green Version]
- Bonkowski, M. Protozoa and plant growth: The microbial loop in soil revisited. New Phytol. 2004, 162, 617–631. [Google Scholar] [CrossRef]
- Adl, S.M.; Coleman, D.C. Dynamics of soil protozoa using a direct count method. Biol. Fertil. Soils 2005, 42, 168–171. [Google Scholar] [CrossRef]
- Bartošová, P.; Tirjaková, E. Diversity and ecology of ciliates (Alveolata: Ciliophora) living in the bark and decaying wood mass in Slovakia. Acta Protozool. 2008, 47, 173–187. [Google Scholar]
- Lara, E.; Berney, C.; Harms, H.; Chatzinotas, A. Cultivation-independent analysis reveals a shift in ciliate 18S rRNA gene diversity in a polycyclic aromatic hydrocarbon-polluted soil. FEMS Microbiol. Ecol. 2007, 62, 365–373. [Google Scholar] [CrossRef] [Green Version]
- Jousset, A.; Lara, E.; Nikolausz, M.; Harms, H.; Chatzinotas, A. Application of the denaturing gradient gel electrophoresis (DGGE) technique as an efficient diagnostic tool for ciliate communities in soil. Sci. Total Environ. 2010, 408, 1221–1225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gora, E.M.; Lucas, J.M.; Yanoviak, S.P. Microbial composition and wood decomposition rates vary with microclimate from the ground to the canopy in a tropical forest. Ecosystems 2019, 22, 1206–1219. [Google Scholar] [CrossRef]
- Jaroszewicz, B.; Cholewińska, O.; Chećko, E.; Wrzosek, M. Predictors of diversity of deadwood-dwelling macrofungi in a European natural forest. For. Ecol. Manag. 2021, 490, 119123. [Google Scholar] [CrossRef]
- Bani, A.; Pioli, S.; Ventura, M.; Panzacchi, P.; Borruso, L.; Tognetti, R.; Tonon, G.; Brusetti, L. The role of microbial community in the decomposition of leaf litter and deadwood. Appl. Soil Ecol. 2018, 126, 75–84. [Google Scholar] [CrossRef]
- Baldrian, P.; Zrůstová, P.; Tláskal, V.; Davidová, A.; Merhautová, V.; Vrška, T. Fungi associated with decomposing deadwood in a natural beech-dominated forest. Fungal Ecol. 2016, 23, 109–122. [Google Scholar] [CrossRef]
- Arnstadt, T.; Hoppe, B.; Kahl, T.; Kellner, H.; Krüger, D.; Bauhus, J.; Hofrichter, M. Dynamics of fungal community composition, decomposition and resulting deadwood properties in logs of Fagus sylvatica, Picea abies and Pinus sylvestris. For. Ecol. Manag. 2016, 382, 129–142. [Google Scholar] [CrossRef]
- Mäkinen, H.; Hynynen, J.; Siitonen, J.; Sievänen, R. Predicting the decomposition of Scots pine, Norway spruce, and birch stems in Finland. Ecol. Appl. 2006, 16, 1865–1879. [Google Scholar] [CrossRef]
- Russell, M.B.; Fraver, S.; Aakala, T.; Gove, J.H.; Woodall, C.W.; D’amato, A.W.; Ducey, M.J. Quantifying carbon stores and decomposition in deadwood: A review. For. Ecol. Manag. 2015, 350, 107–128. [Google Scholar] [CrossRef]
- Rinne, K.T.; Rajala, T.; Peltoniemi, K.; Chen, J.; Smolander, A.; Mäkipää, R. Accumulation rates and sources of external nitrogen in decaying wood in a Norway spruce dominated forest. Funct. Ecol. 2017, 31, 530–541. [Google Scholar] [CrossRef]
- Palviainen, M.; Finér, L.; Laiho, R.; Shorohova, E.; Kapitsa, E.; Vanha-Majamaa, I. Carbon and nitrogen release from decomposing Scots pine, Norway spruce and silver birch stumps. For. Ecol. Manag. 2010, 259, 390–398. [Google Scholar] [CrossRef]
- Laiho, R.; Prescott, C.E. Decay and nutrient dynamics of coarse woody debris in northern coniferous forests: A synthesis. Can. J. For. Res. 2004, 34, 763–777. [Google Scholar] [CrossRef]
- Birkemoe, T.; Jacobsen, R.M.; Sverdrup-Thygeson, A.; Biedermann, P.H. Insect-fungus interactions in dead wood systems. In Saproxylic Insects; Springer: Cham, Switzerland, 2018; pp. 377–427. [Google Scholar]
- Johnston, S.R.; Boddy, L.; Weightman, A.J. Bacteria in decomposing wood and their interactions with wood-decay fungi. FEMS Microbiol. Ecol. 2016, 92, 179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valášková, V.; de Boer, W.; Klein Gunnewiek, P.J.A.; Pospíšek, M.; Baldrian, P. Phylogenetic composition and properties of bacteria coexisting with the fungus Hypholoma fasciculare in decaying wood. ISME J. 2009, 3, 1218–1221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frey-Klett, P.; Burlinson, P.; Deveau, A.; Barret, M.; Tarkka, M.; Sarniguet, A. Bacterial-fungal interactions: Hyphens between agricultural, clinical, environmental, and food microbiologists. Microbiol. Mol. Biol. Rev. 2011, 75, 583–609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azam, F.; Fenchel, T.; Field, J.G.; Gray, J.S.; Meyer-Reil, L.A.; Thingstad, F. The ecological role of water-column microbes in the sea. Mar. Ecol. Prog. Ser. 1983, 10, 257–263. [Google Scholar] [CrossRef]
- Geisen, S.; Mitchell, E.A.; Adl, S.; Bonkowski, M.; Dunthorn, M.; Ekelund, F.; Fernández, L.D.; Jousset, A.; Krashevska, V.; Singer, D.; et al. Soil protists: A fertile frontier in soil biology research. FEMS Microbiol. Rev. 2018, 42, 293–323. [Google Scholar] [CrossRef] [PubMed]
- Weedon, J.T.; Cornwell, W.K.; Cornelissen, J.H.; Zanne, A.E.; Wirth, C.; Coomes, D. Global meta-analysis of wood decomposition rates: A role for trait variation among tree species? Ecol. Lett. 2009, 12, 45–56. [Google Scholar] [CrossRef] [PubMed]
- Gough, C.M.; Vogel, C.S.; Kazanski, C.; Nagel, L.; Flower, C.E.; Curtis, P.S. Coarse woody debris and the carbon balance of a north temperate forest. For. Ecol. Manag. 2007, 244, 60–67. [Google Scholar] [CrossRef]
- Treseder, K.K.; Bent, E.; Borneman, J.; McGuire, K.L. Shifts in fungal communities during decomposition of boreal forest litter. Fungal Ecol. 2014, 10, 58–69. [Google Scholar] [CrossRef]
- Kuzyakov, Y. Prime time for microbes. Nat. Clim. Chang. 2011, 1, 295–297. [Google Scholar] [CrossRef]
- Baldrian, P.; Šnajdr, J.; Merhautová, V.; Dobiášová, P.; Cajthaml, T.; Valášková, V. Responses of the extracellular enzyme activities in hardwood forest to soil temperature and seasonality and the potential effects of climate change. Soil Biol. Biochem. 2013, 56, 60–68. [Google Scholar] [CrossRef]
- Bebber, D.P.; Watkinson, S.C.; Boddy, L.; Darrah, P.R. Simulated nitrogen deposition affects wood decomposition by cord-forming fungi. Oecologia 2011, 167, 1177–1184. [Google Scholar] [CrossRef] [PubMed]
- Waldrop, M.P.; Zak, D.R.; Sinsabaugh, R.L.; Gallo, M.; Lauber, C. Nitrogen deposition modifies soil carbon storage through changes in microbial enzymatic activity. Ecol. Appl. 2004, 14, 1172–1177. [Google Scholar] [CrossRef]
- Covey, K.R.; Wood, S.A.; Warren, R.J.; Lee, X.; Bradford, M.A. Elevated methane concentrations in trees of an upland forest. Geophys. Res. Lett. 2012, 39, L15705. [Google Scholar] [CrossRef] [Green Version]
- Lenhart, K.; Bunge, M.; Ratering, S.; Neu, T.R.; Schüttmann, I.; Greule, M.; Kammann, C.; Schnell, S.; Müller, C.; Zorn, Z.; et al. Evidence for methane production by saprotrophic fungi. Nat. Commun. 2012, 3, 1046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, B.K.; Bardgett, R.D.; Smith, P.; Reay, D.S. Microorganisms and climate change: Terrestrial feedbacks and mitigation options. Nat. Rev. Microbiol. 2010, 8, 779–790. [Google Scholar] [CrossRef] [PubMed]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pastorelli, R.; De Meo, I.; Lagomarsino, A. The Necrobiome of Deadwood: The Life after Death. Ecologies 2023, 4, 20-38. https://doi.org/10.3390/ecologies4010003
Pastorelli R, De Meo I, Lagomarsino A. The Necrobiome of Deadwood: The Life after Death. Ecologies. 2023; 4(1):20-38. https://doi.org/10.3390/ecologies4010003
Chicago/Turabian StylePastorelli, Roberta, Isabella De Meo, and Alessandra Lagomarsino. 2023. "The Necrobiome of Deadwood: The Life after Death" Ecologies 4, no. 1: 20-38. https://doi.org/10.3390/ecologies4010003