
Pollinator Communities of Planted and Feral Pyrus calleryana

 , , ,
, , ,
Abstract
:1. Introduction
2. Methods
2.1. Study Area
2.2. Tree Identification, Selection, and Bloom Period
2.3. Pollinator Sample Collection
2.4. Expected Pollinator Community Data Acquisition
2.5. Data Analysis
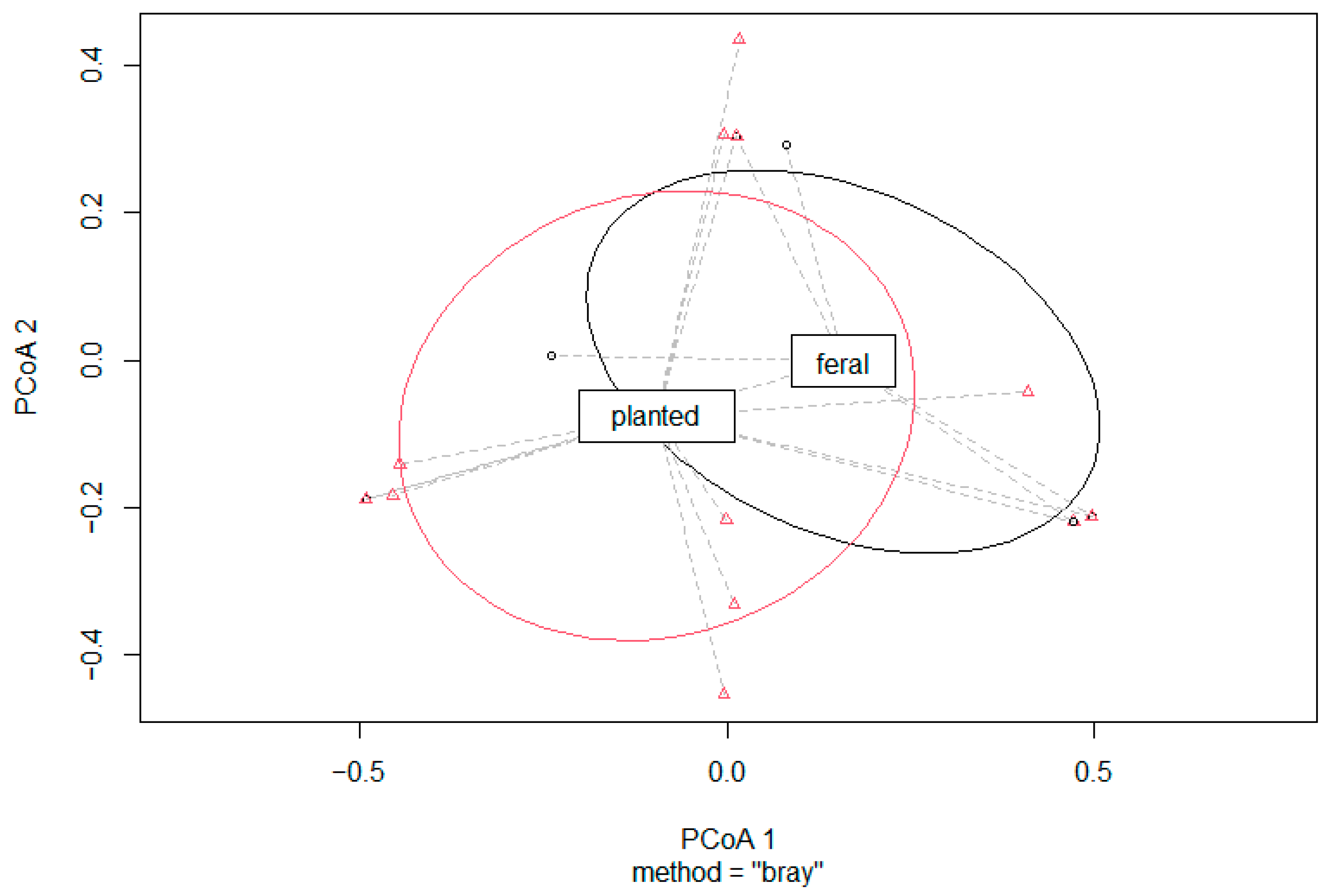
3. Results
3.1. Observed Pollinator Community
3.2. Observed and Expected Community Comparisons
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- IUCN. Guidelines for the Prevention of Biodiversity Loss Caused by Alien Invasive Species Prepared by the SSC Invasive Species Specialist Group. (International Union of Conservation of Nature Report 2000-052). In Proceedings of the 51st Meeting of the IUCN Council, Gland, Switzerland, 9 February 2000. [Google Scholar]
- Vilà, M.; Espinar, J.L.; Hejda, M.; Hulme, P.E.; Jarošík, V.; Maron, J.L.; Pergl, J.; Schaffner, U.; Sun, Y.; Pyšek, P. Ecological Impacts of Invasive Alien Plants: A Meta-Analysis of Their Effects on Species, Communities and Ecosystems. Ecol. Lett. 2011, 14, 702–708. [Google Scholar] [CrossRef] [PubMed]
- Lehan, N.E.; Murphy, J.R.; Thorburn, L.P.; Bradley, B.A. Accidental Introductions Are an Important Source of Invasive Plants in the Continental United States. Am. J. Bot. 2013, 100, 1287–1293. [Google Scholar] [CrossRef] [PubMed]
- Pimentel, D.; Lach, L.; Zuniga, R.; Morrison, D. Environmental and Economic Costs of Nonindigenous Species in the United States. Bioscience 2000, 50, 53–65. [Google Scholar] [CrossRef] [Green Version]
- Culley, T.M.; Hardiman, N.A. The Beginning of a New Invasive Plant: A History of the Ornamental Callery Pear in the United States. Bioscience 2007, 57, 956–964. [Google Scholar] [CrossRef] [Green Version]
- Hartshorn, J.A.; Palmer, J.F.; Coyle, D.R. Into the Wild: Evidence for the Enemy Release Hypothesis in the Invasive Callery Pear (Pyrus calleryana) (Rosales: Rosaceae). Environ. Entomol. 2022, 51, 216–221. [Google Scholar] [CrossRef] [PubMed]
- Woods, M.J.; Attea, G.K.; McEwan, R.W. Resprouting of the Woody Plant Pyrus calleryana Influences Soil Ecology during Invasion of Grasslands in the American Midwest. Appl. Soil Ecol. 2021, 166, 103989. [Google Scholar] [CrossRef]
- Culley, T. The Rise and Fall of the Ornamental Callery Pear Tree. Arnoldia 2017, 74, 2–11. [Google Scholar]
- Clark, O. A Starling in a Pear Tree: Assessing the Influence of Bird Dispersal on Callery Pear (Pyrus calleryana). Undergraduate Honors Thesis 347, University of Dayton, Dayton, OH, USA, 2022. [Google Scholar]
- Swearingen, C.; Slattery, B.; Reshetiloff, K.; Zwicker, S. Plant Invaders of Mid-Atlantic Natural Areas, 4th ed.; National Park Service and U.S. Fish and Wildlife Service: Washington, DC, USA, 2010; p. 168. [Google Scholar]
- Gilman, E.F.; Watson, D.G.; Klein, R.W.; Koeser, A.K.; Hilbert, D.R.; McLean, D.C. Pyrus Calleryana: “Bradford” Callery Pear; Extension Publication ENH-695; Southern Trees Facts Sheets; University of Florida: Gainesville, FL, USA, 2019. [Google Scholar]
- Serota, T.H.; Culley, T.M. Seed Germination and Seedling Survival of Invasive Callery Pear (Pyrus calleryana Decne.) 11 Years After Fruit Collection. Castanea 2019, 84, 47–52. [Google Scholar] [CrossRef]
- Ferrer, M.M.; Good, S.v. Self-Sterility in Flowering Plants: Preventing Self-Fertilization Increases Family Diversification Rates. Ann. Bot. 2012, 110, 535. [Google Scholar] [CrossRef] [Green Version]
- Fountain, M.T.; Mateos-Fierro, Z.; Shaw, B.; Brain, P.; Delgado, A. Insect Pollinators of Conference Pear (Pyrus communis L.) and Their Contribution to Fruit Quality. J. Pollinat. Ecol 2019, 25, 103–114. [Google Scholar] [CrossRef]
- Quinet, M.; Warzée, M.; Vanderplanck, M.; Michez, D.; Lognay, G.; Jacquemart, A.L. Do Floral Resources Influence Pollination Rates and Subsequent Fruit Set in Pear (Pyrus communis L.) and Apple (Malus × Domestica Borkh) Cultivars? Eur. J. Agron. 2016, 77, 59–69. [Google Scholar] [CrossRef]
- Kaluza, B.F.; Wallace, H.; Keller, A.; Heard, T.A.; Jeffers, B.; Drescher, N.; Blüthgen, N.; Leonhardt, S.D. Generalist Social Bees Maximize Diversity Intake in Plant Species-Rich and Resource-Abundant Environments. Ecosphere 2017, 8, e01758. [Google Scholar] [CrossRef] [Green Version]
- Davis, E.S.; Kelly, R.; Maggs, C.A.; Stout, J.C. Contrasting Impacts of Highly Invasive Plant Species on Flower-Visiting Insect Communities. Biodivers Conserv. 2018, 27, 2069–2085. [Google Scholar] [CrossRef]
- Bartomeus, I.; Vilà, M.; Santamaría, L. Contrasting Effects of Invasive Plants in Plant-Pollinator Networks. Oecologia 2008, 155, 761–770. [Google Scholar] [CrossRef] [PubMed]
- Burkle, L.A.; Marlin, J.C.; Knight, T.M. Plant-Pollinator Interactions over 120 Years: Loss of Species, Co-Occurrence, and Function. Science 2013, 340, 1611–1615. [Google Scholar] [CrossRef] [Green Version]
- Tepedino, V.J.; Bradley, B.A.; Griswold, T.L. Might Flowers of Invasive Plants Increase Native Bee Carrying Capacity? Intimations from Capitol Reef National Park, Utah. Nat. Areas J. 2008, 28, 44–50. [Google Scholar] [CrossRef]
- Stout, J.C.; Tiedeken, E.J. Direct Interactions between Invasive Plants and Native Pollinators: Evidence, Impacts and Approaches. Funct. Ecol. 2017, 31, 38–46. [Google Scholar] [CrossRef] [Green Version]
- Giaquinto, A. Biodiversity, Ecosystem Function and the Pollination Ecology of Urban Gardens in Dayton, Ohio. Undergraduate. Honors Thesis, University of Dayton, Dayton, OH, USA, 2016. [Google Scholar]
- Tucker, E.M.; Rehan, S.M. Farming for Bees: Annual Variation in Pollinator Populations across Agricultural Landscapes. Agric. For. Entomol. 2018, 20, 541–548. [Google Scholar] [CrossRef]
- Hernandez, J.; Frankie, G.; Thorp, R. Ecology of Urban Bees: A Review of Current Knowledge and Directions for Future Study. Cities Environ. (CATE) 2009, 2, 3. [Google Scholar]
- Eaton, E.R. Common Bees & Wasps of Ohio Field Guide, 1st ed.; Ohio DNR Division of Wildlife: Columbus, OH, USA, 2016. [Google Scholar]
- North American Native Bee Collaborative. Bees of Ohio: A Field Guide; Schnebelin, A., Spring, M., Ellsworth, D., Eds.; North American Native Bee Collaborative: Washington, DC, USA, 2020. [Google Scholar]
- Omernik, J.M.; Griffith, G.E. Ecoregions of the Conterminous United States: Evolution of a Hierarchical Spatial Framework. Environ. Manag. 2014, 54, 1249–1266. [Google Scholar] [CrossRef]
- Köppen, W.; Volken, E.; Brönnimann, S. The Thermal Zones of the Earth According to the Duration of Hot, Moderate and Cold Periods and to the Impact of Heat on the Organic World. Meteorol. Z. 2011, 20, 351–360. [Google Scholar] [CrossRef]
- Soil Survey Staff. Web Soil Survey. Available online: http://websoilsurvey.sc.egov.usda.gov/ (accessed on 4 October 2022).
- U.S. Census Bureau QuickFacts. Available online: https://www.census.gov/quickfacts/fact/table/ashlandcityohio,findlaycityohio/PST045221 (accessed on 27 November 2022).
- Ellsworth, D. Ohio State Phenology Calendar. Available online: https://weather.cfaes.osu.edu/gdd/default.asp (accessed on 4 October 2022).
- NCEI. U.S. Climate Normals. Available online: https://www.ncei.noaa.gov/products/land-based-station/us-climate-normals (accessed on 4 October 2022).
- iNaturalist Contributors. iNaturalist Research-Grade Observations. Available online: https://www.inaturalist.org (accessed on 1 October 2022).
- Barve, V.; Hart, E. Rinat: Access “iNaturalist” Data Through APIs. R Package Version 0.1.8. Available online: https://CRAN.R-project.org/package=rinat (accessed on 1 October 2021).
- R Core Team R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing: Vienna, Austria. Available online: https://www.R-project.org/ (accessed on 1 October 2021).
- RStudio Team. RStudio: Integrated Development Environment for R, version 2021.09.2. Available online: https://www.rstudio.com/ (accessed on 1 October 2021).
- Simpson, E.H. Measurement of Diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Anderson, M.J. A New Method for Non-Parametric Multivariate Analysis of Variance. Austral. Ecol. 2001, 26, 32–46. [Google Scholar] [CrossRef]
- Dufrene, M.; Legendre, P. Species Assemblages and Indicator Species: The Need for a Flexible Asymmetrical Approach. Ecol. Monogr. 1997, 67, 345–366. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. R Package Version 2.5-7. Available online: https://CRAN.R-project.org/package=vegan (accessed on 1 October 2021).
- Roberts, D.W. labdsv: Ordination and Multivariate Analysis for Ecology. R Package Version 2.0-1. Available online: https://CRAN.R-project.org/package=labdsv (accessed on 1 October 2021).
- Kovács-Hostyánszki, A.; Szigeti, V.; Miholcsa, Z.; Sándor, D.; Soltész, Z.; Török, E.; Fenesi, A. Threats and Benefits of Invasive Alien Plant Species on Pollinators. Basic Appl. Ecol. 2022, 64, 89–102. [Google Scholar] [CrossRef]
- Makimura, F.; Tsuruta, M.; Yamasaki, K.; Mukai, Y. A Study of Pollinators of the Endangered Species Pyrus Calleryana Decne. in the Natural Habitat of the Callery Pear in Mie Prefecture, Japan. Conserv. Ecol. Res. 2015, 20, 197–202. [Google Scholar] [CrossRef]
- Marcos-Garcia, M.A.; Garcia-Lopez, A.; Zumbado, M.A.; Rotheray, G.E. Sampling Methods for Assessing Syrphid Biodiversity (Diptera: Syrphidae) in Tropical Forests. Environ. Entomol. 2012, 41, 1544–1552. [Google Scholar] [CrossRef] [Green Version]
- Bjerknes, A.L.; Totland, Ø.; Hegland, S.J.; Nielsen, A. Do Alien Plant Invasions Really Affect Pollination Success in Native Plant Species? Biol. Conserv. 2007, 138, 1–12. [Google Scholar] [CrossRef]
- Schmidt, J.O. Feeding Preferences of Apis Mellifera L. (Hymenoptera: Apidae): Individual versus Mixed Pollen Species. J. Kans. Entomol. Soc. 1984, 57, 323–327. [Google Scholar]
- Wood, T.J.; Roberts, S.P.M. An Assessment of Historical and Contemporary Diet Breadth in Polylectic Andrena Bee Species. Biol. Conserv. 2017, 215, 72–80. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Planted Trees | Feral Trees | Collection Days |
|---|---|---|---|
| 2018 | 7 | 0 | 3 |
| 2019 | 0 | 0 | 0 |
| 2020 | 2 | 1 | 2 |
| 2021 | 0 | 1 | 1 |
| 2022 | 10 | 7 | 6 |
| Total | 19 | 9 | 12 |
| Order: Family | Lower Classification | Planted | Feral |
|---|---|---|---|
| Hymenoptera: Apidae | Apis mellifera, L. | 0.24 | 0.11 |
| Hymenoptera: Andrenidae | Andrena spp., Fabricius | 0.20 | 0.15 |
| Hymenoptera: Vespidae | Polistes spp., Latreille | 0.02 | 0.06 |
| Diptera: Tachinidae | Epalpus signifier, Walker | 0.02 | 0.06 |
| Hymenoptera: Megachilidae | Megachile spp., Latreille | 0.11 | |
| Hymenoptera: Apidae | Bombus spp., Latreille | 0.11 | |
| Diptera: Syrphidae | Helophilus fasciatus, Walker | 0.05 | |
| Hymenoptera: Megachilidae | Osmia spp., Panzer | 0.05 | |
| Hymenoptera: Halictidae | Halictus spp., Latreille | 0.05 | |
| Hymenoptera: Apidae | Xylocopa virginica, L. | 0.11 | |
| Hymenoptera: Halictidae | Augochlora pura, Say | 0.11 | |
| Coleoptera: Coccinellidae | Harmonia axyridis, Pallas | 0.11 | |
| Diptera: Syrphidae | unknown | 0.11 | |
| Hemiptera: Cydnidae | unknown | 0.11 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dolan, B.J.; Kalkstein, A.L.; Haase, M.; Chernisky, P.; Niese, A.; Hicks, A.N.; Miller, D. Pollinator Communities of Planted and Feral Pyrus calleryana. Ecologies 2023, 4, 1-10. https://doi.org/10.3390/ecologies4010001
Dolan BJ, Kalkstein AL, Haase M, Chernisky P, Niese A, Hicks AN, Miller D. Pollinator Communities of Planted and Feral Pyrus calleryana. Ecologies. 2023; 4(1):1-10. https://doi.org/10.3390/ecologies4010001
Chicago/Turabian StyleDolan, Benjamin J., Abby L. Kalkstein, Madeline Haase, Paige Chernisky, Alivia Niese, Ashlie N. Hicks, and Dolly Miller. 2023. "Pollinator Communities of Planted and Feral Pyrus calleryana" Ecologies 4, no. 1: 1-10. https://doi.org/10.3390/ecologies4010001