
Contribution of smFRET to Chromatin Research

1
Wexner Medical Center, Ohio State University, Columbus, OH 43210, USA
2
Department of Chemistry, Pennsylvania State University, University Park, PA 16801, USA
*
Author to whom correspondence should be addressed.
Biophysica 2023, 3(1), 93-108; https://doi.org/10.3390/biophysica3010007
Submission received: 30 November 2022
/
Revised: 27 January 2023
/
Accepted: 31 January 2023
/
Published: 8 February 2023
(This article belongs to the Collection Feature Papers in Biophysics)
Abstract
:Chromatins are structural components of chromosomes and consist of DNA and histone proteins. The structure, dynamics, and function of chromatins are important in regulating genetic processes. Several different experimental and theoretical tools have been employed to understand chromatins better. In this review, we will focus on the literatures engrossed in understanding of chromatins using single-molecule Förster resonance energy transfer (smFRET). smFRET is a single-molecule fluorescence microscopic technique that can furnish information regarding the distance between two points in space. This has been utilized to efficiently unveil the structural details of chromatins.
1. Introduction
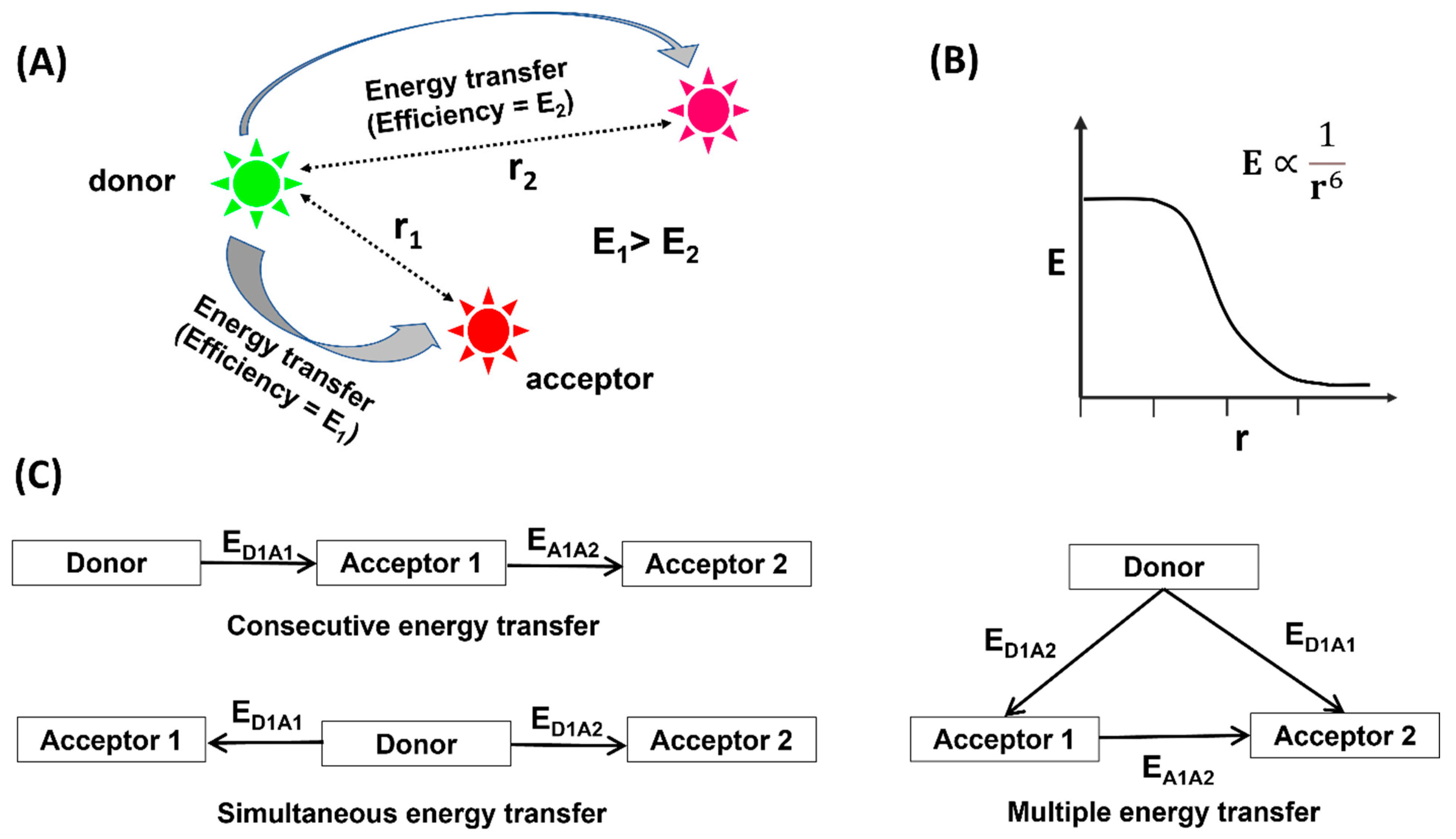
FRET, or Förster resonance energy transfer, defines the phenomena of energy transfer between two fluorescent molecules residing within a specific distance. The idea that energy can be transferred from one particle (atom or molecule) to the other without physical contact was realized around the year 1900. Several theoretical and experimental contributions came from eminent scientists such as Heisenberg, Schrödinger and Dirac, and Jean-Baptiste Perrin and his son Francis Perrin during the first half of the 20th century [1], among which was the first experiment of energy transfer on condensed systems by Perrins in 1920 [1]. Although Perrin was able to see the distance dependence of the energy transfer process, he was unable to determine the appropriate range for effective energy transfer. In 1946 Theodor Förster came up with his groundbreaking work communicating the proper distance dependence of the energy transfer [2].
To honor Förster’s contribution, the term Förster resonance energy transfer (FRET) was coined. Although sometimes an erroneous illustration, fluorescence resonance energy transfer is also used for easier understanding of the larger scientific community. The basic requirement for the occurrence of FRET is to have two fluorophores having appropriate spectral properties for effective energy transfer within a specific distance. Upon excitation, the donor fluorophore will absorb the energy and relay it to the acceptor (Figure 1A). The distance between two fluorophores can be calculated from its dependence on the efficiency of energy transfer (E) using a proper mathematical expression (Figure 1B). During the second half of the 20th century, a noteworthy amount of research articles was published on the practical application of FRET [3,4,5,6,7,8,9,10]. FRET was applied to diverse systems including metal complexes [11,12,13], proteins [14,15], nucleic acids [16,17,18], and several other systems [8,19,20,21] in solution. Soon, it was realized that the ensemble average of the observables significantly influenced the conclusions drawn therein. To circumvent this, the idea of a single-molecule FRET was conceived. The first report on smFRET came in 1996 from T. J. Ha and coworkers [22]. Soon, smFRET gained immediate attention from researchers around the world owing to its potential capability to extract crucial information regarding structural dynamics of large molecules, dynamics of intra- and intermolecular interactions, the mechanisms of complex cellular processes, and more. The domain of biosciences was largely benefitted by smFRET [23,24,25,26,27,28,29,30,31,32]. This is mostly accredited to the fact that smFRET works around a distance range that is biologically relevant. Hence, the molecular dynamics and intra- and intermolecular interactions, which occur around 1–10 nm distance, can all be effectively probed using smFRET. Furthermore, smFRET operates in the picomolar range, resulting in low sample requirement. Unlike structure-based measurements such as CryoEM, smFRET deals with solution-based measurements that significantly reduce the sample preparation time. In addition, real-time dynamics of the nucleosome over cellular processes can be visualized in the solution. Lastly, the data collection is straightforward, and the non-radiative energy transfer makes the data analysis much less complicated. Among the various biological systems that were experimented on using smFRET, chromatins form a major subgroup.
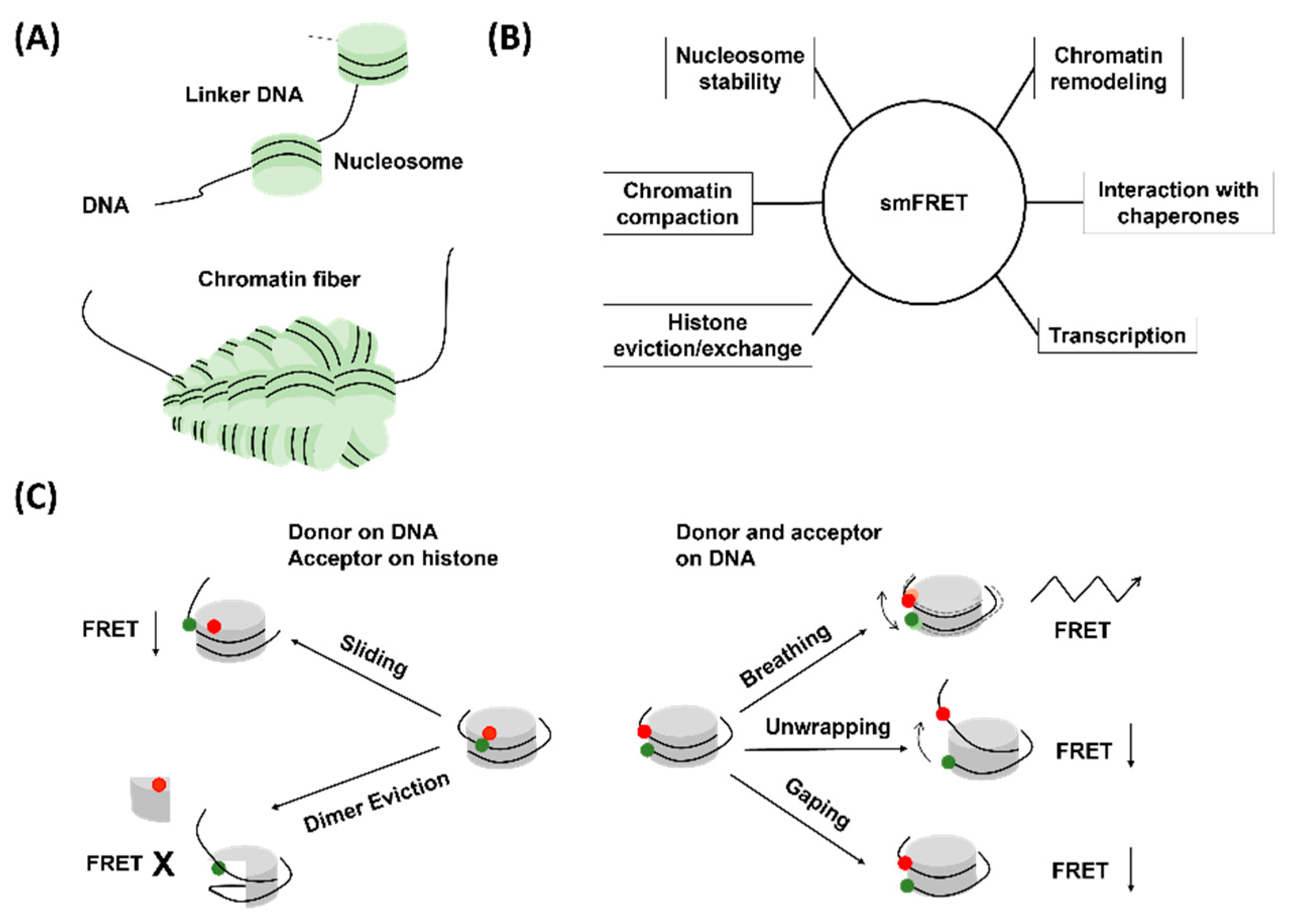
Chromatins are the structural constituents of eukaryotic chromosomes and have a “beads on a string” structure [33]. The “beads” therein are scientifically termed nucleosomes and are composed of DNA and histone proteins, whereas the string is the linker DNA (Figure 2). Chromatins are widely studied owing to the fact that chromatin structures and dynamics are central to understanding the process of gene regulation. Although the existence of genes and chromosomes was registered in the late 19th century, the proposal of the beads on a string structure came up in 1974 from R. D. Kornberg [34]. Based on contemporary research and available results, Kornberg suggested that DNA wraps around the histone core and forms beads, each of which contains about 200 base pairs of DNA [34]. The histone core consists of eight histone proteins in total, which comprises one copy of a histone (H3-H4)2 tetramer and two copies of histone H2A-H2B dimers [35]. The beads are the structural units of chromatin and are termed as nucleosomes. In 1997, the pioneering work from K. Luger reporting the crystal structure of nucleosomes at 2.8 Å resolution was published [35]. From late 20th century, chromatin research started going uphill. Diverse scientific techniques such as imaging techniques (FRET, SAX, AFM, cryo-EM) and genome-wide accessibility assays (MNase-seq, DNase-seq, ATAC-seq) were applied to better understand chromatin structure, dynamics, and behavior [36,37,38,39]. As time passed and the idea of a single molecule started surfacing, it did not take long for scientists to implement it for chromatin research.
As already mentioned, in the event of FRET, distance-dependent energy transfer happens between the donor and the acceptor fluorophore molecules (Figure 1) [40]. By selectively inserting a suitable donor–acceptor couple to two locations on a large biomolecule, the relative changes between these two locations can be observed. This information helps in unveiling information about conformational changes in the molecule. The labeling of the biomolecule with the fluorophores plays a pivotal role in smFRET (Figure 1C). The labeling locations must be decided based on a few important facts. First, the labeling must not interfere with the biological function of the protein. Second, the distance must be between the working distance of the corresponding FRET pair. If one is interested in studying the structural dynamics, they must work with sites that undergo distance change upon rearrangement. It is also common to work with multiple labeling positions to extract maximum information. Most of the time, fluorescence labeling requires site-directed mutagenesis of the biomolecules. It is essential that the mutagenesis does not significantly perturb the structure and function of the molecule. In the case of chromatins, different combinations of labeling schemes have been practiced. The labels must be inserted in such a way that the compactness of the nucleosome, the DNA wrapping around the histone core, and the histone–DNA interactions are unperturbed. In addition, it is preferred to have the label in the non-template DNA strand to avoid interference in biological processes such as translation and transcription. Putting both the fluorophores on the DNA strand delivers information about the DNA unwrapping/rewrapping, whereas inserting one of them in the histone core helps in understanding the relative changes in the DNA and histone core, which might be caused by dimer eviction, dimer exchange, DNA sliding, and so on. Therefore, just by changing the labeling locations, one can bring out information about several processes associated with chromatins. The generally accepted labeling scheme for the smFRET study of nucleosomes corresponds to a high FRET efficiency in the native structure (DNA properly wrapped around the histone core) of the nucleosome. Upon any structural change, such as unwrapping of DNA, sliding or dimer eviction, the FRET efficiency usually decreases, and the extent of change is reflective of the extent of structural alteration. However, this is not universal, and exceptions exist. Apart from the simplest case of FRET between two fluorophores, complex systems have also been studied with three- or four-color FRET, where simultaneous or consecutive energy transfer between different fluorophore pairs occur [41,42,43,44,45,46].
In this review, we have attempted to summarize all the research works concentrated on the use of smFRET on chromatin systems (Table 1). The plethora of articles have been segregated into two segments:
- Chromatin dynamics and compaction;
- Interaction with chaperones and other protein machinery.
2. Chromatin Structures and Dynamics
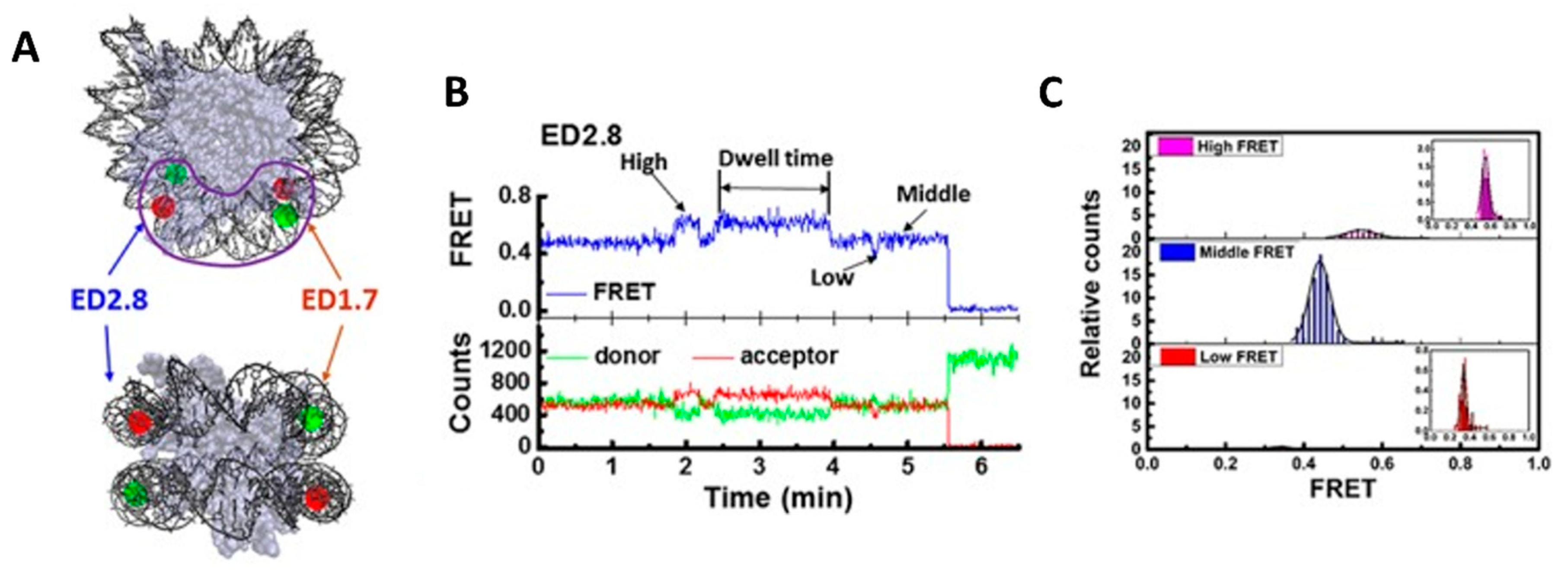
Structure and dynamics of native nucleosome: The first attempt to study nucleosomes using single-molecule fluorescence was made by J. Widom. Widom employed fluorescence correlation spectroscopy (FCS) to understand the conformational fluctuations of the nucleosomes and was pretty successful [47]. Soon after this, researchers realized that the nucleosomal DNA can be labeled site-specifically in such a way that the unwrapping of the nucleosomal DNA will induce a significant decrease in the FRET efficiency. M. Tomschik and his group reported the conformational dynamics of nucleosomes between the closed and the open conformation (DNA loosely bound) using single-molecule FRET [48,81]. This marks the first of many research articles focused on the study of chromatins using smFRET. The closed conformation is defined by the DNA tightly wrapped around the histone core, and the open conformation is defined by significantly loosened nucleosomal DNA around the histones. In 2009, Gansen et al. was able to see three (sub)nucleosome populations by monitoring the distance between a FRET pair on nucleosomal DNA: an intact tightly bound nucleosome, a partially unwrapped intermediate, and another partially unwrapped intermediate with a higher degree of DNA opening [49]. The existence of these (sub)nucleosome populations was explained by the transient unwrapping of DNA ends, which can lead to the displacement of the H2A-H2B dimer. In the same year, Koopmans et al. published another article reporting the equilibrium constants of DNA unwrapping using single-molecule FRET [50]. Later in 2015, Ngo et al. once more confirmed the presence of three (sub)nucleosome conformations [51]. By labeling DNA with two different FRET pairs on nucleosomal DNA to monitor the changes at both DNA ends, Ngo et al. revealed that nucleosomes undergo slow spontaneous conformation switching from one state to another beyond tens of seconds, and these conformational changes occur at both DNA ends with different extents of unwrapping (Figure 3) [51]. In a different study to investigate how nucleosomes undergo spontaneous unwrapping, Bohm et al. utilized a FRET pair where DNA and either histone H2A-H2B dimer or histone (H3-H4)2 tetramer were labeled [52]. This study observed DNA spontaneously opening before the dissociation of the histone H2A-H2B dimer, which substantiates previous studies on (sub)nucleosome intermediates with partially unwrapped DNA due to the potential loss of DNA–histone contacts. It also reported that nucleosomes undergo stepwise disassembly and reassembly [52]. During the disassembly process, the H2A-H2B dimer dissociates before the H3-H4 tetramer, and the order is reversed during reassembly [52].
The fact that intrinsic properties of nucleosome structure and dynamics are highly dependent on the environmental conditions has also been tested with smFRET. Gansen et al. reported that nucleosomes are destabilized at low nucleosome concentrations (~pM) and high ionic conditions [49]. Changes in nucleosome structure were observed within a wide range of monovalent salt concentrations. Within 5–100 mM NaCl concentrations, the authors demonstrated that increased salt concentration leads to an increased population of partially unwrapped nucleosomes [49]. The destabilizing effects of high ionic condition were experimented with in another paper by Ngo et al., where 50 mM to 1 M NaCl concentrations were utilized instead [51]. In this paper, the authors tried to investigate whether loss of DNA–histone contacts is responsible for the nucleosome-destabilizing effects of high salt concentrations [51]. The authors found that the salt-induced nucleosome disassembly process is reversible, which leads to the conclusion that histone H2A-H2B dimer(s) do not become completely evicted and are still retained in the nucleosome core [51]. Another interesting article came up from the same group where they tried to bridge the gap between ensemble and single-molecule FRET measurements by finding a way to operate single-molecule measurements at ensemble concentrations and vice versa. Through this work, they were able to find an intermediate conformational state, which the acetylated nucleosome adopts while unwrapping [53]. The histone eviction process has also been studied by Lee et al. using three-color smFRET, where they found that in the absence of any chaperone, the nucleosomal DNA needs to unwrap before the dimer can be evicted [54].
Overall, a few key points elucidated from the abovementioned reports are: (1) nucleosome structure is dynamic, (2) nucleosome can undergo reversible disassembly process, and (3) nucleosome stability is dependent on both nucleosome and salt concentrations.
Apart from investigating mononucleosomes, smFRET has also been used to unveil details about higher-order structures, such as the nucleosomal array. Understanding the driving force of chromatin compaction is extremely important considering that this is the key to accommodation of the long DNA in the cell. In 2018, Kilic et al. published the effects of HP1α on chromatin compaction [55]. Using three different combinations of site-specific tagging locations of the donor and the acceptor, they observed the transient formation of tetrameric structures. They also established that this event is triggered by the interaction of the HP1α protein with methylated histone tails, and the timescale depends on the lifetime of this interaction [55]. Using a similar experimental approach, Langowski’s group studied the effects of H3 N-terminal tails (H3NtT) on salt-dependent nucleosome stability and the interplay of chromatin compaction and accessibility of DNA [56]. They disclosed that spatial movement or diffusion of H3NtT is coordinated with DNA movement. Furthermore, they were successful in recording the ability of H3NtT to perceive charge-altering mutations to the histone core.
Effects of DNA modifications on nucleosome structure: The ~150 base pair long DNA wrapping around the histone core is called the nucleosome positioning sequence (NPS), and the DNA connecting the nucleosome beads is called the linker DNA. The tightly bound compact structure of the nucleosome is managed by the electrostatic interactions of DNA with histones. Therefore, modifications to DNA can alter these electrostatic interactions, resulting in a transformation of the nucleosome structure and dynamics. This fact has also been tested with the use of smFRET.
In 2015, Lee et al. published a paper on the role of DNA methylation on nucleosome dynamics [57]. The authors utilized a three-color labeling scheme where DNA was labeled with a donor and an acceptor, and the histone core was labeled with a second acceptor. By monitoring the histone deposition on the DNA template and the DNA rewrapping, the authors reported that CpG methylation on DNA facilitates stable tetrasome (DNA-(H3-H4)2) formation due to efficient DNA rewrapping while inhibiting the incorporation of the histone H2A-H2B dimer on the same CpG-methylated DNA [57]. The authors came up with possible pathways where CpG methylation assists nucleosome assembly: either by the complete histone octamer deposition followed by DNA rewrapping, or by the sequential steps of (H3-H4)2 tetramer deposition, and then by DNA rewrapping followed by the recruitment of histone H2A-H2B dimer [57].
Effects of histone PTMs (post-translational modifications) on nucleosome structure and dynamics: Histone components in nucleosomes are not universal. Nucleosomes can undergo post-translational modifications during the cell cycle such as phosphorylation, acetylation, ubiquitylation, and many more. These modifications play a vital role in regulating chromatin structure and function because they are usually coupled with certain biological processes that either lead to enhanced or repressed gene expression. Histone modifications have been a point of interest because of their abundance near active or repressed genes.
Histone acetylation markers have been coupled with gene activation, and deficiency in the acetylation marker can lead to neuronal and memory disorders. In 2009, Neumann et al. studied the effects of histone acetylation on nucleosome dynamics [58]. The authors generated acetylated histone H3 at Lysine 56 position (in short, H3K56ac) and utilized a FRET pair on the DNA near the nucleosome entry site in order to observe the structural changes in the nucleosomes [58]. The authors reported a seven-times increase in nucleosome “breathing” dynamics, meaning that H3K56ac induces enhanced spontaneous DNA opening at the DNA ends [58]. This observation complies with the known function of histone acetylation to promote gene expression.
Another important histone PTM is H3K122 succinylation. This marker has been found to be abundant near the active genes and is proven to increase the output of transcriptional products. In 2020, Shahidian et al. used smFRET to show that H3K122 succinylated nucleosomes have significantly less stability as compared to the wild-type nucleosomes [59]. Using smFRET and biochemical assays, they compared the acetylation and succinylation to show that the transcriptional output is not only regulated by the nucleosome structure but also by a consortium of other factors [59].
Recently, Lee’s group published an article that reports the effects of ubiquitylation on the nucleosome structure and dinucleosome formation [60]. Ubiquitylation is also proven to facilitate the transcription process. As per their findings, ubiquitylation at the 34th lysine in H2B destabilizes the nucleosome by incorporating perturbations to the native structure [60]. At the same time, it stabilizes the internucleosomal interactions, which might relate to stable higher-order structure formation [60].
Besides histone post-translational modifications, past studies have suggested that the linker histone H1 can regulate the nucleosome conformations in a higher order of chromatin. In 2016, Fang et al. explored the functions of linker histone H1 by looking at its conformational changes in the presence of nucleosomes [61]. The authors revealed three distant histone H1 populations by monitoring the distance between a FRET pair on the histone H1 C-terminal domain: H1-bound mononucleosome, H1-bound nucleosome array, and unbound H1, thus providing additional information on how linker histone H1 enacts its reorganization effects on chromatin [61].
3. Interaction of Chromatins with Protein Machineries
Nucleosomes undergo multiple biological processes such as DNA replication, transcription, and DNA repair. All these actions follow a common mechanism of DNA unwrapping or distortion of the native structure, followed by the genetic process and finally DNA rewrapping around the histone core. These steps are directly controlled by designated multimodular proteins and are assisted by specific molecular chaperones. In the next section we will discuss some scientific works centered on the interactions of these proteins with nucleosomes.
Interaction with remodelers: Chromatin remodelers help in increasing the accessibility of the DNA or the histone core to other proteins or enzymes by altering the native structure. On the other hand, they help to maintain the original structure of nucleosomes by assisting in structural rearrangements after the genetic procedure is over. This multitasking capability makes chromatin remodelers crucial for the fitness of the cell, and their defects can lead to cancers and lethal diseases. ATP-dependent chromatin remodelers only work in the presence of ATP and are classified into four distinct subsets based on their structure and functions: SWI/SNF (SWItch/sucrose non-fermentable), ISWI (imitation SWI/SNF), Chd (chromodomain-helicase DNA-binding), and INO80 (INOsitol-requiring mutant 80). In this mini-review, we will discuss some examples of chromatin remodelers that have been investigated by the single-molecule FRET method.
In 2009, Blosser et al. published on the actions of ACF (ATP-dependent chromatin assembly and remodeling factor) on the nucleosome structure and dynamics during remodeling [62]. ACF belongs to the SWI/SNF family, and it has been known to translocate the histone core to help in nucleosome reassembly. The authors labeled the histone core and DNA with a FRET pair and monitored its relative changes during nucleosome remodeling [62]. They found that nucleosome remodeling undergoes two steps, which are ATP-dependent ACF binding to the nucleosome core and ATP-dependent ACF translocation activity [62]. ACF translocation is not continuous, and it is a repeated cycle where the translocation of the nucleosomes occurs with a short DNA distance of 7 or 3–4 base pairs in succession, and then, it follows by a stable pause in between each translocation event [62]. By altering the fluorophore locations on the DNA to observe changes in both DNA ends, the authors also found that ACF can translocate the histone octamer both upstream and downstream of the original position [62].
In 2013, Deindl et al. assessed a different family of chromatin remodelers—ISWI—using similar methods [63]. ISWI functions to translocate nucleosomes, and its mechanisms were unclear at the time. Similar observations to ACF were obtained, which means that ISWI also translocates nucleosomes in short DNA bursts followed by rest periods [63]. The authors looked closely into the DNA movements from both DNA ends and discovered that the seven-base-pair translocation at the DNA exit end is made up of multiple one-base-pair steps where the three-base-pair translocation is a single compound step [63]. They came up with a translocation model where the nucleosome builds pressure during seven-base-pair translocation and releases it during three-base-pair translocation. In 2019, a very interesting work came up from the same group [64]. They showed that there is a lag between the sliding from the entry and exit side for Chd1 and SNF2h remodelers, and this lag is attributed to the absorption of 1–3 base pairs by the remodeler [64]. Based on their three-color smFRET results, they propose that this DNA buffering stabilizes the nucleosome during remodeling and helps in communication between two remodelers working on the opposite sides [64].
In 2016, Harada et al. investigated the mechanism of RSC (remodeling of the structure of chromatin) during the remodeling process [65]. RSC also belongs to the SWI/SNF family, and in the project, the authors employed various labeling schemes (either on DNA or on histone components in the nucleosome structure) to monitor RSC-induced nucleosome reposition [65]. This work provides additional proof to validate that SWI/SNF chromatin remodelers function bidirectionally and independently of DNA sequence. Furthermore, it reveals that all histone components retain and move along the DNA together during the translocation process without overall significant nucleosomal DNA distortions [65]. Consistent with previously mentioned studies, Harada et al. demonstrates that these chromatin remodelers are likely to translocate around 1–2 DNA base pairs each time, but they are still open to the possibilities that the translocation step size may go beyond two base pairs [65].
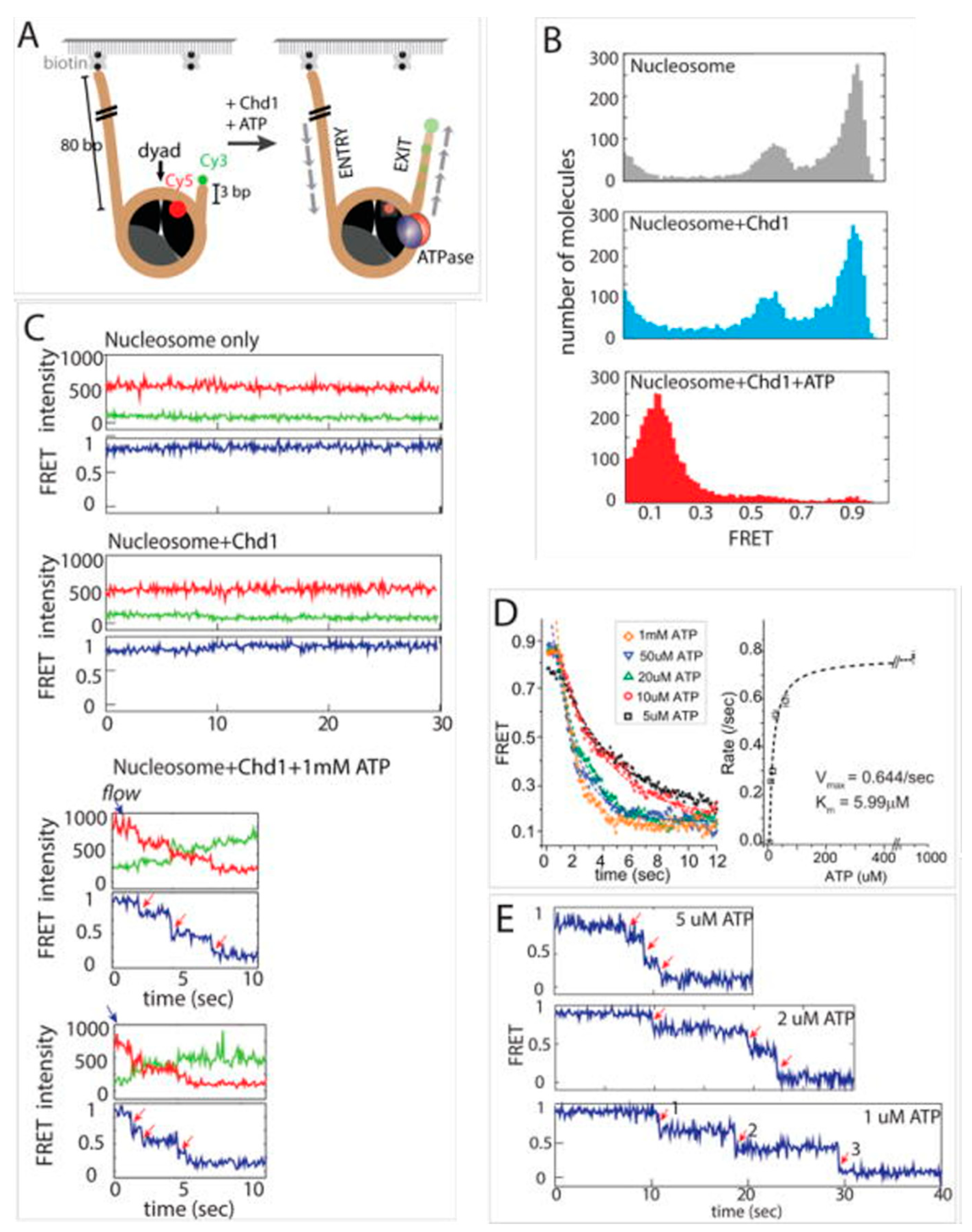
The following year, Qiu et al. published an article on the mechanism of Chd1 (monomeric)-mediated remodeling of chromatins [66]. Using smFRET, Qiu et al. showed that the DNA binding domains help Chd1 to interact with chromatin and slide it bidirectionally [66]. During this sliding, unstable intermediates are generated, which eventually relax to a stable remodeled structure (Figure 4). They also proved that the presence of N-terminal chromodomain helps it to distinguish between hexasomes and nucleosomes and to selectively remodel nucleosomes only [66].
Narlikar’s group published an article in 2018 showing the significance of the nucleosome acidic patch formed by H2A and H2B on the remodeling process [67]. Using smFRET and other biochemical assays, they proved that both ISWI and INO80 remodelers require the presence of this acidic patch to become activated [67]. Additionally, their smFRET data show how the acidic patch is crucial for chromatin remodelers to escape the pause phase and to move into the translocation phase [67]. This pause phase is generated by the autoinhibitory behavior of the AutoN and NegC motifs, and the acidic patch interacts with these two motifs to overcome the pause barrier [67]. Soon after this, the same group came up with another article which was structurally similar to the 2017 work by Qiu et al. but which deals with the INO80 remodeler [68]. They showed that the Nhp10 module of INO80 has autoinhibitory effects that can be switched off upon interaction with the flanking DNA [68]. The sliding is highly dependent on the flanking DNA length and increases 100-fold when the DNA length is increased from 40 base pairs to 60 base pairs. Although this length does not affect the sliding steps, as in both cases, the INO80 slides the DNA by 20 base pairs. Another article was published in 2022 by the same group on chromatin remodeling by the SNF2 dimer [69]. They used a synthetic dimer of the SNF2 motor to show that the directionality of nucleosome sliding is governed by ATP hydrolysis of both ATP hydrolyzing motors [69]. The two motors coordinate with each other to decide the direction of sliding upon ATP hydrolysis [69].
Schwarz et al. shed further light on INO80-initiated chromatin remodeling [70]. They studied the effects of histone tails in the remodeling process, as it was established that tailless nucleosomes exhibit increased remodeling [70]. They showed that the initial conformation of DNA in the entry region remains the same in both wild-type and tailless nucleosomes [70]. However, the ADP-bound states are much more heterogenous in the case of tailless nucleosomes [70]. Based on their results and previous reports, they proposed that the increased remodeling for tailless constructs may arise due to the reduced nucleosome stability [70]. In 2022, Wu’s group published an article on the mechanistic details and timescale of the histone dimer exchange [71]. The histone H2A-H2B dimer is replaced by the H2A.Z-H2B variant at the +1 nucleosome downstream of the transcription start site. Using three-color FRET, they evaluated that the whole process of dimer exchange happens within 2.3 s [71]. During this time, SWR1-assisted DNA unwrapping and dimer exchange happen from the same face of the nucleosome [71].
From all single-molecule FRET studies mentioned above, there are some commonalities in the mechanisms of chromatin remodelers: (1) chromatin remodelers can translocate nucleosomes similarly in both ways and (2) they function in a repeated cycle between a short stretch of DNA translocation and a stable rest stage.
Interaction with other protein machineries: Apart from the remodelers, nucleosomes also interact with other protein machineries such as replication proteins, polymerases, transcription factors and many more. We have indicated significant works in this aspect.
Poirier’s group demonstrated FRET efficiency fluctuations of the labeled nucleosome in the presence of transcription factors. They showed that the binding rate reduces dramatically in the entry–exit regions of the nucleosomes as compared to duplex DNA. On the other hand, the dissociation rate increases by three orders of magnitude [72]. They labeled the DNA with Cy3 in the entry–exit region and H2A with Cy5 to study LexA and Gal4 binding to the nucleosome [72]. Furthermore, using single-molecule methods, they were able to calculate the binding and dissociation constants of LexA and Gal4 with nucleosomes [73].
In 2017, Reese’s group reported on the role of the elongation factor Spt4/5 on the transcription process using two-color FRET [74]. This work delivers that Spt4/5 interacts with the nucleotides and RNA Polymerase II (Pol II) to facilitate Pol II escape from pause near the H3-H4 and DNA contact points [74]. The usage of different DNA sequences shows that the effect and requirement of Spt4/5 highly depend on the strength of H3-H4 to DNA interaction [74]. They also observed a significant assistance from Spt4/5 in DNA rewrapping behind Pol II [74].
In 2017, Fishel’s group showed that nucleosomal DNA can undergo stochastic stepwise unwrapping in the presence of ATP and Rad51 aggregated on double-stranded DNA [75]. This unwrapping was found to be independent of the nucleosomal DNA sequence [75]. There is a battle between attachment of the Rad51 assembly to the DNA and the rewrapping of the DNA, which makes the process reversible [75].
Dot1L (disruptor of telomeric silencing 1-like) is a histone (H3K79) methyltransferase, which is assisted by H2BK120 ubiquitylation. In 2019, Jang et al. used smFRET combined with cryo-EM to show the destabilization of the nucleosome structure upon Dot1L binding [76]. They also observed cross-talk between H2BK120 ubiquitylation and Dot1L, which they believed plays a critical role in the destabilization process [76]. Recently, McGinty’s group published another report where they used two-color smFRET to observe the mechanistic details of the interactions of Dot1L with the acidic patch of the nucleosomal surface and with H2BK120 ubiquitin [77]. They spotted that Dot1L needs to have direct interaction with the ubiquitin to form a stable acidic-patch-bound intermediate [77]. Moreover, they detected a contribution of S-adenosyl methionine (SAM) to induce a conformational change of the Dot1L–nucleosome complex to initiate methylation [77].
Rap1 (Repressor activator protein 1) is a pioneer transcription factor (pTF) that works as a precursor for the subsequent binding of downstream factors and chromatin remodelers. In 2020, Fierz’s group used two-color FRET to report the binding of Rap1 to DNA [78]. Based on their results, they proposed the transient binding and unbinding of Rap1 to the compact chromatin fiber [78]. This transient interaction introduces some disruptions on the internucleosomal contacts, which eventually lead to the loosening of the chromatin fiber. They also showed that in the presence of the chromatin remodeler RSC, which displaces the promoter nucleosomes, Rap1 can form stable bound states with the exposed DNA [78].
In 2020, the Lee group published an article reporting the effects of acetylation on the transcription process [79]. They used two-color smFRET to show that acetylation at K56 of H3 greatly reduces the Pol II pause duration near the entry site of the nucleosome [79]. This way, H3K56ac promotes a more efficient transcription. They also tested the effects of histone chaperone Nap1 (nucleosome assembly protein 1) on the DNA rewrapping but failed to see any assistance from Nap1 in absence of other factors [79].
FACT (facilitates chromatin transcription) is a histone chaperone, the yeast homolog of which is a heterodimer and comprises Pob3 and Spt6 subunits. FACT acts in the presence of the DNA-binding protein Nhp6. Recently, Duderstadt’s group reported the effects of different domains of each subunit with the help of two-color FRET [80]. They observed significant decreases in FACT activity when middle and C-terminal domains of Pob3 and Spt6 are compromised but when the N-terminal domain does not have a significant role in the chaperoning process [80].
4. Summary and Future Directions
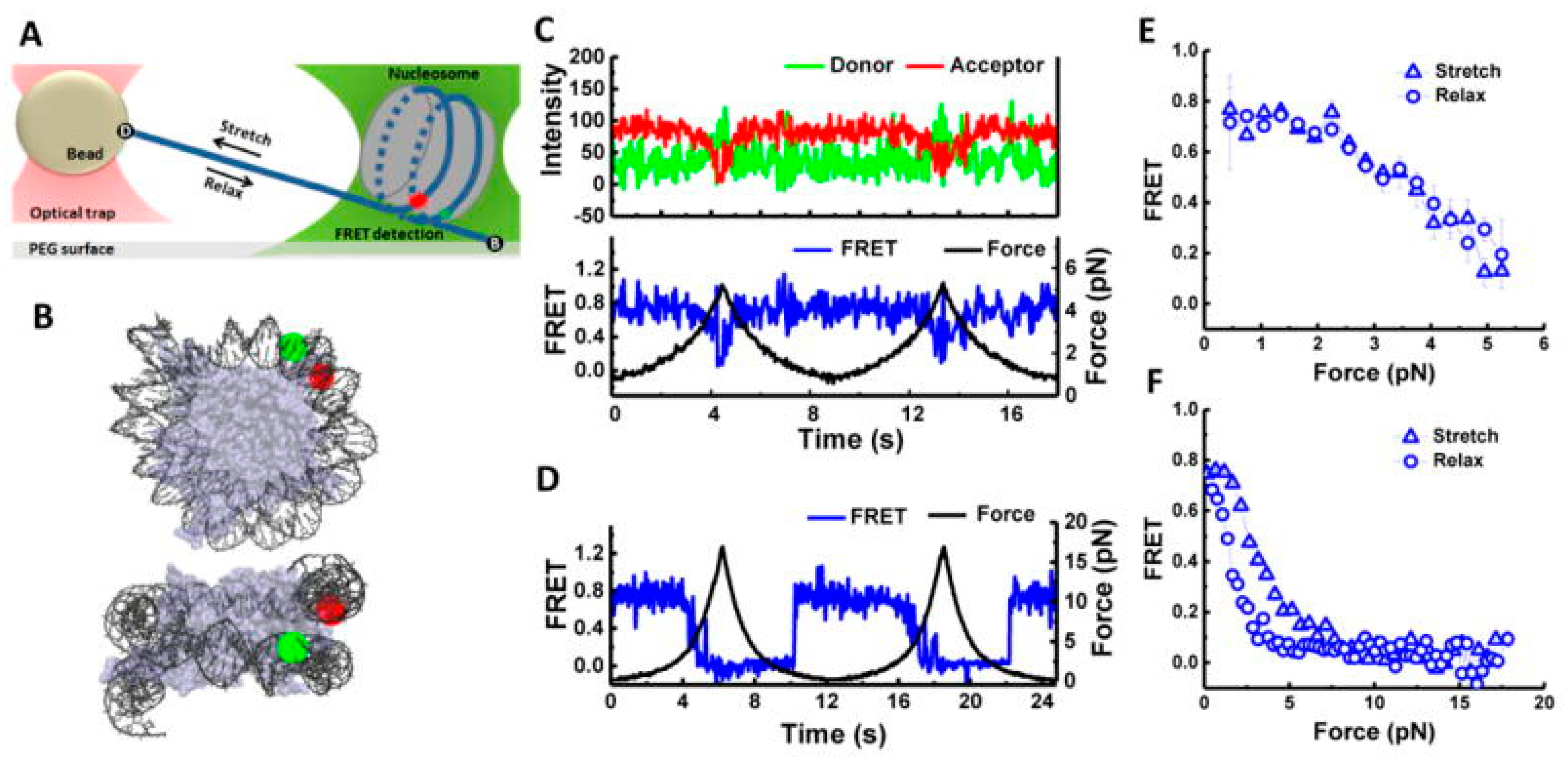
We have discussed the research works published to date on the use of smFRET to understand chromatin structure, function, and dynamics. We have only included works that deal with nucleosomes or chromatin directly. A lot of information regarding chromatins is already known, yet a lot remains unexplained. Like every experimental technique, smFRET also has limitations that restrict its use in certain cases. Especially, ultrafast processes were difficult to observe with smFRET owing to the manual error(s) and limitation(s) of the electronic components. First and foremost, the process of FRET is highly controlled by the fluorophore; thus, the extractable information is constrained by the behavior of the donor–acceptor pair. FRET only operates in a given distance range specific to the donor–acceptor pair. Any crucial details that happen beyond this range cannot be studied using FRET. If the experimental conditions are not optimized properly, the interference of the dye photophysics and bleaching can make the data analysis significantly complicated. Due to irreversible bleaching of the dyes, it is practically impossible to study the mechanism of lengthy processes. The site-specific dye labeling of protein is often strenuous due to the complex architecture and chances of multiple labelings due to the amino acid sequence. The need for multiple-site-directed mutagenesis to get the labels to work often increases the workload and delays an effective outcome. In some unfortunate cases, after successful labeling, sometimes the protein loses its activity due to chemical alteration, and the system is no longer suitable for experimentation. Despite all these conditions, smFRET is thriving in the scientific community as the most reliable tool owing to its efficiency in successfully predicting finer details about the structural reformations. As time proceeds, advancements and improvements are being made to the instrumentation, enabling the capture of the finest details of the chromatin processes [82]. A perfect example would be the use of multiparameter fluorescence detection (MFD) with smFRET by Langowski’s group. Unlike the conventional smFRET measurement, MFD includes the FRET properties of the FRET donor as well as donor–acceptor stoichiometry of the FRET pair. This work shows the stepwise breakdown of the nucleosome structure upon exposure to a high salt concentration. As per their group, the starting point of the degradation is breathing-motion-induced DNA unwrapping followed by histone dimer eviction [83]. Lamond’s group also conducted pronounced work on using multiphoton FLIM (fluorescence lifetime imaging microscopy) to measure the chromatin compaction using a different set of donor- and acceptor-labeled nucleosomes [84]. Furthermore, smFRET is being coupled with other structure measurement techniques and simulations, which can deliver crucial structural insights into the chromatins. Back in 2007, in a classic work, Levitus and coworkers used fluorescence correlation spectroscopy (FCS) with smFRET to study the conformational dynamics of nucleosomes [85]. They tested the accuracy of the technique by matching the outcome with previous results. In 2015, Ha and coworkers used optical tweezers with smFRET to observe the unwrapping of nucleosomal DNA (Figure 5). Using DNA with different sequences, they proved that unwrapping is governed by DNA flexibility [86]. There are other studies in the literature that report force-based measurements on nucleosomes and support the findings of the smFRET results [87,88]. We look forward to a time when we will be able to obtain a complete structural landscape along with proper kinetic data of the nucleosomes and chromatins.
Author Contributions
Conceptualization, B.S.; literature survey, M.H.; writing, B.S and M.H.; review and editing, B.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Clegg, R. Reviews in Fluorescence. 2006. Available online: https://link.springer.com/book/10.1007/0-387-33016-X (accessed on 20 November 2022).
- Sun, Y.; Wallrabe, H.; Seo, S.-A.; Periasamy, A. FRET microscopy in 2010: The legacy of Theodor Förster on the 100th anniversary of his birth. Chemphyschem: A Eur. J. Chem. Phys. Phys. Chem. 2011, 12, 462–474. [Google Scholar] [CrossRef] [PubMed]
- Periasamy, A.; Kay, S.; Day, R. Fluorescence Resonance Energy Transfer (FRET) Imaging of a Single Living Cell Using Green Fluorescent Protein; SPIE: San Jose, CA, USA, 1997; Volume 2983. [Google Scholar]
- Chirio-Lebrun, M.-C.; Prats, M. Fluorescence resonance energy transfer (FRET): Theory and experiments. Biochem. Educ. 1998, 26, 320–323. [Google Scholar] [CrossRef]
- Selvin, P.R. The renaissance of fluorescence resonance energy transfer. Nat. Struct. Biol. 2000, 7, 730–734. [Google Scholar] [CrossRef]
- Selvin, P.R. Fluorescence resonance energy transfer. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1995; Volume 246, pp. 300–334. [Google Scholar]
- Szöllosi, J.; Damjanovich, S.; Mátyus, L. Application of fluorescence resonance energy transfer in the clinical laboratory: Routine and research. Cytometry 1998, 34, 159–179. [Google Scholar] [CrossRef]
- Ju Youn, H.; Terpetschnig, E.; Szmacinski, H.; Lakowicz, J.R. Fluorescence Energy Transfer Immunoassay Based on a Long-Lifetime Luminescent Metal–Ligand Complex. Anal. Biochem. 1995, 232, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.P.; Censullo, M.L.; Kim, H.G.; Knutson, J.R.; Han, M.K. Characterization of Endonucleolytic Activity of HIV-1 Integrase Using a Fluorogenic Substrate. Anal. Biochem. 1995, 227, 295–301. [Google Scholar] [CrossRef] [PubMed]
- Jovin, T.M.; Arndt-Jovin, D.J. 5—FRET Microscopy: Digital Imaging of Fluorescence Resonance Energy Transfer. Application in Cell Biology. In Cell Structure and Function by Microspectrofluorometry; Kohen, E., Ed.; Academic Press: Cambridge, MA, USA, 1989; pp. 99–117. [Google Scholar]
- Pearce, D.A.; Walkup, G.K.; Imperiali, B. Peptidyl chemosensors incorporating a fret mechanism for detection of Ni(II). Bioorganic Med. Chem. Lett. 1998, 8, 1963–1968. [Google Scholar] [CrossRef]
- Aladekomo, J.; Geddes, C.D.; Rolinski, O.J.; Birch, D.J.S. Metal Ion Detection Using Fluorescence Lifetime Sensors. In Proceedings of the European Quantum Electronics Conference, Glasgow, Scotland, 14 September 1998; p. QTuG61. [Google Scholar]
- Olaf, J.R.; David, J.S.B. A fluorescence lifetime sensor for Cu(I) ions. Meas. Sci. Technol. 1999, 10, 127. [Google Scholar] [CrossRef]
- Brown, M.P.; Royer, C. Fluorescence spectroscopy as a tool to investigate protein interactions. Curr. Opin. Biotechnol. 1997, 8, 45–49. [Google Scholar] [CrossRef]
- Heyduk, T. Measuring protein conformational changes by FRET/LRET. Curr. Opin. Biotechnol. 2002, 13, 292–296. [Google Scholar] [CrossRef]
- Quan, K.; Yi, C.; Yang, X.; He, X.; Huang, J.; Wang, K. FRET-based nucleic acid probes: Basic designs and applications in bioimaging. TrAC Trends Anal. Chem. 2020, 124, 115784. [Google Scholar] [CrossRef]
- Füchtbauer, A.F.; Wranne, M.S.; Bood, M.; Weis, E.; Pfeiffer, P.; Nilsson, J.R.; Dahlén, A.; Grøtli, M.; Wilhelmsson, L.M. Interbase FRET in RNA: From A to Z. Nucleic Acids Res. 2019, 47, 9990–9997. [Google Scholar] [CrossRef] [PubMed]
- Clegg, R.M. Fluorescence resonance energy transfer and nucleic acids. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1992; Volume 211, pp. 353–388. [Google Scholar]
- Imani, M.; Mohajeri, N.; Rastegar, M.; Zarghami, N. Recent advances in FRET-Based biosensors for biomedical applications. Anal. Biochem. 2021, 630, 114323. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Hu, Y.; Yang, X.; Tang, Y.; Han, S.; Kang, A.; Deng, H.; Chi, Y.; Zhu, D.; Lu, Y. FÖrster resonance energy transfer (FRET)-based biosensors for biological applications. Biosens. Bioelectron. 2019, 138, 111314. [Google Scholar] [CrossRef] [PubMed]
- Pollok, B.A.; Heim, R. Using GFP in FRET-based applications. Trends Cell Biol. 1999, 9, 57–60. [Google Scholar] [CrossRef] [PubMed]
- Ha, T.; Enderle, T.; Ogletree, D.F.; Chemla, D.S.; Selvin, P.R.; Weiss, S. Probing the interaction between two single molecules: Fluorescence resonance energy transfer between a single donor and a single acceptor. Proc. Natl. Acad. Sci. USA 1996, 93, 6264–6268. [Google Scholar] [CrossRef] [PubMed]
- Krüger, A.C.; Hildebrandt, L.L.; Kragh, S.L.; Birkedal, V. Structural dynamics of nucleic acids by single-molecule FRET. Methods Cell Biol. 2013, 113, 1–37. [Google Scholar] [CrossRef] [PubMed]
- Lamichhane, R.; Solem, A.; Black, W.; Rueda, D. Single-molecule FRET of protein-nucleic acid and protein-protein complexes: Surface passivation and immobilization. Methods 2010, 52, 192–200. [Google Scholar] [CrossRef]
- Kaur, A.; Sapkota, K.; Dhakal, S. Multiplexed Nucleic Acid Sensing with Single-Molecule FRET. ACS Sens. 2019, 4, 623–633. [Google Scholar] [CrossRef]
- Mazal, H.; Haran, G. Single-molecule FRET methods to study the dynamics of proteins at work. Curr. Opin. Biomed. Eng. 2019, 12, 8–17. [Google Scholar] [CrossRef]
- Tan, Y.W.; Hanson, J.A.; Chu, J.W.; Yang, H. Confocal single-molecule FRET for protein conformational dynamics. Methods Mol. Biol. 2014, 1084, 51–62. [Google Scholar] [CrossRef]
- Metskas, L.A.; Rhoades, E. Single-Molecule FRET of Intrinsically Disordered Proteins. Annu. Rev. Phys. Chem. 2020, 71, 391–414. [Google Scholar] [CrossRef] [PubMed]
- Gauer, J.W.; LeBlanc, S.; Hao, P.; Qiu, R.; Case, B.C.; Sakato, M.; Hingorani, M.M.; Erie, D.A.; Weninger, K.R. Single-Molecule FRET to Measure Conformational Dynamics of DNA Mismatch Repair Proteins. Methods Enzymol. 2016, 581, 285–315. [Google Scholar] [CrossRef]
- Wu, S.; Liu, J.; Wang, W. Dissecting the Conformational Dynamics-Modulated Enzyme Catalysis with Single-Molecule FRET. J. Phys. Chem. B 2018, 122, 6179–6187. [Google Scholar] [CrossRef] [PubMed]
- Huynh, M.; Sengupta, B. Analysis of Enzyme Conformation Dynamics Using Single-Molecule Förster Resonance Energy Transfer (smFRET). Biophysica 2022, 2, 123–134. [Google Scholar] [CrossRef]
- Deindl, S.; Zhuang, X. Monitoring conformational dynamics with single-molecule fluorescence energy transfer: Applications in nucleosome remodeling. Methods Enzymol. 2012, 513, 59–86. [Google Scholar] [CrossRef]
- Deichmann, U. Chromatin: Its history, current research, and the seminal researchers and their philosophy. Perspect Biol. Med. 2015, 58, 143–164. [Google Scholar] [CrossRef] [PubMed]
- Kornberg, R.D. Chromatin Structure: A Repeating Unit of Histones and DNA. Science 1974, 184, 868–871. [Google Scholar] [CrossRef]
- Luger, K.; Mäder, A.W.; Richmond, R.K.; Sargent, D.F.; Richmond, T.J. Crystal structure of the nucleosome core particle at 2.8 A resolution. Nature 1997, 389, 251–260. [Google Scholar] [CrossRef]
- Kornberg, R.D.; Lorch, Y. Twenty-five years of the nucleosome, fundamental particle of the eukaryote chromosome. Cell 1999, 98, 285–294. [Google Scholar] [CrossRef] [Green Version]
- Sajan, S.A.; Hawkins, R.D. Methods for identifying higher-order chromatin structure. Annu. Rev. Genom. Hum. Genet. 2012, 13, 59–82. [Google Scholar] [CrossRef]
- Zhou, K.; Gaullier, G.; Luger, K. Nucleosome structure and dynamics are coming of age. Nat. Struct. Mol. Biol. 2019, 26, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Mansisidor, A.R.; Risca, V.I. Chromatin accessibility: Methods, mechanisms, and biological insights. Nucleus 2022, 13, 236–276. [Google Scholar] [CrossRef] [PubMed]
- Albrecht, C. Joseph R. Lakowicz: Principles of fluorescence spectroscopy. Anal. Bioanal. Chem. 2008, 390, 1223–1224. [Google Scholar] [CrossRef]
- Hohng, S.; Joo, C.; Ha, T. Single-molecule three-color FRET. Biophys. J. 2004, 87, 1328–1337. [Google Scholar] [CrossRef]
- Yoo, J.; Louis, J.M.; Gopich, I.V.; Chung, H.S. Three-Color Single-Molecule FRET and Fluorescence Lifetime Analysis of Fast Protein Folding. J. Phys. Chem. B 2018, 122, 11702–11720. [Google Scholar] [CrossRef]
- Voith von Voithenberg, L.; Barth, A.; Trauschke, V.; Demarco, B.; Tyagi, S.; Koehler, C.; Lemke, E.A.; Lamb, D.C. Comparative analysis of the coordinated motion of Hsp70s from different organelles observed by single-molecule three-color FRET. Proc. Natl. Acad. Sci. 2021, 118, e2025578118. [Google Scholar] [CrossRef]
- Lee, J.; Lee, S.; Ragunathan, K.; Joo, C.; Ha, T.; Hohng, S. Single-molecule four-color FRET. Angew Chem. Int. Ed. Engl. 2010, 49, 9922–9925. [Google Scholar] [CrossRef]
- Four-color FRET follows Hsp90 states. Nat. Methods 2014, 11, 791. [CrossRef]
- Stein, I.H.; Steinhauer, C.; Tinnefeld, P. Single-Molecule Four-Color FRET Visualizes Energy-Transfer Paths on DNA Origami. J. Am. Chem. Soc. 2011, 133, 4193–4195. [Google Scholar] [CrossRef]
- Li, G.; Levitus, M.; Bustamante, C.; Widom, J. Rapid spontaneous accessibility of nucleosomal DNA. Nat. Struct. Mol. Biol. 2005, 12, 46–53. [Google Scholar] [CrossRef]
- Tomschik, M.; Zheng, H.; van Holde, K.; Zlatanova, J.; Leuba, S.H. Fast, long-range, reversible conformational fluctuations in nucleosomes revealed by single-pair fluorescence resonance energy transfer. Proc. Natl. Acad. Sci. USA 2005, 102, 3278–3283. [Google Scholar] [CrossRef]
- Gansen, A.; Valeri, A.; Hauger, F.; Felekyan, S.; Kalinin, S.; Tóth, K.; Langowski, J.; Seidel, C.A.M. Nucleosome disassembly intermediates characterized by single-molecule FRET. Proc. Natl. Acad. Sci. USA 2009, 106, 15308–15313. [Google Scholar] [CrossRef]
- Koopmans, W.J.A.; Buning, R.; Schmidt, T.; van Noort, J. spFRET Using Alternating Excitation and FCS Reveals Progressive DNA Unwrapping in Nucleosomes. Biophys. J. 2009, 97, 195–204. [Google Scholar] [CrossRef]
- Ngo, T.T.; Ha, T. Nucleosomes undergo slow spontaneous gaping. Nucleic Acids Res. 2015, 43, 3964–3971. [Google Scholar] [CrossRef]
- Böhm, V.; Hieb, A.R.; Andrews, A.J.; Gansen, A.; Rocker, A.; Tóth, K.; Luger, K.; Langowski, J. Nucleosome accessibility governed by the dimer/tetramer interface. Nucleic Acids Res. 2011, 39, 3093–3102. [Google Scholar] [CrossRef]
- Gansen, A.; Hieb, A.R.; Böhm, V.; Tóth, K.; Langowski, J. Closing the gap between single molecule and bulk FRET analysis of nucleosomes. PLoS ONE 2013, 8, e57018. [Google Scholar] [CrossRef]
- Lee, J.; Lee, T.H. Single-Molecule Investigations on Histone H2A-H2B Dynamics in the Nucleosome. Biochemistry 2017, 56, 977–985. [Google Scholar] [CrossRef]
- Kilic, S.; Felekyan, S.; Doroshenko, O.; Boichenko, I.; Dimura, M.; Vardanyan, H.; Bryan, L.C.; Arya, G.; Seidel, C.A.M.; Fierz, B. Single-molecule FRET reveals multiscale chromatin dynamics modulated by HP1α. Nat. Commun. 2018, 9, 235. [Google Scholar] [CrossRef]
- Lehmann, K.; Felekyan, S.; Kühnemuth, R.; Dimura, M.; Tóth, K.; Seidel, C.A.M.; Langowski, J. Dynamics of the nucleosomal histone H3 N-terminal tail revealed by high precision single-molecule FRET. Nucleic Acids Res. 2020, 48, 1551–1571. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.Y.; Lee, J.; Yue, H.; Lee, T.H. Dynamics of nucleosome assembly and effects of DNA methylation. J. Biol. Chem. 2015, 290, 4291–4303. [Google Scholar] [CrossRef] [PubMed]
- Neumann, H.; Hancock, S.M.; Buning, R.; Routh, A.; Chapman, L.; Somers, J.; Owen-Hughes, T.; van Noort, J.; Rhodes, D.; Chin, J.W. A method for genetically installing site-specific acetylation in recombinant histones defines the effects of H3 K56 acetylation. Mol. Cell 2009, 36, 153–163. [Google Scholar] [CrossRef] [PubMed]
- Zorro Shahidian, L.; Haas, M.; Le Gras, S.; Nitsch, S.; Mourão, A.; Geerlof, A.; Margueron, R.; Michaelis, J.; Daujat, S.; Schneider, R. Succinylation of H3K122 destabilizes nucleosomes and enhances transcription. EMBO Rep. 2021, 22, e51009. [Google Scholar] [CrossRef]
- Sengupta, B.; Huynh, M.; Smith, C.B.; McGinty, R.K.; Krajewski, W.; Lee, T.H. The Effects of Histone H2B Ubiquitylations on the Nucleosome Structure and Internucleosomal Interactions. Biochemistry 2022, 61, 2198–2205. [Google Scholar] [CrossRef] [PubMed]
- Fang, H.; Wei, S.; Lee, T.H.; Hayes, J.J. Chromatin structure-dependent conformations of the H1 CTD. Nucleic Acids Res. 2016, 44, 9131–9141. [Google Scholar] [CrossRef]
- Blosser, T.R.; Yang, J.G.; Stone, M.D.; Narlikar, G.J.; Zhuang, X. Dynamics of nucleosome remodelling by individual ACF complexes. Nature 2009, 462, 1022–1027. [Google Scholar] [CrossRef]
- Deindl, S.; Hwang, W.L.; Hota, S.K.; Blosser, T.R.; Prasad, P.; Bartholomew, B.; Zhuang, X. ISWI remodelers slide nucleosomes with coordinated multi-base-pair entry steps and single-base-pair exit steps. Cell 2013, 152, 442–452. [Google Scholar] [CrossRef]
- Sabantsev, A.; Levendosky, R.F.; Zhuang, X.; Bowman, G.D.; Deindl, S. Direct observation of coordinated DNA movements on the nucleosome during chromatin remodelling. Nat. Commun. 2019, 10, 1720. [Google Scholar] [CrossRef]
- Harada, B.T.; Hwang, W.L.; Deindl, S.; Chatterjee, N.; Bartholomew, B.; Zhuang, X. Stepwise nucleosome translocation by RSC remodeling complexes. Elife 2016, 5. [Google Scholar] [CrossRef]
- Qiu, Y.; Levendosky, R.F.; Chakravarthy, S.; Patel, A.; Bowman, G.D.; Myong, S. The Chd1 Chromatin Remodeler Shifts Nucleosomal DNA Bidirectionally as a Monomer. Molecular Cell 2017, 68, 76–88.e76. [Google Scholar] [CrossRef] [Green Version]
- Gamarra, N.; Johnson, S.L.; Trnka, M.J.; Burlingame, A.L.; Narlikar, G.J. The nucleosomal acidic patch relieves auto-inhibition by the ISWI remodeler SNF2h. Elife 2018, 7. [Google Scholar] [CrossRef]
- Zhou, C.Y.; Johnson, S.L.; Lee, L.J.; Longhurst, A.D.; Beckwith, S.L.; Johnson, M.J.; Morrison, A.J.; Narlikar, G.J. The Yeast INO80 Complex Operates as a Tunable DNA Length-Sensitive Switch to Regulate Nucleosome Sliding. Mol. Cell 2018, 69, 677–688.e679. [Google Scholar] [CrossRef] [PubMed]
- Johnson, S.L.; Narlikar, G.J. ATP Hydrolysis Coordinates the Activities of Two Motors in a Dimeric Chromatin Remodeling Enzyme. J. Mol. Biol. 2022, 434, 167653. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, M.; Schall, K.; Kallis, E.; Eustermann, S.; Guariento, M.; Moldt, M.; Hopfner, K.P.; Michaelis, J. Single-molecule nucleosome remodeling by INO80 and effects of histone tails. FEBS Lett. 2018, 592, 318–331. [Google Scholar] [CrossRef]
- Poyton, M.F.; Feng, X.A.; Ranjan, A.; Lei, Q.; Wang, F.; Zarb, J.S.; Louder, R.K.; Park, G.; Jo, M.H.; Ye, J.; et al. Coordinated DNA and histone dynamics drive accurate histone H2A.Z exchange. Sci. Adv. 2022, 8, eabj5509. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; North, J.A.; Rose, S.D.; Poirier, M.G. Nucleosomes accelerate transcription factor dissociation. Nucleic Acids Res. 2014, 42, 3017–3027. [Google Scholar] [CrossRef]
- Luo, Y.; North, J.A.; Poirier, M.G. Single molecule fluorescence methodologies for investigating transcription factor binding kinetics to nucleosomes and DNA. Methods 2014, 70, 108–118. [Google Scholar] [CrossRef]
- Crickard, J.B.; Lee, J.; Lee, T.H.; Reese, J.C. The elongation factor Spt4/5 regulates RNA polymerase II transcription through the nucleosome. Nucleic Acids Res. 2017, 45, 6362–6374. [Google Scholar] [CrossRef]
- Senavirathne, G.; Mahto, S.K.; Hanne, J.; O’Brian, D.; Fishel, R. Dynamic unwrapping of nucleosomes by HsRAD51 that includes sliding and rotational motion of histone octamers. Nucleic Acids Res. 2016, 45, 685–698. [Google Scholar] [CrossRef]
- Jang, S.; Kang, C.; Yang, H.S.; Jung, T.; Hebert, H.; Chung, K.Y.; Kim, S.J.; Hohng, S.; Song, J.J. Structural basis of recognition and destabilization of the histone H2B ubiquitinated nucleosome by the DOT1L histone H3 Lys79 methyltransferase. Genes Dev. 2019, 33, 620–625. [Google Scholar] [CrossRef] [Green Version]
- Spangler, C.J.; Yadav, S.P.; Li, D.; Geil, C.N.; Smith, C.B.; Wang, G.G.; Lee, T.-H.; McGinty, R.K. DOT1L activity in leukemia cells requires interaction with ubiquitylated H2B that promotes productive nucleosome binding. Cell Rep. 2022, 38, 110369. [Google Scholar] [CrossRef] [PubMed]
- Mivelaz, M.; Cao, A.M.; Kubik, S.; Zencir, S.; Hovius, R.; Boichenko, I.; Stachowicz, A.M.; Kurat, C.F.; Shore, D.; Fierz, B. Chromatin Fiber Invasion and Nucleosome Displacement by the Rap1 Transcription Factor. Mol. Cell 2020, 77, 488–500.e489. [Google Scholar] [CrossRef]
- Huynh, M.T.; Yadav, S.P.; Reese, J.C.; Lee, T.-H. Nucleosome Dynamics during Transcription Elongation. ACS Chem. Biol. 2020, 15, 3133–3142. [Google Scholar] [CrossRef] [PubMed]
- Safaric, B.; Chacin, E.; Scherr, M.J.; Rajappa, L.; Gebhardt, C.; Kurat, C.F.; Cordes, T.; Duderstadt, K.E. The fork protection complex recruits FACT to reorganize nucleosomes during replication. Nucleic Acids Res. 2022, 50, 1317–1334. [Google Scholar] [CrossRef] [PubMed]
- Tomschik, M.; van Holde, K.; Zlatanova, J. Nucleosome dynamics as studied by single-pair fluorescence resonance energy transfer: A reevaluation. J. Fluoresc. 2009, 19, 53–62. [Google Scholar] [CrossRef]
- Bacic, L.; Sabantsev, A.; Deindl, S. Recent advances in single-molecule fluorescence microscopy render structural biology dynamic. Curr. Opin. Struct Biol. 2020, 65, 61–68. [Google Scholar] [CrossRef]
- Gansen, A.; Felekyan, S.; Kühnemuth, R.; Lehmann, K.; Tóth, K.; Seidel, C.A.M.; Langowski, J. High precision FRET studies reveal reversible transitions in nucleosomes between microseconds and minutes. Nat. Commun. 2018, 9, 4628. [Google Scholar] [CrossRef]
- Llères, D.; James, J.; Swift, S.; Norman, D.G.; Lamond, A.I. Quantitative analysis of chromatin compaction in living cells using FLIM-FRET. J. Cell. Biol. 2009, 187, 481–496. [Google Scholar] [CrossRef]
- Torres, T.; Levitus, M. Measuring conformational dynamics: A new FCS-FRET approach. J. Phys. Chem. B 2007, 111, 7392–7400. [Google Scholar] [CrossRef]
- Ngo, T.T.; Zhang, Q.; Zhou, R.; Yodh, J.G.; Ha, T. Asymmetric unwrapping of nucleosomes under tension directed by DNA local flexibility. Cell 2015, 160, 1135–1144. [Google Scholar] [CrossRef] [Green Version]
- Hall, M.A.; Shundrovsky, A.; Bai, L.; Fulbright, R.M.; Lis, J.T.; Wang, M.D. High-resolution dynamic mapping of histone-DNA interactions in a nucleosome. Nat. Struct Mol. Biol. 2009, 16, 124–129. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Bustamante, C. Pulling a single chromatin fiber reveals the forces that maintain its higher-order structure. Proc. Natl. Acad. Sci. USA 2000, 97, 127–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Schematic of FRET. (A) The energy is transferred from a donor fluorophore (green) to acceptor fluorophore (red/pink) in a distance-dependent fashion. (B) The change in efficiency of energy transfer (E) with the distance between the fluorophores. (C) Different cases of energy transfer for more than two fluorophores.
Figure 1.
Schematic of FRET. (A) The energy is transferred from a donor fluorophore (green) to acceptor fluorophore (red/pink) in a distance-dependent fashion. (B) The change in efficiency of energy transfer (E) with the distance between the fluorophores. (C) Different cases of energy transfer for more than two fluorophores.

Figure 2.
(A) Schematic of chromatin structure. (B) Different processes studied with smFRET. Research works focused on these processes have been discussed in detail in the main text. (C) Different cases of nucleosome structural modification resulting in a change in FRET efficiency. The green and red dots represent the donor and the acceptor dye respectively.
Figure 2.
(A) Schematic of chromatin structure. (B) Different processes studied with smFRET. Research works focused on these processes have been discussed in detail in the main text. (C) Different cases of nucleosome structural modification resulting in a change in FRET efficiency. The green and red dots represent the donor and the acceptor dye respectively.

Figure 3.
(A) A labeling scheme to study the conformational dynamics of the nucleosome. While in the native state, the FRET efficiency is expected to be high, whereas in the loosened state, the FRET efficiency decreases owing to the increased distance between two gyres of DNA. (B) A representative FRET trace showing nucleosome dynamics. The green, red and blue lines represent donor intensity, acceptor intensity and FRET efficiency, respectively. The trace shows the transition of nucleosomes between three conformational states corresponding to the high, mid, and low FRET states observed therein. The rate of transition from one state to another can be calculated from the dwell time of each state. (C) The FRET efficiency histograms calculated from the FRET traces (top: high; middle: mid; bottom: low). The area under the curve gives the population of the corresponding FRET state. Evidently, in this case, the mid FRET state is the most populated one. The peak positions denote the FRET efficiency of the maximum number of molecules. Within the scope of the measurement, the authors could not assign any specific FRET state to the native structure. However, by using varied labeling positions, they could conclude that the FRET states represent conformations generated by the gaping motion of the DNA perpendicular to the nucleosome plane. The figures have been adapted from [47] with permission.
Figure 3.
(A) A labeling scheme to study the conformational dynamics of the nucleosome. While in the native state, the FRET efficiency is expected to be high, whereas in the loosened state, the FRET efficiency decreases owing to the increased distance between two gyres of DNA. (B) A representative FRET trace showing nucleosome dynamics. The green, red and blue lines represent donor intensity, acceptor intensity and FRET efficiency, respectively. The trace shows the transition of nucleosomes between three conformational states corresponding to the high, mid, and low FRET states observed therein. The rate of transition from one state to another can be calculated from the dwell time of each state. (C) The FRET efficiency histograms calculated from the FRET traces (top: high; middle: mid; bottom: low). The area under the curve gives the population of the corresponding FRET state. Evidently, in this case, the mid FRET state is the most populated one. The peak positions denote the FRET efficiency of the maximum number of molecules. Within the scope of the measurement, the authors could not assign any specific FRET state to the native structure. However, by using varied labeling positions, they could conclude that the FRET states represent conformations generated by the gaping motion of the DNA perpendicular to the nucleosome plane. The figures have been adapted from [47] with permission.

Figure 4.
Interaction of Chd1 with nucleosome studied with smFRET. (A) The nucleosomal DNA is labeled with Cy3 on the exit side. There are two Cy5 molecules one on each histone H2A(T120C). In the figure, only the proximal Cy5 is shown. (B) The FRET efficiency histogram of the nucleosome (top), nucleosome after the addition of Chd1 without ATP (middle) and nucleosome after addition of Chd1 in the presence of ATP. In the absence of ATP, Chd1 does not influence the FRET efficiency histogram, whereas in the presence of ATP, almost the entire population of the nucleosome shows low FRET efficiency. (C) Representative smFRET traces for each condition mentioned in (B). In the presence of ATP, the stepwise depletion of the high FRET state. Green, red and blue denote donor emission, acceptor emission and FRET efficiency, respectively. (D) FRET efficiency as a function of time in the presence of different concentrations of ATP. Rates of ATP-dependent repositioning were calculated from these variations. These rates are then plotted against the ATP concentration and fitted with the Michaelis–Menten equation to calculate Vmax and KM. (E) smFRET traces at low ATP concentration to show the stepwise decrease in FRET efficiency clearly. Figure is adapted with permission from [66].
Figure 4.
Interaction of Chd1 with nucleosome studied with smFRET. (A) The nucleosomal DNA is labeled with Cy3 on the exit side. There are two Cy5 molecules one on each histone H2A(T120C). In the figure, only the proximal Cy5 is shown. (B) The FRET efficiency histogram of the nucleosome (top), nucleosome after the addition of Chd1 without ATP (middle) and nucleosome after addition of Chd1 in the presence of ATP. In the absence of ATP, Chd1 does not influence the FRET efficiency histogram, whereas in the presence of ATP, almost the entire population of the nucleosome shows low FRET efficiency. (C) Representative smFRET traces for each condition mentioned in (B). In the presence of ATP, the stepwise depletion of the high FRET state. Green, red and blue denote donor emission, acceptor emission and FRET efficiency, respectively. (D) FRET efficiency as a function of time in the presence of different concentrations of ATP. Rates of ATP-dependent repositioning were calculated from these variations. These rates are then plotted against the ATP concentration and fitted with the Michaelis–Menten equation to calculate Vmax and KM. (E) smFRET traces at low ATP concentration to show the stepwise decrease in FRET efficiency clearly. Figure is adapted with permission from [66].

Figure 5.
Combination of smFRET and force measurements to understand the conformational changes of the nucleosome under tension. (A) Scheme: Nucleosome was attached to the microscope via a dsDNA extension. The other end of the DNA was held by an optical trap that applies force. Green and red dots denote the positions of donor and acceptor dye, respectively. Local conformational changes were monitored through FRET between the donor (Cy3) and acceptor (Cy5). (B) Two different views of the labeling positions for the smFRET measurement. Nucleosome structure file: PDB, 3MVD. (C) Representative traces for smFRET measurement and force measurement at a stage speed of 455 nm/s with maximum force set at ~6 pN. Top: smFRET trace; green and red denote the donor and acceptor emissions, respectively. Bottom: overlap of FRET efficiency (blue) and force (black). (D) Overlap of smFRET (blue) and force (black) time traces similar to (C) with a maximum force set at ~20 pN. (E) Average FRET vs. force plot when maximum force was ~6 pN. (F) Average FRET vs. force plot when maximum force was ~20 pN. Figure is adapted from [86] with permission.
Figure 5.
Combination of smFRET and force measurements to understand the conformational changes of the nucleosome under tension. (A) Scheme: Nucleosome was attached to the microscope via a dsDNA extension. The other end of the DNA was held by an optical trap that applies force. Green and red dots denote the positions of donor and acceptor dye, respectively. Local conformational changes were monitored through FRET between the donor (Cy3) and acceptor (Cy5). (B) Two different views of the labeling positions for the smFRET measurement. Nucleosome structure file: PDB, 3MVD. (C) Representative traces for smFRET measurement and force measurement at a stage speed of 455 nm/s with maximum force set at ~6 pN. Top: smFRET trace; green and red denote the donor and acceptor emissions, respectively. Bottom: overlap of FRET efficiency (blue) and force (black). (D) Overlap of smFRET (blue) and force (black) time traces similar to (C) with a maximum force set at ~20 pN. (E) Average FRET vs. force plot when maximum force was ~6 pN. (F) Average FRET vs. force plot when maximum force was ~20 pN. Figure is adapted from [86] with permission.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Summary of research works focused on the application of smFRET to understand the structure, dynamics and behavior of nucleosomes.
Table 1.
Summary of research works focused on the application of smFRET to understand the structure, dynamics and behavior of nucleosomes.
| Topic | Subtopic | Details | Reference |
|---|---|---|---|
| Chromatin structure and dynamics | Native state | Spontaneous confirmational dynamics between closed and open state | [47,48] |
| Conformational dynamics between three states: closed, partially open, and open | [49] | ||
| Equilibrium constants of DNA unwrapping | [50] | ||
| Conformational switching time is different for different DNA ends | [51] | ||
| Stepwise spontaneous DNA unwrapping resulting in histone eviction | [52] | ||
| Effect of salt | Increasing salt concentration triggers DNA unwrapping | [49] | |
| Salt-induced DNA unwrapping is reversible | [51] | ||
| Nucleosomes containing acetylated histones reflect additional conformational states as compared to native | [53] | ||
| DNA needs to unwrap before dimer eviction | [54] | ||
| Higher-order nucleosome structure | Transient formation of tetrameric structure, interaction of HP1α with methylated histone tails | [55] | |
| Motion of H3 N-terminal tail | [56] | ||
| Effect of DNA modification | CpG-methylation assists nucleosome assembly, formation of stable tetrasomes | [57] | |
| Effect of histone post translational modification | Acetylation results in increased nucleosome breathing | [58] | |
| Succinylation decreases nucleosome stability | [59] | ||
| Ubiquitylation destabilizes nucleosome, facilitates dinucleosome formation | [60] | ||
| Effect of H1 linker histone | H1 influences higher-order nucleosome structure | [61] | |
| Interaction of nucleosome with protein machineries | Interaction with remodelers | Nucleosome remodeling comprises two steps: ACF binding and ACF translocation | [62] |
| Stepwise ISWF-directed translocation of nucleosome | [63] | ||
| Entry and exit side time lag of nucleosomeal DNA translocation during Chd1- and SNF2h-induced remodeling | [64] | ||
| SWI/SNF directed remodeling is bidirectional and sequence independent, and histone octamer is retained with DNA throughout the process | [65] | ||
| Stable remodeled nucleosomes are generated via unstable intermediates for Chd1 controlled remodeling | [66] | ||
| Nucleosomal acidic patch plays crucial role in ISWI and INO80 controlled remodeling. | [67] | ||
| DNA sliding by INO80 depends on the length of flanking DNA | [68] | ||
| Directionality of nucleosome sliding is governed by the coordination of ATP hydrolyzing motors of two units of the dimeric remodeler. | [69] | ||
| Effect of histone tails on the remodeling process | [70] | ||
| Histone dimer exchange happens in seconds timescale. | [71] | ||
| Interaction with other protein machineries | Transcription factors binds to duplex DNA more effectively than mononucleosome or nucleosome array. | [72] | |
| Single-molecule measurements can accurately measure the binding constant for the interaction between the nucleosome and transcription factors | [73] | ||
| Spt4/5 helps polymerase to escape the pause during transcription and facilitates DNA rewrapping behind it | [74] | ||
| Rad51-mediated unwrapping of nucleosomal DNA happens stepwise | [75] | ||
| Dot1L binding destabilizes nucleosomes | [76] | ||
| Dot1L interacts directly with ubiquitin on H3K120, which leads to an effective acidic patch binding | [77] | ||
| Rap1 loosens up chromatin fiber by transient interaction with it | [78] | ||
| Acetylation at K56 of H3 greatly reduces the Pol II pause duration near the entry site of the nucleosome | [79] | ||
| FACT activity is highly dependent on the middle and C terminal domains of the subunits Pob3 and Spt6 | [80] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Sengupta, B.; Huynh, M. Contribution of smFRET to Chromatin Research. Biophysica 2023, 3, 93-108. https://doi.org/10.3390/biophysica3010007
AMA Style
Sengupta B, Huynh M. Contribution of smFRET to Chromatin Research. Biophysica. 2023; 3(1):93-108. https://doi.org/10.3390/biophysica3010007
Chicago/Turabian StyleSengupta, Bhaswati, and Mai Huynh. 2023. "Contribution of smFRET to Chromatin Research" Biophysica 3, no. 1: 93-108. https://doi.org/10.3390/biophysica3010007