
Current Opinion on the Use of c-Fos in Neuroscience
 , ,
, , 
 ,
,  , ,
, ,  ,
,
Abstract
:1. Introduction
2. Methodology: Selection Criteria
- (a).
- Records identified in GoogleScholar (n = 3000).
- (b).
- Records identified in SciELO and Pubmed (n = 245).
- (c).
- Total papers (n = 3245).
- (d).
- Papers excluded based on title/being duplicates/being deleted (n = 2888).
- (e).
- Papers selected to assess their eligibility (n = 357).
- (f).
- Papers included in this review (n = 82).
3. Development
3.1. The Expression of Immediate Early Genes (IEGs)
3.2. Fos
3.2.1. c-Fos Gene
3.2.2. Fos Protein
3.2.3. AP-1, MAPK, and c-Fos Activation

3.2.4. c-Fos Degradation
3.3. Fos Mapping outside the Central Nervous System
3.4. Fos Activation in Glial Cells
3.5. Stimuli That Trigger Fos Expression in Mammalian Animal Models
3.6. Stimuli That Trigger Fos Expression in Non-Mammalian Animal Models
4. Perspectives and Suggestions
5. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cruz, F.; Jauregui, F.; Aguilar, A.; García, J.; Luquin, S. Immediate Early Gene c-fos in the Brain: Focus on Glial Cells. Brain Sci. 2022, 12, 687. [Google Scholar] [CrossRef] [PubMed]
- Herrera, D.; Robertson, H. Activation of c-fos in the brain. Prog. Neurobiol. 1996, 50, 83–107. [Google Scholar] [CrossRef] [PubMed]
- Saha, R.; Dudek, S. Splitting Hares and Tortoises: A classification of neuronal immediate early gene transcription based on poised RNA polymerase II. Neuroscience 2013, 247, 175–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, L. A Brief Introduction to the Transduction of Neural Activity into Fos Signal. Dev. Reprod. 2015, 19, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Minatohara, K.; Akiyoshi, M.; Okuno, H. Role of immediate-early genes in synaptic plasticity and neuronal ensembles underlying the memory trace. Front. Mol. Neurosci. 2016, 8, 78. [Google Scholar] [CrossRef] [Green Version]
- Xiao, Q.; Castillo, S.; Nikodem, V. Distribution of messenger RNAs for the orphan nuclear receptors Nurr1 and Nur77 (NGFI-B) in adult rat brain using in situ hybridization. Neuroscience 1996, 75, 221–230. [Google Scholar] [CrossRef]
- Hughes, P.; Lawlor, P.; Dragunow, M. Basal expression of Fos, Fos-related, Jun, and Krox 24 proteins in rat hippocampus. Mol. Brain Res. 1992, 13, 355–357. [Google Scholar] [CrossRef]
- Zhou, L.; Xue, C.; Chen, C.; Jiang, W.; He, S.; Zhang, X. c-Fos is a mechanosensor that regulates inflammatory responses and lung barrier dysfunction during ventilator-induced acute lung injury. BMC Pulm. Med. 2022, 22, 9. [Google Scholar] [CrossRef]
- Inuzuka, H.; Nanbu, R.; Masuho, Y.; Muramatsu, M.; Tojo, H.; Wakao, H. Differential regulation of immediate early gene expression in preadipocyte cells through multiple signaling pathways. Biochem. Biophys. Res. Commun. 1999, 265, 664–668. [Google Scholar] [CrossRef]
- Lin, J.; Kornhauser, J.; Singh, N.; Mayo, K.; Takahashi, J. Visual sensitivities of nur77 (NGFI-B) and zif268 (NGFI-A) induction in the suprachiasmatic nucleus are dissociated from c-fos induction and behavioral phase-shifting responses. Mol. Brain Res. 1997, 46, 303–310. [Google Scholar] [CrossRef]
- Ryseck, R.; Macdonald, H.; Mattéi, M.; Ruppert, S.; Bravo, R. Structure, mapping and expression of a growth factor inducible gene encoding a putative nuclear hormonal binding receptor. EMBO J. 1989, 11, 3327–3335. [Google Scholar] [CrossRef] [PubMed]
- Farioli, S.; Ceccarelli, M.; Saraulli, D.; Micheli, L.; Cannas, S.; D’Alessandro, F.; Scardigli, R.; Leonardi, L.; Cinà, I.; Costanzi, M.; et al. Tis21 is required for adult neurogenesis in the subventricular zone and for olfactory behavior regulating cyclins, BMP4, Hes1/5, and Ids. Front. Cell Neurosci. 2014, 8, 98. [Google Scholar] [CrossRef]
- Resovia, A.; Pinessia, D.; Chiorino, G.; Giulia, T. Current understanding of the thrombospondin-1 interactome. Matrix Biol. 2014, 37, 83–91. [Google Scholar] [CrossRef]
- Bozon, B.; Davis, S.; Laroche, S. Regulated transcription of the immediate-early gene Zif268: Mechanisms and gene dosage-dependent function in synaptic plasticity and memory formation. Hippocampus 2002, 12, 570–577. [Google Scholar] [CrossRef]
- Hyeon, J.; Chul, M.; Ha, J.; Hye, I.; Lee, J.; Chun, B.; Koo, I.; Myung, S.; Yun, J.; Geun, T.; et al. Increased immunoreactivity of c-Fos in the spinal cord of the aged mouse and dog. Mol. Med. Rep. 2015, 11, 1043–1048. [Google Scholar] [CrossRef]
- Morgan, J.; Curran, T. Stimulus-transcription coupling in the nervous system: Involvement of the inducible proto-oncogenes fos and jun. Annu. Rev. Neurosci. 1991, 14, 421–451. [Google Scholar] [CrossRef] [PubMed]
- Perrin, A.; Jeton, F.; Pichon, A.; Frugière, A.; Richalet, J.; Bodineau, L.; Voituron, N. The c-FOS protein immunohistological detection: A useful tool as a marker of central pathways involved in specific physiological responses in vivo and ex vivo. J. Vis. Exp. 2016, 2016, e53613. [Google Scholar] [CrossRef] [Green Version]
- Meza, J.; Montaño, A.; Aguayo, A. Las Bases Moleculares del Cáncer. Acta Univ. 2006, 58, 56–70. [Google Scholar]
- Pang, Y.; Kiba, H.; Jayaraman, A. Acute nicotine injections induce c-fos mostly in non-dopaminergic neurons of the midbrain of the rat. Mol. Brain Res. 1993, 20, 162–170. [Google Scholar] [CrossRef]
- Dragunow, M.; Robertson, H. Brain injury induces c-fos protein(s) in nerve and glial-like cells in adult mammalian brain. Brain Res. 1988, 455, 295–299. [Google Scholar] [CrossRef]
- Hai, T.; Curran, T. Cross-family dimerization of transcription factors Fos/Jun and ATF/CREB alters DNA binding specificity. Proc. Natl. Acad. Sci. USA 1991, 88, 3720–3724. [Google Scholar] [CrossRef] [PubMed]
- Seldeen, K.; Deegan, B.; Bhat, V.; Mikles, D.; McDonald, C.; Farooq, A. Energetic coupling along an allosteric communication channel drives the binding of Jun-Fos heterodimeric transcription factor to DNA. FEBS J. 2011, 278, 2090–2104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsujino, T.; Sano, H.; Kubota, Y.; Hsieh, S.; Miyajima, T.; Saito, K.; Nakajima, M.; Saito, N.; Yokoyama, M. Expression of Fos-like immunoreactivity by yohimbine and clonidine in the rat brain. Eur. J. Pharmacol. Mol. Pharmacol. 1992, 226, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Onodera, H.; Kogure, K.; Ono, Y.; Igarashi, K.; Kiyota, Y.; Nagaoka, A. Proto-oncogene c-fos is transiently induced in the rat cerebral cortex after forebrain ischemia. Neurosci. Lett. 1989, 98, 101–104. [Google Scholar] [CrossRef] [PubMed]
- Curran, T.; Miller, A.; Zokas, L.; Verma, I. Viral and cellular fos proteins: A comparative analysis. Cell 1984, 36, 259–268. [Google Scholar] [CrossRef]
- Rahmsdorf, H.; Schönthal, A.; Angel, P.; Litfin, M.; Rüther, U.; Herrlich, P. Posttranscriptional regulation of c-fos mRNA expression. Nucleic Acids Res. 1987, 15, 1643–1659. [Google Scholar] [CrossRef] [Green Version]
- Herdegen, T.; Kovary, K.; Buhl, A.; Bravo, R.; Zimmermann, M.; Gass, P. Basal expression of the inducible transcription factors c-Jun, JunB, JunD, c-Fos, FosB, and Krox-24 in the adult rat brain. J. Comp. Neurol. 1995, 354, 39–56. [Google Scholar] [CrossRef]
- Appleyard, S. Lighting Up Neuronal Pathways: The Development of a Novel Transgenic Rat that Identifies Fos-Activated Neurons Using a Red Fluorescent Protein. Endocrinology 2009, 150, 5199–5201. [Google Scholar] [CrossRef] [Green Version]
- Jackson, M.; Papa, S.; Bola, J.; Bruckdorfer, R.; Carlsen, H.; Elliott, R.; Flier, J.; Griffiths, H.; Heales, S.; Holst, B.; et al. Antioxidants, reactive oxygen and nitrogen species. Mol. Asp. Med. 2002, 23, 209–285. [Google Scholar] [CrossRef]
- Groves, A.; Kihara, Y.; Jonnalagadda, D.; Rivera, R.; Kennedy, G.; Mayford, M.; Chun, J. A functionally defined in vivo astrocyte population identified by c-Fos activation in a mouse model of multiple sclerosis modulated by S1P signaling: Immediate-early astrocytes (ieAstrocytes). eNeuro 2018, 5, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Barros, V.; Mundim, M.; Galindo, L.; Bittencourt, S.; Porcionatto, M.; Mello, L. The pattern of c-Fos expression and its refractory period in the brain of rats and monkeys. Front. Cell Neurosci. 2015, 9, 72. [Google Scholar] [CrossRef]
- Scammell, T.; Price, K.; Sagar, S. Hyperthermia induces c-fos expression in the preoptic area. Brain Res. 1993, 618, 303–307. [Google Scholar] [CrossRef]
- Yokoyama, K.; Hiyama, A.; Arai, F.; Nukaga, T.; Sakai, D.; Mochida, J. C-Fos regulation by the MAPK and PKC pathways in intervertebral disc cells. PLoS ONE 2013, 8, e73210. [Google Scholar] [CrossRef] [Green Version]
- Del-Bel, E.; Borges, C.; Delfino, H.; Guimaräes, F. Induction of Fos protein immunoreactivity by spinal cord contusion. Braz. J. Med. Biol. Res. 2000, 33, 521–528. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Zheng, Z.; Ping, K.; Qian, Z. Effects of spinal cord injury on c-fos expression in hypothalamic paraventricular nucleus and supraoptic nucleus in rats. Brain Res. 2006, 1087, 175–179. [Google Scholar] [CrossRef]
- Hu, V.; Black, G.; Torres, A.; Abramson, F. 3H-thymidine is a defective tool with which to measure rates of DNA synthesis. FASEB J. 2002, 16, 1456–1457. [Google Scholar] [CrossRef]
- Brennan, A.; Leech, J.; Kad, N.; Mason, J. Selective antagonism of cJun for cancer therapy. J. Exp. Clin. Cancer Res. 2020, 39, 184. [Google Scholar] [CrossRef] [PubMed]
- Gustems, M.; Woellmer, A.; Rothbauer, U.; Eck, S.; Wieland, T.; Lutter, D.; Hammerschmidt, W. C-Jun/c-Fos heterodimers regulate cellular genes via a newly identified class of methylated DNA sequence motifs. Nucleic Acids Res. 2014, 42, 3059–3072. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.; Kikuchi, K.; Kondo, M. Activation of mitogen-activated protein kinase kinase (MEK)/extracellular signal regulated kinase (ERK) signaling pathway is involved in myeloid lineage commitment. Blood 2007, 110, 1420–1428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roux, P.; Blenis, J. ERK and p38 MAPK-Activated Protein Kinases: A Family of Protein Kinases with Diverse Biological Functions. Microbiol. Mol. Biol. Rev. 2004, 68, 320–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hudson, A. Genetic reporters of neuronal activity: C-Fos and G-CaMP6. Methods Enzymol. 2018, 603, 197. [Google Scholar] [CrossRef] [PubMed]
- Yao, W.; Wu, C. Distinct roles of CaMKII and PKA in regulation of firing patterns and K(+) currents in Drosophila neurons. J. Neurophysiol. 2001, 85, 1384–1394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gille, H.; Strahl, T.; Shaw, P. Activation of ternary complex factor Elk-1 by stress-activated protein kinases. Curr. Biol. 1995, 5, 1191–1200. [Google Scholar] [CrossRef] [Green Version]
- Hanlon, M.; Bundy, L.; Sealy, L. C/EBPBeta and Elk-1 synergistically transactivate the c-fos serum response element. BMC Cell Biol. 2000, 1, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vamosi, G.; Baudendistel, N.; Von Der Lieth, C.; Szalóki, N.; Mocsár, G.; Müller, G.; Brázda, P.; Waldeck, W.; Damjanovich, S.; Langowski, J.; et al. Conformation of the c-Fos/c-Jun complex in vivo: A combined FRET, FCCS, and MD-modeling study. Biophys. J. 2008, 94, 2859–2868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stancovski, I.; Gonen, H.; Orian, A.; Schwartz, A.L.; Ciechanover, A. Degradation of the proto-oncogene product c-Fos by the ubiquitin proteolytic system in vivo and in vitro: Identification and characterization of the conjugating enzymes. Mol. Cell Biol. 1995, 15, 7106–7116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perrin, F.; Najib, H. Serotonergic mechanisms in spinal cord injury. Exp. Neurol. 2019, 318, 174–191. [Google Scholar] [CrossRef]
- Manzo, J.; Miquel, M.; Toledo, R.; Mayor, J.; Garcia, L.; Aranda, G.; Caba, M.; Elena, M. Fos expression at the cerebellum following non-contact arousal and mating behavior in male rats. Physiol. Behav. 2008, 93, 357–363. [Google Scholar] [CrossRef] [Green Version]
- Kim, O.; Kim, D.; Lee, M.; Park, S.; Yamada, W.; Eun, S.; Lee, J. Standardized Edible Bird’s Nest Extract Prevents UVB Irradiation-Mediated Oxidative Stress and Photoaging in the Skin. Antioxidants 2021, 9, 1452. [Google Scholar] [CrossRef]
- Edling, Y.; Ingelman, M.; Simi, A. Glutamate Activates c-fos in Glial Cells via a Novel Mechanism Involving the Glutamate Receptor Subtype mGlu5 and the Transcriptional Repressor DREAM. Glia 2007, 55, 328–340. [Google Scholar] [CrossRef]
- Wu, X.; Yoo, S.; Wrathall, J. Real-time quantitative PCR analysis of temporal-spatial alterations in gene expression after spinal cord contusion. J. Neurochem. 2005, 93, 943–952. [Google Scholar] [CrossRef] [PubMed]
- Holstein, G.; Friedrich, V.; Martinelli, G. Glutamate and GABA in vestibulo-sympathetic pathway neurons. Front. Neuroanat. 2016, 10, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paul, M.; Graybiel, A.; David, J.; Robertson, H. D1-like and D2-like dopamine receptors synergistically activate rotation and c-fos expression in the dopamine-depleted striatum in a rat model of Parkinson’s disease. J. Neurosci. 1992, 12, 3729–3742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richard, D.; Rivest, S.; Rivier, C. The 5-hydroxytryptamine agonist fenfluramine increases fos-like immunoreactivity in the brain. Brain Res. 2002, 594, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Chang, E. Expresión de C-Fos En El Cerebelo Después de La Lesión Espinal de La Rata Macho. Ph.D. Thesis, Universidad Veracruzana, Xalapa, Mexico, 2013. [Google Scholar]
- Haltera, B.; Ildari, N.; Cline, M.; Gilbert, E. Ferulic acid, a phytochemical with transient anorexigenic effects in birds. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2021, 259, 111015. [Google Scholar] [CrossRef]
- Yang, M.; Wu, D.; Cheng, S.; Dong, Y.; Wu, C.; Wanga, Z.; Du, M. Inhibitory effects of Atlantic cod (Gadus morhua) peptides on RANKL-induced osteoclastogenesis in vitro and osteoporosis in ovariectomized mice. Food Funct. 2022, 13, 1975–1988. [Google Scholar] [CrossRef]
- Alsrhani, A.; Raman, R.; Jagadeeswaran, P. Trypsin induces an aversive response in zebrafish by PAR2 activation in keratinocytes. PLoS ONE 2021, 10, e0257774. [Google Scholar] [CrossRef]
- Dos Santos, R. Imunorreatividade de C-Fos Em Áreas Telencefálicas Do Lagarto Tropidurus Hispidus (SPIX, 1825) Submetido a Um Estímulo Aversivo. Bachelor’s Thesis, Universidade Federal de Sergipe, San Cristobal, Brazil, 2021. [Google Scholar]

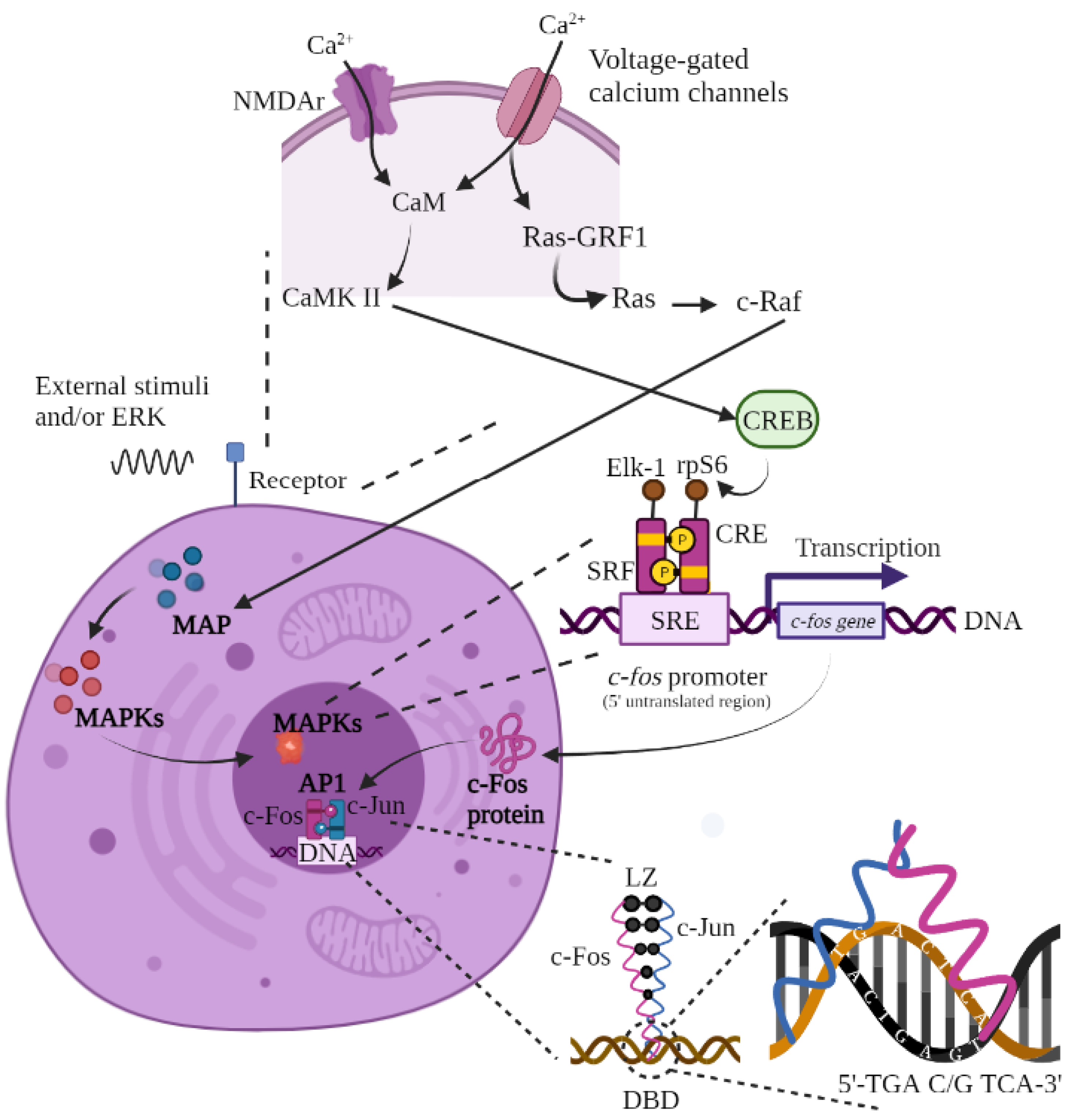{kind=link}
| IEGs | Function |
|---|---|
| Arc (Arg3.1) | It regulates specific neurons associated with learning and memory. These functions are also associated with c-fos and egr-1. |
| c-fos c-jun | Cell activation (cell proliferation and differentiation) and protein transcriptions. For c-fos: sensory information processing, pain regulation mechanisms, long-term potentiation, neural plasticity, learning and memory, direct control of the expression of inflammatory cytokines, mediation of neuronal excitability by regulating the expression of the kainic acid receptor and GluR6, brain-derived neurotrophic factor. |
| cyr 61 | Cell adhesion, migration, and angiogenesis. |
| egr-1 (tis 8) | Functioning and development of the central nervous system, as well as the development of prostate cancer, thymic T cells. Determines the fate of hematopoietic cells. |
| erp or MKP-1 (dual-specificity MAPK phosphatase) | Dephosphorylation of MAPs, essential for the regulation of cell growth. |
| Homer1a | Long-term potentiation, memory consolidation processes. and modification of glutamatergic signaling pathways. |
| krox-24 | Long-term potentiation, neural plasticity, learning and memory. |
| c-myc | Normal cell development, cell malignancy, cell proliferation and apoptosis. |
| nur1 nur77 (NGFI-B) | Vision (dorsal lateral geniculate nuclei and visual cortex), sensory information processing (primary olfactory cortex and anterior olfactory nuclei), pain regulation mechanisms (greater raphe nuclei), auditory stimulation and stress mechanisms (facial, spinal trigeminal, medial vestibular, lateral vestibular, and dorsal cochlear nuclei). |
| N-10 (thyroid hormone receptor) | Cell differentiation and proliferation. |
| pip92, Chx1 or ETR101 | Neuronal differentiation and cell death. |
| tis10 (cox-2) | Formation of PG-G2 from arachidonic acid and two oxygen molecules, which is then reduced to PGH2 and, in turn, acts as a precursor to several eicosanoids, including PGE2, PGF2α, PGD2, prostacyclin, and thromboxane A2. |
| tis11 | Gene expression at the transcriptional level, promotes destabilization of cytokine mRNAs and promotes ARE (adenylate-uridylate-rich elements)-induced mRNA decay and decay. |
| Glutamate receptor 6 (GluR6), mitogen-activated proteins (MAPs), PG (prostaglandin) [3,4,5,6,7,8,9,10] | |
| tis21 | Proliferation and differentiation of neural stem and progenitor cells (for example, in the cerebellum, hippocampus, or dentate gyrus). |
| TSP-1 (thrombospondin-1) | Activation of receptors and signaling pathways, formation of multimolecular complexes, sequestration and inactivation of growth factors and enzymes, alterations in protein localization, proteolytic processing and internalization, and effects on receptor/ligand balance and downstream signaling (depending on their ability to bind to different ligands). |
| Stra13 zf9 | Early phase of preadipocyte cell differentiation. |
| zif268 (NGFI-A) | Synaptic plasticity and long-term memory. |
| [8,9,11,12,13] | |
| IEGs | Stimulus | Animal Model |
|---|---|---|
| Arc (Arg3.1) | Learning and memory | Rats |
| c-fos c-jun c-myc | Seizures Cell differentiation Neural arousal Electrical stimulation Surgical injuries/nerve transections Devascularization Cerebral ischemia Abstinence Nociceptive and peripheral stimulation Heat stress Light stimulation Old age | Mice Rats Cats Monkeys Humans Dogs |
| egr-1 (tis 8) | Phorbol esters Growth factors (e.g., NGF, EGF) Cellular differentiation (cardiac and neuronal) Electrical stimulation | Drosophila fruit fly Rats Mice |
| egr-2 (krox20) cyr 61 nur77 (NGFI-B) | Cell differentiation Growth factors | Developing mice Rats |
| krox-24 N-10 zif268 (NGFI-A) | Phorbol esters Growth factors (e.g., NGF) Long-term object recognition and memory tasks (24 h) | Drosophila fruit fly Mice |
| [4,10,13,14,15,16] | ||
| Basal Expression * | After Stimuli Expression ** | |
|---|---|---|
| mRNA transcription | ≈first 5 min | ≈first 15 min |
| mRNA increase | ≈15–20 min | ≈30–60 min |
| mRNA peak | ≈30 min | ≈30–45 min |
| mRNA half-life (metabolization) | ≈180 min | ≈10–20 min |
| c-Fos transcription | --- | ≈20–90 min |
| c-Fos increase | --- | Depends on type of stimuli |
| c-Fos peak | --- | Depends on type of stimuli |
| c-Fos half-life (metabolization) | --- | ≈2–5 h |
| [15,16,26,27] | ||
| Stimuli | Model |
|---|---|
| Neurotransmitters | |
| Glutamate depolarization (NMDA and AMPA) | Rats/blood pressure and posture |
| Dopamine (D1) | Rats/Parkinson’s |
| Noradrenaline and adrenaline (α2) | Rats/neuroplasticity |
| Acetylcholine (nicotinics) | Rats/visual stimuli |
| Serotonine (5-HT1c and 5-HT2) | Rats/obesity |
| Physiological mechanisms | |
| Alzheimer’s | Rats, humans |
| Mechanical brain injuries | Rats (neurons: 1, 6, 12, and 72 h later; glia: 12, 24, and 72 h later *) |
| Ischemia | Mongolian gerbils, rats (15–60 min, 3 days *) |
| Heatstroke | Rats, rabbits |
| Seizures | Rats (30–60 min, 3–4 h *), marmosets (6 h *) |
| Learning and memory | Rats, mice |
| Osmotic stimulation | Rats (30–60 min, 180 min, 1–2 h, 4 h *) |
| Stress | Rats |
| Cardiac rhythms | Rats, hamsters |
| Alcohol intake | Mice, rats (2–4 h, 8 h *) |
| Depression | Mice (7 d) |
| Old age (neuropathic pain) | Beagles, mice |
| [1,14,19,23,34,52,53,54] | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lara Aparicio, S.Y.; Laureani Fierro, Á.d.J.; Aranda Abreu, G.E.; Toledo Cárdenas, R.; García Hernández, L.I.; Coria Ávila, G.A.; Rojas Durán, F.; Aguilar, M.E.H.; Manzo Denes, J.; Chi-Castañeda, L.D.; et al. Current Opinion on the Use of c-Fos in Neuroscience. NeuroSci 2022, 3, 687-702. https://doi.org/10.3390/neurosci3040050
Lara Aparicio SY, Laureani Fierro ÁdJ, Aranda Abreu GE, Toledo Cárdenas R, García Hernández LI, Coria Ávila GA, Rojas Durán F, Aguilar MEH, Manzo Denes J, Chi-Castañeda LD, et al. Current Opinion on the Use of c-Fos in Neuroscience. NeuroSci. 2022; 3(4):687-702. https://doi.org/10.3390/neurosci3040050
Chicago/Turabian StyleLara Aparicio, Sandra Yasbeth, Ángel de Jesús Laureani Fierro, Gonzalo Emiliano Aranda Abreu, Rebeca Toledo Cárdenas, Luis Isauro García Hernández, Genaro Alfonso Coria Ávila, Fausto Rojas Durán, María Elena Hernández Aguilar, Jorge Manzo Denes, Lizbeth Donají Chi-Castañeda, and et al. 2022. "Current Opinion on the Use of c-Fos in Neuroscience" NeuroSci 3, no. 4: 687-702. https://doi.org/10.3390/neurosci3040050