
Roles of Two F-Box Proteins: FBXL14 in the Periosteum and FBXW2 at Elastic Fibers

Department of Biomaterials, Osaka Dental University, 8-1, Kuzuhahanazono-cho, Hirakata 573-1121, Osaka, Japan
Osteology 2023, 3(1), 1-10; https://doi.org/10.3390/osteology3010001
Submission received: 26 October 2022
/
Revised: 28 November 2022
/
Accepted: 3 January 2023
/
Published: 5 January 2023
Abstract
:I previously reported that F-box/leucine-rich repeat protein 14 (FBXL14) expressed in periosteum-derived cells, and F-box and WD-40 domain-containing protein 2 (FBXW2) in the periosteum form a fiber-like structure. Here, two culture medium conditions, that is, media with and without ascorbic acid, were compared during explant culture. In the absence of ascorbic acid, the expression patterns of osteocalcin, FBXW2, and elastin were compared using fluorescent immunostaining during weeks 3–5. By observing the periosteum, cambium layer and bone, I demonstrated FBXL14 expression in micro-vessels and bone lacuna. Fluorescent immunostaining revealed that, without ascorbic acid, the FBXL14 layer was thin. Conversely, in the presence of ascorbic acid, FBXL14 formed a thick membrane-like structure inside the periosteum, and the multilayer of periosteum-derived cells (PDCs) was strong. The expression patterns of osteocalcin and FBXW2 were similar. Elastin retained its fiber structure for up to five weeks. Although osteocalcin and FBXW2 were expressed in regions similar to elastin, they could not retain their fiber structures. In conclusion, FBXL14 appears to play a role in preparing a native scaffold for forming a multilayered sheet of PDCs inside the periosteum. FBXW2 and osteocalcin appear to separate from elastic fibers during calcification.
1. Introduction
F-box/leucine-rich repeat protein 14 (FBXL14) contains the E3 ubiquitin ligase Skp, Cullin and F-box (SCF) complex for proteolysis [1,2,3,4]. Recently, it was reported that FBXL14 is associated with lung cancer [5], breast cancer [6] and distraction osteogenesis [7]. FBXL14 is reportedly a regulator of epithelial–mesenchymal transition pathway [8]. Mason and Laman [9] reviewed three-dimensional structures of the FBXL family members, but they described only structure prediction of FBXL14. In 2006, Akiyama et al. [10] investigated bovine periosteum-derived cells (PDCs) for bone regeneration and reported that FBXL14 is expressed in periosteum-derived cells [11]. In 2009, Akiyama and Nakamura [12] compared two Culture Medium 199 conditions, that is, media with and without ascorbic acid [12]. They found that PDCs without ascorbic acid could not form a multilayered cell sheet and that bone regeneration failed. To form a three-dimensional sheet of PDCs in vitro, supportive proteins as a natural scaffold are needed. In the presence of ascorbic acid, collagen fibers support PDCs [12]. In the present study, I sought for non-collagenous supportive proteins that function only with ascorbic acid. Unlike FBXL14, F-box and WD-40 domain-containing protein 2 (FBXW2) is reportedly expressed in the periosteum but not in PDCs [11]. FBXW2 regulates cancer stemness and is important for cancer therapy [13]. In the periosteum, FBXW2 and osteocalcin have been found to localize to the same fiber structures [14], but the relationship between FBXW2 and osteocalcin calcification remains unknown. The aim of this study was to examine the role of the two F-box proteins, FBXL14 and FBXW2. Formation of a multilayered cell sheet of PDCs resulted in the scaffold-free bone regeneration [10], and FBXL14 may be related to this mechanism. Additionally, the osteogenic differentiation of PDCs induced by FBXW2 and osteocalcin was investigated.
2. Materials and Methods
2.1. Cell Culture and Sample Preparation
Periosteum for explant culture and bones with periosteum from four hind and front legs of 30-month-old Japanese Black Cattle (Kobe Chuo Chikusan, Kobe, Japan) were randomly sampled from a slaughterhouse. This study was performed within 24 h of slaughtering the cattle. All protocols adhered to the Osaka Dental University Regulations on Animal Care and Use (Approval No. 22-01002). No live animals were used in this study. Periosteum from eight cattle was used in explant culture for up to five weeks in Medium 199 (12340-030; Gibco, Grand Island, NY, USA) with 10% fetal bovine serum and penicillin/streptomycin (168-23191; FUJIFILM Wako Pure Chemical Corporation, Osaka, Japan), as previously described [15]. Periosteum from four cattle was cultured with ascorbic acid, and that of the other four cattle was cultured without ascorbic acid. To prepare paraffin-embedded tissue blocks and sections, the periosteum after explant culture and bone with periosteum were fixed in 4% paraformaldehyde.
2.2. Fluorescent Immunostaining
Fluorescent immunostaining for FBXL14, von Willebrand factor, elastin, osteocalcin and FBXW2 in bovine periosteum-derived cells cultured with or without ascorbic acid was performed.
Table 1 shows the conditions for antigen retrieval, primary and secondary antibodies, manufacturers and culture medium for each target protein. Proteinase K (Dako Cytomation, Glostrup, Denmark) was used for antigen retrieval. The primary antibodies used were as follows: anti-FBXL14, anti-von Willebrand factor, anti-elastin, anti-osteocalcin and anti-FBXW2. The secondary antibodies used were as follows: Alexa Fluor 594 anti-rabbit IgG (H+L), Alexa Fluor™ 488 anti-mouse IgG (H+L) and anti-goat IgG-CFL 594. Additionally, 4′,6-diamidino-2-phenylindole (DAPI) was used for counterstaining. The microscope (BZ-9000; Keyence Japan, Osaka, Japan), BZ-II Viewer software (Version 1.1, Keyence Japan) and BZ-II Analyzer software (Version 1.1; Keyence Japan) were used for visualization and photography.
3. Results
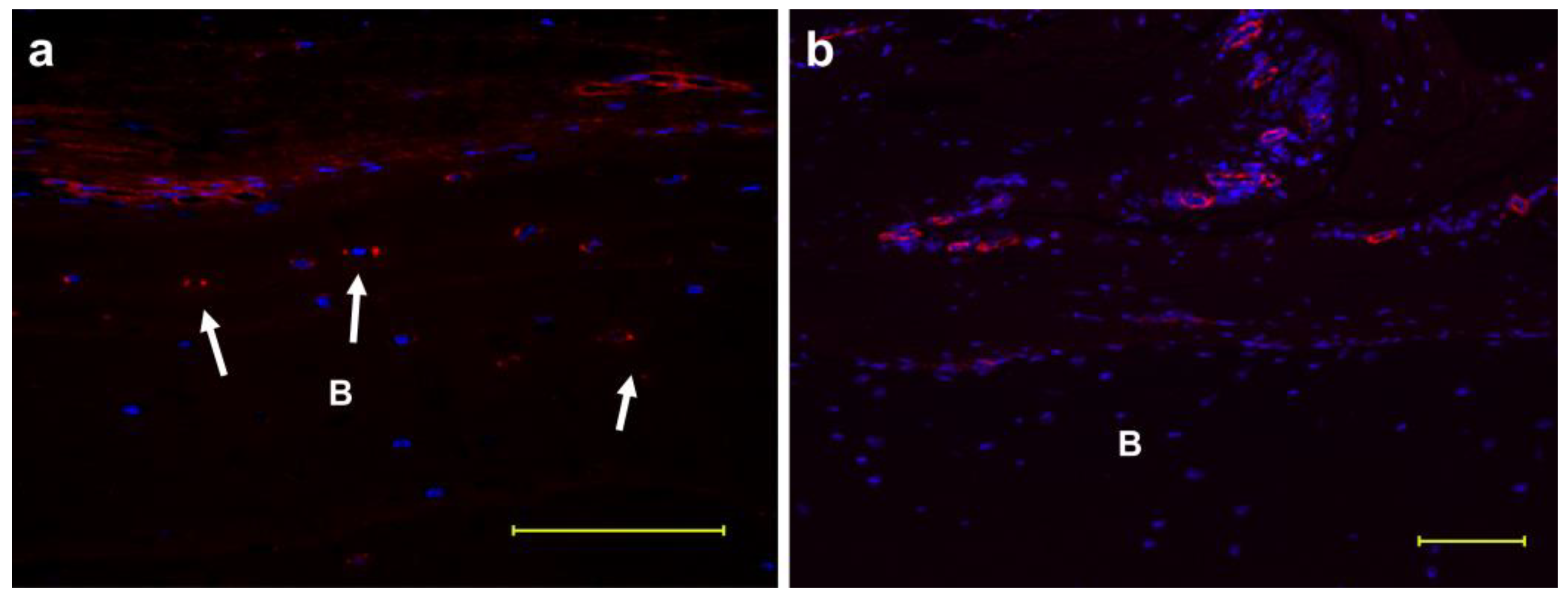
Figure 1 shows the fluorescent immunostaining of the bone with periosteum. von Willebrand factor, which indicates the presence of endothelial cells, and FBXL14 were expressed around micro-vessels (Figure 1a,b). FBXL14 was also expressed in bone lacunae (Figure 1, arrow).
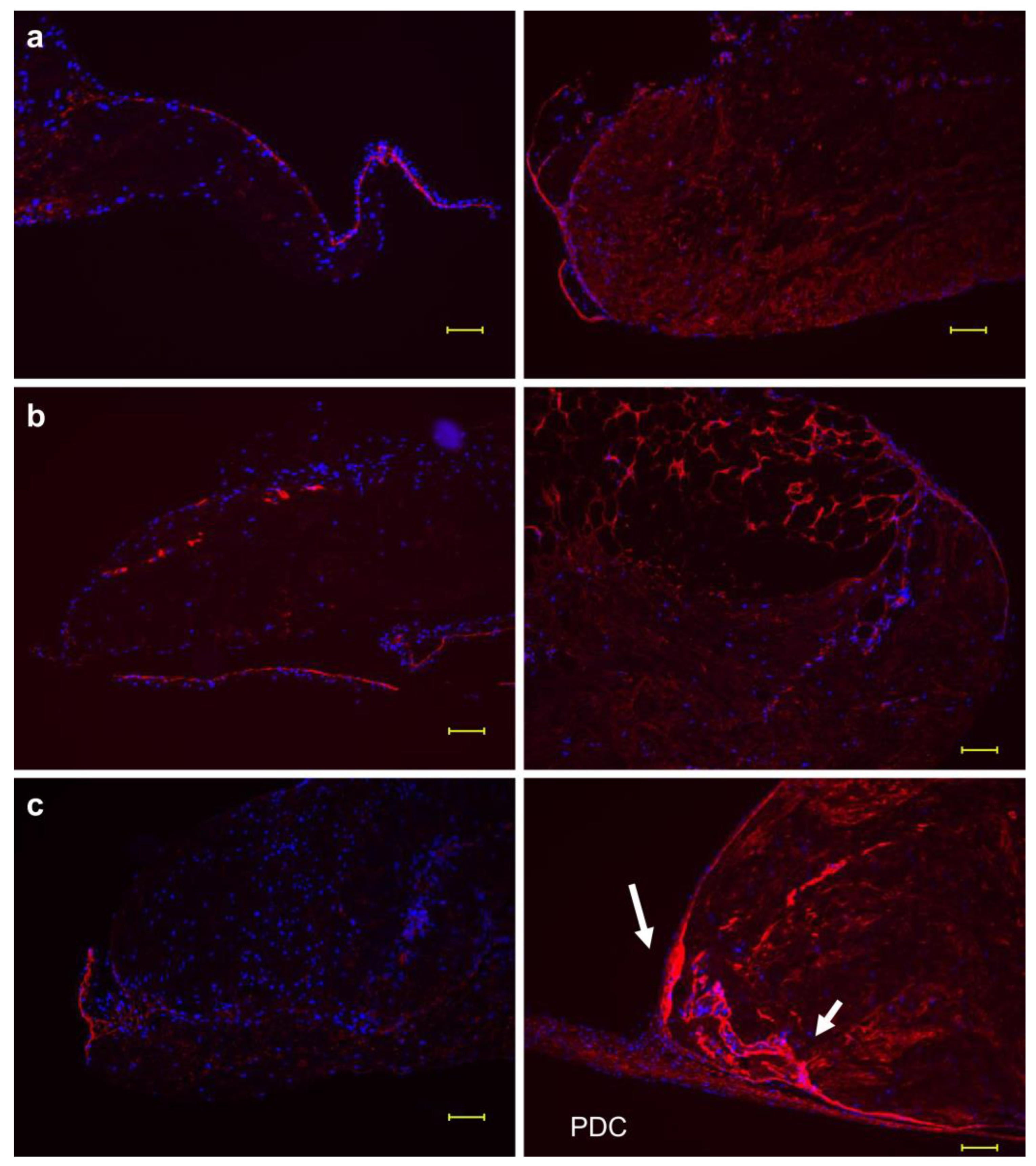
The developed multilayered sheet of PDCs outgrew from the original periosteum and periosteal cells during explant culture in vitro with only ascorbic acid. To seek necessary proteins for multilayered cell sheet formation and the subsequent bone regeneration, two conditions, that is, with and without ascorbic acid, were examined. Figure 2a–c shows the fluorescent immunostaining of FBXL14 in the periosteum following explant culture at weeks three, four and five under both culture medium conditions (with and without ascorbic acid). Without ascorbic acid, FBXL14 and PDCs formed a thin monolayer structure during weeks 3–5. However, with ascorbic acid, FBXL14 and PDCs formed thick multilayered cell sheets for up to five weeks. FBXL14 formed a strong membrane-like structure inside the periosteum (Figure 2, arrow), and periosteal cells inside moved and extended till outside the periosteum and PDCs were formed during week 5. Figure S1 shows the formation of a multilayer structure by FBXL14 and PDCs. Figure S1a–c are images of samples from different cows from Figure 2. PDCs became apparent mostly after three weeks (Figure S1a–c). Figure S1d is the negative control of Figure 2c (right). Elastica van Gieson staining (Figure S1e) revealed the formation of PDCs towards the outer side of the periosteum. Inside the periosteum, bundles of periosteal cells were supported by elastic fibers (arrow) from both sides, and PDCs were released outside. FBXL14 supported cells from inside to outside the periosteum (Figure 2 and Figure S1).
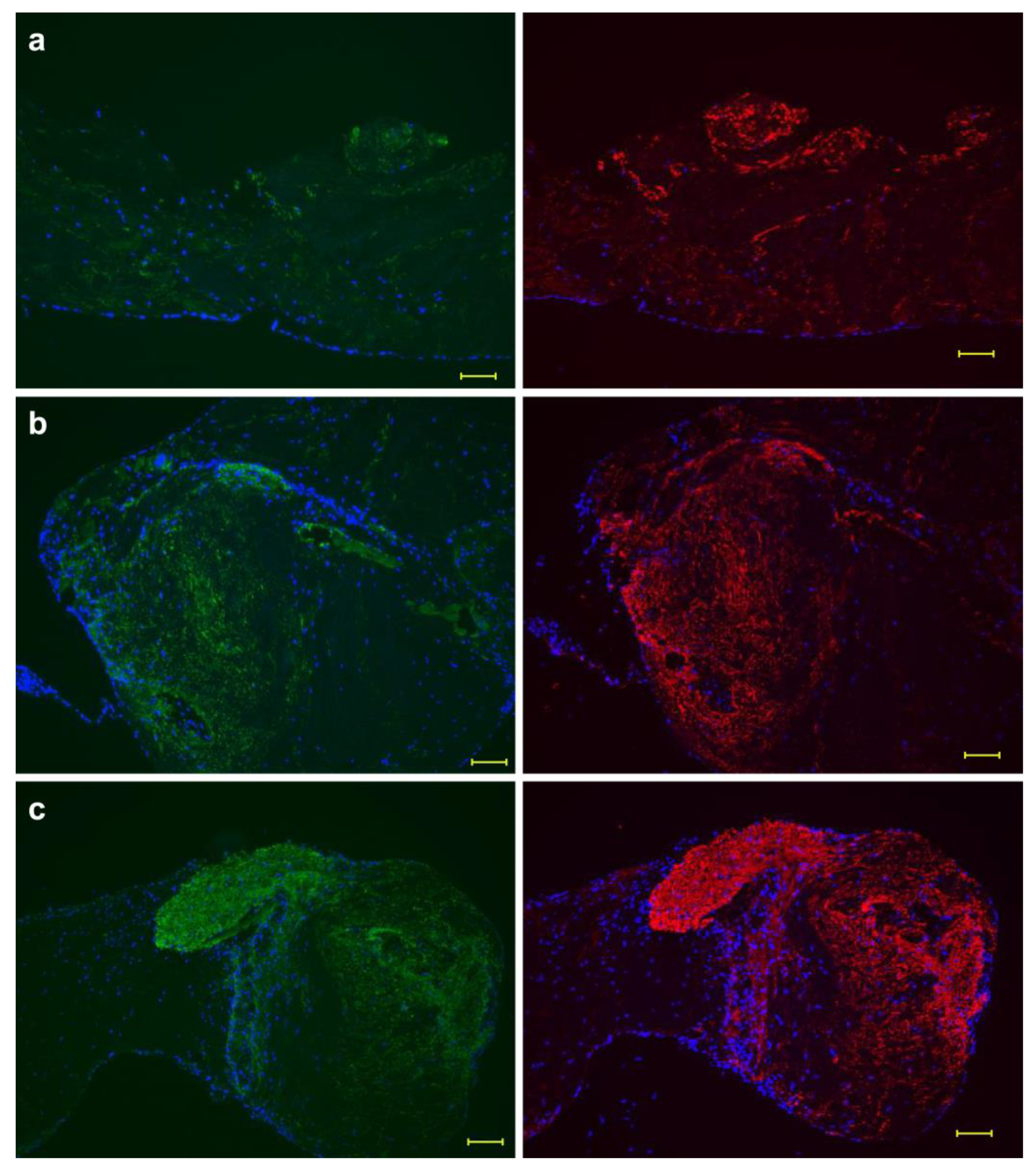
Figure 3a–c and Figure 4a–c show the periosteum in the absence of ascorbic acid. A previous study reported that osteocalcin and FBXW2 were expressed inside the periosteum in the presence of ascorbic acid [15]. To detect major proteins in bone regeneration, the condition without ascorbic acid was tested. Contrary to expectations, both osteocalcin and FBXW2 were expressed in the absence of ascorbic acid. Although osteocalcin expression was weaker than FBXW2 expression, their expression patterns were the same (Figure 3a–c). In the presence of ascorbic acid, not only osteocalcin and FBXW2 [15] but also FBXW2 and elastin were expressed in the same region [16] then osteocalcin and elastin expression was compared in the absence of ascorbic acid. However, the osteocalcin and elastin expression patterns were not the same, even though both proteins were localized in similar regions (Figure 4a–c). Elastin retained its fiber structure, whereas osteocalcin was separated from elastic fibers and showed a pattern spread over a wide area. Figure S2a–c shows the expression pattern of osteocalcin, FBXW2 and elastin at five weeks in the absence of ascorbic acid, using samples of different cows from Figure 3c and Figure 4c. Figure S2 reveals that the expression pattern of osteocalcin and FBXW2 was the same and the structure of fiber became obscure, while the insoluble elastin maintained the fiber structure as shown in Figure 3 and Figure 4.
4. Discussion
This study revealed that von Willebrand factor and FBXL14 are associated with blood vessels, but only FBXL14 is associated with bone lacuna (Figure 1a). Zhang et al. [7] reported that FBXL14 mRNA in bone marrow mesenchymal stem cells may play a key role in distraction osteogenesis. Cui et al. [5,6] evaluated the role of FBXL14 in ubiquitination. However, the relationship between the FBXL14 and bone remains unclear. In this study, FBXL14 formed a fine line around the periosteum during weeks 3–5 in the absence of ascorbic acid and a thick membrane inside the periosteum for up to 5 weeks in the presence of ascorbic acid (Figure 2a–c). For the clinical use of PDCs, many culture systems use spinner flasks [17], orbital shakers [18], passaged cells [19] or mechanical stretch [20]. Meanwhile, only primary PDCs were cultured in still dishes in this study. The process from periosteal cell to PDC formation with an extracellular matrix is of interest [21]. The results of this study suggest that FBXL14 may play a role in the scaffold preparation for forming the multilayered sheet of PDCs inside the periosteum.
I previously reported that osteocalcin localizes to the same region as FBXW2 in the presence of ascorbic acid [14]. Moreover, during explant culture, osteocalcin is deposited onto fibers from FBXW2 [15]. In this study, I compared the expression patterns of osteocalcin, FBXW2 and elastin in the absence of ascorbic acid. The results indicate that the expression patterns of osteocalcin and FBXW2 were similar, whereas those of osteocalcin and elastin were different (Figure 3 and Figure 4). In another study [16], I found that FBXW2 was associated with the elastic fibers. In the presence of ascorbic acid, FBXW2 and osteocalcin form fibrous structures [14,15]. It was inferred that FBXW2 and osteocalcin separated from the elastic fibers in the absence of ascorbic acid and could not retain their fiber shapes. A comparison of osteocalcin and FBXW2 and a comparison of osteocalcin and elastin were performed using single immunostaining; hence, no double-immunostaining was required. Two reasons for this are as follows: (1) preliminary experiments revealed that elastin was not clear with double-immunostaining and (2) in a previous study, regions of osteocalcin and FBXW2 spread and covered the fibers of elastin (Figure 4 and Figure S2).
Several studies have investigated elastin degradation and elastic fiber calcification [22,23,24,25,26,27,28,29,30]. Peranskunas et al. [31] reported the PDC secretome and found that, under osteogenic differentiation conditions, fibrillin-1 (not insoluble elastin), a major component of elastic fibers, was detected. The relationship between elastin degradation and osteocalcin remains unknown. However, the results of this study indicate that elastin is insoluble and shows low degradation, whereas FBXW2 is more flexible. As two conditions, that is, with and without ascorbic acid, modulated collagen deposition [12], osteocalcin was expressed in the absence of ascorbic acid (Figure 3 and Figure 4) and showed a pattern that spread over a wide area from elastic fibers. However, a mass of osteocalcin [15] was not observed, unlike that in the presence of ascorbic acid (Figure 3 and Figure 4). In the absence of ascorbic acid, osteocalcin was expressed. Osteocalcin expression does not always mean bone regeneration for the following reasons. First, before transplantation in vivo, the periosteum was cut off, and only PDCs were applied; calcification of the periosteum itself resulted in an insufficient supply of osteocalcin for osteogenic differentiation of PDCs. Second, the amount of FBXL14 is low. FBXL14 plays a role of bundling periosteal cells inside the periosteum. FBXL14 expression decreases in the absence of ascorbic acid as collagen synthesis decreases [12]; however, the relationship between FBXL14 and collagen is not clear. Two F-box proteins, FBXL14 and FBXW2, were contrastive. Here, FBXL14 levels increased around the periosteum during explant culture, while FBXW2 was originally present in the cambium layer of the periosteum.
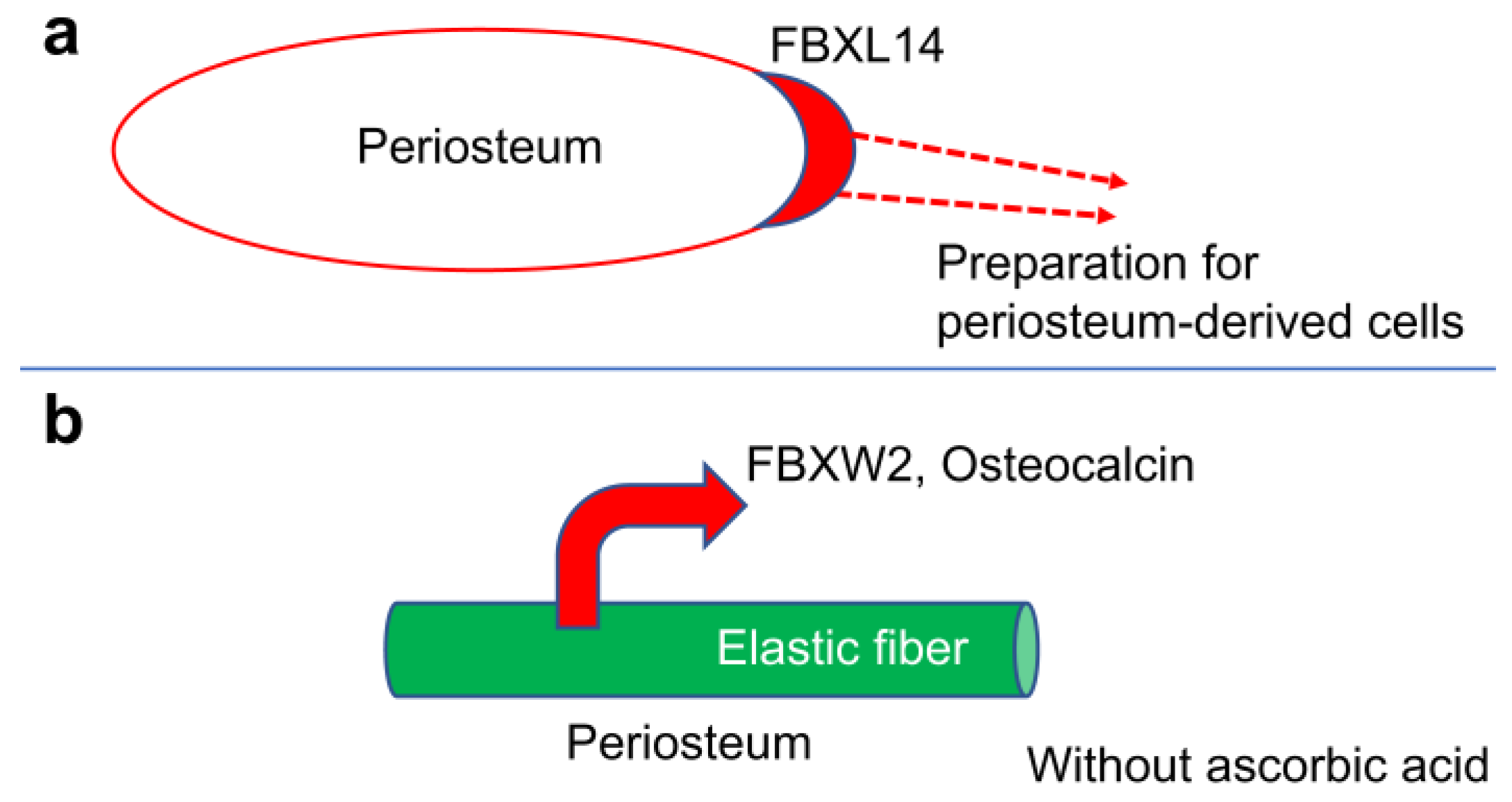
Gene knockdown using siRNA was not performed in this study. Only collagen synthesis was affected in the absence of ascorbic acid, and its effect on FBXL14 is unknown. Similar sections were used for immunostaining instead of double immunostaining. However, single immunostaining revealed differences between osteocalcin and elastin, and FBXW2 and elastin, whereas double immunostaining made elastin unclear. Here, FBXL14 appeared naturally; however, in future studies, a combination of FBXL14 and other biomaterials, such as collagen gel and decellular matrix, needs to be investigated as new scaffolds. A binding or departure system among FBXW2, elastin and osteocalcin is required for clarification. Osteocalcin deposition was not related to the formation of multilayered PDCs. FBXL14 in periosteum-derived cells appears to play a role in preparing a native scaffold for the multilayered cell sheet formation inside the periosteum (Figure 5a).
FBXW2 and osteocalcin spread from the elastin, and calcification occurred around elastin in the absence of ascorbic acid. These results indicate that the separation of FBXW2 from elastic fibers may result in bone regeneration failure instead of osteocalcin expression (Figure 5b). Clarifying the roles of FBXL14 and FBXW2 is important for the scaffold-free bone regeneration in clinical practice.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/osteology3010001/s1. Figure S1. Formation of a multilayer structure of periosteum-derived cells. Figure S2. Comparison of osteocalcin, FBXW2, and elastin at 5 weeks in the absence of ascorbic acid.
Funding
This research was funded by the Japan Society for the Promotion of Science (JSPS) KAKENHI, grant number [21K09947].
Institutional Review Board Statement
The animal study protocol was approved by the Institutional Ethics Committee of Osaka Dental University (protocol code: 22-01002, date of approval: 3 February 2022–31 March 2023).
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were generated or analyzed in this study. Data sharing is not applicable to this article.
Acknowledgments
The author thanks Kobe Chuo Chikusan for providing bovine legs and KAC Co., Ltd. for the unstained sections.
Conflicts of Interest
The author declares no conflict of interest. The funder had no role in the design of the study; in the collection, analyses or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Viñas-Castells, R.; Beltran, M.; Valls, G.; Gómez, I.; García, J.M.; Montserrat-Sentís, B.; Baulida, J.; Bonilla, F.; de Herreros, A.G.; Díaz, V.M. The hypoxia-controlled FBXL14 ubiquitin ligase targets SNAIL1 for proteasome degradation. J. Biol. Chem. 2010, 285, 3794–3805. [Google Scholar] [CrossRef] [Green Version]
- Zheng, H.; Du, Y.; Hua, Y.; Wu, Z.; Yan, Y.; Li, Y. Essential role of Fbxl14 ubiquitin ligase in regulation of vertebrate axis formation through modulating Mkp3 level. Cell Res. 2012, 22, 936–940. [Google Scholar] [CrossRef] [Green Version]
- Chen, F.; Zhang, C.; Wu, H.; Ma, Y.; Luo, X.; Gong, X.; Jiang, F.; Gui, Y.; Zhang, H.; Lu, F. The E3 ubiquitin ligase SCF(FBXL14) complex stimulates neuronal differentiation by targeting the Notch signaling factor HES1 for proteolysis. J. Biol. Chem. 2017, 292, 20100–20112. [Google Scholar] [CrossRef] [Green Version]
- Fang, X.; Zhou, W.; Wu, Q.; Huang, Z.; Shi, Y.; Yang, K.; Chen, C.; Xie, Q.; Mack, S.C.; Wang, X.; et al. Deubiquitinase USP13 maintains glioblastoma stem cells by antagonizing FBXL14-mediated Myc ubiquitination. J. Exp. Med. 2017, 214, 245–267. [Google Scholar] [CrossRef] [Green Version]
- Cui, Y.H.; Kang, J.H.; Suh, Y.; Zhao, Y.; Yi, J.M.; Bae, I.H.; Lee, H.J.; Park, D.W.; Kim, M.J.; Lee, S.J. Loss of FBXL14 promotes mesenchymal shift and radioresistance of non-small cell lung cancer by TWIST1 stabilization. Signal Transduct. Target Ther. 2021, 6, 272. [Google Scholar] [CrossRef]
- Cui, Y.H.; Kim, H.; Lee, M.; Yi, J.M.; Kim, R.K.; Uddin, N.; Yoo, K.C.; Kang, J.H.; Choi, M.Y.; Cha, H.J.; et al. FBXL14 abolishes breast cancer progression by targeting CDCP1 for proteasomal degradation. Oncogene 2018, 37, 5794–5809. [Google Scholar] [CrossRef]
- Zhang, T.; Jiang, W.; Liao, F.; Zhu, P.; Guo, L.; Zhao, Z.; Liu, Y.; Huang, X.; Zhou, N. Identification of the key exosomal lncRNAs/mRNAs in the serum during distraction osteogenesis. J. Orthop. Surg. Res. 2022, 17, 291. [Google Scholar] [CrossRef]
- Tekcham, D.S.; Chen, D.; Liu, Y.; Ling, T.; Zhang, Y.; Chen, H.; Wang, W.; Otkur, W.; Qi, H.; Xia, T.; et al. F-box proteins and cancer: An update from functional and regulatory mechanism to therapeutic clinical prospects. Theranostics 2020, 10, 4150–4167. [Google Scholar] [CrossRef]
- Mason, B.; Laman, H. The FBXL family of F-box proteins: Variations on a theme. Open Biol. 2020, 10, 200319. [Google Scholar] [CrossRef]
- Akiyama, M.; Nonomura, H.; Kamil, S.H.; Ignotz, R.A. Periosteal cell pellet culture system: A new technique for bone engineering. Cell Transplant. 2006, 15, 521–532. [Google Scholar] [CrossRef]
- Akiyama, M. Characterization of the F-box proteins FBXW2 and FBXL14 in the initiation of bone regeneration in transplants given to nude mice. Open Biomed. Eng. J. 2018, 12, 75–89. [Google Scholar] [CrossRef] [PubMed]
- Akiyama, M.; Nakamura, M. Bone regeneration and neovascularization processes in a pellet culture system for periosteal cells. Cell Transplant. 2009, 18, 443–452. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.Q.; Al-Tamimi, M.; Uddin, S.; Steinhoff, M. F-box proteins in cancer stemness: An emerging prognostic and therapeutic target. Drug Discov. Today 2021, 26, 2905–2914. [Google Scholar] [CrossRef] [PubMed]
- Akiyama, M. FBXW2 localizes with osteocalcin in bovine periosteum on culture dishes as visualized by double immunostaining. Heliyon 2018, 4, e00782. [Google Scholar] [CrossRef]
- Akiyama, M. Role of FBXW2 in explant cultures of bovine periosteum-derived cells. BMC Res. Notes 2021, 14, 410. [Google Scholar] [CrossRef]
- Akiyama, M. Elastic fibers and F-Box and WD-40 domain-containing protein 2 in bovine periosteum and blood vessels. Biomimetics 2023, 8, 7. [Google Scholar] [CrossRef]
- Van Beylen, K.; Papantoniou, I.; Aerts, J.M. Microcarrier screening and evaluation for dynamic expansion of human periosteum-derived progenitor cells in a xenogeneic free medium. Front. Bioeng. Biotechnol. 2021, 9, 624890. [Google Scholar] [CrossRef]
- Cha, H.M.; Kim, S.M.; Choi, Y.S.; Kim, D.I. Scaffold-free three-dimensional culture systems for mass production of periosteum-derived progenitor cells. J. Biosci. Bioeng. 2015, 120, 218–222. [Google Scholar] [CrossRef]
- Choi, Y.S.; Noh, S.E.; Lim, S.M.; Lee, C.W.; Kim, C.S.; Im, M.W.; Lee, M.H.; Kim, D.I. Multipotency and growth characteristic of periosteum-derived progenitor cells for chondrogenic, osteogenic, and adipogenic differentiation. Biotechnol. Lett. 2008, 30, 593–601. [Google Scholar] [CrossRef]
- Ito, R.; Matsumiya, T.; Kon, T.; Narita, N.; Kubota, K.; Sakaki, H.; Ozaki, T.; Imaizumi, T.; Kobayashi, W.; Kimura, H. Periosteum-derived cells respond to mechanical stretch and activate Wnt and BMP signaling pathways. Biomed. Res. 2014, 35, 69–79. [Google Scholar] [CrossRef]
- Zhang, W.; Wang, N.; Yang, M.; Sun, T.; Zhang, J.; Zhao, Y.; Huo, N.; Li, Z. Periosteum and development of the tissue-engineered periosteum for guided bone regeneration. J. Orthop. Transl. 2022, 33, 41–54. [Google Scholar] [CrossRef] [PubMed]
- Basalyga, D.M.; Simionescu, D.T.; Xiong, W.; Baxter, B.T.; Starcher, B.C.; Vyavahare, N.R. Elastin degradation and calcification in an abdominal aorta injury model: Role of matrix metalloproteinases. Circulation 2004, 110, 3480–3487. [Google Scholar] [CrossRef] [PubMed]
- Simionescu, A.; Simionescu, D.T.; Vyavahare, N.R. Osteogenic responses in fibroblasts activated by elastin degradation products and transforming growth factor-beta1: Role of myofibroblasts in vascular calcification. Am. J. Pathol. 2007, 171, 116–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hosaka, N.; Mizobuchi, M.; Ogata, H.; Kumata, C.; Kondo, F.; Koiwa, F.; Kinugasa, E.; Akizawa, T. Elastin degradation accelerates phosphate-induced mineralization of vascular smooth muscle cells. Calcif. Tissue Int. 2009, 85, 523–529. [Google Scholar] [CrossRef] [PubMed]
- Pai, A.; Leaf, E.M.; El-Abbadi, M.; Giachelli, C.M. Elastin degradation and vascular smooth muscle cell phenotype change precede cell loss and arterial medial calcification in a uremic mouse model of chronic kidney disease. Am. J. Pathol. 2011, 178, 764–773. [Google Scholar] [CrossRef]
- Sinha, A.; Vyavahare, N.R. High-glucose levels and elastin degradation products accelerate osteogenesis in vascular smooth muscle cells. Diab. Vasc. Dis. Res. 2013, 10, 410–419. [Google Scholar] [CrossRef] [Green Version]
- Janssen, R. Magnesium to counteract elastin degradation and vascular calcification in chronic obstructive pulmonary disease. Med. Hypotheses 2017, 107, 74–77. [Google Scholar] [CrossRef]
- Bartstra, J.W.; Spiering, W.; van den Ouweland, J.M.W.; Mali, W.; Janssen, R.; de Jong, P.A. Increased elastin degradation in pseudoxanthoma elasticum is associated with peripheral arterial disease independent of calcification. J. Clin. Med. 2020, 9, 2771. [Google Scholar] [CrossRef]
- Boraldi, F.; Moscarelli, P.; Lofaro, F.D.; Sabia, C.; Quaglino, D. The mineralization process of insoluble elastin fibrillar structures: Ionic environment vs. degradation. Int. J. Biol. Macromol. 2020, 149, 693–706. [Google Scholar] [CrossRef]
- Zhao, S.; Cao, J.; Li, J.; Yang, X.; Cao, P.; Lan, J.; Lu, G. Association between serum elastin-derived peptides and abdominal aortic calcification in peritoneal dialysis patients: A cross-sectional study. Ren. Fail. 2021, 43, 860–868. [Google Scholar] [CrossRef]
- Pranskunas, M.; Simoliunas, E.; Alksne, M.; Kaupinis, A.; Juodzbalys, G. Periosteum-derived mesenchymal stem cells secretome—Cell-free strategy for endogenous bone regeneration: Proteomic analysis in vitro. J. Oral Maxillofac. Res. 2021, 12, e2. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Expression of FBXL14 in blood vessels and bone lacunae. Staining of FBXL14 and von Willebrand factor in the bone with periosteum. FBXL14 was expressed in the periosteum and bone lacuna blood vessels. (a) FBXL14: red; 4′,6-diamidino-2-phenylindole (DAPI): blue; arrow: bone lacuna. (b) Von Willebrand factor: red; DAPI: blue; B: bone. Scale bar: 100 μm.
Figure 1.
Expression of FBXL14 in blood vessels and bone lacunae. Staining of FBXL14 and von Willebrand factor in the bone with periosteum. FBXL14 was expressed in the periosteum and bone lacuna blood vessels. (a) FBXL14: red; 4′,6-diamidino-2-phenylindole (DAPI): blue; arrow: bone lacuna. (b) Von Willebrand factor: red; DAPI: blue; B: bone. Scale bar: 100 μm.

Figure 2.
Role of FBXL14 in the periosteum. Fluorescent immunostaining of FBXL14. (left) Without ascorbic acid. (right) With ascorbic acid. FBXL14: red; 4′,6-diamidino-2-phenylindole (DAPI): blue. (a): At week 3; (b): at week 4; (c): at week 5. PDCs: periosteum-derived cells; arrow: a strong membrane-like structure. Scale bar: 100 μm.
Figure 2.
Role of FBXL14 in the periosteum. Fluorescent immunostaining of FBXL14. (left) Without ascorbic acid. (right) With ascorbic acid. FBXL14: red; 4′,6-diamidino-2-phenylindole (DAPI): blue. (a): At week 3; (b): at week 4; (c): at week 5. PDCs: periosteum-derived cells; arrow: a strong membrane-like structure. Scale bar: 100 μm.

Figure 3.
Expression patterns of osteocalcin and FBXW2. A comparison of osteocalcin (left) and FBXW2 (right) in the absence of ascorbic acid. Expression patterns of osteocalcin and FBXW2 were similar. Osteocalcin: green; FBXW2: red; 4′,6-diamidino-2-phenylindole (DAPI): blue. (a) At week 3; (b) at week 4; (c) at week 5. Scale bar: 100 μm.
Figure 3.
Expression patterns of osteocalcin and FBXW2. A comparison of osteocalcin (left) and FBXW2 (right) in the absence of ascorbic acid. Expression patterns of osteocalcin and FBXW2 were similar. Osteocalcin: green; FBXW2: red; 4′,6-diamidino-2-phenylindole (DAPI): blue. (a) At week 3; (b) at week 4; (c) at week 5. Scale bar: 100 μm.

Figure 4.
Expression patterns of osteocalcin and elastin. A comparison of osteocalcin (left) and elastin (right) in the absence of ascorbic acid. Their expression patterns are different. Osteocalcin: green (left); elastin: green (right); 4′,6-diamidino-2-phenylindole (DAPI): blue. (a) At week 3; (b) at week 4; (c) at week 5. Scale bar: 100 μm.
Figure 4.
Expression patterns of osteocalcin and elastin. A comparison of osteocalcin (left) and elastin (right) in the absence of ascorbic acid. Their expression patterns are different. Osteocalcin: green (left); elastin: green (right); 4′,6-diamidino-2-phenylindole (DAPI): blue. (a) At week 3; (b) at week 4; (c) at week 5. Scale bar: 100 μm.

Figure 5.
Summary of this study. (a) Role of FBXL14 for PDCs: inside the periosteum, FBXL14 prepares a native scaffold for the multilayered cell sheet formation. (b) Role of FBXW2 in the calcification process: FBXW2 and osteocalcin separate from the elastic fiber in the absence of ascorbic acid.
Figure 5.
Summary of this study. (a) Role of FBXL14 for PDCs: inside the periosteum, FBXL14 prepares a native scaffold for the multilayered cell sheet formation. (b) Role of FBXW2 in the calcification process: FBXW2 and osteocalcin separate from the elastic fiber in the absence of ascorbic acid.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Conditions for antigen retrieval, primary and secondary antibodies, and culture medium for each target protein.
Table 1.
Conditions for antigen retrieval, primary and secondary antibodies, and culture medium for each target protein.
| Protein | Antigen Retrieval | Primary Antibody | Secondary Antibody |
|---|---|---|---|
| FBXL14 (with and without ascorbic acid) | Proteinase K (Room Temperature, 10 min) | #SAB2103691 1:500 4 h (Sigma-Aldrich, Saint Louis, MO, USA) | Alexa FluorTM 594 goat anti-rabbit IgG (H+L) #A11037 1:200 1 h (Invitrogen, Eugene, OR, USA) |
| Von Willebrand factor | Proteinase K (Room Temperature, 10 min) | #ab6994 1:300 4 h (Abcam, Cambridge, UK) | |
| Elastin (without ascorbic acid) | Proteinase K (Room Temperature, 10 min) | Elastin (BA-4) #sc-58756 1:200 4 h (Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA) | Alexa FluorTM 488 goat anti-mouse IgG (H+L) #A32731 1:200 1 h (Invitrogen) |
| Osteocalcin (without ascorbic acid) | Proteinase K (Room Temperature, 10 min) | Monoclonal antibody to osteocalcin #M042 1:500 4 h (Takara Bio Inc., Shiga, Japan) | |
| FBXW2 (without ascorbic acid) | Proteinase K (Room Temperature, 10 min) | #PA5-18189 1:200 4 h (Invitrogen) | Mouse anti-goat IgG-CFL 594 #sc516243 1:200 1 h (Santa Cruz Biotechnology, Inc.) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Akiyama, M. Roles of Two F-Box Proteins: FBXL14 in the Periosteum and FBXW2 at Elastic Fibers. Osteology 2023, 3, 1-10. https://doi.org/10.3390/osteology3010001
AMA Style
Akiyama M. Roles of Two F-Box Proteins: FBXL14 in the Periosteum and FBXW2 at Elastic Fibers. Osteology. 2023; 3(1):1-10. https://doi.org/10.3390/osteology3010001
Chicago/Turabian StyleAkiyama, Mari. 2023. "Roles of Two F-Box Proteins: FBXL14 in the Periosteum and FBXW2 at Elastic Fibers" Osteology 3, no. 1: 1-10. https://doi.org/10.3390/osteology3010001